The Human Cytomegalovirus
UL116
Gene Encodes an Envelope
Glycoprotein Forming a Complex with gH Independently from gL
Stefano Caló,aMirko Cortese,a*Claudio Ciferri,b*Luca Bruno,a*Rachel Gerrein,bBarbara Benucci,aGiuseppina Monda,a* Michela Gentile,aTobias Kessler,a*Yasushi Uematsu,aDomenico Maione,aAnders E. Lilja,b*Andrea Carfí,bMarcello Merolaa,c
GSK Vaccines, Siena, Italya
; GSK Vaccines, Cambridge, Massachusetts, USAb
; Department of Biology, University of Naples Federico II, Naples, Italyc
ABSTRACT
Human cytomegalovirus (HCMV) is a major cause of morbidity and mortality in transplant patients and is the leading viral
cause of birth defects after congenital infection. HCMV infection relies on the recognition of cell-specific receptors by one of the
viral envelope glycoprotein complexes. Either the gH/gL/gO or the gH/gL/UL128/UL130/UL131A (Pentamer) complex has been
found to fulfill this role, accounting for HCMV entry into almost all cell types. We have studied the
UL116
gene product, a
puta-tive open reading frame identified by
in silico
analysis and predicted to code for a secreted protein. Virus infection experiments
in mammalian cells demonstrated that UL116 is expressed late in the HCMV replication cycle and is a heavily glycosylated
pro-tein that first localizes to the cellular site of virus assembly and then inserts into the virion envelope. Transient-transfection
studies revealed that UL116 is efficiently transported to the plasma membrane when coexpressed with gH and that gL competes
with UL116 for gH binding. Further evidence for gH/UL116 complex formation was obtained by coimmunoprecipitation
experi-ments on both transfected and infected cells and biochemical characterization of the purified complex. In summary, our results
show that the product of the
UL116
gene is an HCMV envelope glycoprotein that forms a novel gH-based complex alternative to
gH/gL. Remarkably, the gH/UL116 complex is the first herpesvirus gH-based gL-less complex.
IMPORTANCE
HCMV infection can cause severe disease in immunocompromised adults and infants infected
in utero
. The dissection of the
HCMV entry machinery is important to understand the mechanism of viral infection and to identify new vaccine antigens. The
gH/gL/gO and gH/gL/UL128/UL130/UL131 (Pentamer) complexes play a key role in HCMV cell entry and tropism. Both
com-plexes are formed by an invariant gH/gL scaffold on which the other subunits assemble. Here, we show that the
UL116
gene
product is expressed in infected cells and forms a heterodimer with gH. The gH/UL116 complex is carried on the infectious
viri-ons, although in smaller amounts than gH/gL complexes. No gH/UL116/gL ternary complex formed in transfected cells,
suggest-ing that the gH/UL116 complex is independent from gL. This new gH-based gL-free complex represents a potential target for a
protective HCMV vaccine and opens new perspectives on the comprehension of the HCMV cell entry mechanism and tropism.
H
uman cytomegalovirus (HCMV) is a ubiquitous
betaherpes-virus infecting 40 to 60% of the human population (
1
).
HCMV infection usually is mild or asymptomatic in
immuno-competent individuals, while it can cause severe disease in
immu-nocompromised adults. Infants infected
in utero
can suffer from
disseminated HCMV disease and impaired neurological
develop-ment (
2
). Transplant patients do not adequately control HCMV
infection or reactivation, often resulting in graft rejection and
death (
3
). Preventing primary HCMV infection could
signifi-cantly decrease the frequency of organ rejection in seronegative
recipients of solid-organ transplants as well as prevent congenital
infection (
4
). The Institute of Medicine has identified the
devel-opment of an anti-HCMV vaccine protecting against congenital
infection as a top priority (
5
).
The HCMV genome from clinical isolates encodes a minimum
of 165 functional open reading frames (ORFs) (
6–9
). Out of the
total number of proteins composing the HCMV virion,
approxi-mately 20 have been identified as envelope-associated proteins
(
10–12
). Notably, all glycoproteins involved in HCMV cell entry
were shown to be essential for virus replication
in vitro
in different
cell types: glycoproteins B (gB), M (gM), N (gN), L (gL), H (gH),
UL128, UL130, and UL131A (
13
). Although not originally
in-cluded in this list, studies conducted with viruses deleted of the
UL74
gene, whose product is known as gO, have shown that this
glycoprotein is a crucial component of the fusion-promoting
ma-chinery (
14–16
).
HCMV infection begins with the low-affinity tethering of the
virion to cell surface heparin sulfate proteoglycans (HSPGs), an
event mediated by both gB and the gM/gN complex (
13
,
17
). gB
and gH/gL are part of the conserved herpesvirus fusion machinery
and are required for cell fusion and viral entry (
18
). Current
evi-Received2 October 2015Accepted22 February 2016
Accepted manuscript posted online2 March 2016
CitationCaló S, Cortese M, Ciferri C, Bruno L, Gerrein R, Benucci B, Monda G, Gentile M, Kessler T, Uematsu Y, Maione D, Lilja AE, Carfí A, Merola M. 2016. The human cytomegalovirusUL116gene encodes an envelope glycoprotein forming a complex with gH independently from gL. J Virol 90:4926 –4938.
doi:10.1128/JVI.02517-15.
Editor:K. Frueh, Oregon Health Sciences University
Address correspondence to Marcello Merola, marcello.x.merola@gsk.com.
*Present address: Mirko Cortese, Department of Infectious Diseases, Molecular Virology, Heidelberg University, Heidelberg, Germany; Claudio Ciferri, Genentech Inc., South San Francisco, California, USA; Luca Bruno, Advent srl, Pomezia (Rome), Italy; Tobias Kessler, Sandoz GmbH, Sandoz Development Center, Kundl, Austria; Giuseppina Monda, Institute of Cellular Pharmacology Ltd., Mosta, Malta; Anders E. Lilja, Hookipa Biotech AG, Vienna, Austria.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
on November 7, 2019 by guest
http://jvi.asm.org/
dence suggests that the binding of a complex between gH, gL, and
other viral envelope glycoproteins to a cell surface receptor
trig-gers conformational changes in gB, the viral fusogen, resulting in
membrane fusion (
13
). Compositions of the gH/gL-based
com-plexes and cell-specific receptors determine viral tropism and
mechanism of entry (
19–22
). The gH/gL/gO complex appears to
be sufficient for HCMV entry and replication into fibroblasts (
23
,
24
) but still is required for proper growth and for fusion events in
all cell types (
15
,
16
,
25
). The more recently discovered gH/gL/
UL128/UL130/UL131A complex (Pentamer) instead is required
for endothelial/epithelial cell, leukocyte, and monocyte tropism
(
26–28
). On the cellular side, several putative HCMV receptors
have been described, but none of them completely fulfills the
char-acteristics of an entry receptor (
17
,
29
).
It has been estimated that in sera of HCMV-seropositive
indi-viduals, approximately 50% of antibodies neutralizing fibroblast
infection are directed against gB (
30
,
31
). gB has been tested,
to-gether with the adjuvant MF59, in human trials as a potential
HCMV vaccine (
30
,
32
,
33
). The vaccine conferred partial
protec-tion in healthy seronegative subjects (
34
). The observation that
MF59-adjuvanted gB was able to partially protect from primary
infection suggests that a subunit HCMV vaccine is feasible, but it
also has renewed interest for new targets among the exposed viral
envelope glycoproteins. Recently, the analysis of human plasma
from seropositive subjects has identified the pentamer complex as
the main target of antibodies neutralizing infection in epithelial/
endothelial cells (
31
). Moreover, immunization with
Pentamer-containing vaccines elicited strong and broadly neutralizing
re-sponses in small animals and nonhuman primates (NHPs) (
35–
37
). Therefore, the pentamer complex represents a promising
candidate for a potent HCMV vaccine (
35–41
).
HCMV structural glycoproteins can be divided into those that
are conserved among members of the
Herpesviridae
(including gB,
gH, gL, gM, and gN) and subgenus-specific glycoproteins without
homology to other herpesviruses. The latter include, among
oth-ers, gpRL13 (
11
,
42
), gpTRL10 (
43
), gpUL132 (
44
),
UL74-en-coded gO (
45
), UL4-encoded gp48 (
46
), US27 (
47
), and UL33
(
48
). In contrast to the conserved glycoproteins, knockout of
many of the HCMV-specific structural proteins does not prevent
in vitro
replication in fibroblast, epithelial, or endothelial cells and
presumably participates in cell and tissue tropism or
pathogenic-ity (
23
,
24
). The functional characterization of these proteins may
be beneficial for the dissection of the mechanism of HCMV entry
into the broad range of cell types susceptible to HCMV infection.
Here, we report the characterization of the
betaherpesvirus-specific product of the
UL116
gene from the HCMV TR strain as
an envelope glycoprotein forming a stable complex with gH. In
infected cells, UL116 showed late kinetics of expression,
under-went extensive glycosylation, and localized at the site of virus
as-sembly and secondary envelopment. Electron microscopy (EM)
and phase separation experiments indicated that UL116 is part of
the virion envelope. Coimmunoprecipitation experiments and
multiangle light scattering (MALS) analysis of purified proteins
confirmed the formation of a complex between UL116 and gH.
