Expression of the Varicella-Zoster Virus
Origin-Binding
Protein
and
Analysis of Its
Site-Specific DNA-Binding Properties
DANCHENANDPAULD. OLIVO*
Departmentof Medicine, Divisionof Infectious Diseases, Washington University Schoolof Medicine, St. Louis, Missouri63110
Received18 November 1993/Accepted 1 March1994
The varicella-zoster virus (VZV)genome contains homologs to each of the seven herpes simplex virus(HSV) genes that arerequired for viral DNAsynthesis.VZVgene51 ishomologous toHSVUL9, whichencodes an originof DNAreplication binding protein(OBP).It waspreviously shown, by usingaproteinAfusionprotein, that the productofgene51 is a site-specificDNA-binding proteinwhich binds to sequences within the VZV origin(Stow et al., Virology177:570-577, 1990).In thisreport, gene51 wasexpressedin an in vitro translation system.Rabbit antiserumraisedagainstthecarboxyl-terminal 20 amino acidswasusedtoconfirm expression of thefull-length gene51protein, andsite-specific DNA-bindingactivitywasdemonstratedin agel retardation assay.Theorigin-bindingdomain waslocated within a 263-amino-acidregionofthecarboxyl terminus byusing aseries of deletion mutants.Theaffinity ofbinding ofthe VZVOBPto thethreebinding sitesintheVZVorigin wasfoundtobe similar.Inaddition, as withUL9, a CGC triplet within a 10-bp consensus sequence is critical totheinteraction between the OBP and the origin. The HSV andVZVOBPs, therefore, appear to have
virtually
identical recognition sequences despiteonly 33%identityand 44%similarityin the primary structure oftheir site-specific DNA-binding domains.Mammalian DNAviruses have proven to beuseful model systems forthe study of eukaryotic DNAreplication (3, 20). Herpesviruses, which have large linear DNA genomes, offer manyadvantages for DNA replication studies. Herpes simplex virus(HSV) is the best characterized of the herpesviruses. Its genomeencodes about 72 genes, of which 7 have been shown to be required for viral DNA synthesis (2, 15, 18, 26). In
addition, three origins ofDNA replication have been
identi-fied, onecopy of
o0L
andtwo copies ofons
(19, 21, 24,25).Varicella-zoster virus (VZV) is a human herpesvirus which, like HSV, belongs to the subfamily
Alphaherpesvirinae
(14).Despite significantdifferences in their biologicbehavior, VZV and HSV share many structural and molecular biological features. Sequence analysis has revealed that the 125-kb linear double-stranded DNA genome ofVZV has homologs to all sevenof the essential HSVDNAreplicationgenes(5, 15).The VZV genome has no equivalent to HSVOnL, but, like HSV, theVZVgenomehastwocopiesof
onis
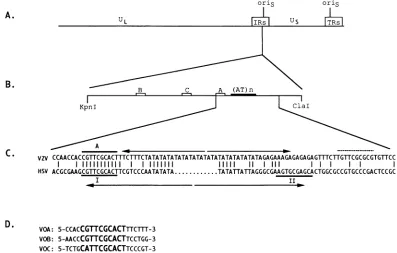
in theinverted repeatsflankingthe shortunique region of the genome (Fig. 1A).
VZVgene 51 ishomologoustoHSV UL9 which encodes a
protein thatbindstothe viralDNAorigins of replication and isthoughttoplayanimportantrole ininitiationof viralDNA
synthesis (9, 17). The predicted amino acid sequence of the gene51 proteinwasshowntobe44%identicaltoUL9, and it was expected that the product of gene 51 would be a VZV
origin-binding protein (OBP) (5, 15).Consistent with thisidea,
a proteinA-gene 51 fusionproteinwasshown to bind to the VZV origin, and, aswith UL9, the origin-binding domain of the VZV OBP mapped to the carboxyl terminus (23). It is likely that the VZVOBPplays the samerole as UL9 during viral DNA replication; however, there are no genetic data to support this idea and biochemical studies of the VZV OBP
*Corresponding author. Mailing address: Department ofMedicine,
Division of Infectious Diseases, Washington University School of Medicine,Box8051,660 S. EuclidAve., St.Louis,MO63110. Phone: (314)362-5718. Fax:(314)362-9230.Electronic mail address:Olivo@ borcim.wustl.edu.
have been limited to the fusionproteinmentioned above.An
origin-binding activity has not been identified in extracts of VZV-infected cells, and expression of the full-length VZV OBPinaheterologous system has notbeenreported.
Despite both structural and functional similarities, the HSV and VZV origins differ in the arrangement of their OBP
binding sites (Fig. 1B and C). HSV
ons
has two high-affinity bindingsites whicharepartofanimperfect palindromewitha central AT-rich region. VZVoris
also contains an AT-rich region, but it has three OBP binding sites which are all located onone side of the AT-rich region(4, 22, 23) (Fig. 1C). Thesignificanceofthis structural difference between the VZV and HSV origins remains to be determined, but it is likely that
comparative studies of the HSV and VZV DNA replication
systems willcontribute to ourknowledge of the mechanisms involved inherpesvirusDNAsynthesis. Specifically,studieson the VZV OBPnotonlywilldefineits role in the initiation of VZVDNAreplication,but will advanceourunderstandingof the role of UL9 in HSV DNA replication. This approach is initiated in this report, in which theexpression and functional characterization of the VZVOBParedescribed.
