Vol.44, No.1 JOURNALOFVIROLOGY, Oct.1982,p.67-77
0022-538X/82/100067-11$02.00/0
Copyright©1982, American Society for Microbiology
Structural
and
Biological Analysis of Integrated Polyoma
Virus DNA
and
Its
Adjacent
Host
Sequences Cloned
from
Transformed
Rat
Cells
ADRIANHAYDAY,tH. EARL RULEY,AND MIKE FRIED*
Departmentof TumourVirusGenetics,Imperial Cancer Research Fund, Lincoln'sInnFields, London WC2A
3PX,England
Received 5 February1982/Accepted 22 June 1982
EcoRIfragments containing integrated viral and adjacent hostsequences were cloned from two polyoma virus-transformed cell lines (7axT and 7axB) which eachcontainasingle insert ofpolyoma virus DNA. ClonedDNAfragments which containedacompletecodingcapacity for the polyoma virus middle and small T-antigenswerecapable oftransforming ratcells invitro. Analysis oftheflanking sequences indicated thatratDNAhad been reorganized ordeleted atthe sitesof polyoma virus integration, but none of the hallmarks of retroviral integration,
such as the duplication of host DNA, were apparent. There was no obvious similarity of DNA sequencesin the four virus-hostjoins. In one case the virus-hostjunctionsequencepredicted the virus-host fusion protein whichwasdetected in thetransformed cell line. DNAhomologoustotheflanking sequencesof three outof four of thejoins waspresent in single copy in untransformed cells. One copy of the flanking host sequences existed in an unaltered form in the two transformed cell lines, indicating that a haploid copy of the viral transforming
sequences is sufficient to maintain transformation. The flanking sequencesfrom onecelllinewerefurther usedas aprobetoisolateatargetsite(unoccupied site) forpolyoma virus integrationfrom uninfected cellularDNA.Therestriction map of this DNA was in agreement with that of the flanking sequences, but the sequenceof theunoccupied siteindicated that viralintegrationdidnotinvolvea
simple recombination event between viral and cellular sequences. Instead,
sequencerearrangements oralterations occurredimmediatelyadjacenttotheviral insert, possiblyas aconsequenceof theintegration of viral DNA.
Polyoma virus (Py)cantransformavarietyof cell types to a neoplastic state. In virus-trans-formed cells, Py sequences are retained and expressed as part of the host cell genome
(re-viewedinreference34).Thestudyofintegrated
viral sequences has been complementaryto oth-er approaches in understanding the interaction
of the virus with the host cell. These studies, whichwere largely dependenton Southern blot analysis, indicated that the retention of the cod-ing capacities for small and middle T-antigens (T-Ags) is sufficienttomaintain cell transforma-tion (17,19,21,22, 27). Retention ofthelargeT coding capacity is not necessary to maintain transformation (20), although the large T does
playaroleininitiatingtransformation after viral
infection (10-12). In transformed cells Py se-quenceshavebeen observedtobecome
integrat-ed at different chromosomal locations (19, 22,
t Present address: Center for CancerResearch, Massachu-setts Institute of Technology, 77 Massachusetts Avenue, Cambridge,MA02139.
23).Thequestions which defiedprevious analy-seswere whether the chromosomal locations of
integrated viral sequences fell into a subset of
the hostgenome (e.g., highly reiterated DNA); whether the adjacent cellular DNA modified the biological activity of the viral DNA; whether there was a sequence specificity to integration manifested by similar sequences at the
virus-hostjoins;whetherhomology between viral and
host DNA mediated integration; and whether
theintegrationof viralsequencesperturbed
(du-plicated/deleted/inverted) the host cell
se-quences at the site ofintegration. These ques-tions requirecloning of the integrated viralDNA from transformed cells andofthecellular DNA at the site of viral insertion from uninfected cells. This approach has been followed here.
MATERIALS AND METHODS
Cell culture and biological assays. The isolation of cell lines 7axT and 7axB has previously been
de-scribed (22, 23). Cellsweregrown inDulbecco modi-67
on November 10, 2019 by guest
http://jvi.asm.org/
68 HAYDAY, RULEY, AND FRIED fied Eagle medium containing 5% fetal calf serum. Whole mouse embryo cells were used for plaque assays aspreviously described (20). For transforma-tion assays, 0.5 ,ug of Py-pAT153 recombinant plasmid DNAs in the presence of 20 p.gof rat carrier DNA were applied to semiconfluent Rat-1 cells (about 106
perdish) in calcium phosphate precipitates as previ-ouslydescribed (13, 39). Cell sheets were stained for transformed foci 2 weeks later with Leishman stain.
Preparation, restriction, and fractionation of
eucary-oticcellDNA.Total cell DNAwasprepared essentially aspreviously described (20, 21, 23).ForSouthern (31) analysis of cell DNA, endonuclease digestions were performed on about 10 ,ug of cell DNA for 3 to6h, depending on the enzyme used. The enzymes were
purchased from New England Biolabs, and the pre-scribedbuffers were used.Preparative fractionation of endonuclease-cleaved cell DNA was on 10 to 40%
sucrosevelocity gradients,asdescribed by Maniatiset
al.(25).Fractions were assayed for the presence of Py sequencesbySouthernblotanalysis.
Gelelectrophoresis, transfer,andhybridization
pro-cedures. DNA waselectrophoresed in agarosegels of 0.6 to2%with E buffer(18). The DNA was denatured insitu, transferred to nitrocellulose filters(Schleicher andSchuell, BA85), andhybridized tovirion-purified
[32P]DNAnick-translatedto aspecificactivity of 1 x
108 to 2 x 108 cpm/,ug (20, 21, 23). Blots were autoradiographed on presensitized Fuji Rx film at
-70°Cwithintensityscreens(Dupont).
Cloning eucaryotic DNA into procaryotic vectors.
Fractions ofEcoRI-cleaved7axBand 7axT cell DNA which contained Py sequences were prepared as de-scribed above and ligated to the purified arms of Agt.WES.Ab. In the standard reaction, between 0.15 and 0.25 ,ug of cell DNA was ligated to 1.5 ,ug of X
arms. T4 DNA ligasewas supplied byNew England
Biolabs. Packaging of A DNA in vitrowasperformed
aspreviouslydescribed(15);between2 x105 and1 x 106plaques perp.gofX arms wereobtained. Thephage
[image:2.496.58.251.493.645.2]were plated on Escherichia coli LE392 (33) and screened by the method of Benton and Davis (3).