Finally, cotransfection experiments and negative-staining EM
coupled with single-particle analysis of purified complexes suggest
that UL116 and gL form alternative, mutually exclusive complexes
with gH. Overall, we identify a novel HCMV gH-based complex
potentially playing a role in viral infection and dissemination.
MATERIALS AND METHODS
Cell lines, plasmids, and viruses. A bacterial artificial chromosome (BAC) containing the genome of the HCMV TR strain was obtained from Oregon Health Science University. TR, a clinical HCMV strain derived from an ocular vitreous fluid sample from a patient with HIV disease (49), was cloned into a BAC after limited passage in fibroblasts (50). HCMV strain TR and recombinant UL116-Flag-TR virus were propagated in hu-man foreskin fibroblasts (HFF-1; ATCC SCRC-1041) grown in minimal essential medium (Invitrogen) supplemented with 10% fetal calf serum, glutamine (100 mg/liter), and gentamicin (350 mg/liter). Virions were isolated by glycerol-tartrate gradient centrifugation as previously de-scribed (51). HEK293T cells were cultured in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum, glutamine, and genta-micin. Lipofectamine 2000 (Invitrogen) was used to transfect HEK293T cells. Human codon-optimizedUL116, gH (UL75), gL (UL115), and gB (UL55) HCMV genes based on the TR strain sequence were synthesized by GeneArt and cloned into the pcDNA3.1(⫺)/myc-His C vector (Invitro-gen) in frame with C-terminal myc and the six-histidine tag sequences. Single-point mutations were introduced with the QuikChange mutagen-esis kit (Stratagene), resulting in the mono-tag versions and the tag-less versions of these genes.
Generation of UL116 polyclonal antiserum.Mouse antiserum recog-nizing UL116 was developed in-house. Briefly, His-tagged peptides en-compassing the amino acid regions of residues 32 to 265 of UL116 from the TR strain were produced inEscherichia coliand purified through metal ion affinity chromatography. Purified peptides were used to immunize mice.
Construction and generation of UL116-Flag TR BAC.Insertion of the Flag tag at the C terminus of the UL116 ORF in the TR BAC was achieved using the two-step red-mediated recombination method (52). The primer pair used to amplify the kanamycin insertion cassette was 5=-TTC GGC GCC AAC TGG CTC CTT ACC GTC ACA CTC TCA TCG TGC CGC AGA CTG ATT ACA AGG ATG ACG ACG ATA AG-3=and 5=-TAT CAC CGG TCC AGG TGA GAA AGA GAA GCC GCA ATC CGG GCG GCG GCA CAT CA CTT ATC GTC GTC ATC CTT GTA ATC AGT CTG CGG CAC GAT GAG CAA CCA AAT TAA ACCA ATT CTG ATT TAG-3=, where the underlined base pairs encode the Flag peptide.
Reconstitution of infectious virus.To reconstitute the virus, 2g of the BAC-HCMV DNA and 1g of a pp71 expression plasmid were trans-fected into MRC-5 cells by electroporation. Culture medium was changed 24 h later, and the cells were split and cultured until the appearance of cytopathic effect. The virus stock was prepared by harvesting cell-free culture supernatant when extensive cytopathic effect was visible.
Purification of HCMV TR virions.HCMV particles in cell superna-tants were separated into virion, dense-body, and noninfectious envel-oped particle (NIEP) fractions by positive density/negative viscosity gra-dient centrifugation as described previously (53). Particle concentrations in the preparations were estimated by counting negatively stained samples by EM in relation to a standard concentration of latex beads. To separate virion envelope proteins from capsid and tegument proteins, 108particles
were mixed 1:1 with envelope stripping buffer (2% Nonidet-P40 in phos-phate-buffered saline [PBS]) and incubated for 15 min at 4°C. Particles were pelleted (12,000⫻gfor 5 min at 4°C), and the soluble envelope fraction was harvested. The insoluble capsid/tegument material was washed twice with envelope-stripping buffer and once in PBS before being solubilized in SDS-PAGE sample buffer.
Flow cytometry.For the detection of membrane-exposed UL116, HEK293T cells transiently transfected with vectors coding for UL116, gH, gB, and empty vector were trypsin detached 48 h posttransfection, incu-bated for 30 min at room temperature (RT) with Live/Dead Aqua (Invit-rogen), diluted 1:400 in PBS, and incubated with different dilutions of anti-UL116 polyclonal mouse sera for 60 min on ice. After three washes in PBS, the Alexa Fluor 647-conjugated goat anti-mouse secondary antibody was added at a 1:200 dilution and incubated for 30 min on ice. A total of 104cells were analyzed for each histogram using FACSCanto II (Becton
HCMV gH and UL116 Form a Complex Alternative to gH/gL
on November 7, 2019 by guest
http://jvi.asm.org/
Dickinson, Heidelberg, Germany). Experiments were performed in trip-licate for statistical consistency; means and standard deviations were an-alyzed and plotted using GraphPad Prism software.
Immunogold EM.Purified virions were air dried to the surface of Formvar-coated EM grids. The grids were treated with mouse anti-Flag antibody (F3165; Sigma) for 4 h at RT, washed 3 times with PBS, and incubated with goat anti-mouse antibody conjugated to 5-nm gold parti-cles for 1 h at RT. After further washing in PBS, the grids were negatively stained with phosphotungstic acid and subjected to EM analysis.
Purification of gH/UL116 and gH/gL-C144A Fab complexes.The gH/UL116 complex was purified from supernatants of HEK293 cells bly transfected with plasmids encoding UL116 and gH containing a dou-ble Strep-tag and 6His tag at their C termini, respectively (UL116-strep and gH-His). The medium then was applied to StrepTrap HP 1-ml col-umns (GE Life Sciences), and the complex was eluted with the same buffer containing 2.5 mM desthiobiotin. The gH/gL complex harboring the gL-C144A mutation was purified from supernatants of HEK293 cells doubly transfected with plasmids encoding gH-His and untagged gL-C144A as previously described (54). Purified gH/gL-C144A or gH/UL116 was mixed at 1:1.2 with either 3G16 or MSL-109 Fabs. The ternary complexes were isolated by size exclusion chromatography (SEC).
MALS analysis.Purified gH/UL116 complex, at two different dilu-tions, was analyzed by SEC on a Superose-6 10/30 column (GE Health-care) using 25 mM Tris, pH 8.0, 150 mM NaCl as a running buffer. The SEC system was coupled in-line with the following calibrated detection systems: (i) HP1 1050 Hewlett-Packard UV detector (Norwalk, CT); (ii) MiniDawn Treos multiangle light scattering (MALS) detector (Wyatt Corporation, CA); (iii) quasielastic light scattering detector (Wyatt Cor-poration, CA); and (iv) Optilab T-reX refractive index (RI) detector (Wy-att Corporation). Briefly, these combined measurements allow both the absolute molar mass of an eluting glycoprotein and the individual contri-butions made by protein and carbohydrate components to be determined (55–57). The carbohydrate contribution to the molar mass can be assessed because glycans contribute directly to the RI signal but not the UV signal.
Competition between gL and UL116 for gH binding and formation of higher-order complexes.HEK293T cells were transfected with DNA encoding gH-His and either gL, UL116-strep, or both. The samples in-volving UL116 were purified by Strep-tag affinity, while the gH/gL com-plex was purified by Ni affinity as described above. The elution fractions were run on SDS-PAGE with previously purified gH/gL and gH/UL116 as controls. To detect higher-order complex formation, HEK293T cells were transfected with DNA encoding soluble gH extracellular domain in four different combinations: (i) gH/gL/UL128/strep-UL130/UL131, (ii) gH/ strep-UL116/UL128/UL130/UL131, (iii) gH/gL/His-gO, and (iv) gH/ strep-UL116/gO. The formation of pentamer in supernatant was assessed by Western blotting with anti-UL128 antibody, whereas anti-His was used to reveal secreted gO. The samples involving UL116 were purified by Strep-tag affinity, and associated proteins were revealed by Coomassie staining of the gel.
Negative-staining electron microscopy and single-particle analysis.
Five microliters of purified gH/UL116/3G16, gH/UL116/MSL-109, or gH/gL-C144A/3G16 samples (approximately 10 ng) were placed onto a freshly glow-discharged holey carbon grid covered with a thin layer of continuous carbon. The grid was stained with sequential 75-l drops of a freshly prepared 2% (wt/vol) uranyl formate solution. Samples were im-aged using a Tecnai Spirit T12 transmission electron microscope operat-ing at 120 keV at a nominal magnification of⫻49,000 (1.57 Å/pixel at the detector level) using a defocus range of⫺0.8 to⫺1.2m. Images were recorded under low-dose conditions on a Gatan 4,096- by 4,096-pixel charge-coupled-device (CCD) camera. Particles were manually picked using e2boxer (EMAN2) (58) and extracted using a 224-pixel box size. The two data sets were band-pass filtered with a 200-Å high-pass cutoff and a 20-Å low-pass cutoff. For two-dimensional (2D) classification, we used iterative multivariate statistical analysis (MSA) and multireference alignment (MRA) in Imagic (59). 2D classes from gH/gL and gH/UL116
Fab complexes were aligned and compared by cross-correlation using the SPIDER AP SH command.