MATERIALSAND METHODS
Cloningofgene 51 into a vector for in vitrotranscription and translation. The entire gene 51 open reading frame was cloned intopTM1,aplasmid originallyconstructed formaking
recombinant vaccinia viruses (16). pTM1 contains,in a 5'-3'
orientation, the bacteriophage T7 promoter, the
encephalo-myocarditis virus internal ribosome entry site which facilitates
cap-independent ribosome binding, a multiple cloning site,
translation stop codonsinall threereadingframes,and theT7 transcription terminal signal.A uniqueNcoI site, which
con-tainsanATG, is 5' ofall otherrestriction enzymesites within themultiple cloningsite. Priortocloninggene51 intopTM1,
theNcoI site and ATG were removedby digestionwithNcoI,
digestion of the 5' protruding end with mung bean
nuclease,
andreligationof the bluntendstogeneratepMH4.TheVZV3841
on November 9, 2019 by guest
http://jvi.asm.org/
3842 CHEN AND OLIVO
oris
A. u US
B.\
KpnI kClal
VZV
CCAACCACCGTTCGCACTTCTTITCTATATATATATATATATATATATATATATATAGAGAAAGAGAGAGAGTTITCTTGTTCGCGCGTGTTCC
I 1 11111111111 111111111 11111 11 1i111 1 1I
HSV ACGCGAAGCGTTCGCACTTCGTCCCAATATATA...TATATTATTAGGGCGAAGTGCGAGCACTGGCGCCGTGCCCGACTCCGC
I II,_
D.
VOA: 5-CCACCGTTCGCACTTTCT1T-3
VOB: 5-AACCCGTTCGCACTTCCTGG-3
VOC: 5-TCTGCATTCGCACTTCCCGT-3
FIG. 1. Schematic of the VZV origin.(A)Location of the two copies ofonsin VZV genome. (B) Structure of
on.s
BoxeslabeledA, B, and Cindicate the three 51 geneproductbindingsites whichweredeterminedbyStowetal.(23). (AT)nrepresents theAT-richregion.(C) Comparison of VZV and HSVoriginsequences.The46-bp palindromicsequence inthe VZVoriginis markedby divergentarrowsabove the top line. Note that thepalindromedoesnotinclude site A which isindicated byaheavyline above the VZV sequence. HSV site I and site IIaremarkedbya heavy line, and the palindrome, whichincludesboth siteIand siteII,is marked bydivergent arrows below the HSV sequence. The central dots inthe HSV sequenceindicate theshorter AT-richregion. Thedashedline abovetheVZV sequence totheright of the AT-richregionshowsa potential 51 binding site. (D) VOA, VOB, and VOC are three 20-meroligonucleotides which contain the threebindingsites A,B, and C, respectively, andwhichwereused in thecompetitive bindingassaysinFig.8. The 10 bases inbold indicatethelikelyOBPrecognitionsequence.gene 51 was obtained from a VZV genomic DNA library kindly provided by L.Gelb (WashingtonUniversity). Priorto cloning gene 51 into the pMH4 vector, a BamHI site was createdjustupstreamof thestartofthe gene 51 openreading frame byusing PCR-mediated mutagenesis. The entire gene 51,fromthisBamHI site (nucleotide sequence no. 87888) to a
StyI site (nucleotide sequence no. 90513), was cloned, using BamHI linkers, into the BamHI site of pMH4 to generate pMH4-51.
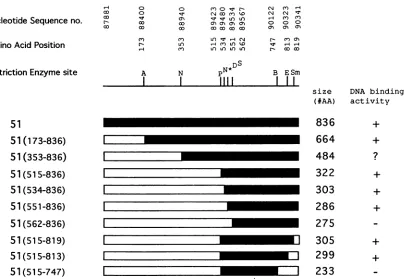
Construction of deletion mutants of the gene 51. Nine deletion mutants of gene51 wereconstructed in pTM1.Figure 2 shows schematically the position of each deletion. Prior to making the deletion mutations,anNcoI sitewascreatedatthe 5' end of the gene 51 open reading frame by using PCR-mediated mutagenesis. Gene 51 was then cloned into theNcoI andBamHI sites ofpTM1 togeneratepTM151. Five gene51 amino-terminal deletion mutants were generated from pTM151 by digestion withNcoI and five restriction enzymes with siteswithin the gene 51 openreading frame (AatII, NdeI, PvuII, Dral, and Spel). An additional deletion mutant was generated by using an NcoI site created by PCR-mediated mutagenesis. Three carboxyl-terminal deletion mutants were generated starting with pTM151-PvuII, one of the amino-terminal mutants, by digestion with one of three restriction sites within the gene 51 open reading frame (BglII, Eco47III,
andSmaI)andaStul siteatthe3' end of the multiple cloning site in the vector, followed byreligation. The positions of these restriction sites within the gene 51 open reading frame are shown inFig.2.
Production of antisera against the C terminus of the gene 51 product.A 20-amino-acid peptide, CELSPGSWPRINGAVN FESL, whose sequence was derived from the predicted car-boxyl terminus of the gene 51 open reading frame, was
synthesizedby the Protein ChemistryLaboratoryat Washing-tonUniversity. This peptidewasconjugatedtokeyhole limpet hemocyanin by using
m-maleimidobenzoyl-N-hydroxysuccin-imide ester (Pierce Corp., Rockford, Ill.). The conjugated
peptidewasusedtoimmunizerabbits(1mg perrabbit in three injections). The activity of the antiserum was tested by immu-noblot analysis using gene 51 protein made in insect cells infected with a recombinant baculovirus. One such rabbit antiserum (R309) and the preimmunization serum from the samerabbit(pre-R309)wereused for allimmunoprecipitation and"double-retardation" experiments described.
