0022-538X/81/080510-09$02.00/0
Application
of Arrhenius Kinetic
Theory
to Viral
Eclipse:
Selection of
Bacteriophage
4X174 Mutants
N. L.INCARDONA
DepartmentofMicrobiology&Immunology, University ofTennessee Centerforthe Health Sciences, Memphis, Tennessee 38163
Received17February 1981/Accepted15April 1981
Analysis of the
bacteriophage
4X174eclipse period
in terms of Arrheniuskinetic theorysuggeststhefollowing hypothesis: mutantsshould exist with two
concomitant
physiological
characteristicsastheirphenotype.
Theseare aneclipserate lower than that of the wild type at
permissive
temperatures for plaqueformation and an
eclipse
rate toolow atlower temperatures topermit
plaquedevelopment. Thus, enrichment ofa
mutagenized
viruspopulation
formutantsthatfailto
eclipse during
ashortperiod
atpermissive
temperatures shouldyieldeclipse mutants with the cold-sensitive
(cs;
nonpermissive
temperature, 25°C),andnotthetemperature-sensitive (ts; nonpermissivetemperature,
420C),
plaquephenotype.Inseveral
trials,
thefrequency of thecsphenotype
inthepopulation
increased from less than 0.2%to between2 and 4% after the enrichment step,
whereasthe
frequency
of thetsphenotype
remainedunchanged (less
than0.2%).Moreover,80% of thesecs mutantshave
eclipse
ratesthatare3-to40-fold lowerthan that ofthe wildtype atboth
370C
and250C.
Thesuccessfulapplication
ofthe Arrhenius theory to 4X
eclipse
mayprovide insights
into the molecularmechanismwhereby the
4X174
genomeisdelivered into the hostcell. Since theeclipse
kinetics of other nonenveloped viruses are similar to those of4X174,
kinetictheorymaybe
broadly applicable
intheselection and characterization ofviraleclipsemutants.
Since viral
nucleocapsids
areconstructed fromprotein subunits and nucleic acid molecules,
noncovalent interactions makeasignificant
con-tribution to the structural stability of viruses
and
play
animportant
role in theirreplication.
The
importance
of noncovalent bonds in virusassembly has been
extensively
documented (3),and the principles that govern their formation
arebeginningtoemerge (21). However, the
dis-ruption of these interactions must also play a
part invirus
replication
atthepoint where viralgenomesgainentryintohostcells.Yetevidence
for theprecise role played by noncovalent
inter-actions
inthisdelivery
of viralgenomesremainsvery sketchy
(1, 6, 12-15, 18, 27, 29, 30). This ispartlydue to the dearth ofadequate means for
interrupting theprocess todetermine the
num-berandnatureof theinternediate steps. Advan-tage has been taken of the fact that, at low temperatures, many viruses adsorb but fail to enterthe
eclipse
phase (17, 18, 22, 24) and that some viruses requiremetabolically
active cellsforcompleteentryintothe cytoplasm (4, 16, 19).
However, there isa need for selectiontechniques
specificforconditionallylethal mutants that are
defectiveinthe steps subsequent to adsorption
and associated with the entry into the eclipse
phase of the replication cycle. Since the viral
genomeis dissociated fromatleastsomeof the capsid proteins during eclipse, it is likely that
mutants with altered noncovalent interactions
will be generated bysuch procedures.
Structural and kinetic studies onthe eclipse
ofbacteriophage
4X174
provide a backgroundfor thedesign of selection procedures that are
specific for eclipsemutants. One can single out
for study thesteps associatedwith the loss in
infectivity that occurs at the beginning of the
eclipse period bysynchronizing the infection in
starvation buffer at
150C
and shifting to a higher temperature toinitiate theentry into theeclipsephase.Moreover, since EDTA partially disrupts
the outer wall of the cell, viral nucleocapsid structures canberecovered from infected cells
between the attachment and complete DNA
injection steps, inclusively. Use of these
tech-niqueshasrevealed that at the low temperature,
4X174
adsorbs to starvedcells
but does notbegintoeject itsDNA. Therefore, the recovered
virusremainsinfective. Shifting thesevirus-cell
complexesto temperatures above
200C
instar-vation bufferresults in the partial ejection of the
510
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 39, 1981
DNA with the concomitant loss ofinfectivity
that defines thebeginning of theeclipseperiod.
Addition of nutrientstothe starvation buffer is
required for complete injection of the viral ge-nome (22).
The kinetics ofthe lossin infectivity
associ-ated withthe partial ejection process (for
con-venience, termed the eclipse reaction) can be
followed by diluting virus-cell complexesmade
at
150C
into starvation bufferat highertemper-aturesand then stopping the reactionbydiluting
into EDTAat
00C
(23). Therateof thiseclipsereaction for wild-type
OX
and a cold-sensitive(cs) eclipse mutant, cs7O, has been examined
over awide temperature range. At the
nonper-missive temperature
(2500)
usedtoselect thecsmutants of
OX,
cs7O hasaneclipse ratethatisessentiallyzero.This is thedefectivestep that is
responsible for itscsplaque phenotypesince its
eclipse and latent periods, as well as its burst
size,arethesameasthose of thewildtypewhen
virus-cell complexesmade at
370C
in starvationbufferareshifted tonutrient broth at
250C
tocomplete the infection (28). However, the
un-usualpropertyof thismutantis thelowereclipse
rate at all temperaturesbetween 30 and
400C,
the permissive range (8). Analysis of these
ki-netic properties in terms of Arrhenius theory
providesatheoretical basis for the lowereclipse
rate of cs7O at permissive temperatures.
Fur-thermore, since the
theory requires
that sucheclipse
mutantshave the csplaque
phenotype,specific procedures
have beendevised forselect-ing thesemutants.
The
cs7O mutation is believedtobe ingeneF(26).