UL116 expression kinetics and glycosidase digestion.Infected cells were harvested at the indicated time points and lysed in radioimmuno-precipitation assay (RIPA) buffer (Sigma) according to the manufactur-er’s protocol. Late-phase protein expression was inhibited by the use of phosphonoacetic acid (PAA; Sigma-Aldrich). A total of 250g/ml of PAA was added to the medium at the time of infection and maintained throughout infection. For deglycosylation treatments, 20g of protein extract was incubated with 2.5l of endoglycosidase H (Endo H; NEB), 2.5l peptide-N-glycosidase F (PNGase F; NEB), or buffer only for 3 h at 37°C according to the manufacturer’s protocol. Samples were analyzed by Western immunoblotting.
SDS-PAGE and immunoblotting.Proteins were separated by SDS-PAGE on 10 to 15% polyacrylamide gels under standard conditions. Pro-teins were transferred to nitrocellulose membranes, and membranes were blocked with PBS containing 0.1% Tween 20 and 5% powdered milk. Incubations with primary antibodies and sera, diluted in PBS– 0.1% Tween 20 (PBST), were performed at RT for 1 h, followed by 3 washes in PBST before incubation with horseradish peroxidase-conjugated second-ary antibodies (PerkinElmer) for 1 h. Membranes were washed 5 times in PBST prior to detection with an enhanced chemiluminescence detection system (Pierce).
Immunofluorescence.Cells were plated on glass coverslips and in-fected with HCMV. At day 7 postinfection, cells were fixed in 4% para-formaldehyde, permeabilized with 0.5% Triton X-100, preblocked with HCMV seronegative human IgG, and incubated with primary antibody for 1 h at RT. Following washing in PBS, secondary antibodies were incu-bated for 1 h at 37°C, washed again, and mounted with 4= ,6-diamidino-2-phenylindole (DAPI) ProLong safe stain mounting media (Invitrogen). Primary antibodies were rabbit anti-Flag (Sigma), mouse anti-Flag (Sigma), sheep human TGN 46 (Serotec), mouse monoclonal anti-body (MAb) anti-pp28 (Abcam), mouse anti-gH (AbD Serotech), and rabbit anti-gL (OHSU). Secondary antibodies were Alexa Fluor 488-, 568-, and 647-conjugated goat anti-mouse and anti-rabbit (Invitrogen). The intracellular locations of antibody-tagged proteins were examined under laser illumination in a Zeiss LMS 700 confocal microscope, and images were captured using ZEN 2009 software. Image analysis was per-formed with Fiji (NIH) and Imaris 8.1 (Bitplane). Deconvolution was performed using Autoquant X3 (MediaCybernetics) with fixed point spread function (PSF) modeling.
Immunoprecipitations.HFF-1 cells were infected with HCMV TR and UL116-Flag-TR. Protein expression was allowed to proceed for 72 h, and then cells were washed in 1⫻PBS and lysed with a lysis buffer (25 mM Tris, 150 mM NaCl, 1 mM EDTA, 1% NP-40, 5% glycerol, pH 7.4) in the presence of protease inhibitors (Complete EDTA-free; Roche). Five hun-dred micrograms of total protein was incubated overnight at 4°C with 5
g of the anti-gH MSL-109 monoclonal antibody. Complexes were im-munoprecipitated using Protein G Dynabeads (Invitrogen) according to the manufacturer’s protocol. The beads were washed four times in lysis buffer and then boiled for 5 min in SDS-PAGE loading buffer. Proteins were separated on SDS-PAGE, and immunoblotting was performed as described above. A similar procedure was adopted for immunoprecipita-tion from supernatants of transiently transfected HEK293T cells. Com-plexes were captured in parallel experiments with covalently linked anti-His antibody magnetic beads (GenScript) and anti-c-myc magnetic beads (Pierce) to avoid background signals in the eluted materials.
RESULTS
Primary structure of UL116 gene product.
In all sequenced
HCMV genomes, the
UL116
gene is located in the unique-long
(UL) region between the
UL115
(gL) and
UL117
genes on the
antisense coding strand (
Fig. 1A
). The
UL116
mRNA was
previ-ously shown to arise in the true-late stage of AD169 infection as
part of the
UL119-UL115
transcription unit, but the gene product
on November 7, 2019 by guest
http://jvi.asm.org/
was never analyzed (
60
). The
UL116
gene is not conserved among
alpha and gamma herpesviruses; thus, it does not belong to the
core herpesvirus genes (
61
). Orthologous genes are present in
cy-tomegaloviruses infecting nonhuman primates (
62
), mouse, rat,
and guinea pig (
63
,
64
). The
UL116
ORF from the HCMV TR
strain is predicted to encode a 313-amino-acid glycoprotein
com-prising a signal peptide (amino acid positions 1 to 24) (
Fig. 1B
, SP)
and a threonine-rich domain (amino acids 27 to 157), with the
resulting polypeptide backbone predicted to have a molecular
mass of 34.2 kDa. Fourteen consensus sites for N-linked
glycosy-FIG 1UL116position in the HCMV TR genome and sequence conservation among laboratory-adapted and clinical HCMV strains. (A) ORF map of the TR BAC clone used in this work. Arrows indicate the relative orientations of the repeated and unique ORF blocks. TheUL116gene, in boldface, is located betweenUL115(gL) andUL117genes on the antisense strand. (B) T-Coffee primary amino acid sequence multialignment showing 98%UL116gene conservation among members of a consistent pool of HCMV strains. Asterisks indicate the predicted 14 N-glycosylation sites. SP indicates the N-terminal predicted signal peptide. RL, repeat long; UL, unique long; IRS, internal repeat short; US, unique short; TRS, terminal repeat short; B, BAC inserts.
HCMV gH and UL116 Form a Complex Alternative to gH/gL
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.47.534.89.585.2]lation are predicted, whereas the O-glycosylation acceptor
poten-tial sites are very numerous (
⬎
70). Moreover, UL116 lacks
mem-brane anchor sequences and therefore is expected to be a secreted
protein. A multisequence alignment of UL116 from clinical and
laboratory-adapted HCMV strains reveals 98% sequence identity
(
Fig. 1B
), suggesting a conserved functional role.
Kinetics of UL116 expression during HCMV replication.
To
investigate the expression kinetics of UL116 during productive
HCMV infection, we generated a recombinant HCMV, derived
from the BAC clone of the TR strain, with a Flag tag fused to the
UL116 C-terminal end. The reconstituted TR UL116-Flag virus,
which showed growth kinetics identical to those of the parental
virus (
Fig. 2A
), was used to infect HFF-1 cells. Extracts were
pre-pared from infected cells at different time points, ranging from 4 to
120 h postinfection (hpi), and the expression kinetics of UL116 were
monitored by immunoblotting using an anti-Flag antibody. UL116
migrated as two species, a faster-migrating protein of 76 kDa and a
slower-migrating protein of about 125 kDa (
Fig. 2B
). The latter
ap-peared to be the mature product of the 76-kDa species, as indicated by
the increased accumulation of the 125-kDa species from 72 through
120 hpi compared to that of the 76-kDa band (
Fig. 2B
). As a control,
expression of the immediate-early protein IE1 pp72 (UL123) and the
late phosphoprotein pp28 (UL99) were monitored in the same
sam-ples (
Fig. 2B
). The detection of UL116 expression paralleled that of
pp28, a late-phase marker (
65
). Consistent with the observed kinetic
pattern, the inhibition of viral DNA synthesis, a prerequisite for late
gene expression, with PAA resulted in the disappearance of the
UL116 bands (
Fig. 2C
).
The high number of putative N- and O-linked glycosylation
sites predicted on UL116 and the difference between the apparent
FIG 2UL116expression kinetics and carbohydrate addition in HCMV-infected fibroblasts. (A) Multiple-step growth curve analysis of the reconstituted virus UL116-Flag TR and of the parental HCMV strain TR. Time point 0 titers represent input inocula, and each data point represents averages from three independent wells. (B) Uninfected (mock) and TR-UL116-Flag-infected HFF-1 cells at an multiplicity of infection of 5 were harvested at the indicated times postinfection. Equivalent amounts of cell lysates were subjected to SDS-PAGE under reducing conditions and analyzed by immunoblotting with anti-Flag, HCMV IE1, and pp28 protein antibodies as indicated on the left. Actin detection was used as a protein loading control. (C) TR-UL116-Flag infections of HFF cells were performed in the presence (right) or absence (left) of phosphonoacetic acid (PAA), an inhibitor of HCMV late-phase protein expression. Five days postinfection, lysates were prepared from infected cells, and UL116 and pp28 expression was detected by Western blotting using anti-Flag and anti-pp28 antibodies, respectively. (D) Equal amounts of cell lysates of TR UL116-Flag-infected HFF-1 cells (72 h p.i.) were subjected to glycosidase treatments: left lane, untreated control; middle and right lanes, endoglycosidase H (Endo H) and PNGase F treatments, respectively. Proteins were separated on SDS-PAGE under reducing conditions, and UL116 was detected by immunoblot analysis using an anti-Flag antibody.on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.89.507.64.440.2]molecular mass observed (125 kDa) by immunoblotting and the
calculated molecular mass (34.2 kDa) of the polypeptide
back-bone suggested that UL116 undergoes an extensive
posttransla-tional glycosylation process. The trafficking of glycoproteins to
the Golgi apparatus is associated with the processing of N-linked
oligosaccharides to complex oligosaccharides that are resistant to
the action of Endo H (
66
). Endo H completely cleaves only
endo-plasmic reticulum (ER)-type N-linked carbohydrates, while it
does not trim hybrid and complex high-mannose N-linked
car-bohydrates that form when the glycoproteins reach the Golgi
ap-paratus. PNGase F is able to digest both ER and Golgi-type
high-mannose carbohydrate moieties of glycoproteins (
67
). To analyze
the glycosylation of UL116 during the virus life cycle, HFF-1 cells
were infected with TR UL116-Flag for 3 days, followed by
glyco-sidase digestion of the cell extract. As shown in
Fig. 2D
, digestion
with Endo H had several effects: (i) the 125-kDa band was only
slightly affected, and its apparent molecular mass was reduced by
about 10 kDa; (ii) the 75-kDa band collapsed to
⬃
38 kDa, a value
very close to the predicted mass based on sequence (
Fig. 2C
,
com-pare lanes 1 and 2). PNGase F digestion caused the disappearance
of the faster-migrating 76-kDa band that migrated to
approxi-mately 35 kDa, while the apparent mass of the 125-kDa band was
reduced at
⬃
78 kDa (
Fig. 2D
, compare lanes 1 and 3). These
results are consistent with the presence of an immature species
entirely modified with ER-type glycan and of a mature form
car-rying mostly Golgi-type carbohydrates.