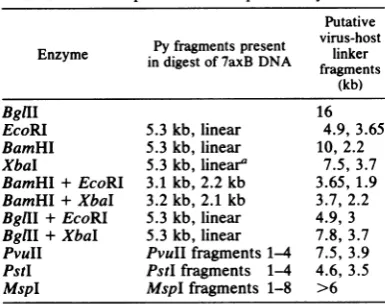
TABLE 1. Summary ofSouthernblot data derived fromanalysisof 7axB cell DNA with the enzymes
indicated andaprobe ofvirion-purifiedPy DNA
Putative Pyfrgmensprsent virus-host
Enzyme indigestof 7axB DNA linker
(kb)
BgllI 16
EcoRI 5.3kb, linear 4.9, 3.65
BamHI 5.3kb,linear 10,2.2
XbaI 5.3kb,linear' 7.5,3.7
BamHI+ EcoRI 3.1 kb, 2.2kb 3.65,1.9
BamHI+ XbaI 3.2kb, 2.1 kb 3.7,2.2
BglII+ EcoRI 5.3kb, linear 4.9,3
BgIII+ XbaI 5.3kb, linear 7.8,3.7
PvuII PvuIIfragments1-4 7.5,3.9 PstI PstIfragments 1-4 4.6,3.5
MspI MspI fragments 1-8 >6
aXbal cuts Py DNA twice, but since one of the fragments is 45 bp long,XbaIisregardedas a
single-cutenzyme whenusedtoanalyze genomicblots.
Positive plaques were picked andpurified, and their
DNAwas isolated by the method of Maniatis et al. (25). Cloned Py DNA fragments were ligated into EcoRI-cleaved pAT153 (36) DNA and introduced into
E.coli HB101.
Preparationof plasmid DNA. The method of
Birn-boim and Doly (4) was used to obtain yields of recombinant plasmid DNAof >1 mg/liter of culture. After CsCl densitycentrifugation, plasmid DNAs were furtherpurified on sucrose gradients (5 to 20%) con-tainingethidium bromide.
DNA sequence analysis. Restriction endonuclease cleavage siteswere5'-end labeledaftertreatmentwith calf intestine alkaline phosphatase and T4 polynucleo-tide kinase (38). After secondary enzyme cleavage, labeled fragments were fractionated on polyacryl-amidegels andeluted (26). The chemical degradation method of Maxam and Gilbert was employed as de-scribed(26), except the G + Areaction was stopped with 200 ,ul of hydrazine stop buffer and ethanol precipitated. The degradation products were resolved
onthin urea-acrylamide gels as described by Sanger and Coulson (28).DNAsequences wereanalyzed with theSEQ computer program (6).
RESULTS
State of the viral DNA in cell lines 7axB and 7axT. Cell lines 7axB and 7axT were isolated from independent monoclonaltumorsinduced in rats by the injection ofPy-transformed cells of theline7ax(23). Although7axcontained multi-ple copies of both free andintegratedPy DNA, both 7axB and 7axT contained only a single
insert ofPy DNA and lacked free viral forms (23). The structure of the integrated viral se-quences in the 7axT cells as determined by Southern blot analysis has previously been
re-portedtocompriseasingle insert ofcontinuous
viralsequences (0.8 to 0.85genomes in length) extending from 26 to 32 map units(MU) atone end tobetween 12 and14 MUatthe other end (22). A preliminary description of the 7axB insertindicated thepresenceof head-to-tail tan-demly duplicated Py sequences (23). By an analysis ofDNAfragments from acombination ofcleavages with enzymes which cut Py DNA once orseveral times andenzymeswhich donot cutPy DNAwhich used the strategyemployed by Laniaetal.(21) formappingtandem head-to-tail integrated viral sequences, the structure of the 7axB insert was derived. Briefly, enzymes which cleave the Py genome once (EcoRI,
BamHI)generateda 5.3-kilobase (kb)linear Py
DNA fragment. A 5.3-kb fragment was also generated by the cleavage of 7axB DNA with XbaI. Cleavage of7axB DNA with enzymes that
cut Py DNA at multiple sites (PstI, PvuII,
HpaII) generated all the fragments expected
from the cleavage ofthe circular Py genome. From this data the 7axB insert wasjudged to contain a head-to-tail tandem duplication
ex-tendingfrom 59to70MUatthe left endto79to
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
INTEGRATED POLYOMA VIRUS DNA 69
90 MU at theright end. The resultsofsomeof these digestions have beenpreviously presented (23) andarelistedalongwith additional cleavage datain Table1.Theextentofviral sequences at each endwasjudgedasfollows.At the left-hand end, the EcoRI virus-host linker fragment (3.65 kb) hybridizedto apurifiedprobe of PyHpaII-3 (54to70 MU) but didnotcontain the PyBamHI site (59 MU). At the other end, the EcoRI virus-host linker fragment hybridized to a purified probe of Py HpaII-4 (78to 91 MU) but did not contain the Py AvaI siteat 90MU. The resulting maps of Py DNA in 7axT and 7axB cells are shown (Fig. 1). 7axT contains anintact coding region for small and middle T-Ags, whereas 7axB contains two complete coding regions for large, middle, and small T-Ags. The sequence arrangement of the two inserts was studied in more detail after molecular cloning had been
performed.
Isolationandcharacterization of the 7axB and
7axT viral inserts and adjacent host sequences. Theintegrated viralsequencesof7axTand7axB were contained in two and three EcoRI frag-ments, respectively (Fig. 1). Size-fractionated EcoRI-cleaved DNA from both cell lines was
clonedinto
Xgt.
WES.Xbas described above. TheEcoRI fragments werethen subcloned into the
plasmidpAT153 and analyzed. The cloned3.6-,
5.3-, and 4.9-kb EcoRIfragments from 7axBare referred to as 7B-L (left), 7B-M (middle), and 7B-R(right), respectively (Fig. 1). The 4.7- and 5.1-kb cloned EcoRI fragments of 7axT are referred to as 7T-L(left) and7T-R (right) (Fig. 1). These cloned fragments were indistinguish-ableinelectrophoretic mobilityfrom theEcoRI
fragmentsgenerated from total cellular DNA. In
addition, analysisof the clonedfragmentswitha
number ofrestriction enzymes both confirmed their derivation from the integrated viral DNA structures mapped in Fig. 1 and allowed better resolution of the virus-hostjoin regions. In sev-eral cases putative virus-host linker fragments detected by Southern analysis of cellular DNA were verifiedas such byrestriction cleavage of cloned DNA. For 7T-L, the extent of viral sequences attheleft-hand end of the viral insert was 100base pairs (bp)greaterthan had
previ-ouslybeen determinedbySouthern blotanalysis
of cell DNA (22). Thisdiscrepancy resulted from the inability tocleave 7axT DNA at eitherthe HhaIsiteatnucleotide(NT) 2,962orthe HpaII site atNT 2,991. Since MspI (37) cleavesboth
7T-Land 7axTDNA at NT2,991, theinabilityof
HpaIItocleave7axT DNA waspresumablydue
tomethylationatthat site.