MATERIALS AND
METHODS
Viral and bacterial strains. Wild-type 4X174
originallycamefrom thelaboratoryof R. L.
Sinshei-merandwassubsequentlyplaque purified.High-titer stocksareroutinelygrownbyinfecting25-mlcultures ofEscherichiacoliC/1atacelldensityof
108/ml
with randomlyselectedsingleplaques. Lysisisinhibitedby the addition of3mlofsterile2MMgSO4atthetime of infection (8). After3hof incubation at370C,the cellsarecollectedbycentrifugation,suspendedin5ml ofpH9.5saturated sodium boratecontaining0.05M EDTA, and storedat2°Covernightforlysis.Stocks preparedin thismanneralways give plaquesat420C. E. coli C/1 and C/+X have been previously de-scribed(11).Plaque assay. Theprocedure is thesame asthat previouslydescribed(10),withmodificationstoreduce the size of theplaquesforcountingwithaNew Bruns-wick BiotranIIAutomatic ColonyCounter. For the wild type, the top agar concentrationis1.1%;1.0mlof alate-log-phaseculture of E. coliC/1is used perplate,
and theplatesareincubated for1hat370Cand then overnightat230C.With thecsmutants,only0.2ml of
SELECTION OF ECLIPSE MUTANTS 511
bacteria is used with 0.7% top agar. The plates are incubated at 37°C until plaques are faintly visible (between 2 and 4 h) and then shifted to 23°C for overnight incubation.
Chemical mutagenesis. Several high-titer stocks ofwild-type4X174were diluted to1010PFU/ml and mutagenized with hydroxylamine as outlined by Tess-man (31). After incubation at370Cwith 0.10 M hy-droxylamine for7 min,the survivors (20%) were di-luted at least 100-fold in ice-cold medium to quench the chemical mutagenesis. Then an appropriate vol-umeof the diluted survivors was added to a culture of E. coliC/1 at a multiplicity of less than 0.01. The conditions of the infection and lysis were identical to those described above for the high-titer stocks. The low multiplicity was used to reduce the probability of multiply infected cells, and an additional 50-fold dilu-tion ofthe mutagen was also accomplished at this step.
Selection ofmutants. The above-described ly-satesof the survivors' progeny(10'0to 10'lPFU/ml) werediluted with starvation buffer (23) to 109 PFU/ ml and incubated at370Cfor 10 min. At thattime, a 0.1-ml samplewasadded to 0.9 ml of E. coli C/1 in starvation buffer at 150C and stirred for a 30-min adsorptionperiod. The cellsweregrown to2 x108/ml inKC broth (23), washed once with starvation buffer, and concentratedto109/ml.Atthe endof the adsorp-tion period, the reaction flask was transferred to a waterbathshaker at370Cfor a 10-min eclipse period, after which the reactionwasquenched by a 100-fold dilution into ice-cold EDTA elutionbuffer (22). After 10min, the cells were removed by centrifugation at 12,000 xgfor 10 min, and the supernatant solution wascarefullyremovedand stored at20C.At a conven-ienttime, the solution wasplatedonE. coli C/1 at
370C, and individual plaques were transferred with
sterile toothpicksto platesto testfor the cs (2500)
and ts (4200) phenotypes. Eachmutant was plaque
purified several times and retested for thetwo phe-notypes.
Invitro transformation reaction.Virus stocks with titers between5x109 and1x 1010PFU/mlwere diluted 100-fold into starvation buffer maintained at 37.00 + 0.010Ctoinitiate the transformation to the form which adsorbsat low temperatures. At various timeintervals, sampleswere withdrawn and diluted 100-fold into tubescontaining E. coliC/1 in starvation bufferateither2 or150Cataconcentration of2to3
x109/ml.Aftera15-minadsorptionperiod,each
sam-plewascentrifugedfor 1 min inanEppendorf3200 centrifuge. The supernatant solutions were main-tained in ice until the concentration of unadsorbed viruswas measuredbyplaque assay.The zerotime pointwasobtained bydilutingthestock into buffer maintainedinice. Asamplefrom the zero timeflask and thereaction flask(afterthe lasttimepointwas
taken)wasalso diluted intostarvation buffer without cellsas a measureof thermal inactivation of the virus. Nosignificantinactivationwasdetected in anymutant
examined,sothesetwovalueswereaveragedandused tocalculate theunadsorbed fraction.
Invivoeclipsekinetics. Theprocedureof New-bold and Sinsheimer (23) wasused withtwo
on November 10, 2019 by guest
http://jvi.asm.org/
cations. First,thevirus-cell complexes werenot
col-lected after theadsorptionstep.However,thelevelof
unadsorbed viruswasalwayscheckedby
centrifuga-tion of asample atthe endof theadsorption period
and maintainedbelow25%byanappropriate choice
of thepreincubation timeat37°Candadsorptiontime at15°C.Second,theeclipsereactionwasinitiatedby
either a 10- or 100-fold dilution of the adsorption
mixture. The corrected rate constant for the faster component of theeclipserate curve wasobtainedas previously described(8).
RESULTS
Theoretical
analysis.
Historically,
Arrhen-ius kinetic theory was the first successful
at-tempt to account for the fact that chemical
reactionratesincrease with
increasing
temper-ature. The
relationship
between the ratecon-stant, k, and the absolute temperature, T,
(ex-pressed inits
logarithmic form)
is:Ink =InA_-E ()
R\T
Here
E.,
the activation energy,isobtained fromtheslope of the linear
plot
oflog
kversus1/T,
and the preexponential
factor, A,
comes fromthe
intercept
with theyaxisatinfinite temper-ature (R is the ideal gas constant).Although
recenttheories haveattemptedtorelate thetwofundamental parameters,EaandA,tothe
struc-tural properties of atoms and
molecules,
theabove form will be sufficient for
application
toviral
eclipse.