Confocal microscopy analysis of HCMV-infected human
fi-broblasts reveals colocalization of UL116 with the viral
assem-bly complex.
Confocal microscopy was used to investigate the
localization of UL116 in HCMV-infected cells. An Flag
anti-body was employed together with antibodies specific for cellular
compartments to trace the subcellular distribution of UL116.
Hu-man fibroblasts were infected with TR UL116-Flag and at 72 hpi
were fixed and stained for confocal analysis. Among the cellular
FIG 3Intracellular localization ofUL116in infected HFF-1 cells. (A) HFF-1 cells were infected with the recombinant virus TR-UL116-Flag at an MOI of 5 for 72 h. Cells then were fixed, permeabilized, and stained. UL116 detection was achieved by anti-Flag (in red) in combination with either the cellular marker TGN 46 (upper) or the HCMV tegument protein pp28 (lower). Cell nuclei are stained blue. The merge panels show colocalization of the red and green signals. (B) Same experiment as that described for panel A, except that mouse anti-gH was used. A 3D reconstruction and the colocalization surface of the UL116 and gH signals are shown in the lower images.HCMV gH and UL116 Form a Complex Alternative to gH/gL
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.133.448.69.465.2]markers used, UL116 showed colocalization with the
trans
-Golgi
marker TGN 46 (
Fig. 3A
, upper). Based on the cytoplasmic
com-partmentalization of UL116, we asked whether the structural
HCMV proteins colocalized with UL116. We found a distinct
co-localization of UL116 with tegument phosphoprotein pp28 and
gH in infected HFF-1 cells 72 h after infection with TR UL116-Flag
(
Fig. 3A
and
B
, respectively, bottom). Image deconvolution and
3D reconstruction allowed us to better define the colocalization
surface between UL116 and the glycoprotein gH. As shown in
Fig.
3B
, both proteins strongly accumulate in a cytoplasmic
perinu-clear inclusion and in smaller vesicles scattered in its close
prox-imity. pp28 and gH previously were reported to localize with other
tegument or envelope proteins at the virus assembly complex
(AC) site during the late phase of the infectious cycle (
68
,
69
) and
to be acquired by the nascent virion.
These data are consistent with the trafficking of UL116 to the
site of viral AC and suggest that UL116 can associate with other
envelope glycoproteins.
UL116 is a component of the HCMV virion envelope.
To
ver-ify if UL116 was incorporated into the virion and to establish its
localization, we purified the recombinant TR UL116-Flag viral
particles from the supernatant of infected HFF-1 cells and
per-formed both Western blotting and immuno-EM analysis. TR
UL116-Flag particles purified by positive density/negative
viscos-ity glycerol/tartrate gradient were extracted with detergent to
sep-arate envelope and tegument fractions (
51
), which were probed by
immunoblotting with different antibodies (
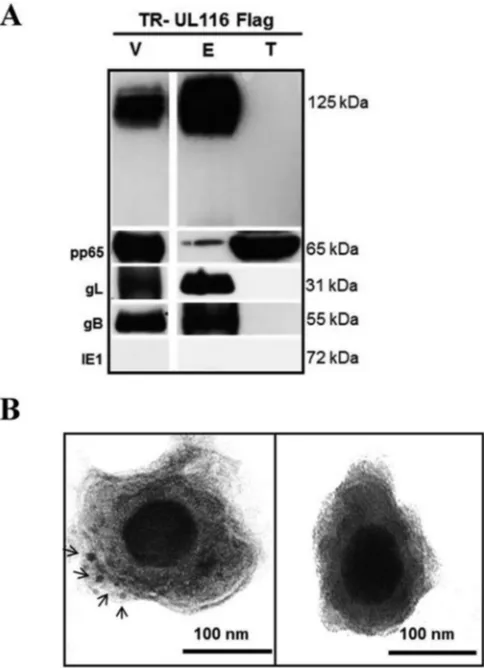
Fig. 4A
). pp65 was
used as a marker of the tegument fraction, and the nonstructural
HCMV protein IE1 was used to exclude samples that were
con-taminated by cellular extracts. As expected, the envelope proteins
gB and gL were detected by specific antibodies in the envelope
fraction (
Fig. 4A
). Notably, probing with the anti-Flag antibody
showed that UL116 was present exclusively in the envelope
tion and was not detectable in the pp65-containing tegument
frac-tion.
To confirm the presence of UL116 on the viral envelope, we
performed immunoelectron microscopy on the purified TR
UL116-Flag particles using a wild-type TR viral preparation as a
control. The 10-nm gold-labeled anti-Flag secondary antibody
displayed distinct labeling of the envelope (
Fig. 4B
). Taken
to-gether, these results are consistent with the localization of UL116
on the surface of the HCMV envelope.
Cotransfection of recombinant UL116 and gH.
To better
characterize the UL116 gene product, we generated a recombinant
myc-His tagged version of the gene that was codon optimized for
efficient expression and cloned in a eukaryotic expression vector.
Surprisingly, transfection in MRC-5 or HEK293T cells did not
result in secretion of the recombinant product or its transport to
the cell surface (
Fig. 5A
). Confocal analysis of MRC-5-transfected
cells showed that UL116 colocalized almost completely with
pro-tein disulfide-isomerase (PDI), a marker of the endoplasmic
retic-ulum (
Fig. 5A
). We hypothesized that correct localization of
UL116 can be achieved following association with one or more of
the major HCMV envelope glycoproteins (
70
). To investigate this
possibility, we performed binary cotransfection experiments in
HEK293T cells of TR-UL116 with well-characterized HCMV
en-velope glycoproteins and analyzed the membrane localization of
UL116 by cytofluorimetric assay on nonpermeabilized cells with a
polyclonal mouse UL116 antisera and a fluorescein isothiocyanate
(FITC)-conjugated secondary anti-mouse antibody. To exclude
possible nonspecific detection, we included
single-gene-trans-fected cells as negative controls. The results from a representative
experiment using gH/UL116 and gB/UL116 are shown in
Fig. 5B
.
A strong UL116 plasma membrane signal was observed only in
gH/UL116-coexpressing cells. The cotransfection of UL116 with
other HCMV proteins, such as gO and UL10, did not allow the
transport of UL116 on the plasma membrane (unpublished
re-sults). These data are consistent with UL116 depending on the
formation of a heterodimeric complex with gH for translocation
to the cell surface.
To test the formation of a gH/UL116 complex, we generated
eukaryotic expression vectors carrying singly tagged gH and
UL116 versions of the codon-optimized genes. We first performed
coimmunoprecipitation experiments on extracts of HEK293T
cells coexpressing gH-myc/UL116-His or gB/UL116-His. Extracts
from single-gene transfection with gH-myc and gB were used as
controls. Coimmunoprecipitations were performed with an
anti-His antibody to detect proteins associated with UL116 and by
anti-myc to reveal species associated with gH. Each
immunopre-cipitated sample was divided into three aliquots and treated for
Western blotting using anti-His (for UL116), anti-myc (for gH),
FIG 4Localization ofUL116on the virion envelope. (A) Western blotting was performed on purified virions from virus expressing Flag-tagged UL116. The virion total lysate (V), envelope fraction (E), and tegument/ capsid fraction (T) were probed for the indicated antigens. (B) Immuno-gold EM staining for anti-Flag MAb of purified TR-UL116Flag and TR wild type (left and right, respectively). Black arrows indicate gold particles. Original magnification,⫻40,000.on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.301.543.68.402.2]and anti-gB as probes. Pulldown of UL116 resulted in association
with gH only and not with gB (
Fig. 5C
, bottom, lanes 5 and 10,
respectively). Conversely, immunoprecipitation of gH resulted in
the coimmunoprecipitation of UL116 (
Fig. 5C
, middle, lane 1).