Invitrotranscription and translationofgene 51.All invitro transcription-translation reactions were done by using theTnT coupled reticulocyte lysate system and T7 RNA polymerase
according to the manufacturer's recommendations (Promega
Corp., Madison, Wis.). Fourmicroliters ofL-[35S]methionine
(1,000 Ci/mmol;AmershamCorp.,ArlingtonHeights,Ill.)was added to the50-pI reaction togenerate radiolabeled invitro synthesizedproteins. Thesameprotocolwasusedtosynthesize unlabeled gene 51 protein for use in gel retardation assays except thatadditional L-methioninewasusedbyadding1
RI
of the leucine-minus amino acid mixprovidedby thesupplier inplace of
L-[35S]-methinone.
SDS.PAGE and autoradiography. Sodium dodecyl sulfate
(SDS)-polyacrylamide gel electrophoresis (PAGE) ofL-[35S]
J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.106.503.78.331.2]Nucleotide Sequenceno.
AminoAcid Position
0 o nos
0a 0 v 0 O) )
- a 0 0) 0) 0)an
OX a, a, 00, co
CY) CY) *nf v -iN N- c
N LO 'A LO0'I V rrs
C') Ur) LO) LO LO r- 0 O
RestrictionEnzymesite A N PN
N*DS
I
I
1111
BI II
ESmsize DNA binding (#AA) activity
51
51 (173-836)
51 (353-836)
51(515-836)
51(534-836)
51
(551-836)51
(562-836)51(515-819)
51
(515-813)51
(515-747)I
I
I
FIG. 2. Schematic representation ofVZVgene51deletionmutants.White barsindicate the regions deleted. Nucleotidesequence no.refers
to thegenomicsequenceof Davison and Scott(5).Aminoacidposition,in thecaseoftheamino-terminal deletions,indicatestheresidue which immediately follows the initiation methioninederivedfrom thevector.Inthecaseof thecarboxyl-terminaldeletions, amino acidpositionindicates the lastresidue of themutantpolypeptide derived from the 51sequence. Restriction sites usedtogeneratethemutantsare shownby letters.A, AatII; N,NdeI; P, PvuII; S, SpeI; D, DraI; B, BglII; E, Eco47III; Sm, SmaI.N*representsanNcoIsitewhich isnotpresentin thewild-typesequence
ofgene51 but whichwasgenerated by PCR mutagenesistoderive the51(5348-36)deletionmutant.The sizesrepresentthe number of amino acids inthemutanttranslation products.TheDNA-binding activitywasdetermined bygelretardationassaysasshown in FIG. 3 and FIG. 6.+,binding activity; -,nobindingactivity; ?, ambiguousresult.
methionine-labeled in vitro-synthesized proteins was carried
out as described in Promega's Protocols And Applications
Guide. After electrophoresis, the gel was fixed, soaked in
Amplify solution (Amersham Corp.), dried, and exposed to X-ray film (Fuji RX film; Fisher Scientific, St. Louis, Mo.) at
-700C.
Immunoprecipitation. One microliter of antiserum R309or
preimmune antiserum (pre-R309) was coupled to 20 of protein A-Sepharose (Sigma Co., St. Louis, Mo.) in 300 of phosphate-buffered salinewith 0.1% Tween 20(PT buffer) in arotatorovernightat40C.Thecoupled Sepharose beadswere
washed three times with PT buffer and once with PT buffer
containing 5mgof bovineserum albuminperml(PTB buffer). The L-[35S]methionine-labeled in vitro translation products
wereaddedtoatubecontaining the antiserum-coupled
Sepha-rosein300 of PTB buffer. After being rotated for 2 hat4°C, theSepharose beadswerewashed threetimeswith PTB buffer
andoncewith PT buffer.SDS-PAGE loading bufferwasadded
tothe pellet, and after being boiled for 2 min, theSepharose beads were centrifuged and the supernatant wasanalyzed by SDS-PAGE (10%acrylamide) and autoradiography.
Oligonucleotide probes usedin thegel retardationassays.A 26-base oligonucleotide containing site A of the VZV replica-tion origin (VOA-26f) (5'-CCAACCACCGYFCGCACTlFTC
TTTCT-3') and a complementary oligonucleotide (VOA26r)
were synthesized by the Protein Chemistry Laboratory of Washington University. VOA-26fwas radiolabeled at the 5'
end with [y-32P]ATP (4,500 Ci/mmol; Amersham Corp.) and T4polynucleotidekinase(Promega Corp.).Adouble-stranded site Aoligonucleotide (VOA-26)wasprepared by incubating
radiolabeled VOA-26f with excess unlabeled VOA-26r in
annealing buffer (10 mM Tris, pH 7.4, 100 mMNaCl, 5 mM MgCl2),for10minat65°C,10minat55°C,15minat42°C,and 60 min at 37°C. Unlabeled 20-mer double-stranded oligonu-cleotides usedin the competition gel retardation assays were
annealedin an identical manner.
Gel retardation assays.The DNA-protein binding reaction
wasperformed atroomtemperaturefor 15min. Each reaction
wasdone with 2to4 p.l of in vitro-synthesized protein, 104cpm
of radiolabeledprobe (0.8 x 106to1.0 x 106cpm/,ug)and 1 pLg
ofpoly(dI-dC) (Pharmacia Corp., Piscataway, N.J.) in a final volume of 25 ,u1. The incubation buffer (10 mM HEPES [N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid], [pH 7.9], 5 mM KCl, 2.5 mM MgCl2, 33.3 mM NaCl, 20 mMTris [pH 7.8], 3.8 mM EDTA, 5% glycerol, 0.03% Tween 20, 1.2 mM2-mercaptoethanol) andelectrophoresisbuffer(28.54gof glycine,6.04gofTrisbase,4 mlof 0.5 M EDTAperliter)were
made according to a commercial protocol (Stratagene, La
Jolla,Calif.).Afterincubation,sampleswereloadedontoa6% native polyacrylamide gel (acrylamide:bisacrylamide ratio of 80:1). After electrophoresis at 4°C for 1 h with a constant current of 20mA,thegelwasdried andexposedtoX-rayfilm. Fordouble-retardation experiments,0.2to0.5 pL. of antiserum wasaddedat thestart of the reaction.