One needsonlytorealizethatnu-mericalvalues for thesetwoparameters are
de-terminedby the molecularstructureofthe
reac-tantsandthe activated
complex,
aswellas themechanism ofthereaction. TheArrhenius
equa-tion, therefore, reveals the contributions that
thesestructuralproperties maketothereaction
rate.
Inthe caseof
4XX174,
the kinetics associated with theeclipse
period ofwild-type
virus and theeclipse
mutant,cs7O,
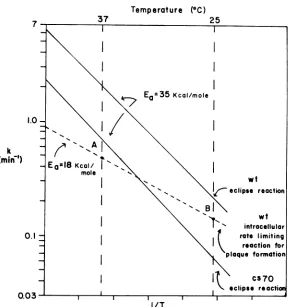
can be analyzed intermsof theArrhenius equation.Concentrating
firston theviral
eclipse
step, theparallel
solidlines in Fig. 1 are the Arrhenius plots for the
wild-type andcs7O eclipse reactions,whose rate
constantshave beenmeasured between 30 and
400C (8). For the purpose ofthis analysis, the
experimental
lines
have beenextended over awidertemperature rangetoinclude the nonper-missive temperatures of the ts and cs plaque mutants of
OX.
IntermsoftheArrheniusequa-tion, the cs7O mutant eclipse reaction has the
sameactivation energy (slope) as does the wild
type,but it has athreefold-lower preexponential
factor (y intercept) (8). This means that the
threefold-lower eclipse rate ofthe mutant
ob-served in the permissive temperature range
should also
apply
at250C,
thenonpermissive
temperaturefor
plaque
formation(Fig. 1).Also
important
totheanalysisarethesimilar-ities between cs7O and wild-type intracellular
development after the eclipse step. Since the
mutantand the wildtypehave thesameeclipse
and latent
periods
at400C,
and at250C
whenashift from the permissive temperature occurs after the eclipse reaction (28), no intracellular
stepsaftercomplete DNAinjectionare
signifi-cantly
impairedin the mutant. Thus,therate-limiting
stepintheintracellulardevelopmentof the mutant is the same as that for the wildtypeat all temperatures between 40 and250C, and
only
oneArrheniusplotisrequiredtodefine thetemperaturedependenceofits rate(dashedline
in Fig. 1). A comparison of the rates for the
intracellular and eclipse reactions is necessary
tounderstand why the csplaquephenotype is
theoretically linkedtothelowereclipserate at
permissive temperatures. Theactivation energy
for thisintracellularreaction wasestimatedfrom the three- to fourfold increase in thelength of
the eclipse period between 40 and 250C
(28).
However,sincethereis not anabsolute valuefor
the rateof this reactionat even onetemperature,
one mustpostulateavaluefor the
preexponen-tial factor. This will determine where the plot
interceptstheyaxis and theposition oftheplot
in
Fig.
1relative to the twoplotsfor theeclipse
reactions.
The value for A can beestablished bymaking
onesimplepostulate: the intracellularreaction
is the
rate-determining
step for plaqueforma-tion. If the eclipse rateisgreater thantherate
oftheintracellular reaction, plaques will form.
If it isless, theuninfected cells surrounding the
infectedcell canestablishsurviving colonies
be-fore the virus burst, and no plaques will be
visible. Therefore, the rateof the intracellular
reaction must be less than both the cs7O and
wild-type eclipse rate at 370C since both give
plaquesatthis temperature. However,at
250C,
therateof theintracellular reaction must beless
than only the wild-type eclipse ratesince only
the wildtype givesplaquesatthisnonpermissive
temperature. Thisreasoningthenrequiresthat
the Arrheniusplotfortheintracellular reaction
inFig. 1must crossbelowbotheclipse reaction
lines atthe
370C
ordinatebutbetweenthe twoeclipse linesatthe250C ordinate,thus
establish-inganapproximate value forA.
Tosummarizethe experimental basis of Fig.
1, the twoArrheniusplots forthecs7Oand
wild-typeeclipse reactionswereestablishedfrom
ex-perimentally measuredrateconstantssothat no
assumptionsarerequiredtoestablish the values
for
E.
and A (8). Anestimateforthe activationenergyof therate-limitingreaction for the
on November 10, 2019 by guest
http://jvi.asm.org/
SELECTION OF ECLIPSE MUTANTS 513 Temperature (OC)
k
(minf')
I/T
FIG. 1. Theoretical basis for the cs plaque phenotype of4X174 eclipse mutants. The solid lines are extrapolations of the experimentallydeterminedArrhenius plots for the eclipse reactions of the wild type and thecs7O eclipsemutant(8).The dashed line is a postulated Arrhenius plot for the rate-limiting intracellular reaction based on the measured eclipse periods at 40 and 25°C (28). Since thecs70 eclipse rate is greater than therateof the hypothetical intracellular reaction at 37°C, plaques areformedat this temperature. However, at25°C,theeclipse reaction ofcs7Obecomes rate limiting, and the uninfected cells outgrow the virus so that theplaquesare notvisible. wt, Wild type.
cellulardevelopment of bothmutantand
wild-typevirus wasobtainedfromexperimental data
at twotemperatures (28).Only the
preexponen-tial factor for this intracellular reaction has a
postulatedvalue, based onthe hypothesis that
therateofintracellular
development
of the virusis therate-determiningstepin
plaque
formation.Test for
predicted
plaquephenotype.
Since thevalidity of
applying
Arrheniustheoryto 4X
eclipse
rests on asimple,
butcrucial,
assumption,one must find logicalconsequences
that can be tested by experimental data. The firstprediction can beillustrated byan exami-nationofFig. 1in lightof the abovepostulate.
Mutantsshouldexist witheclipseratesthatare
lessthanwild-type valuesatpermissive
temper-atures, such as
370C.