To confirm the formation of the gH-UL116 complex and to
ex-clude potential artifacts induced by the overexpression system, we
decided to test the interaction between the two proteins in
HCMV-infected cells. To this aim, we performed
coimmunopre-cipitation experiments using the anti-gH human monoclonal
an-tibody MSL-109. Extracts from TR UL116-Flag-infected HFF-1
cells at 96 hpi were prepared and submitted to anti-gH
immuno-precipitation. Cell lysate from wild-type-TR-infected HFF-1 cells
was used as a negative control. The gH-associated proteins were
separated by SDS-PAGE, and as a positive control, gL was probed
by Western blotting. Anti-Flag antibody was used as the probe to
reveal the presence of UL116. As expected, gL was detected in
extracts from cells infected with both viruses (
Fig. 5D
). Notably, a
clear signal for UL116-Flag was detected in extracts from cells
infected with TR UL116-Flag but not when wild-type TR virus was
used for infection (
Fig. 5D
).
Finally, HEK293T cells were transfected with plasmids
encod-ing a soluble form of gH and UL116 containencod-ing a Strep-tag at the
C terminus. HEK293T cell supernatants were loaded on a
Strep-Tactin resin, and the eluted material was analyzed using SEC,
mul-tiangle light scattering, and SDS-PAGE. SEC and SDS-PAGE
re-vealed that gH and UL116 form a stable complex that migrates as
FIG 5UL116interaction with gH in HEK293T transfected cells. (A) ARPE-19 cells transiently expressing UL116-myc were fixed, permeabilized, and stained with anti-myc (green) and anti-PDI (red) specific antibodies prior to confocal immunofluorescence microscopy. Scale bars represent 10m. (B) Detection of UL116 by fluorescence-activated cell sorter (FACS) analysis on nonpermeabilized HEK293T cells. Cells were transfected with expression vectors for UL116 as well as gH and gB, both alone and in combination, as indicated. Forty-eight hours posttransfection, cells were stained at 4°C with anti-UL116 polyclonal mouse serum at different dilutions. Excess probe was removed by washing in PBS, and then cells were fixed and stained with Alexa Fluor 488-conjugated anti-mouse antibody. For each point, 10,000 cells were analyzed and the mean fluorescent intensity of Alexa Fluor 488-positive cells was reported. Secondary antibody only was used as a negative control. Experiments were performed in triplicate. (C) UL116-gH coimmunoprecipitation. Lysates from HEK293T cells transiently expressing UL116-his/gH-myc, UL116-his, gB, and UL116-his/gB were subjected to parallel immunoprecipitation (IP) experiments (antibodies used are specified on the left) with both covalently linked magnetic anti-His and anti-myc beads. Total lysates (input) and eluted samples were separated by SDS-PAGE and analyzed by immunoblotting for both the His and myc tag (indicated at the bottom). (D) Coimmunoprecipitation (Co-IP) of the UL116-gH complex in infected cells. Cell lysates were prepared from HFF-1 infected with HCMV-TR UL116 Flag and wt TR separately (5 dpi). Complexes were captured using the human monoclonal antibody MSL-109. Elutions and crude extracts were subjected to immunoblotting using a rabbit anti-gH polyclonal serum, an anti-Flag M2 clone monoclonal MAb, and a rabbit polyclonal serum specific for gL.HCMV gH and UL116 Form a Complex Alternative to gH/gL
on November 7, 2019 by guest
http://jvi.asm.org/
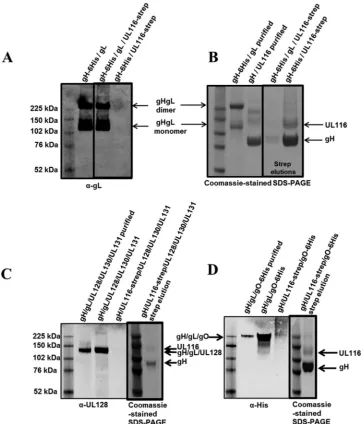
[image:8.585.43.539.66.427.2]a single peak. MALS analysis suggested that the mass of the gH/
UL116 complex is 161 kDa (i.e., glycans account for
⬃
37% of the
total mass of the complex), consistent with a 1-to-1 stoichiometry
(
Fig. 6A
). The heterodimer is not disulfide linked and dissociates
in single subunits in SDS-PAGE under nonreducing conditions
(
Fig. 6B
).
These results demonstrate the formation of a gH/UL116
com-plex in transfected cells that does not require the presence of gL to
form. Moreover, we also were able to confirm the formation of the
complex during a productive HCMV infection.
gL competes with UL116 for gH binding in transfected cells.
To assess if gH, gL, and UL116 can form a ternary complex, we
transfected HEK293T cells with a construct corresponding to the
soluble gH extracellular domain together with either gL or
UL116-strep or both gL and UL116-UL116-strep. Western blot analysis of the cell
supernatants with an anti-gL antibody showed the formation of
monomers and homodimers of the gH/gL heterodimers in cells
transfected with either gH and gL or gH, gL, and UL116 (
Fig. 7A
).
The formation of disulfide-linked monomers and homodimers of
soluble gH/gL is consistent with what has been described
previ-ously (
35
,
54
,
71
). The Strep-tag at the C terminus of UL116
al-lowed us to isolate gH/UL116-containing complexes from cell
su-pernatants. SDS-PAGE analysis of the eluted fraction revealed that
when gH, gL, and UL116 are transfected together, only a very
minor amount of gH/UL116 complex forms and no ternary
com-plex could be detected (
Fig. 7B
). Together, these data suggest that
gL competes with UL116 for gH binding. It has to be noted that gH
and gL form disulfide-linked heterodimers, and perhaps once this
complex forms UL116 is not able to displace gL.
The gL subunit is not only disulfide linked to gH but also is
engaged in disulfide bond formation with gO, in the gH/gL/gO
complex, or with UL128 in the pentamer (
54
). Although the
ab-sence of gL from the gH/UL116 dimer suggested that neither gO
nor UL128 can associate with this heterodimer, we sought to
ver-ify this hypothesis. To assess if UL116 could replace gL as a scaffold
for the formation of these complexes, we transfected HEK293 cells
with constructs encoding the soluble extracellular gH domain
with the following combinations: (i) gH/gL/UL128/strep-UL130/
UL131; (ii) gH/strep-UL116/UL128/UL130/UL131; (iii) gH/gL/
His-gO; and (iv) gH/strep-UL116/His-gO. Secreted proteins then
were detected by Western blotting, and Strep-tag pulldown was
used to reveal subunits associated with UL116. As can be seen in
Fig. 7B
and
C
, UL128 is secreted only when gL is present and not
when UL116 is expressed. Similarly, gO does not form a complex
when UL116 replaces gL (
Fig. 7D
). Thus, the association of UL116
with gH competes with the formation of both pentamer and gO
subunits.
Negative-stain EM analysis of the purified UL116/gH
com-plex.
We previously used single-particle negative-stain EM to
characterize gH/gL, gH/gL/gO, and Pentamer complexes bound
to neutralizing antibodies (
54
,
72
). Our data revealed that HCMV
gH/gL has a boot-shaped structure similar to that of HSV-2 gH/gL
(
73
) and that gO and the UL128/UL130/UL131A subunits bind to
the N-terminal end of gH/gL in gH/gL/gO and Pentamer,
respec-tively. We also engineered a gL mutant, C144A, which prevents
gH/gL homodimerization.
Here, we extended the EM analysis to the purified gH/
UL116 complex bound to either MSL-109 or 3G16 antibody
(
41
,
72
). The trimeric gH/UL116/3G16 and gH/UL116/MSL-109
complexes were analyzed by negative-stain EM and single-particle
analysis. 2D class averages were generated and compared to the
cor-responding 2D class averages for the gH/gL-C144A/3G16 complex
(
Fig. 8
). This analysis revealed that the gH/UL116 complex has a
boot-shaped structure similar to that of gH/gL. 3G16 and
MSL-109 Fabs bind to the C-terminal domain and to the “heel” of
the gH molecule, respectively, as previously described for
gH/gL (
54
,
72
). Furthermore, the EM analysis and binding data
(unpublished results) suggest that UL116 occupies a position
similar to that of gL at the N-terminal end of gH (
Fig. 8
) and
that the presence of UL116 does not affect the 3G16 and
MSL-109 binding sites.
DISCUSSION
HCMV envelope glycoproteins are potential components of a
prophylactic HCMV vaccine. Among them, gB is the first
recom-binant antigen used for clinical immunization studies (
74
). In
FIG 6MALS analysis of purified gH/ULL116 complex. (A) SEC profile of the gH/UL116 complex at two different concentrations (gray and orange traces) and molecular mass estimates for the polypeptide only (110 kDa) and for the glycosylated complex (160 kDa) determined by MALS (pink and cyan lines, respec-tively). The leftyaxis pertains to UV absorbance (Abs), while the rightyaxis represents the molecular weight. (B) SDS-PAGE analysis under nonreducing conditions of the gH/UL116 complex used in panel A.on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.97.497.67.235.2]more recent years the focus for preclinical vaccination studies has
shifted to the components of the gH-based complexes gH, gL, gO,
UL128, UL130, and UL131A. Here, we describe the initial
charac-terization of the product of the HCMV
UL116
gene from the TR
strain that we have identified as a virion envelope glycoprotein.
Our results suggest that UL116 forms a noncovalent heterodimer
with gH, independently from gL, which is present on the virion
surface.