(N4 C')14
(N (N sr
CD C' C')
o o o
0) cn 0
836
664
484
322
303
286
275
305
299
233
+
+
+
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.118.525.72.351.2]3844 CHEN AND OLIVO
1 2 3 4 5 6
200-
97-
69-4
6-
30-Z3 a
't_
.. F ....I
..-...I
....4
O46C-
w
~
S
w
2
1-1
4-FIG. 3. Immunoprecipitation andSDS-polyacrylamide gel analysis ofL-[35S]methionine-labeled 51 geneproductsynthesized in vitro by usingareticulocytelysate. Lane 1, full-length product of VZVgene51; lane 2,immunoprecipitation of full-length product of VZVgene51by anti-51 antiserum R309; lane 3, immunoprecipitation of full-length product of VZVgene51 usingpreimmune antiserum (pre-R309); lane 4,mutant 51(5,5-836)product (Fig. 2); lane 5, immunoprecipitation of
51(5,5-836)
product by R309 antiserum; lane 6, immunoprecipitation of 51(515836) product by preimmune antiserum (pre-R309). The sizes (kilodaltons) and positions of molecular weight markersareindicatedon the left.
Competitive gel retardation assays. Competitive gel retar-dation experimentswere performed with the same conditions
asstandard gel retardation assaysexceptthat unlabeled
com-petitoroligonucleotideswere addedatthestartof thebinding reaction. The dried gels were submitted to a BetaScope 603
Blot Analyzer (Betagen, A Division of Intelligenetics, Inc., Mountain View, Calif.) to quantify the radioactivity in the
1 2 3 4 5 6 7 8
_ ^ ^~~-C2
[image:4.612.88.263.72.234.2]-"~~~~~~.C
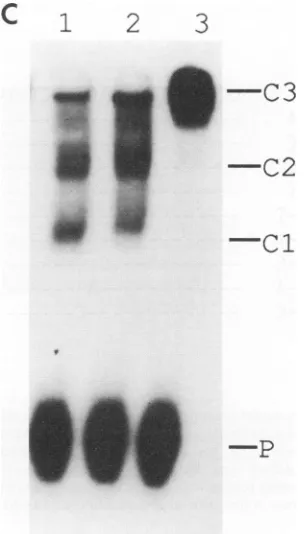
FIG. 4. Gel retardationassaydemonstrating the site-specific
bind-ing activity of in vitro translated product of VZVgene51. Theprobe
is a26-mer double-stranded oligonucleotide (VOA-26) which
corre-spondstothe DNAsequenceatsite A(see Materials and Methods).
Lane 1,controllysate;lanes 2and3, controllysatewith 0.2 and 0.5 ,ul ofanti-51 antiserum R309,respectively; lane 4, control lysate withgene
51 product; lanes 5 and 6, gene51 product with 0.2 and 0.5 .1l of control antiserum (pre-R309), respectively; lanes 7 and 8, 51 gene
[image:4.612.316.549.73.227.2]product with 0.2 and 0.5 ,ul ofanti-51 antiserum (R309), respectively. P, free probe; Cl, DNA protein complex; C2, DNA probe-protein-antibody complex.
FIG. 5. SDS-polyacrylamide gel analysis of
L-[35S]methionine-la-beledproductsofgene51deletionmutantssynthesizedin vitro.Lysate (2
[l)
wasloaded into each lane. Lane 1,full-lengthgene51product; lane2,51(173.836)
product; lane 3,51(353_836)product; lane4,51(l515836)
product; lane 5,51(534
8,36);
lane 6,51(55l_836);lane 7,51(562_836);lane8, 51(515-XI9); lane 9, 51(5,5-813); lane 107 51(515-747)-The sizes (kilodal-tons) andpositionsof themolecularweightmarkersare indicatedontheleft.
protein-DNAcomplex. Theanalysiswasperformed according
tothe manufacturer'sinstructions, with arecordingtime of40 min for eachgel.
RESULTS
In vitro transcription and translation of VZV gene 51. A 2.7-kb fragment from the VZV genomecontaining the entire gene51 codingsequencewassubcloned into avectorsuitable for in vitro transcription and translation as described in Materials and Methods section. The
L-[35S]methionine-labeled
translation product of gene 51 was analyzed by SDS-PAGE and immunoprecipitation followed by SDS-PAGE. The mostslowlymigratingproteinband(Fig. 3, lane 1) correspondsto a
molecularweight of 85,000. This invitro synthesized protein comigrated with gene 51 protein expressed in insect cells infected witharecombinant baculovirus(datanotshown).The predicted molecular size ofgene51proteinis94,314,however, and the possibility that the 85-kDa polypeptide was derived from proteolyticdegradation orinternal translation initiation wasconsidered. The
35S-labeled
translation productwas then immunoprecipitated with a rabbit antiserum (R309) which recognizes the carboxyl-terminal 20 amino acids of gene 51 protein. The 85-kDaproteinwasimmunoprecipitatedbyR309 butnotbyapreimmune antiserum from the samerabbit(Fig.3, lanes 2 and3). R309 antiserum also immunoprecipitated a 322-amino-acid polypeptide from the carboxyl terminus
[51(515-836); Fig.2and Fig. 3, lane5] butnotapeptide witha
deleted carboxyl terminus
[51(515.819));
Fig. 2 and data notshown], showing the specificity ofthe R309 antisera for the carboxylterminus.Amino-terminalamino acid sequence anal-ysiswasperformedonthe85-kDa gene 51proteinexpressedin recombinant baculovirus-infected insect cells. Nine of ten
residues from the amino terminuswere determined
unambig-uously and were identical to the predicted amino acid se-quenceencodedbygene51 (5).The 85-kDaproteintherefore represents the full-length product ofgene51.