Nature of the adjacent host sequences and cloning of an unoccupied site DNA from Rat-1 cellularDNA.The clones7B-R,7B-L, 7T-R, and 7T-L were used to probe untransformed Rat-1
cellular DNAtoestimatethecopynumberof the adjacent host sequences in the cellulargenome (Fig. 2). The host sequences in three of the clones were foundto berepresented only once in EcoRI-cleaved Rat-1 DNA (7B-L in a frag-mentof6.2kb; 7B-R, 2kb; 7T-L,0.75kb). The host sequences in 7T-R were found in at least twoEcoRIfragments ofRat-1 DNAof5and 1.3 kb, respectively. The clonescontaining flanking
cellular DNA were also used to probe EcoRI-and XbaI-cleaved DNA from 7axB and 7axT cells. In each case the probes detected two classes of fragments: (i) the fragment(s) due to viralsequences incommon (whichrepresentall or part of the proviral inserts), and (ii) the fragment(s) detected by that probe in
untrans-formed cell DNA. The presence of class ii fragments suggested that theintegration of viral DNA is haploid in the respective transformed cell lines. The observation that probes from 7axB cellshybridizedtodifferentfragmentsthan did probes from 7axT cells confirms that the Py
DNA in these cell lines isintegrated into differ-entsites in the hostgenome.That theright-hand
flanking sequences ofeither 7axT or 7axB
hy-bridized to fragments in Rat-1 DNA distinct from those detected by the left-hand flanking
sequences shows that the viral DNA has not
merely been inserted into the host DNA, but that cellular sequences have been rearranged
(e.g.,deleted)atthe site ofintegration.
The EcoRIfragment homologousto7B-Lwas
clonedfrom untransformedRat-1 cells intoXgt. WES.Xb and subcloned into pAT153 (pAU-L). Arestrictionmapfor theunoccupiedsite, pAU-L, was determined by complete and partial
digestion with more than a dozen restriction enzymes(Fig. 3). For about 1.3 kbfrom the left-hand EcoRI site, pAU-L had arestriction map identicalto7B-L.Atabout thatpoint,the
flank-ing sequences in 7B-L becamejoined to viral DNA.
DNAsequences of the virus-hostjoins and the unoccupied-site DNA. The DNA sequences around thevirus-hostjoins of 7B-L, 7B-R, 7T-L, and7T-R areshown inFig. 4. Continuous viral sequences were detected in 7B-L up to NT 4,682, in 7B-R up to NT916, in 7T-L up to NT 2,838, and in 7T-R up toNT 2,252. At each of thesepointsthere was anabruptdiscontinuityin theviralsequences.Thisisafeature ofotherPy virus-host joins that have been studied (27a; H. E. Ruley,unpublished data).Acloser analy-sis of the sequence of 7T-L revealed that the discontinuity in viralsequencescouldhave been due to avirus-virus rather thanavirus-hostjoin. Theeleven nucleotideswhichabut thejoin could be derivedfromPyDNA(NTs 2,943 to 2,932). This arrangement of the viral sequence could have resultedfrom deletion of viralDNAfroma VOL.44, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
70 HAYDAY, RULEY, AND FRIED
7
m_
co
i
I_
2
It.. m rv
co
I
0
U.
_
8
Lu-~ m
10
04
N
-aF-Iq
W)
Cl)
10
C14
N
Iq
tn Cl)
i
I
lii
0
-U,)
i.
I
r.l Iq
-i
r-AI
I---(U
I
(N4
N
'IV
C..)
.1
4
C) O;; ,
O0°,5-X
.0 0
C13
Cd) CZ
z0j
-0
Dn r t
cCZ ru
t
o°
-4)4
E-5 0.0 c- 4)24)
x c13
0 o
.0
m
<=
b C) 4)r
U,
-4)-'
cr2
Zo.0o
c^s 04
0
CI C: 4
J.VIROL.
J
8D-x
on November 10, 2019 by guest
http://jvi.asm.org/
INTEGRATED POLYOMA VIRUS DNA 71
7B-L 7BM
U7axB1xTAd2I 7axBlxTP
..4 -b
.
*11 ar- 5-3kb
a
2kb-7B-R 71R 7T-L
U XaxB 7axT 7RpAT1 U17axT 7axT U
to
5-1kbo- m5L 5kb53kb.-..kbf
451kb
37kb.-.
41
3b755bpM
FIG. 2. Detection of flankinghost DNAsequencesinnormal andratcellDNA, indicatingthehaploidnature of theinsertions ofPy sequencesand rearrangements atthe site ofintegrationin 7axT and 7axB cells. Total cellular DNA from7axT, 7axB,anduntransformed cells(U)wascleaved withEcoRI, separatedona1%agarose gel, transferredtonitrocellulose paper,andprobed with thepAT153 recombinant DNAs. The captionabove
eachpanelindicates theprobe used,and the notation above each lane indicates theEcoRI-cleavedDNAprobed (pAT, pAT153DNA; Py, Py DNA). The bands detected with the7B-M probe(contains only Py sequences)
identifiedfragments of 7axT (4.7and 5.1 kb) and 7axB (3.6, 5.3, and 4.9kb) cellular DNAs containingviral
sequences (see Fig. 1).Bands in addition tothese fragmentsdetected inuntransformed, 7axB, and 7axTcell DNAidentified host DNAsequenceshomologousto ratcellularsequencesinpAU-Lortheflankingsequences of7B-L, 7B-R, 7T-L,or7T-R. The 6.15-kb rat cell DNAfragment (sequencescontained in theunoccupiedsite ratcellularDNAclone pAU-L)wasdetected inuntransformed,7axB,and 7axTcell DNAs wheneitherpAU-L
or7B-Lwasusedasprobe. In 7axB DNA this host 6.15-kbfragmentwasdetectedinadditiontothe 3.6-kb 7B-L
fragment with the pAU-L and 7B-L probes. When 7B-R wasused as aprobe, the 2-kb host DNAfound in untransformed cell DNAwasdetected in 7axB DNA in addition tothe4.9-kb 7B-Rfragment.When 7T-Rwas
usedas aprobe, the 5- and 1.3-kbratDNAfragmentsfound in untransformed cell DNAweredetected in7axT DNA in addition to the 5.1-kb 7T-R fragment. When 7T-L was used as a probe, the 750 bp found in untransformed cell DNAwasdetected in 7axT DNA inaddition to the 4.7-kb 7T-Lfragment. Ad2, HindIll-cleaved adenovirus 2 DNA usedas amarker.
tandem duplication of Py sequences. Because
the probability that 11 out of 13 nucleotides
around the virus-host join would occur by
chanceonce oneitherstrand of thePygenomeis
less than 1 in300(6),wefavor therepresentation
of the7T-Lvirus-hostjoin shown inFig. 5.