Our analysis showed that the mature form of UL116 is heavily
glycosylated, principally by Golgi-type carbohydrate chains. An
ER-type glycosylated UL116 of lower mass is constantly present
over the infection cycle, likely as an immature precursor, but it was
not found to be present on the virion. Following translocation
from the ER, UL116 is targeted to the intracellular virion AC and
is incorporated into virions. Transient expression experiments
with UL116 expressed as single HCMV species revealed that the
UL116 protein is retained in the ER and not secreted or targeted to
the plasma membrane. Among the HCMV envelope proteins we
tried in cotransfection, gH was the only one that was able to release
UL116 ER retention, suggesting the formation of a heterodimeric
gH/UL116 complex. This hypothesis was confirmed by
experi-ments in HCMV-infected cells.
So far the only dimeric complex known for HCMV gH is the
disulfide-linked gH/gL. Although it was reported to be sufficient
(with gB) for entry into fibroblasts (
75
), a more recent analysis has
shown that on the surface of different HCMV strains there are two
FIG 7Western blotting and SDS-PAGE for gH/gL, gH/gL/UL116, and gH/UL116 transfections. (A) Anti-gL Western blotting (nonreducing) on conditioned media from Expi293 cells transiently transfected with DNA encoding the indicated proteins. (B) gH-6His/UL116-strep and gH-6His/gL/UL116-strep samples were Strep-tag affinity purified from the medium, and the elution fraction was run in SDS-PAGE. Coomassie-stained nonreducing SDS-PAGE gels show purified gH/gL and gH/UL116 (controls) followed by strep elution samples. (C and D) Anti-UL128 (C) and anti-His (D) Western blotting (nonreducing) on purified protein alongside conditioned medium from Expi293 cells transiently transfected with the DNA encoding the indicated proteins. Samples containing UL116-strep then were Strep-tag affinity purified from the medium, and the elution fractions were run in SDS-PAGE. Coomassie-stained nonreducing SDS-PAGE gel showing elution fractions are in thickly lined boxes in panels C and D.HCMV gH and UL116 Form a Complex Alternative to gH/gL
on November 7, 2019 by guest
http://jvi.asm.org/
[image:10.585.111.475.66.490.2]distinct complexes assembling on the gH/gL scaffold: the trimeric
gH/gL/gO and the pentameric gH/gL/UL128/UL130/UL131A
complex (
76
). The efficiency of infection of specific cell types,
perhaps by distinct entry pathways, could depend on relative
ra-tios between these complexes on the envelope (
76
). Replication in
distinctive cell types generates viral populations carrying different
ratios of these complexes on their envelope (
77
). The association
of gH/gL with these alternative complexes relies on the formation
of a disulfide bond with either gO or UL128. Both proteins target
the same Cys residue on gL, thus forming mutually exclusive
gH/gL complexes (
54
). Both gO and UL128/UL130/UL131A
oc-cupy the N-terminal region of gH and were proposed to govern
the specificity of receptor interaction (
54
). EM analysis of purified
gH/UL116 indicated that the complex has the same overall
orga-nization of gH/gL and that UL116 might share the gH binding site
of gL, although gH binding to UL116 is not mediated by disulfide
bond formation. Peculiarly, the
UL116
gene is adjacent to that
encoding gL (
UL115
). Although not a rule, it has been noticed that
in the HCMV genome, genes with similar functions are in close
proximity to each other (
8
). Complex formation analysis by the
coexpression of gH, gL, and UL116 and subsequent SEC and
im-munoprecipitation experiments confirmed that gL did not
asso-ciate with the gH/UL116 heterodimer. Furthermore, UL116 not
only displaces gL from gH but also prevents binding of gO and
UL128/130/131.
A very recent report described how the production of
alterna-tive gH-based complexes is regulated. The product of the
UL148
gene is an ER glycoprotein that, in this compartment, interacts
with the gH/gL heterodimer in the absence of either gO or UL128
and favors the formation of the gH/gL/gO complex (
78
). The
as-sociation of UL116 with gH appears to take place in the ER, so we
cannot exclude that a similar complex switch is driven by UL116
and that the heterodimer gH/UL116 represents the core scaffold
for an unidentified complex that excludes gL but could recruit
additional factors. However, we showed that unlike UL148,
UL116 is present in the virion.
gH/UL116 is a novel gH-based complex, and it is tempting to
speculate that it could act as a ligand for a different receptor, thus
amplifying the potential for the specific recognition of human cell
types. Intriguingly, the restoration of Pentamer expression in the
AD169 laboratory strains induces the increased expression of
UL116 (
38
), although the biological significance has not been
fur-ther investigated. One can speculate that the increased expression
of UL116 somehow is linked to the acquisition of a functional
Pentamer and the concomitant expansion of cellular tropism. Our
coimmunoprecipitation experiments performed in HFF-1 cells
infected with the TR strain suggest that the amount of gH/UL116
complex is considerably lower than that of gH/gL complexes.
Al-though we did not explore this observation further, it would be
interesting to assess the ratio of gH/UL116 to gH/gL complexes in
viruses with different genetic backgrounds or grown in different
cell lines.
In conclusion, our results add a new layer of complexity by
identifying for the first time a stable herpesvirus protein complex
that contains gH but not gL. Given that gH-containing complexes
have great potential as vaccine antigens, a better understanding of
these antibody targets may guide the design of improved vaccine
antigens or antibody-based therapies.
ACKNOWLEDGMENTS
The SEC-LS/UV/RI instrumentation was supported by NIH award 1S10RR023748-01. The MALS experiments and analysis were performed at the Biophysics Resource of the Keck Facility at Yale University.
The content of this paper is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
FUNDING INFORMATION
This work, including the efforts of Stefano Caló, Mirko Cortese, Claudio Ciferri, Luca Bruno, Rachel Gerrein, Barbara Benucci, Giuseppina Monda, Tobias Kessler, Yasushi Uematsu, Domenico Maione, Anders E. Lilja, and Andrea Carfí, was funded by GSK Vaccines (previously Novartis Vaccines).
REFERENCES
1.Mocarski JE, Shenk T, Griffiths PD, Pass RF.2013. Cytomegaloviruses, p 1960 –2014.InKnipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed. Lip-pincott Williams & Wilkins, Philadelphia, PA.
2.Cannon MJ.2009. Congenital cytomegalovirus (CMV) epidemiology and awareness. J Clin Virol46(Suppl 4):S6 –S10.http://dx.doi.org/10.1016/j .jcv.2009.09.002.
3.Britt W.2008. Manifestations of human cytomegalovirus infection: pro-posed mechanisms of acute and chronic disease. Curr Topics Microbiol Immunol325:417– 470.
4.Krause PR, Bialek SR, Boppana SB, Griffiths PD, Laughlin CA, Ljung-man P, Mocarski ES, Pass RF, Read JS, Schleiss MR, Plotkin SA.2013. Priorities for CMV vaccine development. Vaccine32:4 –10.http://dx.doi .org/10.1016/j.vaccine.2013.09.042.
5.Stratton KR, Durch JS, Lawrence RS.2001. Vaccines for the 21st century: a tool for a decisionmaking. National Academy Press, Washington, DC. 6.Murphy E, Yu D, Grimwood J, Schmutz J, Dickson M, Jarvis MA, Hahn
G, Nelson JA, Myers RM, Shenk TE.2003. Coding potential of labora-tory and clinical strains of human cytomegalovirus. Proc Natl Acad Sci U S A100:14976 –14981.http://dx.doi.org/10.1073/pnas.2136652100. 7.Davison AJ, Dolan A, Akter P, Addison C, Dargan DJ, Alcendor DJ,
McGeoch DJ, Hayward GS.2003. The human cytomegalovirus genome revisited: comparison with the chimpanzee cytomegalovirus genome. J Gen Virol84:17–28.
8.Murphy E, Shenk T.2008. Human cytomegalovirus genome. Curr Top-ics Microbiol Immunol325:1–19.
9.Dolan A, Cunningham C, Hector RD, Hassan-Walker AF, Lee L, FIG 8Single-particle EM of the gH/UL116 complex. Purified gH/UL116/
MSL-109 (A), gH/UL116/3G16 (B), and gH/gL_C144S/3G16 (C) ternary complexes were analyzed by single-particle electron microscopy. Individual particles, representing a specific 2D view of the complex, were aligned and used to generate 2D class averages. Comparison among the class averages of these 3 complexes and also with previous ones obtained for gH/gL and gH/gL/gO bound to MSL-109 or 3G16 (72) suggest the position of individual subunits and Fabs.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:11.585.64.264.64.227.2]Addison C, Dargan DJ, McGeoch DJ, Gatherer D, Emery VC, Griffiths PD, Sinzger C, McSharry BP, Wilkinson GW, Davison AJ.2004. Genetic content of wild-type human cytomegalovirus. J Gen Virol85:1301–1312. 10. Shikhagaie M, Merce-Maldonado E, Isern E, Muntasell A, Alba MM, Lopez-Botet M, Hengel H, Angulo A.2012. The human cytomegalovi-rus-specific UL1 gene encodes a late-phase glycoprotein incorporated in the virion envelope. J Virol86:4091– 4101.http://dx.doi.org/10.1128/JVI .06291-11.
11. Stanton RJ, Baluchova K, Dargan DJ, Cunningham C, Sheehy O, Seirafian S, McSharry BP, Neale ML, Davies JA, Tomasec P, Davison AJ, Wilkinson GW.2010. Reconstruction of the complete human cyto-megalovirus genome in a BAC reveals RL13 to be a potent inhibitor of replication. J Clin Investig 120:3191–3208. http://dx.doi.org/10.1172 /JCI42955.