DNA-binding activity of in vitro-synthesized gene 51 pro-tein.The 85-kDa gene 51 product obtained from extractsof insectcellsinfectedwith arecombinantbaculoviruswas insol-J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.94.265.454.617.2]VZV ORIGIN-BINDING PROTEIN 3845
C
1
2
3
-C3
7
_.
..-
..-1 2 3 e
.. .
1
2
3
4
5
6
&-
-C2
:s aS
-CI
-P
...
_E.
1.~~~~~~~~~..
I
ubleandthereforenotsuitableforfunctionalstudies. Thegene
51 protein synthesized in vitro was tested, therefore, for
site-specific DNA-binding activity inagel retardationassay.A
5'32P-end-labeled 26-merprobe(VOA-26)wasusedin the gel
retardationassay(see MaterialsandMethods). Figure 4shows
that the rabbit reticulocyte extract containing in vitro-trans-lated gene 51 protein (lane 4) caused a retardation of the
probe whereasthe controlrabbitreticulocyte lysate displayed
noretarded bands(lane 1).Theretardedcomplex(es) didnot yield adiscrete band, butaddition of anti-51 antisera (R309) ledtoadoubleretardation ofall thecomplex(es) (lanes7and
8).
Determination of the boundaries of the site-specific DNA-binding domain of the VZV OBP. The site-specific DNA-binding domain of the gene 51 product was mapped to the carboxyl-terminal 322 amino acids with a protein A fusion
protein (23).Toconfirmthis inthe invitro translationsystem andto determinewhether theDNA-bindingdomaincould be
-C2
-.c1
.
WV,-
_
_s_
-PFIG. 6. Gel retardationassays using in vitro synthesized products
ofgene51 deletion mutants. Theprobe (VOA-26)wasthesameas
used inFig. 3. (A) Lane 1, control lysate; lane 2,51(515_836)product; lane3,51(515-747)product; lane 4,51(515-813)product; lane5,51(515819)
product; lane 6,51(562836)product; lane 7, 51(551.836) product; lane 8,
51(534836) product. P, free probe; C, probe-protein complex. (B) Doubleretardationusing51(515_836)mutant product. Lane 1,control lysate; lane 2, control lysatewith 0.2 ofpre-R309; lane 3, control lysate with 0.2 p.1ofR309; lane 4,lysatewith
51(515836)
product; lane5,
51(515836)
product with 0.2 p.l ofpre-R309;lane6,51(5
5-836) prod-uct with 0.2 p.1 of R309. P,free probe; Cl, probe-protein complex;C2, protein-antibody probe complex. (C) Double retardation with
51(173-836) mutant product. Lane 1, 51(173-836) product; lane 2, 51
(173-836) product with 0.2 of control serum (pre-R309); lane 3,
51(173.36) product with 0.2 ofanti-51 antiserum (R309). P, free
probe; Cl and C2, protein-probe complexes; C3, protein-antibody-probe complex.
expressedas astablepolypeptide, aseries ofdeletionmutants of the gene51 wereconstructed. Figure 2 shows aschematic mapof thesemutants andtheirrelationship tothefull-length OBP. Figure 5 shows an autoradiograph of the
L-[35S]methi-onine-labeled in vitro translation products of each deletion mutant analyzed by SDS-PAGE. Although multiple radiola-beled productswere seen witheach mutant, themost slowly migrating band corresponds to the predicted size of each
mutantpolypeptide.
To determine theboundary of the DNA-binding domain of theVZVOBP, eachmutantproductwastested forsite-specific
DNA-binding activity. Figure 6 shows an autoradiograph of
the gel retardation results which are summarized in Fig. 2.
Thesmallerpolypeptidesgaveasingleretardedcomplex (Fig.
6A, lanes 2, 4, 5, 7, and 8,and Fig. 6B, lanes 4 and5). The larger polypeptides [e.g.,
51(353836)]
did not yield a definiteprotein-DNA complex (data notshown; indicatedby a
ques-tion mark inFig.2)oryieldedseveral indistinct retardedbands
[51(173-836);Fig. 6C, lane 1]. To demonstrate thatthese
indis-tinctly retarded complexes contained truncated gene 51
A
B
VOL.68, 1994
10
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.366.519.74.341.2]3846 CHEN AND OLIVO
Fold excess competitor
10
5'-CCACCGTTCGCACTTTCTTT-3'
----A---
---T--
-
---T---
---A---C
---I
I
100 50 100 50
%ControlBinding %ControlBinding
100 50
%ControlBinding FIG. 7. Mutational analysis of the VZV OBP recognitionsequence.Bindingassays wereperformed with the51(515-836) product and analyzed byacompetitive gel retardationassay asdescribed in Materials and Methods. The radiolabeled probewasVOA-26.Sequence of the pointmutants
isshownontheleft. Thetop line indicatesthesequenceof the wild-type site A 20-mer oligonucleotide (VOA)withthe10-bp UL9 recognition
sequence inboldface type. Belowarethesequencesofa series ofpointmutants. Only the mutated base is indicated. Graphic displayof the competitive bindingresults with eachVOApointmutantsisshownontheright.Horizontal bars indicatebindinginthepresenceof the indicated
amountsofexcesscompetitoroligonucleotide expressedasapercentageof binding withnocompetitor (control binding).