When theflanking DNAsequenceswere
com-pared to the displaced viral sequences (viral
sequences adjacent tothose at the join but no
longer present), various amounts of patchy ho-mologywereobserved (Fig. 4 and 5). Inspection
ofthe 7T-Rsequence revealed an openreading
frameof 24nucleotides (eight amino acids)
con-tinuouswiththe large T-Ag reading frame.This
could encode ahybrid protein of about 76 kilo-daltons(kd) (565virus-encoded amino acids plus 8 flanking DNA-encoded aminoacids [Fig. 6]). Thisis ingoodagreementwith the 75-kd protein detected in 7axT cells (22). Inspection of 7B-R
sequencespredicted theformation of both large and middle T-Ag-related hybrid proteins. The
sequenceresultsindicated that thereare2
flank-ing DNA-coded amino acids in the middle T
reading frame and 34flanking DNA-coded ami-no acids in the large Treading frame(data not
shown). In 7axB cells, only the normal-size large,middle, and small T-Agsaredetected(23).
DNA sequences forpAU-L weredetermined
in eitherdirection fromthe AccI sitelocated 1.3 kb from the left-hand EcoRI site (Fig. 3). The
sequencethrough the AccI sitewasdetermined fromanEcoRI linkerintroducedapproximately
30 bp beyond the AccI site in the flanking cellular DNA(Fig. 3). Thiswasaccomplished by
controlled digestion with exonuclease BaI31 from the XbaI sitelocatedapproximately500bp fromthe AccI site and the insertionofanEcoRI
linker. Approximately 180 bp of the sequence was determined to lie in the region where Py DNA recombined with Rat-1 DNA in pAU-L. Partof thesequencewascomparedtothatofthe
homologous region of7B-L and also tothat of
Py DNA (NTs 4,696 to 4,630) (Fig. 7). In the
case ofa simple recombination event between
pAIJ-L
?ax U
II
-615kt
.-36kb
VOL.44,1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.496.49.450.74.275.2]72 HAYDAY, RULEY, AND FRIED
A
pAU- L EcoRI
pAT
L
Kpnl (770) AccI
1300
Kpnl(4170)
Accl AccI EcoRI
50305440 6pAT
5030 5440 6150
B
pAU-L
I
A.
k~~~~~I
)
}\I_
EcoRI Bgll KpnI XbaI EcoRI' AccI KpnI
7B-L FLANKING DNA IPY
DNA7l
RAT- PY JOIN
- I S NI,.
FIG. 3. (A) Restrictionendonuclease map of pAU-L. TheEcoRIfragment containing the cellular sequences flanking the left end of the 7axB viral insert (see Fig. 1) is drawn in relation to theAccIrestriction endonuclease cleavage sites. (B) Comparison of pAU-L and 7B-L DNAs and strategy for DNA sequencing. For over 1.3 kb from the leftwardEcoRIsite, the map ofpAU-L was the same as 7B-L. A comparison of this region in the two DNAsis shown until the site at which 7B-L becomes joined to Py DNA. Restriction endonuclease cleavage sites were5'-end-labeled, and the arrows refer to the direction and extent of the sequence determined from each labeled site. The sequence through theAccIsite in pAU-L was determined from anEcoRIlinker inserted within approximately 30 bp.
viralorcellularDNAs, the transition from viral tonon-viralsequence in 7B-L would be
expect-ed to coincide with the beginning of homology between the flanking DNA sequence in 7B-L
and pAU-L. Instead, homology did not begin until 37 nucleotides beyond the last nucleotide (NT 4,682) colinear with the viral insert, after which perfect homology between the two se-quencesbegan and continued formorethan100
bp (the extent to which both sequences were
determined). The origin of the 37 bp immediately adjacenttothe viral insertendingatNT4,682is notknown. Thepatchy homology noted (Fig. 7)
may be significant, in which case the first 16
nucleotidesbeyond the joinatNT4,682maybe
largely virus derived; likewise, some of the
remaining sequence maybehost derived. How-ever,there still remain some7to11nucleotides whichappearedtobederived from neither Pyor
pAU-L sequences.
The biological activity ofthesequences cloned
from 7axB and 7axT cells. The presence and expression of integrated viral sequences in Py-transformed cells is assumed to be responsible for cell transformation (34). Cloning the
frag-ments containing integrated viral DNA allows theirbiological activitytobetestedby transfec-tionofRat-1 cells(seeabove). Clones containing the complete coding regions for the Py middle andsmallT-Ags(7B-L, 7B-M, 7T-L)were
capa-ble of transforming rat cells with efficiencies
equaltothatofacompletePy genomeclonedvia theEcoRI site into pAT153DNA(Table2).This result provided assurance that viral sequences werefaithfully maintained during their isolation from cell DNA. Clones 7B-R and7T-Rdid not transform cells. Although 7B-R contained the complete coding region for Py small T-Ag, it may be unable to express this protein because theviralpolyadenylation signals for thismRNA have beenremoved by fusiontohostsequences. However, within 7B-Rthemajor Py early poly-adenylation signal occurs 5' to the small T
coding region. By recloning the Py insert
through theBamHI site inpAT153 DNAitwas
possibletobringthispolyadenylation signal3' to
thesmall Tcodingregion. This reoriented
plas-mid did nottransformratcells.
TheEcoRI fragment of7B-M(thefull-sizePy
genome cloned fromthe tandemPyDNA insert in 7axB cells) was recircularized with T4 DNA ligase and tested for its ability to produce
plaqueson mouse cells. Transfection with 1 ,ug of this fragment produced no plaques. This compares with the105 plaquesper
pLg
formedby
Py DNA aftercleavage and recircularization of the Py genome cloned in the EcoRI site of pAT153. It was subsequently found that 7B-M wasdefectivefor DNAreplicationbecauseofa
singlebase change in the largeTcoding region
(A. C. Hayday, F. Chaudry, and M. Fried,
submitted forpublication). This lesion was tan-J. VIROL.