12. Varnum SM, Streblow DN, Monroe ME, Smith P, Auberry KJ, Pasa-Tolic L, Wang D, Camp DG, II, Rodland K, Wiley S, Britt W, Shenk T, Smith RD, Nelson JA.2004. Identification of proteins in human cyto-megalovirus (HCMV) particles: the HCMV proteome. J Virol78:10960 – 10966.http://dx.doi.org/10.1128/JVI.78.20.10960-10966.2004. 13. Isaacson MK, Juckem LK, Compton T.2008. Virus entry and innate
immune activation. Curr Topics Microbiol Immunol325:85–100. 14. Jiang XJ, Adler B, Sampaio KL, Digel M, Jahn G, Ettischer N, Stierhof
YD, Scrivano L, Koszinowski U, Mach M, Sinzger C.2008. UL74 of human cytomegalovirus contributes to virus release by promoting sec-ondary envelopment of virions. J Virol82:2802–2812.http://dx.doi.org /10.1128/JVI.01550-07.
15. Wille PT, Knoche AJ, Nelson JA, Jarvis MA, Johnson DC. 2010. A human cytomegalovirus gO-null mutant fails to incorporate gH/gL into the virion envelope and is unable to enter fibroblasts and epithelial and endothelial cells. J Virol 84:2585–2596. http://dx.doi.org/10.1128/JVI .02249-09.
16. Zhou M, Lanchy JM, Ryckman BJ.2015. Human cytomegalovirus gH/ gL/gO promotes the fusion step of entry into all cell types, whereas gH/gL/ UL128-131 broadens virus tropism through a distinct mechanism. J Virol
89:8999 –9009.http://dx.doi.org/10.1128/JVI.01325-15.
17. Compton T.2004. Receptors and immune sensors: the complex entry path of human cytomegalovirus. Trends Cell Biol14:5– 8.http://dx.doi .org/10.1016/j.tcb.2003.10.009.
18. Pellett PE, Roizman B.2013. Herpesviridae, p 1802–1822.InKnipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed. Lippincott Williams & Wilkins, Philadelphia, PA.
19. Compton T, Nepomuceno RR, Nowlin DM.1992. Human cytomegalo-virus penetrates host cells by pH-independent fusion at the cell surface. Virology191:387–395.http://dx.doi.org/10.1016/0042-6822(92)90200-9. 20. Haspot F, Lavault A, Sinzger C, Laib Sampaio K, Stierhof YD, Pilet P, Bressolette-Bodin C, Halary F.2012. Human cytomegalovirus entry into dendritic cells occurs via a macropinocytosis-like pathway in a pH-independent and cholesterol-dependent manner. PLoS One7:e34795.
http://dx.doi.org/10.1371/journal.pone.0034795.
21. Ryckman BJ, Jarvis MA, Drummond DD, Nelson JA, Johnson DC.
2006. Human cytomegalovirus entry into epithelial and endothelial cells depends on genes UL128 to UL150 and occurs by endocytosis and low-pH fusion. J Virol 80:710 –722. http://dx.doi.org/10.1128/JVI.80.2.710-722 .2006.
22. Wang D, Yu QC, Schroer J, Murphy E, Shenk T.2007. Human cyto-megalovirus uses two distinct pathways to enter retinal pigmented epithe-lial cells. Proc Natl Acad Sci U S A104:20037–20042.http://dx.doi.org/10 .1073/pnas.0709704104.
23. Dunn W, Chou C, Li H, Hai R, Patterson D, Stolc V, Zhu H, Liu F.
2003. Functional profiling of a human cytomegalovirus genome. Proc Natl Acad Sci U S A100:14223–14228.http://dx.doi.org/10.1073/pnas .2334032100.
24. Yu D, Silva MC, Shenk T.2003. Functional map of human cytomegalo-virus AD169 defined by global mutational analysis. Proc Natl Acad Sci U S A100:12396 –12401.http://dx.doi.org/10.1073/pnas.1635160100. 25. Hobom U, Brune W, Messerle M, Hahn G, Koszinowski UH.2000. Fast
screening procedures for random transposon libraries of cloned herpesvi-rus genomes: mutational analysis of human cytomegaloviherpesvi-rus envelope glycoprotein genes. J Virol74:7720 –7729.http://dx.doi.org/10.1128/JVI .74.17.7720-7729.2000.
26. Wang D, Shenk T.2005. Human cytomegalovirus virion protein complex
required for epithelial and endothelial cell tropism. Proc Natl Acad Sci U S A102:18153–18158.http://dx.doi.org/10.1073/pnas.0509201102. 27. Hahn G, Revello MG, Patrone M, Percivalle E, Campanini G, Sarasini
A, Wagner M, Gallina A, Milanesi G, Koszinowski U, Baldanti F, Gerna G.2004. Human cytomegalovirus UL131-128 genes are indispensable for virus growth in endothelial cells and virus transfer to leukocytes. J Virol
78:10023–10033.http://dx.doi.org/10.1128/JVI.78.18.10023-10033.2004. 28. Straschewski S, Patrone M, Walther P, Gallina A, Mertens T, Frascaroli G.2011. Protein pUL128 of human cytomegalovirus is necessary for monocyte infection and blocking of migration. J Virol85:5150 –5158.
http://dx.doi.org/10.1128/JVI.02100-10.
29. Li Q, Wilkie AR, Weller M, Liu X, Cohen JI.2015. THY-1 cell surface antigen (CD90) has an important role in the initial stage of human cyto-megalovirus infection. PLoS Pathog 11:e1004999.http://dx.doi.org/10 .1371/journal.ppat.1004999.
30. Britt WJ, Vugler L, Butfiloski EJ, Stephens EB.1990. Cell surface ex-pression of human cytomegalovirus (HCMV) gp55-116 (gB): use of HCMV-recombinant vaccinia virus-infected cells in analysis of the hu-man neutralizing antibody response. J Virol64:1079 –1085.
31. Fouts AE, Chan P, Stephan JP, Vandlen R, Feierbach B.2012. Antibod-ies against the gH/gL/UL128/UL130/UL131 complex comprise the major-ity of the anti-cytomegalovirus (anti-CMV) neutralizing antibody re-sponse in CMV hyperimmune globulin. J Virol86:7444 –7447.http://dx .doi.org/10.1128/JVI.00467-12.
32. Cranage MP, Kouzarides T, Bankier AT, Satchwell S, Weston K, Tom-linson P, Barrell B, Hart H, Bell SE, Minson AC.1986. Identification of the human cytomegalovirus glycoprotein B gene and induction of neu-tralizing antibodies via its expression in recombinant vaccinia virus. EMBO J5:3057–3063.
33. Lilja AE, Mason PW.2012. The next generation recombinant human cytomegalovirus vaccine candidates-beyond gB. Vaccine30:6980 – 6990.
http://dx.doi.org/10.1016/j.vaccine.2012.09.056.
34. Pass RF, Zhang C, Evans A, Simpson T, Andrews W, Huang ML, Corey L, Hill J, Davis E, Flanigan C, Cloud G.2009. Vaccine prevention of maternal cytomegalovirus infection. N Engl J Med360:1191–1199.http: //dx.doi.org/10.1056/NEJMoa0804749.
35. Wen Y, Monroe J, Linton C, Archer J, Beard CW, Barnett SW, Palladino G, Mason PW, Carfi A, Lilja AE.2014. Human cytomeg-alovirus gH/gL/UL128/UL130/UL131A complex elicits potently neu-tralizing antibodies in mice. Vaccine32:3796 –3804.http://dx.doi.org /10.1016/j.vaccine.2014.05.004.
36. Wussow F, Chiuppesi F, Martinez J, Campo J, Johnson E, Flechsig C, Newell M, Tran E, Ortiz J, La Rosa C, Herrmann A, Longmate J, Chakraborty R, Barry PA, Diamond DJ.2014. Human cytomegalovirus vaccine based on the envelope gH/gL pentamer complex. PLoS Pathog
10:e1004524.http://dx.doi.org/10.1371/journal.ppat.1004524.
37. Kabanova A, Perez L, Lilleri D, Marcandalli J, Agatic G, Becattini S, Preite S, Fuschillo D, Percivalle E, Sallusto F, Gerna G, Corti D, Lanzavecchia A.2014. Antibody-driven design of a human cytomegalo-virus gHgLpUL128L subunit vaccine that selectively elicits potent neutral-izing antibodies. Proc Natl Acad Sci U S A111:17965–17970.http://dx.doi .org/10.1073/pnas.1415310111.
38. Freed DC, Tang Q, Tang A, Li F, He X, Huang Z, Meng W, Xia L, Finnefrock AC, Durr E, Espeseth AS, Casimiro DR, Zhang N, Shiver JW, Wang D, An Z, Fu TM.2013. Pentameric complex of viral glyco-protein H is the primary target for potent neutralization by a human cytomegalovirus vaccine. Proc Natl Acad Sci U S A110:E4997–E5005.
http://dx.doi.org/10.1073/pnas.1316517110.
39. Lilleri D, Kabanova A, Revello MG, Percivalle E, Sarasini A, Genini E, Sallusto F, Lanzavecchia A, Corti D, Gerna G. 2013. Fetal human cytomegalovirus transmission correlates with delayed maternal antibodies to gH/gL/pUL128-130-131 complex during primary infection. PLoS One
8:e59863.http://dx.doi.org/10.1371/journal.pone.0059863.