For this to occur, the mutation mustresultinachangeinEa orA orbothsuch that theArrheniusplot forthemutant
eclipse reactioncrossesthe
370C
ordinate belowthewild-type
eclipse
line. Those thatcrossboththe 37 and
250C
ordinateabove the plot for theintracellular reaction will give plaques at
all
temperaturesbetween25and
420C
andwillbeindistinguishable fromthewild type in the
typ-icalscreenfor thecsandtsplaque phenotypes. However,thosethat crossabove the
intracellu-lar lineat
370C
butbelowit at250C
mustalsocrossthe
420C
ordinate above theintracellularreaction
plot.
Thus, these latter mutants willyield
plaques
at370C
and have the cs,butnotthets,
plaque phenotype.
Those thatcrossboththe37and
250C
ordinate below theintracellularline will be lost if 37°C is used to obtain the
plaquesforscreeningattheothertwo
tempera-tures.
A testofthispredictionisa
procedure
which enriches a mutagenizedpopulation
of 4X foreclipsemutantsthat formplaquesat
37°C,
fol-lowedbyascreenforboth thecsandts
pheno-types. If theprediction
isvalid,
thereshouldbe alarger
increase in thefrequency
of the csVOL. 39, 1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.116.405.72.379.2]phenotype in the population. Twotrials of the following schemewereperformed.After
chemi-cal mutagenesis, the survivors weregrownata
lowmultiplicitytoexpressthemutantgenotype in the capsid proteins. Anenrichment for
mu-tantswithalowereclipseratewasaccomplished
byadsorbing the mixed population ofprogeny
onto sensitive cells at 150C and shifting the
reaction flaskto37°Cfora10-mineclipseperiod.
The complexeswere then diluted into ice-cold
EDTA-borate elution buffer to dissociate the unecipsed virus fromthe cells. Over 99% of the wild-type virus, and those mutants with an
eclipse ratethe same asthat of the wild type, areinactivated in thistime interval at370C (8), butasignificant fractionof thepotentialeclipse
mutant population is still infectious due to a
lower rate of eclipse. After centrifugation to
remove all cells that may contain completely
injected
4X
DNA, the unecipsed populationwasthen screened for thecsandtsphenotypes by platingat370Candtesting individual plaques at25 and420C.The results(trialsAlandA2 in Table 1)clearly showatleasta10-foldincrease
in only the cs phenotype. In a third trial, the
enrichment stepwas a45-mineclipseperiodat
250C (thereasonforselecting thistemperature willbe discussedbelow), followed bya screenof plaques obtainedat370Cfortheplaque pheno-types. Since the desired mutantshave alower
eclipseratethan the wildtypeat250C (Fig. 1), theycanbe enriched in thepopulation by
inac-tivating the wild type during an eclipse period
at250C, as well asat370C. Ofcourse, theycan
be recovered by plating the survivors at370C
(Fig. 1). Trial B in Table 1 shows that the results
are again a 10-fold enrichment ofonly the cs
phenotype. Thus, the prediction concerning the plaque phenotype is confirmed.
Test for predicted classes ofeclipse
mu-tants.Asecondpredictionarises whenone
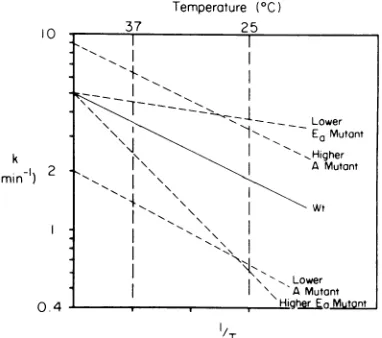
con-siders all of the possible ways of altering the values of Ea and Atoobtain the Arrheniusplots forpossibleeclipsemutants.InFig.2,four ofthe eight possibilities are imustrated and represent the caseswhere onlyone of theArrhenius
pa-rameters is changed from the wild-type value. Since cs7O is an example ofa single mutation
altering onlyoneof the two parameters, there is
precedent forinitially considering only the sim-plestcasesinFig. 2. For thetwo classes above
the wild-type line,one candesignanenrichment
step that takes advantage of their increased eclipserateoverthat of the wild type; but
nei-ther class is expectedtohave either thetsorcs
plaque phenotype for screening the enriched population (compare Fig. 1 and 2). As for the two classes with the eclipse rates slower than that of the wildtype,theenrichment and
screen-ing procedures usedtoobtain thecsmutantsin Table 1 should select for both classes. Thus, examples of both the "higher
E."
and"lower A" mutantsshould bepresentineach population ofcsmutants (compare Fig.1and2).
Totestthisprediction,apreliminary
classifi-cation was obtained by measuring the eclipse
Temperature (°C)
/T
FIG. 2. Theoretical basis for the existence of dif-ferent classes of4XM74eclipsemutants.Thesolid line isanextrapolationoftheexperimentally determined Arrheniusplot for the eclipse reaction ofthe wild-typevirus (8). Thedashed linesareexamples ofsome
postulated classes of eclipse mutants with altered valuesofeithertheArrheniusactivationenergy,Ea, orpreexponentialfactor,A.Wt, Wildtype.