polypeptideswith anintactcarboxyl terminus,a
double-retar-dationassay wasperformed by usingtheanti-carboxyl-terminal
antisera (R309). A double retardation assay with truncated polypeptides
51(515_836)
(Fig. 6B, lane 6) and51(173-836) (Fig. 6C, lane 3) showed that all the complexes appear to be double-retarded by R309 antiserum but not by preimmune antiserum (Fig. 6B, lane 5; Fig. 6C, lane 2). The amino terminus of the DNA-binding domain ofgene 51 protein islocated between amino acids 551 and 562, and the carboxyl terminus is located between amino acids 747 and 813(Fig. 2). The smallestpolypeptide testedwhich retainedDNA-binding activity [51(551_836); Fig. 2] was 286 amino acids, but the
DNA-binding domainmaybe assmall as263 aminoacids.
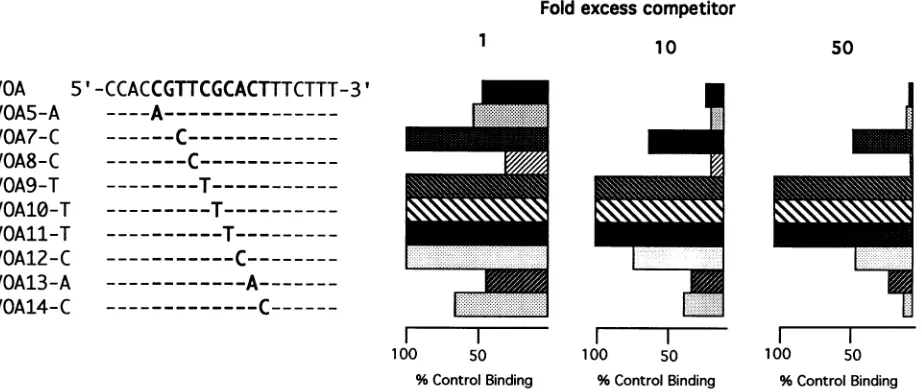
Effect ofsingle-base-pairmutations in the VZVOBP
recog-nition sequence. Elias et al. identified an 11-bp sequence,
5'-CGTTCGCACTT-3' that isrequired forhigh-affinity bind-ing of UL9, and Hazuda et al. refined the UL9 recognition
sequence to 10 bp (CGTTCGCACT) (7, 11). In the VZV
originthis10bpsequenceis foundwithin site A and siteB,and
site C contains 9of the 10 bp. Stowet al. showed that these bases arewellprotected in DNase Ifootprinting experiments using a fusion protein containing 322 amino acids from the
carboxyl terminus of the OBP and a DNA fragment that
contained site Aand site C(23).Inaddition,deletionofthese 10 bases eliminatedbinding. Hazuda etal. performed satura-tion mutagenesis on the 10-bp UL9 recognition sequence to
assess the relative contribution of each base pair. These
authors showed that any point mutation within the 10-bp
sequence reduced binding by 2- to 10-fold, but that any
substitution at positions 5, 6, and 7 (CGC) reduced UL9 binding bymore than 100-fold (11).
In the present work, nine 20-mer oligonucleotides each containing a single point mutation in this 10-base sequence
within thecontextofthe siteAsequenceweremadeand tested
for gene51 protein binding in a competitive gel retardation
assay. In vitro synthesized
51(515_836)
product was incubatedwith radiolabeled VOA-26 in the presence or absence of an
excessofmutantsite Aoligonucleotide,andtheprotein-DNA
complexeswereanalyzed on a nondenaturingacrylamide gel.
Figure7 shows thepositions ofthe mutationsandahistogram
of a quantitation of the results derived from submitting the
driedgeltoaBetascope analysis (seeMaterials andMethods).
Thewild-type site A 20-mer givesthe predicted reduction in binding(Fig. 7,topline).Whereassomeof thepointmutations
havelittleornoeffectonbinding (Fig. 7, VOA5-A, VOA8-C,
VOA13-A, and VOA14-C), others cause at least 10-fold reduced binding (Fig. 7, VOA7-C, and V012-C), but,like the UL9results of Hazudaetal.,substitutionsatbases5, 6, and 7 (CGC)result inover50-fold reducedbinding(Fig. 7, VOA9-T,
VOA10-T, andVOA11-T).