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.496.73.403.60.274.2]INTEGRATED POLYOMA VIRUS DNA 73
JOIN
7,;1>~
INTEGRATED VIAL SEGWENCEGRGCGGTRCCTRCRCRRCGRR
5' 42U 48
FLANKING DNA ES
TGAGTGCTRTGHARTTCTTTGCCACCRCR
3,
GAGGTCCCTACTGTARATAGAAATATGGC
DSPLACED VIRAL SEQUENCES
46S3
7B-R
RGGGRHTGG9RTGRTTTCTT
GCTGTTAHTTGCTAHGGTTGGTHTTCCTT3,
CRGRRRGTGGGRCCCCRGCTRCCRGTCGC
945
7T-L
TGCRCRTTRCRRRGTRTRTC
TGTRTTTCCTTTRRRRRHTAATARTAATT3i
5' 2848 2838
RCRRRRCRRRCTGTRGGRGCRCTCRCTRG
7T-R
TAHCRGRRRRTGTGCCHRAH
GRACTTRRRGGGTTGGGRRCTCCRTRGGH3
d M2242 2252
CGRCGCRRCRCATRCTTTTTRGRGGRCCR
2281 FIG. 4. Sequences of virus-hostjoins in7B-L, 7B-R,7T-L, and 7T-R DNAs. The sequences flankingthe
integratedviral DNAs are shown above thedisplacedviralsequences(i.e.,viralsequences which are nolonger
present as aresult ofrecombinationbetween viral and cellularDNA).Slopinglines indicatehomologiesof .3bp (withinadistanceof4bp)between theflankingDNAsequences anddisplacedviralsequences. The nucleotide numbersofthePygenome areindicated(30).The sequences of 7B-L and 7B-Rweredetermined fromanalysisof bothstrands, and of 7T-L and 7T-R fromoverlapping fragmentsonthesame strand.
demly repeated inthe PyDNA in 7axB, i.e., it
alsooccurred in thelargeTcoding regionof 7B-R.
DISCUSSION
Inthisreportwedescribeexperiments aimed
atunderstanding the recombination events
be-tweenviral and hostsequenceswhich accompa-ny cell transformation by Py. The integrated viral and adjacent host DNA sequences from
twoPy-transformed cell lines were cloned into
procaryotic vectors. The structures of the
cloned DNA sequences were apparently unal-tered by this process. The flanking host DNA sequences at three of the four virus-hostjoins
werefoundtoexist in a singlecopyin untrans-formedratcells. AnEcoRIfragment containing
oneof these unoccupied sites was also cloned.
TheprimaryDNAsequence wasdetermined for
this clone and for eachof the virus-hostjoins.In
7T L
>INTEGRATEDVIRAL
SEQUENCES J
RARRCRRRGR CTGTRTTTCCT
GGRRRTTRRTGTTTRTTCARTARRCTG-
DISPLACED VIRAL"52IH
2Om
7I
\\\\295
I
SEOUENCES
5 CARRGTRTRT
CTGTRTTTCCT,TTRRRRRRA
TRTRRTTCTGAARTRRR-
FLNKING DNA 328481
1 1
1
1 2j39
SEQUENCESCRRRGTRTRT,CHCRRRRCRRR
CTGTRGGRGCRCTCRCTRGCRRACTGG-
DISPLACED VIRALV 2828 2801 SEQUENCES
FIG. 5. Alternativerepresentation of the virus-host join in clone 7T-L. In this representation (see the text), a virus-virus join (J), NTs2,838 to 2,842, precedes the virus-host join (J) at NT 2,932. Vertical and sloping lines indicatehomologies between Py and 7T-L DNA.
VOL.44, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
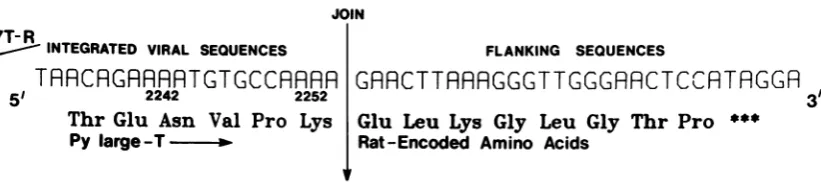
[image:7.496.49.452.60.386.2] [image:7.496.54.451.562.644.2]74 HAYDAY, RULEY, AND FRIED JOIN
R INTEGRATED VIRAL SEQUENCES
TARCRGRRRRTGTGCCRRRR
51 2242 2252
Thr
Glu Asn Val ProLys
Py large T
-FLANKING SEQUENCES
GRRCTTRRRGGGTTGGGRACTCCRTAGGR
3'
Glu
Leu
Lys Gly
LeuGly
Thr Pro+**
Rat-Encoded Amino Acids
FIG. 6. A largeT-Ag-related virus-host fusionproteinpredicted by the DNA sequence across the Py-host join in clone 7T-R. The Virus-hostjoin in 7T-Rinterrupts the coding region for the viral large T-Ag. The cellular sequencesremain openin thelargeT-Agreading frame foranadditional eight amino acids, as shown.
addition, the clones derived from the trans-formed cells were assayed for their biological activity.
Expression of integrated viralsequences. Cells transformed by Py express both the middle and smallT-Ags but not necessarily the full-size Py large T-Ag (19, 21, 23, 27). Recently it has been shown thatexpression of only the middle T-Ag is sufficient for most of the properties of the Py-transformed phenotype (35). Both 7axT and 7axB cells contain at least one unaltered Py coding region for both the small and middle T-Ags, and both of these cell lines produce these proteins (23). The cloned fragments M, 7B-L,and 7T-L, which each encode both small and middle T-Ags, transformed rat cells efficiently.
Neither the 7B-R clone, which contains the
complete coding region for small T-Ag, nor a
modifiedclone with the major viral
polyadenyla-tion signal brought 5' tothe small T-Ag coding
region transformed rat cells. That clones 7B-R and 7T-Rdid not transform cells indicates that the host sequences which are adjacent to the viral inserts inthesecells are non-transforming. 7axT cells do not contain sufficient viral se-quences to encode Py large T-ag. Rather, the sequencedata for 7T-R suggests that alarge T-relatedproteinof76 kdcould beexpressed. An anti-Timmunoreactiveproteinof 75 kd has been detected in 7axT cells (22). The 100-kd anti-T
4882 4662
PY CCTRCRCRCRRCGRRGRGGTCCCTRCTGTRRRTRGRRR
I1111111
I\
I
%" , ,,,...