TABLE 1. Effectiveness of eclipse mutant screen
Procedure Frequency of phenotype No. of eclipse
Growth No.tested mutants/no.of
temp(°C)
Enrichment
cs(2500)
ts(420C) testedControl 37 None 550 0.002 <0.002 NDa
Al 37 Uneclipsed at370C 100 0.02 <0.01 1/2
A2 37 Uneclipsed at
370C
600 0.037 <0.002 4/6B 37 Uneclipsed at 25°C 450 0.040 <0.002 6/6
C 37 Unadsorbed at
150C
400 0.032 0.002 NDaND, Not
determined.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.259.450.288.457.2] [image:5.503.60.449.556.641.2]VOL. 39, 1981
rate at 37and 250C forarepresentative group
from each of the screening procedures. Six of the
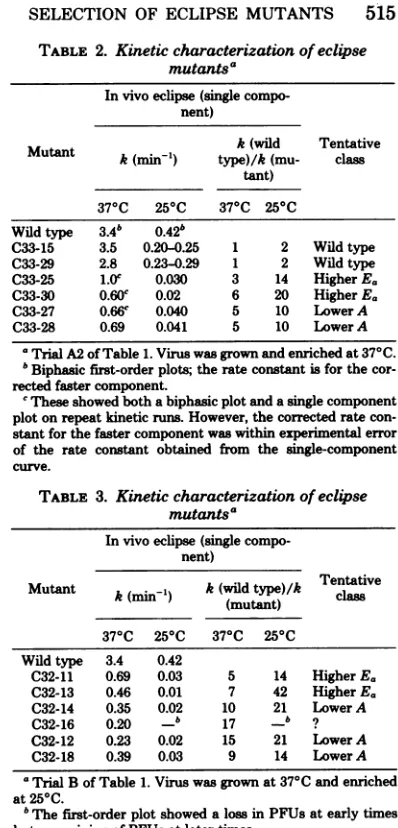
csmutantsfrom the370Cenrichment (trial A2 inTable 1) arecompared with the wildtypein Table 2. The kineticmeasurementswere made
with high-titer crude lysatessinceapreliminary
experiment with wild-type 4X174 (data not
shown) gave an Arrhenius plot with only a
slightly higher A but withan
E.
identicaltothat obtainedwith purified virus. For themost part,asinglekineticrun wasmadeateach tempera-ture so thatmoremutants could be examined. This necessitates a conservative approach in
comparingmutant and wild-type eclipse rates,
sinceuncontrolled variations in cell properties occasionallyleadtolarge differences inratesfor thesamevirus sample. Althougha
reproducibil-ity of 10to 20%canbe obtained from multiple
determinations (8), tentative classification can
be madebyconsidering only ratios ofwild-type to mutant rates that are greater than two as
significantly different. By this criterion,atleast two-thirds of the cs mutants in Table 2 are
defective intheeclipsereaction. Moreover,
ex-amples of both classesarepresent. C33-25and -30probably belongtothehigher Eagroup,since
their ratiosat250Caresignificantlygreaterthan thoseobservedat370C.Onthe otherhand,
C33-27and -28 probablyrepresentthe lower A class in that their ratios at both temperatures are
withinafactor of 2. InTable 3, six of themutants generated by the enrichment at250C are
com-pared. All mutants of this group had a lower
eclipseratethan did thewildtype atboth tem-peratures, andagain, both the higher
E.
(C32-11 and -13) and lower A (C32-12, -14, and -18)
arerepresented. Thus,thesecondpredictionof
theArrheniustheory is also fulfilled.
Ifone considers the possibility that asingle
mutation might simultaneously alter both Ar-rhenius parameters,twoother classesshould be presentinthecspopulations. Mutants with both alower
E.
andalowerAwouldbeselectedby enrichment at either 37 or 250C, since anAr-rheniusplot with thesameyintercept but with asmallerslopethan the lower A mutant inFig. 2would fall between thewild-type and lowerA lines. Moredetailed kinetic dataatseveral tem-peratures will be required to distinguish this class. However, certain mutants with both a
higher
E.
andahigherAcould be revealed by this data. For example, an Arrheniusplotwithahigheryinterceptandasteeperslopethanthe
wild-type line in Fig. 2 could cross the 370C
ordinateatthesamepointasthewild-typeline
but below the wild-type line atthe 250C ordi-nate. Since this mutant would have the same
[image:6.503.257.456.55.469.2]eclipse rate as does the wild type at
370C,
itTABLE 2. Kinetic characterization of eclipse mutants'
In vivo eclipse (single compo-nent)
k(wild Tentative Mutant k
(min-f)
type)/k(mu- classtant) 37°C 25°C 370C 25°C Wild type 3.4 0.42b
C33-15 3.5 0.20-0.25 1 2 Wild type C33-29 2.8 0.23-0.29 1 2 Wild type C33-25 1.0c 0.030 3 14 HigherEa C33-30 0.60c 0.02 6 20 HigherE. 033-27 0.66c 0.040 5 10 Lower A C33-28 0.69 0.041 5 10 LowerA
a
Trial
A2of Table1.Virus was grownand enrichedat37°C. bBiphasic first-orderplots;the rate constant isfor the cor-rected faster component.'These showed bothabiphasicplotandasinglecomponent plotonrepeat kineticruns.However,the correctedrate con-stant for thefastercomponentwaswithinexperimentalerror of the rate constant obtained from the single-component
curve.
TABLE 3. Kineticcharacterizationofeclipse mutantsa
Invivoeclipse(single
compo-nent)
MutantMuat k (min~l)k
(min-')
k(wild(mutant)type)/k Tentativeclass370C 250C
370C
250CWild type 3.4 0.42
C32-11 0.69 0.03 5 14 HigherEa C32-13 0.46 0.01 7 42 HigherEa C32-14 0.35 0.02 10 21 LowerA
C32-16 0.20 -b 17 -b
C32-12 0.23 0.02 15 21 Lower A C32-18 0.39 0.03 9 14 Lower A
a
Trial
Bof Table 1. Virus was grown at370C
and enrichedat250C.
"Thefirst-orderplotshowedaloss in PFUsatearlytimes
butaregaining of PFUs at later times.
would,therefore, be lost during the enrichment
step at
370C
butnot at250C.
Yet thismutantdoesnot seem toberepresented in Table3,since
none of the six examined have arate at
370C
that is closetothat of thewildtype.