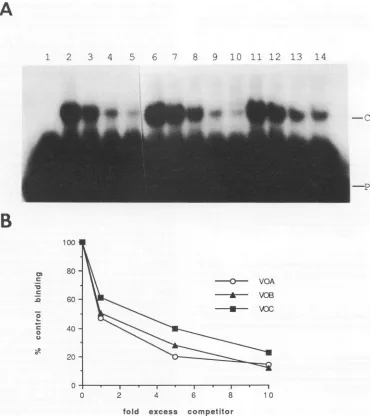
Comparisonof therelativebinding affinityof the VZV OBP toeachof the threebindingsites in the VZVorigin. Figure 1 showsthepositionandsequenceof the three sitesonthe VZV
origintowhichaprotein A-gene51 fusionproteinwasshown
tobind (23).The relative binding affinityofthe VZV OBPto these three sites was analyzed by using a competitive gel
retardation assay. In vitrosynthesized
51(5,5_836)
productwasused for this analysis. Three 20-mer oligonucleotides, which correspond tosite A (VOA), B (VOB), and C(VOC), were
used to compete for binding with a radiolabeled 26-mer
oligonucleotide correspondingtothe DNAsequenceof site A
(VOA-26). Figure 1D shows the DNA sequence of these oligonucleotides. Figure 8A shows the autoradiographic
re-sults, and Fig. 8B shows a graphic display of a quantitative
analysisof the results derived from submitting the driedgelto
a BetaScope. The VZV OBP binds to sites A and B with virtuallyidenticalaffinities, butitbindstositeC withatwo-to threefold loweraffinity.Thissameresultwasobtainedonthree separateexperimentsand by labelingasiteColigonucleotide
(VOC) andcompetingwith VOA(datanotshown). Sequence comparison of the three sites with respect to the 10-base recognition sequence shows that, whereas sites A and B are
identical,the second basepairof the site C10-mer recognition
sequenceis A instead ofG(Fig. 1D).A20-bp oligonucleotide 1
VOA
VOA5-A
VOA7-C
VOA8-C
VOA9-T
VOA10-1
VOAll-1
VOA12-(
VOA13-1
VOA14-(
r
50
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.79.540.76.272.2]A
1 2 3 4 5 6 7 8 1 10 11 12 13 14
-C
-P
B
100
80
C
._
c
'a
a c
0
U
40
20
0
-0-- VOA
-l-- VOB
-n-* Voc
4 6 8 10
[image:7.612.132.503.76.493.2]fold excess competitor
FIG. 8. Comparisonof thebinding affinityof the VZV OBP to the threebindingsites in the VZVorigin.Competitive gel retardation assays wereperformed as described in Materials and Methods. Binding assays were performed with the51(515_836)in vitrotranslation product. VOA, VOB, and VOC 20-mer probes were used as competitor oligonucleotides (Fig. 1D). (A) Autoradiograph. Lane 1, control lysate; lanes 2 and 6,
51(515836)
product without competitor; lane 3, addition of an equimolar amount of unlabeled VOA; lane 4, fivefold excess VOA; lane5, 10-fold excessVOA;lane 7,equimolarunlabeledVOB; lane8, 5-foldexcessVOB; lane9, 10-foldexcessVOB;lane 10, 20-fold excess VOB; lane 11, equimolarVOC; lane12,5-foldexcessVOC; lane 13, 10-fold excessVOC; lane 14, 20-fold excess VOC. P, freeprobe;C, probe-proteincomplex. (B) Graphicdisplay of the competitive gel retardation data derived from the results shown in panel A. The amount ofbinding in the presence of the indicated amounts of excesscompetitoroligonucleotide dividedby the amountofbindingwithnocompetitortimes 100(% controlbinding) isplottedagainst the molar ratio of unlabeled competitor VOA, VOB, and VOC to radiolabeled probe(foldexcesscompetitor).correspondingtosite C except forasingleA toG mutation was made, andby usingacompetitive binding assay, it was found that thebinding affinitywasidenticaltosite A andB (datanot shown). This result is consistent with theidea that the small difference in binding affinity to site C is due to the G to A difference in the 10-bp recognition sequence rather than differences in the flanking sequences.
DISCUSSION
Inthisreport,theexpression and functional identification of the product of VZV gene 51 are described. The full-length gene 51 protein, expressed in a rabbit reticulocyte lysate in
vitrotranscription-translation system, has a molecularweight of 85,000 as determined by SDS-PAGE, despite apredicted
molecularweightof94,314.This is similartoUL9,which also exhibitsanunexplained,faster-than-predictedmobilityin SDS-PAGE. Gel retardation results indicated that the full-length
gene 51 proteinbinds to specificsequences found within the VZVoriginofreplication.However, likeUL9,full-lengthgene 51 proteindoes notyieldadiscrete protein-DNA complexin nativepolyacrylamidegels whereascarboxyl-terminal
polypep-tides ofbothOBPs doyielddiscrete bands inthis assay.Invitro translation products of a series of deletion mutants were
analyzedforsite-specificDNA-bindingactivity,and theresults wereusedtolocalize theorigin-bindingdomaintobetween263
on November 9, 2019 by guest
http://jvi.asm.org/
3848 CHEN AND OLIVO
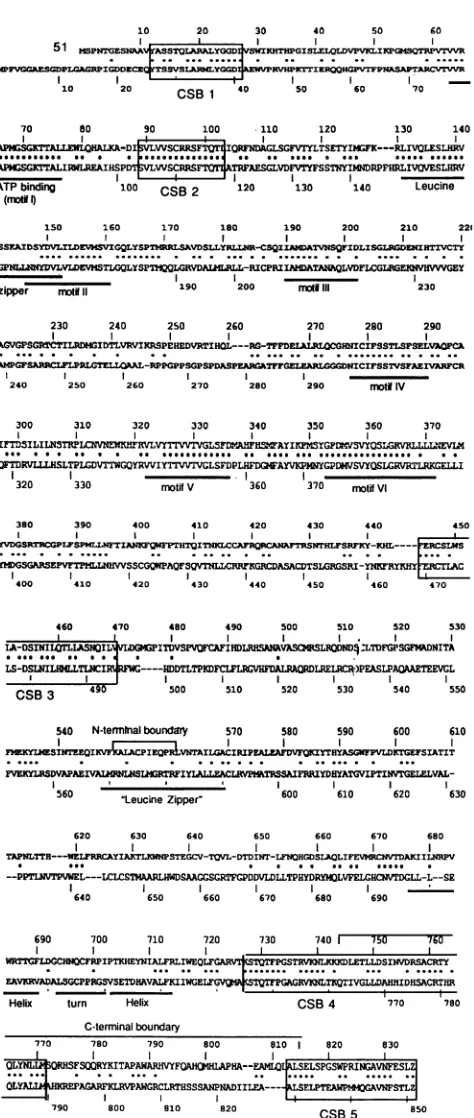
10 20 30
31 M,PTENA SQLBLGGD'VKHTHPG1' bEvGCGAESCDPL PIG0DD=ICK YTS0VSLLAr34YGGIAZVPRVHPRT
10 20 CSB1 40 50
70 80 90 100 110
I~~
APG TrALLELHALKA-DISVLWASCGRSFTQT1jIQRFI7DAISG8Vr AP8ESGXTTALIRWLREAIHSPD7 VIVSCRRS0 AT8FAESGLV0FVT
ATPbinding 100 CSB2 120 131
(mot6
150 160 170 180 190 SSZAXDSYDVLI0E4SJVXGQLYSPTPRLSAVDSLLYRLLNR-CSQIAMI
.... .... ... ... .... . ...