40. Lilleri D, Kabanova A, Lanzavecchia A, Gerna G. 2012. Antibodies against neutralization epitopes of human cytomegalovirus gH/gL/ pUL128-130-131 complex and virus spreading may correlate with virus control in vivo. J Clin Immunol32:1324 –1331.http://dx.doi.org/10.1007 /s10875-012-9739-3.
41. Macagno A, Bernasconi NL, Vanzetta F, Dander E, Sarasini A, Revello MG, Gerna G, Sallusto F, Lanzavecchia A.2010. Isolation of human monoclonal antibodies that potently neutralize human cytomegalovirus infection by targeting different epitopes on the gH/gL/UL128-131A com-plex. J Virol84:1005–1013.http://dx.doi.org/10.1128/JVI.01809-09.
HCMV gH and UL116 Form a Complex Alternative to gH/gL
on November 7, 2019 by guest
http://jvi.asm.org/
42. Cortese M, Calo S, D’Aurizio R, Lilja A, Pacchiani N, Merola M.2012. Recombinant human cytomegalovirus (HCMV) RL13 binds human im-munoglobulin G Fc. PLoS One 7:e50166. http://dx.doi.org/10.1371 /journal.pone.0050166.
43. Spaderna S, Blessing H, Bogner E, Britt W, Mach M.2002. Identifica-tion of glycoprotein gpTRL10 as a structural component of human cyto-megalovirus. J Virol 76:1450 –1460. http://dx.doi.org/10.1128/JVI.76.3 .1450-1460.2002.
44. Spaderna S, Kropff B, Kodel Y, Shen S, Coley S, Lu S, Britt W, Mach M.
2005. Deletion of gpUL132, a structural component of human cytomega-lovirus, results in impaired virus replication in fibroblasts. J Virol79:
11837–11847.http://dx.doi.org/10.1128/JVI.79.18.11837-11847.2005. 45. Huber MT, Compton T.1998. The human cytomegalovirus UL74 gene
encodes the third component of the glycoprotein H-glycoprotein L-con-taining envelope complex. J Virol72:8191– 8197.
46. Chang CP, Vesole DH, Nelson J, Oldstone MB, Stinski MF. 1989. Identification and expression of a human cytomegalovirus early glycopro-tein. J Virol63:3330 –3337.
47. Fraile-Ramos A, Pelchen-Matthews A, Kledal TN, Browne H, Schwartz TW, Marsh M.2002. Localization of HCMV UL33 and US27 in endocytic compartments and viral membranes. Traffic3:218 –232.http://dx.doi.org /10.1034/j.1600-0854.2002.030307.x.
48. Margulies BJ, Browne H, Gibson W.1996. Identification of the human cytomegalovirus G protein-coupled receptor homologue encoded by UL33 in infected cells and enveloped virus particles. Virology225:111– 125.http://dx.doi.org/10.1006/viro.1996.0579.
49. Smith IL, Taskintuna I, Rahhal FM, Powell HC, Ai E, Mueller AJ, Spector SA, Freeman WR.1998. Clinical failure of CMV retinitis with intravitreal cidofovir is associated with antiviral resistance. Arch Ophthal-mol116:178 –185.
50. Murphy E, Rigoutsos I, Shibuya T, Shenk TE.2003. Reevaluation of human cytomegalovirus coding potential. Proc Natl Acad Sci U S A100:
13585–13590.http://dx.doi.org/10.1073/pnas.1735466100.
51. Talbot P, Almeida JD.1977. Human cytomegalovirus: purification of enveloped virions and dense bodies. J Gen Virol36:345–349.
52. Tischer BK, von Einem J, Kaufer B, Osterrieder N.2006. Two-step red-mediated recombination for versatile high-efficiency markerless DNA manipulation in Escherichia coli. Biotechniques40:191–197.http://dx .doi.org/10.2144/000112096.
53. Irmiere A, Gibson W.1983. Isolation and characterization of a nonin-fectious virion-like particle released from cells infected with human strains of cytomegalovirus. Virology130:118 –133.http://dx.doi.org/10 .1016/0042-6822(83)90122-8.
54. Ciferri C, Chandramouli S, Donnarumma D, Nikitin PA, Cianfrocco MA, Gerrein R, Feire AL, Barnett SW, Lilja AE, Rappuoli R, Norais N, Settembre EC, Carfi A. 2015. Structural and biochemical studies of HCMV gH/gL/gO and Pentamer reveal mutually exclusive cell entry com-plexes. Proc Natl Acad Sci U S A112:1767–1772.http://dx.doi.org/10 .1073/pnas.1424818112.
55. Arakawa T, Langley KE, Kameyama K, Takagi T. 1992. Molecular weights of glycosylated and nonglycosylated forms of recombinant human stem cell factor determined by low-angle laser light scattering. Anal Biochem203:53–57.http://dx.doi.org/10.1016/0003-2697(92)90042-6. 56. Hayashi Y, Matsui H, Takagi T. 1989. Membrane protein molecular
weight determined by low-angle laser light-scattering photometry cou-pled with high-performance gel chromatography. Methods Enzymol172:
514 –528.http://dx.doi.org/10.1016/S0076-6879(89)72031-0.
57. Folta-Stogniew E.2006. Oligomeric states of proteins determined by size-exclusion chromatography coupled with light scattering, absorbance, and refractive index detectors. Methods Mol Biol328:97–112.
58. Tang G, Peng L, Baldwin PR, Mann DS, Jiang W, Rees I, Ludtke SJ.2007. EMAN2: an extensible image processing suite for electron microscopy. J Struct Biol157:38 – 46.http://dx.doi.org/10.1016/j.jsb.2006.05.009. 59. van Heel M, Harauz G, Orlova EV, Schmidt R, Schatz M.1996. A new
generation of the IMAGIC image processing system. J Struct Biol116:17– 24.http://dx.doi.org/10.1006/jsbi.1996.0004.
60. Leatham MP, Witte PR, Stinski MF.1991. Alternate promoter selection within a human cytomegalovirus immediate-early and early transcription unit (UL119-115) defines true late transcripts containing open reading frames for putative viral glycoproteins. J Virol65:6144 – 6153.
61. Mockarski JE, Shenk T, Griffiths P, Pass R.2013. Cytomegaloviruses, p
1960 –2014.InKnipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed. Lip-pincott Williams & Wilkins, Philadelphia, PA.
62. Malouli D, Nakayasu ES, Viswanathan K, Camp DG, II, Chang WL, Barry PA, Smith RD, Fruh K.2012. Reevaluation of the coding potential and proteomic analysis of the BAC-derived rhesus cytomegalovirus strain 68-1. J Virol86:8959 – 8973.http://dx.doi.org/10.1128/JVI.01132-12. 63. Kanai K, Yamada S, Yamamoto Y, Fukui Y, Kurane I, Inoue N.2011.
Re-evaluation of the genome sequence of guinea pig cytomegalovirus. J Gen Virol92:1005–1020.
64. Rawlinson WD, Farrell HE, Barrell BG.1996. Analysis of the complete DNA sequence of murine cytomegalovirus. J Virol70:8833– 8849. 65. Kohler CP, Kerry JA, Carter M, Muzithras VP, Jones TR, Stenberg RM.
1994. Use of recombinant virus to assess human cytomegalovirus early and late promoters in the context of the viral genome. J Virol68:6589 – 6597.
66. Johnson DC, Spear PG.1983. O-linked oligosaccharides are acquired by herpes simplex virus glycoproteins in the Golgi apparatus. Cell32:987– 997.http://dx.doi.org/10.1016/0092-8674(83)90083-1.
67. Maley F, Trimble RB, Tarentino AL, Plummer TH, Jr.1989. Charac-terization of glycoproteins and their associated oligosaccharides through the use of endoglycosidases. Anal Biochem180:195–204.http://dx.doi.org /10.1016/0003-2697(89)90115-2.
68. Sanchez V, Greis KD, Sztul E, Britt WJ.2000. Accumulation of virion tegument and envelope proteins in a stable cytoplasmic compartment during human cytomegalovirus replication: characterization of a poten-tial site of virus assembly. J Virol74:975–986.http://dx.doi.org/10.1128 /JVI.74.2.975-986.2000.
69. Sanchez V, Sztul E, Britt WJ. 2000. Human cytomegalovirus pp28 (UL99) localizes to a cytoplasmic compartment which overlaps the endo-plasmic reticulum-Golgi-intermediate compartment. J Virol74:3842– 3851.http://dx.doi.org/10.1128/JVI.74.8.3842-3851.2000.
70. Ryckman BJ, Rainish BL, Chase MC, Borton JA, Nelson JA, Jarvis MA, Johnson DC.2008. Characterization of the human cytomegalovirus gH/ gL/UL128-131 complex that mediates entry into epithelial and endothelial cells. J Virol82:60 –70.http://dx.doi.org/10.1128/JVI.01910-07. 71. Loomis RJ, Lilja AE, Monroe J, Balabanis KA, Brito LA, Palladino G,
Franti M, Mandl CW, Barnett SW, Mason PW. 2013. Vectored co-delivery of human cytomegalovirus gH and gL proteins elicits potent com-plement-independent neutralizing antibodies. Vaccine31:919 –926.http: //dx.doi.org/10.1016/j.vaccine.2012.12.009.
72. Ciferri C, Chandramouli S, Leitner A, Donnarumma D, Cianfoc