Transformation of
OX174
from inactiveto active form. In the course of this
study,
acertain featureof the
-OX
adsorption properties
wasobservedtobealtered in these
eclipse
mu-tants. To insure that thischangeinadsorption
characteristicsdidnotintroduceanartifact into
themeasurement of the eclipseratesexhibited
by these mutants, a
preliminary
study of thewild-type
and eclipse mutantadsorption
wasmade. Since the measurement of the
eclipse
reaction rate
requires
synchronized
infections,
theadsorption
step had to be carried out aton November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.259.457.270.466.2]15°C. When care was taken to maintain the virus samples in ice from the time they were
removed from the refrigerator until the
begin-ning of the adsorption period, only 10% of the
PFUsattachedtosensitivecells.If,however,the
sampleswereinadvertentlyleftat room
temper-ature for a short period of time, the level of
adsorbed viruswasmuchhigher.This variation
was never observed in adsorption studies at
370C,suggestingthat
OX
canexist in an inactiveformduring storage at low temperatures and is
transformedinto theactive format
higher
tem-peratures. Ifthis is the case, theproblemwould
onlybeencounteredin studiesrequiring
adsorp-tionatlow temperatures.
Inapreviousreport(2), atransformation from
a thermally stable form of
OX,
incapable ofadsorbingtocells at low temperatures,to amore
labile form that could adsorb at 00C was
ob-served when the viruswas incubated inpH 7
phosphatebufferat370C.Todetermine whether
this or a similar transformation could account for the variable levels ofadsorptionencountered
atthebeginningof thisstudy, experimentswere
designedto testforsuchatransformation in the
starvation buffer used for the
eclipse
reaction.Anticipating apossible rapidrate of this
trans-formationat370C,the reactionwasinitiatedby
a100-folddilutionof ice-coldvirusstocks intoa
reaction flask containing starvation buffer. At
various time intervals, samples were removed
and diluted 100-fold into tubes containing sen-sitive cells for a 15-min adsorption period at either 2 or
150C.
Toinsure thatadsorptionwasrapid andcomplete inthis timeperiod, the cell
concentration was above109cellspermlandthe
multiplicitywasbelow0.01.Aftercentrifugation,
theconcentration of unadsorbedPFUswas
de-termined in each supernatant solution. Since
control experiments showed that incubation
alone did not leadto virus inactivation, these
titers represent the amount of virus that has
failed to attach to the cells. Over 95% of the
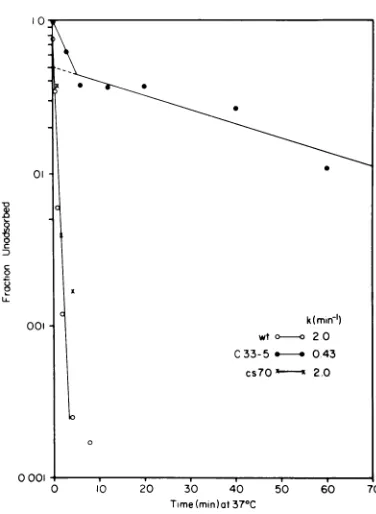
wild-typeandcs7O stockswereconverted within
10 min at 37°C to an active form capable of adsorbing to cellsatlowtemperatures (Fig. 3). However, the oneeclipse mutant from trial Al inTable 1 showed asignificantlylower rate for
this transformation, and a 90-min incubation
wasneeded to convert 90%of thismutant to an adsorbableform.When the 12 csmutants, whose
eclipseratesare shown inTables 2 and 3, were
examinedby thisprocedure, a possible correla-tion emerged. All of the mutants with lower
eclipseratesthan that ofthe wildtype also had
lower rates for thistransformation reaction. The differences in the ratesamong the mutants var-ied from 25- to 200-foldlower than that of the wild type. The two cs mutants whose eclipse
0.01 k(mirrl)
wt o-o 2.0 C 33-5 0.43
cs70W 2.0
0.001
0 10 20 30 40 50 60 70
[image:7.503.258.446.71.328.2]Time(min)at37°C
FIG. 3. Comparison of the in vitrotransformation reactionforwild-type4X174 and its eclipsemutants. Virusstocks maintained continuously at 2°Cwere diluted 100-fold into starvation buffer at 370C. At each timepoint, sample was diluted 100-fold into starvationbuffercontainingE.coliC/1 cellsat2°C. Theconditionsof adsorptionforthisexperimentare 15min at a cell density of3 x 109/ml and at a multiplicityofinfection ofless than0.001. Thezero timepoint wasobtainedbydiluting thevirus stock into starvation bufferat20C, andafter10min, the sample was diluted into buffer containing cells at
2°C.wt, Wild type.
rates are notsignificantlydifferent fromthat of the wild type (C33-150 and -29 inTable2) also
had transformation rates that were similar to
that of the wild type.
This suggests that a slower transformation
rate may be a property of eclipsemutants
se-lectedbytheproceduresdescribed in this report
and could be the reason for their enrichment
during the eclipse step at 37°C. If this is the
case, these same mutants should beselectedby
screeningtheunadsorbed mutagenized
popula-tion of
OX.
Trial C in Table1shows theexpected enrichment ofonlythe csphenotype. Thus, the cs mutants selected in this study survived theeclipse step because they have either a slower
eclipserateafterattachment at150Cora slower
transformation rate to the adsorbableform.
The reversibilityofthis
transformation
reac-tion was tested during the initial experiment
withwild-type
4X.
Asampleofvirus was dilutedat the end of the
370C
incubation period inon November 10, 2019 by guest
http://jvi.asm.org/
39, 1981
starvation buffer at 2°C for a 60-min period
before cellswere added. The level of free virus
in thiscase was the same as that at the 8-min
timne
pointinFig. 3. This means that nosignifi-cant transformation back to the inactive form
occurred in 1 h at low temperatures, and the
data in Fig. 1 represent the concentration of
active virus present at
370C
and during theadsorptionperiod at lowertemperatures.