7B-L CCTRCRCRCRRCGRRTGRGTGCTRTGRRRTTCTTTGCC
4682 1'! .'
pAU-L TTI III I/
immunoreactive protein detected in 7axB cells alsocould not support viral DNA replication due to a mutation in the coding region for the
car-boxy-terminal portionof theprotein (Haydayet al., submitted for publication). The absence of functional Py large T-Ag in these cell lines correlateswith thestabilityof theinserts of viral sequences both uponfurtherpassagein vivo and upon selection for morphological revertants of transformation (2, 23; A. C. Hayday,
unpub-lished data). The DNA sequence of 7B-R pre-dicts large and middle T-Ag-related virus-host fusionproteins.Nosuchproteinsaredetectable
in7axBcells(23). If suchproteinsareproduced,
they may be very unstable,
non-immunoreac-tive,orboth. Alternatively, theirsynthesismay
beblockedat the level oftranscription or
proc-essing. By using the clonedproviral fragments
as probes, it was shown that one copy of the
flankinghost sequences existsunaltered in both
7axB and 7axT cells (Fig. 2). Thus, a haploid
copy ofthe viral transforming gene(s) is suffi-cient to induce transformation, and any theory
of transformation by inactivation of host
se-quencesafterPyinsertion becomes less tenable. Siteand
specificity
ofintegration.
Ithasprevi-ouslybeen shown byrestriction enzyme
analy-sis thatPy DNA is
integrated
in differentchro-mosomalsites in differenttransformed cell lines
(19,21-23). Thequestionariseswhether there is
4642
T TllGCGTi T.I H RGGCGGGRTCiCRG.HIt.H.
111111I
, II III
/,ICF
I
ITLGCRnCRTI.TFI.HC.uHHaC1I-111IHl.I 1
111111111liii111111111111111
IRRTTTTCRTGGTRGRCTGRRTCCTTTRTRTRT AccI
FIG. 7. Analysis of the integration site in pAU-L. The site of recombinant Py and cellular DNA which generated the left-hand join of the viral insert in 7axB is shown by comparing the DNA sequences of Py, 7B-L, and pAU-L. The sequence of the viral insert in 7B-L is colinear with the Py sequences until NT 4,682, as shown.
Sequences flanking the viral insert in 7B-L are not homologous with cellular sequences around the site of integration until4bp before the AccI site, at which point homology between the two sequences abruptly begins andcontinues over the extent that both sequences were determined (100 bp; only 26 bp of this homology is shown). The origin of the37nucleotidesimmediately adjacenttothe colinearextentoftheviral insert in 7B-L is
notknown, butsomeof the sequencesmayhave been derivedfrom small patches of Pyandcellular DNAs, as shownby dotted lines betweenthe sequences.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.496.49.460.75.166.2]INTEGRATED POLYOMA VIRUS DNA 75
TABLE 2. Transforming activitiesofclonedDNAsa
No. of foci Average no. of
DNA
pamdDNA
per0.5ILgof fociper ngofper 0.5 of Py-transforming
plasmidDNA
geneb
7B-L 682 5.6
7B-M 743 7.5
7B-R
7T-L 727 6.9
7T-R
Py-pAT153c 796 8
aThe DNAswere applied to Rat-1
cells
and stained for transformed foci 2 weeks later asdescribedin the text.bCalculatedbyassuming that the oncogenic capaci-tyof each clone lies1.75 kb from theBcIIsite on the Py genome untilthe end of the middle T-Ag coding region.
cPy-pAT153 is an EcoRI-cleaved genome of pAT153 into which EcoRI-linearized Py DNA has beenligated.
any sequence specificity or structural feature commontositesof recombination between viral and host DNA. Forexample, theintegration of retroviral genomes or bacterial transposable ele-ments results in duplication of adjacent host DNA, and transposable elements apparently
havepreferred sites of integration (7, 24, 29). In the cases presented here, the DNA sequences flanking the integrated Py DNA were different and shared no apparentcommon structural fea-ture. This is a consistent finding for other Py-host join sequences (27a; Ruley, unpublished data). Some of the flanking sequences showed homology to displaced viral sequences (7T-L, 7B-L); othersdid not(Fig.4and5). Some ofthe
flanking sequences were A+T rich (e.g., 7T-L,
90% over 20 bp); others were not. Some of the adjacent displaced viral sequences were
purine-CELL
*** ** *****
rich (e.g., 7B-R, 80% over 10 bp); others were not. An extensive hairpin loop structure could be postulated for viral sequences which sur-roundedand includedthe site of recombination of viral and host DNA in 7B-R;for otherjoins this was not possible. In short,
no.
common sequences or structural features were observed at the sites of integrated Py sequences studied here. Similar conclusions werereached in stud-ies on integrated simianvirus 40(SV40)DNA(5, 32).Although no common specificity was ob-served among the different integrated Py se-quences, a unique sequence arrangement was observed foreach complete provirus. Oligonu-cleotides foundat oraround thejoinat oneend
ofthe viral insert were observed in about the correspondingpositionattheotherend(Fig. 8).
Symmetrically placed oligonucleotides have been observed at either end of another Py insert (27a) and at either endof two out of three SV40 proviruses for which the sequence from both ends is available (32). Given the short lengths of sequences involved, the biological significance ofthese symmetries is unclear. However, their occurrence in five out of the six Py and SV40 proviruses for which sequence data from both ends is available suggests that future virus-host
joinsshould beexamined for theiroccurrence.
Mechanism ofintegration. When the flanking
sequences from each cell linewereusedtoprobe
untransformed cell DNA,no commonfragments
weredetected (Fig. 2).Thus, for both 7axB and 7axT, the host sequences which flank the viral inserts were not adjacent in rat cell DNA. This indicates that host DNA sequences have been deleted or rearranged at the site of viral integra-tion. If sequences have been deleted, the mini-mumextent of this deletion is 6.5 kb in 7axB cell DNAand 0.65 kb in 7axT cell DNA. Deletions
VIRUS
CELL
***** ** ***
7RXB TCRTRGCRCTCR TTCGTTGTGTRGGTRCCGC....GGGARTGGRRTGRTTTCTT GCTGTTRRTTGC RGTRTCGTGRGT RRGCRRCRCRTCCRTGGCG.... CCCTTRCCTTRCTRRRGRR CGRCRRTTRRCG
4U2 916
* ** *** *** ***
7HXT RTTRTTTTTTRR RGGRRRTRCRGRTRTACTT....RRCRGRRRRTGTGCCRRRR GRRCTTRRRGGG TRRTRRRRRRTTITCCTTTRTGTCTRTRTGRR.... TTGTCTTTTACRCGGTTTT CTTGRATTTCCC
212 22z26N 2252
FIG. 8. The virus-hostjoinsateitherend of thePy sequencesin 7axB and 7axTcells, respectively. Asterisks indicate thesymmetrical sequenceatthe endsof each viral insert(i.e., inwards from flanking to viral sequences). Numbers arePy nucleotide numbers(30).
VOL.44,1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.496.56.454.496.643.2]76 HAYDAY, RULEY, AND FRIED
have also been observed around the sites of recombination betweenSV40and rat cell DNAs (5);SV40 and adenovirus DNAs (40); SV40 and monkey DNAs in anSV40-defective variant (9);
anddifferentsequencesofintegratedPy DNA in
Py-transformed cells (Ruley, unpublished data).
Thus,deletionsmaybe a general feature
accom-panyingrecombinationbetween twolargely dis-similar sequences in eucaryotic cells.
The Py DNA in 7axB cells was flanked by
single-copyrat DNA. AnEcoRIfragment of cell
DNA,homologous to theflanking DNA in 7B-L, was cloned from syngeneic, untransformed rat cells.Thiscloned DNA(pAU-L) was compared to 7B-L. For1.3 kb from one of theEcoRI sites, 7B-LandpAU-L had identical cleavagepatterns
as determined by analysis with more than a
dozen different restriction enzymes (Fig. 3). Within this region, 7B-L and pAU-L DNA showedidentical sequences for at least 100 bp. For 37 bp adjacent to the virus-host join, the sequence of 7B-Ldiffered from that of pAU-L (Fig. 7). This dissimilarity may be explained in
one of four ways. (i) pAU-L has been cloned
fromthealleleopposite that towhich viral DNA
integrated. If so, the sequencedivergence just at
the virus-host join would be remarkably fortu-itous. (ii) Py DNA transduced a stretch of se-quencewhichistoosmall for its true homolog to bedetectedby hybridization. (iii)The sequences
flanking the viral sequences (7B-L) represent a
patchwork of viral and host sequences, possibly constructedbytemplateswitchingof DNA poly-merase(s) filling agapped duplex at the site of
integration.Some form of template switching for
bacterial DNApolymerase haspreviously been invoked for aberrant replication events (1, 8).
According to this model, parts of the flanking
DNA areviral inorigin,and becauseofthis the
flanking DNA shows homology to viral
se-quences. (iv) Recombination atthe site of
inte-gration has involved other host sequences as
well as theviral sequences.
Asyet, the dataareinsufficientto
distinguish
between these four alternatives. The
flanking
DNA in7B-L did show somehomologytoviral sequences, and thishomologywasgreater inthe 40 bp around the virus-host
join
that it waselsewhere. However, there were
regions
of flanking DNA which were not easily shown tobe derived from either viral or cellular
se-quences. Around the sites ofother virus-host joins, the flanking DNA showed even greater homology to viral DNA than was the case for 7B-L: e.g., Py DNA-rat DNA 7T-L, this study
(Fig. 5);SV40 DNA-rat DNA(32); SV40
DNA-monkeyDNA(9, 16).Onlyananalysisof
unoc-cupiedsites fromcasessuch asthesecanreveal
whether virus-host patchworks or viral
trans-duction may be involved in integration events.
When there isnodetectable homology between viralandflankingDNA (e.g.,7B-R,this study), the virus-host join mayhave been formed by a direct recombination event, or there may have been negligible template switching in filling a
gapped duplex to produce a clean transition from viral tocellular sequences.
From the results presented here it would appear that Pysingle-stranded regions maypair to single-stranded regions of host DNA as a prerequisite to integration. The virus-host mis-matched duplex could then be converted to a homoduplex by the repair systems of the host cell, which in some cases could result inregions ofpatchwork between viral and host sequences. If the two joinsof the viral sequences to the host sequences took place as separate events, the joins could bein nonadjacent regions of the host DNA, causing a rearrangement of host se-quences. A possible candidate for stimulating theopening of the host DNA duplex for entry of the viral strands would be the Py A gene product (large T-Ag) since, after virusinfection, a
func-tionalAgeneisrequiredforefficient
transforma-tion(10-12). The study of more virus-hostjoins and unoccupied-site DNAs should lead to a greater understanding of the mechanism of Py integration.
ACKNOWLEDGMENTS
We thank Brendan Davies and Moira Griffiths for their excellenttechnical assistance and GinaYiangouand Shenaz
Somanifor help in thepreparationof thismanuscript.
LITERATURECITED
1. Backman, K., M. Betlach, H. W.Boyer, and S. Yanofsky. 1978.Geneticandphysicalstudies on the replication of
ColEl-typeplasmids. ColdSpringHarborSymp. Quant. Biol. 43:69-76.
2. Basilico, C., S. Gattoni, D. Zouzias, and G. DeliaVale. 1979. Loss ofintegrated viral DNA sequences in Py-transformed cells is associated with an active viral
A-function. Cell17:645-659.
3. Benton, W. D., and R. W. Davies. 1977. Screening Agt recombinant clonesby hybridization to single plaques in situ.Science196:180-182.
4. Birnbolm, H.,andJ. Doly. 1979. Arapidalkaline extrac-tion procedure for screeningrecombinant plasmid DNA. Nucleic Acids Res.7:1513-1523.
5. Botchan, M.,J. Stringer, T.Mitchison,andJ. Sambrook. 1980. Integrationand excision of SV40 DNA from the chromosome ofatransformedcell.Cell20:143-152. 6. Brutag,D.L., J. Clayton, P. Friedland, and L. H. Kedes.
1982.SEQ: a nucleotide sequence analysis and recombi-nation system. Nucleic Acids Res. 10:279-294. 7. Calos, M., and J.Miller. 1980. Transposable elements.
Cell 20:579-595.
8. Clark,A.J.,andM.R. Volkert.1978.A newclassification ofpathwaysrepairing pyrimidinedimerdamageinDNA, p. 57-72. In P.C.Hanawalt,E.C.Friedberg,andC. F. Fox (ed.), DNA repair mechanisms. Academic Press,
Inc.,New York.
9. Dhruva,B.R.,T.Shenk, and K. N. Subramanian. 1980. Integration in vivo into SV40 DNA of a sequence that resemblesacertainfamilyofgenomic,interspersed, re-J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
INTEGRATED POLYOMA VIRUS DNA 77
peatedsequences.Proc.Natl. Acad. Sci. U.S.A. 77:4514-4518.
10. DiMayorca, G.,J.Callender, G.Marin, and R. Giordano. 1969. Temperature sensitivemutantsofpolyomavirus. Virology38:126-133.