Although the molecularbasisforthe
intercon-version betweenthese forms is not
understood,
the above resultsweretakenintoaccountinthe
design of the selection proceduresandthe
char-acterization of themutantsthatweregenerated
by them. For example, the transformation
reac-tionwasexaminedwitheach mutant so that an
appropriate preincubationperiod at
370C
couldbe used before the adsorptionstep toinsure that
morethan 90% of the viruswasattachedtocells
at the beginning of the eclipse kinetic
experi-ments. Moreover, the level of free virus was
usuallymeasured atthe end ofthe adsorption
period. As for the selection procedures, the
mu-tagenized
wild-type
stock was alwayspreincu-bated at
370C
for15minbefore the additionofcells. These precautionswerenecessary toinsure
that the majority of the infectious virus were
attachedto
cells
during the enrichmentstep.DISCUSSION
The
preliminary
characterization of theeclipse kinetics for thesemutants
clearly
estab-lishes the
selectivity
of the combinedenrich-mentandscreening procedures.Inall fourtrials,
therewas atleasta10-fold increase in the
fre-quencyofthepredictedcs
phenotype. Moreover,
intwo
trials,
the eclipse kinetics of sixmutantswere examinedat two temperatures, and over
two-thirds of the mutants had the
predicted
lower
eclipse
rate. Inmeasuring
theeclipse
ki-netics,
onebegins
the reaction with a 100-folddilution of virus-cell
complexes
fornedat150C.
This design excludes any effect ofaltered
ad-sorption
kineticssince sufficienttime is allowed foradsorption
tobecompleted
before theeclipse
reaction is
begun.
Thus,
the observed lowereclipse rate in the mutants is not a result of
altered
adsorption
properties.
Additionalexper-imentsare
required
todeterminewhethersomeof the mutations may be
pleiotropic
and mayimpart
alteredadsorption
kinetics as well. Ifareversible
adsorption
stepprecedes
theirrevers-ible
eclipse reaction,
more detailed studies arerequiredtodetermine what
contribution,
ifany,anincrease inthe detachmentratemakestothe
observed decrease in the rate at which these
mutantsenterthe
eclipse phase.
Becauseof thispossibility,
theadsorption
kineticsatlow tem-peraturesmustbecarefully
examinedtoestab-MUTANTS 517
lishwhether the firststepleadingto
OX
infectionisreversible. However, it is clearthat the
pro-cedures describedin this report arehighly
selec-tive for mutants with defects in steps thatare
subsequenttoadsorption.
Theselectivity of the technique and the
pre-liminarycharacterizationofthe mutantssuggest
that the invivoeclipsereactionof4X174may,
indeed, be understood in terms of kinetic theory.
Two ofthe threedetectableclassespredicted by
the theoryhave been selected. As pointed out
earlier,mutants with a higherEaand a higher A
were not represented in the group of six that
were examined in detail (Table 3). However,a
single base change which results in an altered
value for both Arrhenius parameters may not
exist or may occur soinfrequently that a large
number ofmutants may have to be examined
for one to be found. The initial success of the
theory inselecting thesemutantswarrantsmore
detailedkinetic and structuralstudies, with the
hope of revealing the molecular propertiesthat
determine the values ofbothArrhenius
param-eters. Although kineticanalysis of biochemical
reactions has yielded values for the
transition-state equivalents of Ea and A, the
OX
eclipsereaction is the firstsystem todemonstrate that
one canalter thevalues of these parametersby
genetic mutation. With the availability of the
aminoacidsequence ofthecapsidproteinsand
complete nucleotide sequence of the 4XDNA
(26),rapid sequencing methods forlocating base
changes in thesemutants will providea
begin-ningin
identiffying
thedomains in theDNA andcapsid structures that are involved in forming
the activatedintermediate for the reaction.
Asfor theapplicability of these selection
pro-cedurestootherviruses, key features of
adsorp-tion and eclipse kinetics of
OX
are shared bymany
nonenveloped
viruses. Forexample,
whereas the
adsorption
rate exhibitsonly
aslight
dependence on temperature (9), theeclipse kinetics showanalmost 100-fold decrease
between37and
150C
(8, 23). That thismay beageneral featureissuggested
by
thediversity
oftheviruses in whichit hasbeen observed. These
include R17 (4), Ti
(25),
T4(16),
lambda (20),poliovirus (7), rhinovirus (18), and adenovirus
(17,24).Thesameholdstruefor thedissociation
ofcertaincapsid
proteins during
thebeginning
of the eclipse period. In additionto4X174,
thephenomenon occurswiththe phagesM13 (13),
R17(15),T4(29),lambda(27),P22
(12),
and429(6) and the following animal viruses: human
rhinovirus (18) and adenovirus (30).
Thus,
ageneticselection
technique
that is basedonsucha commonpropertyasthetemperature
depend-enceof theeclipseratecould beapplicable
toa widerange ofviruses.on November 10, 2019 by guest
http://jvi.asm.org/
ACKNOWLEDGMENTS
This work wassupported in part by General Research Grant RR05423 fromthe National Institutes of Health and grantPCM77-15982 fromtheNationalScience Foundation.
Technical assistancewasprovided byMarshaL.Brown. Thehelpful discussion andsuggestionsof JamesKane,Robert Reeves,and David Kingsburyaregreatlyappreciated.
LITERATURE CITED
1.Arscott, P. G., and E. B. Goldberg. 1976. Cooperative actionof the T4 tailfibersandbaseplateintriggering conformational change and in determining hostrange.
Virology 69:15-22.
2.Bleichrodt, J. F.,and E.R Van Abkoude. 1967. The transition betweentwoforms ofbacteriophage fX174 differing in heat sensitivityandadsorption characteris-tics. Virology 32:93-102.