11. Eckhart, W.1969.Complementation and transformation bytemperature-sensitivemutantsofpolyoma virus.
Virol-ogy38:120-125.
12. Fried, M. 1965.Cell-transforming abilityofatemperature sensitivemutantofpolyomavirus. Proc. Nati.Acad. Sci. U.S.A. 53:486-491.
13. Graham, F. L.,and A.J.vander Eb. 1973. Transforma-tion ofratcells by DNA ofhumanadenovirus5.Virology 52:456-467.
14. Griffin, B. E.,M.Fried, and A. Cowie. 1974. Polyoma DNA-a physical map. Proc. Natl. Acad. Sci. U.S.A. 71:2077-2081.
15. Grosveld, F.G.,H. H.-M. Dahl,E.deBoer, and R. A. Flavell.1981. Isolation of 3-globin-related genesfroma
humancosmid library.Gene 13:227-237.
16. Gutal, M. W., andD.Nathans.1978. Evolutionary vari-antsofSV40: cellular DNAsequencesandsequencesat recombinantjointsofsubstitutedvariants. J. Mol. Biol. 126:275-288.
17. Hassel,J. A., W.Topp,D.Rifkin,and P. Moreau. 1980. Transformation ofratembryofibroblastsbycloned
poly-oma virus DNA fragments containing only part of the early region. Proc. Natl.Acad. Sci.U.S.A. 77:3978-3982. 18. Hayward,G.S.,andM.G. Smith.1972. The chromosome ofbacteriophage T5. Analysisof thesingle-strandedDNA fragments by agarosegel electrophoresis. J. Mol. Biol. 63:383-395.
19. Israel,M.A.,D. F.Vanderryn,M. L.Meltzer,andM. A.
Martin. 1980. Characterisation of polyoma viral DNA
sequencesinpolyoma-inducedhamster tumourcell lines. J.Biol. Chem.255:3798-3805.
20. Lania, L.,M.Griffiths,B.Cooke,Y. Ito,and M. Fried. 1979. Untransformed ratcells containingfreeand inte-gratedDNAofapolyoma non-transforming(hr-t)mutant.
Cell18:793-802.
21. Lania, L.,D.Gandini-Attardi,M.Griffiths,B.Cooke,D. deCicco,and M. Fried. 1980. The polyomavirus 100K large-T antigen is not required for the maintenance of transformation. Virology 101:217-232.
22. Lanla,L.,A.Hayday,G.Bjursell,D.Gandini-Attardi,and M.Fried. 1980.Organizationandexpressionofintegrated polyoma virus DNA sequences in transformed rodent cells. ColdSpringHarborSymp. Quant.Biol. 44:597-603. 23. Lania, L., A. Hayday, and M. Fried. 1981. Loss of functional largeTantigenand free viral geno-^- from cellstransformedin vitroby polyomavirusafterpassage invivoastumorcells. J. Virol.39:422-431.
24. Majors, J. E., and H. E. Varmus. 1981. Nucleotide
sequencesathost-proviral junctionsformousemammary
tumourvirus. Nature(London) 289:253-258.
25. Maniatis, T.,R.Hardison,E.Lacy,J. Lauer,C. O'Con-neil,D.Quon,G. K.She,and A.Efstradiadls. 1978.The
isolationof structuralgenesfromlibraries ofeukaryotic
DNA.Cell15:687-701.
26. Maxam, A. M., and W. Gibert.1977. Sequencing
end-labelled DNA with base-specific chemical cleavages.
Proc.Natl.Acad. Sci. U.S.A. 74:560-563.
27. Novak, U., S.Dilworth,andB. E.Griffin. 1980. Coding
capacity of a35%fragmentof thepolyomavirusgenome is sufficient to initiate and maintain cellular transforma-tion. Proc.Natl.Acad.Sci. U.S.A. 77:3278-3282. 27a.Ruley,H. E.,L. Lania, F.Chaudry, and M. Fried. 1982.
Use of a cellular polyadenylation signal by viral tran-scripts inpolyoma virus transformed cells.NucleicAcids Res.10:4515-4524.
28. Sanger, F., and A. R. Coulson. 1978. The use of thin acrylamidegels for DNA sequencing. FEBS Lett. 87:107-110.
29. Shlmotohno, K., S. Mizutani, and H. M. Temin. 1980.
Sequences of a retrovirus provirus resemble those of b cterial transposable elements. Nature (London)
28 -550-554.
30. Soeda, E., J. Arrand, N. Snolar, J. Walsh, and B. E.
Griffin.1980. Coding potential and regulatorysignals of thepolyoma virus genome. Nature (London) 283:445-463. 31. Southern, E. 1975.Detection of specific sequences among DNAfragments separated by gel electrophoresis.J. Mol. Biol. 98:503-517.
32. Stringer, J. 1981.Integrated simian virus 40 DNA: nucleo-tide sequences at cellular virus recombinant junctions. J. Virol.38:671-679.
33. Tiemeier, D. C., S. M. Tilghman, and P. Leder. 1977. Purification and cloning of a mouse ribosomal gene frag-mentincoliphagelambda. Gene 2:173-191.
34. Tooze, J. (ed.) 1980. DNA tumor viruses. Molecular
biology oftumor viruses, part 2. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
35.Treisman,R., U. Novak, J. Favaloro, and R. Kamen. 1981.
Transformationofratcells by an altered polyoma virus genome expressing only the middle-T antigen. Nature (London)292:595-600.
36. Twigg, A. J., and D. Sherratt. 1980. Trans-amplifiable copy number mutants of plasmidColEl.Nature (London) 283:216-218.
37. WaalwUk,C., and R. A.Flaveel.1978. An isoschizomer of
HpaII whichcleaves both unmethylated and methylated
HpaII sites.Nucleic Acids Res. 5:3231-3236.
38. Weaver, R. F., and C.Weissmann. 1979. Mapping of RNA by a modification of the Berk-Sharp procedure: the 5' terminiof15S ,B-globin mRNA precursor and mature 10S
P-globinmRNA have identical map co-ordinates. Nucleic Acids Res. 7:1175-1193.
39. Wigler,M., R. Sweet, G. K.Sim, B. Wold, A.Pellicer,E. Lacy, T. Maniatis, S. Silverstein, and R. Axel. 1979.
Transformationof mammalian cells with genes from pro-karyotes and eupro-karyotes. Cell18:777-785.
40. Zain, B., and R. Roberts. 1978. Characterisation and sequence analysis of a recombination site in the hybrid virusAd2+ND,.J. Mol. Biol. 120:13-31.
VOL.44, 1982
on November 10, 2019 by guest
http://jvi.asm.org/