3. Casjens, S.,and J.King.1975.Virusassembly.Annu.
Rev.Biochem. 44:555-611.
4. Danziger,R.E.,and W.Paranchych.1970.Stagesin
R17infection. III.Energy requirementsfor theF-pili mediated eclipse of viral infectivity. Virology 40:554-564.
5.Gschwender, H. H., and P. H. Hofschneider. 1969. Lysis inhibition ofOX174-,M12-,andQJ8-infected Esch-erichiacolibacteriaby magnesiumions.Biochimn. Bio-phys. Acta 190:454-459.
6.Hirokava,H.1972.Transfectingdeoxyribonucleicacid of
Bacillusbacteriophage029that is protease sensitive. Proc.Natl. Acad. Sci. U.S.A. 69:1555-1559.
7. Holland, J.J.,and B.H.Hoyer.1962.Earlystages of
enterovirusinfection.ColdSpringHarborSymp.Quant. Biol.27:101-112.
8. Incardona, N. L 1974.Mechanism ofadsorption and eclipse of bacteriophage4X174.III.Comparisonof the activationparameters for the in vitro and in vivoeclipse
reaction withmutantandwild-typevirus. J.Virol. 14: 469-478.
9. Incardona, N. L 1981. The chemicalnature of virus-receptorinteractions,p.157-167. In K.Lonberg-Holm and L.Philpson (ed.),Virus receptors part2,receptors andrecognition, series B, vol. 8. Chapman and Hall, London.
10.Incardona,N. L, R. Blonski, and W. Feeney. 1972. Mechanism of adsorption andeclipseofbacteriophage *X174.I.In vitroconformationalchange under
condi-tions ofeclipse. J. Virol.9:96-101.
11. Incardona,N.L,andL.Selvidge. 1973. Mechanism of
adsorption andeclipseofbacteriophageOX174.II. At-tachmentandeclipsewithisolatedEscherichia colicell walllipopolysaccharide. J. Virol.11:775-782. 12.Israel, V.1977.Eproteins of bacteriophage P22. I.
Iden-tification and ejection from wild-type and defective particles. J. Virol.23:91-97.
13.Jazwinski, S. M., R. Marco, and A. Kornberg.1973.A coatprotein of the bacteriophage M13 virion partici-patesinmembrane-oriented synthesis ofDNA. Proc. Natl.Acad. Sci. U.S.A. 70:205-209.
14.Jazwinski, S. M., R. Marco, andA. Kornberg.1975.
The gene Hspikeprotein ofbacteriophages4sX174and S13.II. Relation tosynthesisof theparental replicative form. Virology 66:294-305.
15. Krahn, P.M., R. J.O'Callaghan, and W.Paranchych. 1972.Stages of phage R17 infection. VI. Injection of a protein and RNA into the host cell.Virology 47:628-637.
16.Labedan, B., and E. B.Goldberg. 1979.Requirement for membranepotentialininjectionofphage T4 DNA. Proc.Natl. Acad. Sci.U.S.A. 76:4669-4673.
17.Lawrence,W.C.,andH. S.Ginsberg. 1967. Intracel-lularuncoating of type 5 adenovirus deoxyribonucleic acid. J. Virol.1:851-867.
18.Lonberg-Holm, K., and B. D. Dorant. 1972. Early in-teraction of rhinoviruses with hostcells. J. Virol. 9:29-40.
19.Lonberg-Holm, K., and N. M. Whiteley. 1976. Physical and metabolic requirements for early interactions of poliovirus and human rhinovirus with HeLa cells. J. Virol.19:857-870.
20.Mackay, D. J., and V.C.Bode. 1976. Events in lambda injection between phage adsorption and DNA entry. Virology 52:154-166.
21. Murialdo, H.,and A. Becker. 1978. Headmorphogenesis incomplex double-stranded deoxyribonucleic acid bac-teriophage.Microbiol. Rev. 42:529-576.
22. Newbold, J. E., and R. L.Sinsheimer.1970.Process of infection with bacteriophage OX174. XXXII. Early steps inthe infection process: attachment,eclipse,and DNApenetration. J. Mol. Biol. 49:49-66.
23. Newbold, J. E., and R.L.Sinsheimer. 1970. Process of infection with bacteriophageOX174.XXXIV.Kinetics of attachmentandeclipsestepsofthe infection J.Virol. 5:427-431.
24. Philipson,L. 1967. Attachment and eclipse ofadenovirus. J. Virol. 1:868-887.
25. Puck, T.T., A.Garen,and J. Cline. 1951. The mecha-nism of virus attachment tohostcells.I. The role of ions in theprimary reaction. J. Exp. Med. 93:65-88. 26. Sanger, F., A. R.Coulson, T. Friedmann, G. M. Air,
B. G. Barrell, N. L. Brown, J. C. Fiddes, C. A.
Hutchisonm,P. M.Slocombe,andM. Smith.1978.
Thenucleotide sequence of bacteriophage fX174. J. Mol. Biol. 125:225-246.
27. Scandella, D., and W. Arber. 1976. Phage A DNA injection into Escherichia coli pel- mutants is restored by mutations in phage genes V or H. Virology 69:206-215.
28. Segal, D. J., and C. E. Dowell. 1974. Cold-sensitive mutantsofbacteriophage
4OX174.
II.Comparison oftwo cold-sensitive mutants. J. Virol. 14:1115-1125. 29. Silverstein,J. L., and E. B.Goldberg. 1976. T4 DNAinjection. II. Protection ofentering DNA from host exonuclease V. Virology72:212-223.
30. Sussenbach, J. S. 1967. Early events in the infection process of adenovirus type 5 inHeLacells.Virology 33: 567-574.
31. Tessman,L. 1968.Mutagenictreatment of double- and single-stranded DNA phages T4 and S13 with
hydrox-ylamine.Virology35:330-332.