GPNLLN0NYDVLVDEVMSTLCQLYSPFTQQILGRVDALIMl8LL-RICPRIIAt4A0
__ _ __ _ _
zipper motiD1119 200 no
230 240 250 260
AGVGPOGRTCTLRDmGIDTLVRV!tOtOPEHEDVRTTHOL---RO-TOTFDELAI A PGFSAPRCI.FP9RLGTELLRPAL-RPPGPPSGPSPDASPEARG6TFF0ELFJ
240 250 260 270 280 2
300 310 320 330 340
IFTDSILILNSTPIE3E0NrVymTrwLVTT5FD0GLSAHFITssAYIz
0FTDRVLLLSLTPIDVTrTGQYRVWQIY7ITTVLSFDPLHFDGMFAYVKPI
320 I3
320 330 motifV 360
380 390 400 410 420 4 YVDGSRTcGPLFSPMLG.TIANFOqWFPTHTQITN0.CCAFTIHRCANAFTR4
YMDGSGWZ0PVFTP0LLNWVVSSCCQPAI0Qr2QVTNLLCaRoGRCBPZACD7
400 410 420 430 440 4
460 470 480 490 500
La-DSINTTLLMLLW ITDVSPVOFCA?IHDLRDSANAVASCM
LS-DSLNILHLLTLCIlIC----HDDTLTPKDFCO.FLRCVHFDALRtAOO
CSB 3 490 500 510 520
540 N-termlnalbounday 570 580
J38EKYLSI4ES.WrIZQIKV~P~IA'PI4TA.L0A.I8ZPELuEALE0PVFQ VWKYI.RSDVAPAEIVAIWILNSLS8GRTrFIYLALLEACLRVP9TESSAIFPI
560 "Leucine Zipper" 600
620 630 640 650
I TAPILTTH---WELFR.r-AYIAXTLGCNSTE.cV-TQvI-DTDINT-INOHGI
--PPTLNVTPVW*RL---LCLCST!4APLHWDSAAG RTFCPDDVLDLTPHYI
II I
640 650 660 670
690 700 710 720 730 WRTTGFLDGCHNOCFRPIPTKHEYNIALFRLIWEQLFGAOTVTLSTVFPGS EAVI!RVA)ALSrCPPflGSVSETDHAVALFKIIWGELFVP42TTPG&GRIV
Helix turn Helix Cc C-terminalboundary
770 780 790
_
800 810
UYNLIPV;QRHSFSOQRYKITAPAWARHVYFOAIO4IIIAPAA--EAMLO0IdS-E
QLYALISEFAGARFKLRVPAWCflLRT0
[image:8.612.56.293.66.625.2]SSANPNADILEA-790 800 810 820
FIG. 9. Alignment of the predicted aminc VZV gene 51 protein and HSV UL9. The to acidsequence of the gene 51 protein, and the UL9sequence. Barsbelow the UL9 sequence sequence motifspreviously noted in UL9 (6, 1 V,and VIrepresent helicasesuperfamily mot C-terminalboundaries indicated above the ge represent the boundaries of the site-specific Theoutermostpoint of each boundary retaine and theinnermost point lackedDNAbinding.
40 50 60 and 286 amino acids at the carboxyl terminus. Finally, an
S LwwKPvSoTr'VV analysis of the VZV OBP recognition sequence showed that
. . .. ...
TIERQOHGPVTFPNeAr
TIARCVVR
the three previously identified binding sites in the VZV origin, '60 70 which share a 5'-CRTTCGCACT-3' sequence, bind with sim-ilar affinities to the OBP and, as with the HSV OBP (UL9),a 120 130 140 CGC triplet is critical for binding. The HSV and VZVOBPs,ILTSETYIHQFK---RLIVQLISLHRV
therefore,
appear to have virtually identical recognitionse-YFSs;nwmOoRPMauVovEsIDmv
quences despite significant differences in their primarystruc-O 140 Leucine tures (see below).
The HSV and VZV OBPs have a 44% identity and 53% 200 210 221 similarity in their predicted primary sequences. An alignment
ATV%SQFIDLXSGLPGDEKNZTZVCTE of their primary sequences is shown in Fig. 9, which also
ATARQLVDFICGLRGE0iVHVVVGEY
indicates the location of sequence motifs which have been230 noted in UL9. Most of the homology between UL9 and the VZV OBP occurs within these motifs. In addition, there are 270 280 290 five regions ofhigh homology between UL9 and the gene 51
Ud=HNICIF
S%PSEL%AOPA protein which do not occur within known sequence motifs____G__ICIFSSTVSFAEIV_WCR (conserved sequence blocks [CSB] 1 to 5 in Fig. 9). The 290
Mot#
IV amino-terminal two-thirds of the gene 51 protein contains all six of the helicase superfamily motifs which have been noted to 350 360 370 be present in UL9 (13) (Fig. 9). Single-amino-acidsubstitu-MSYGFDPTVSVYQ5LGKVLLL1NEVU4 tions in conserved residues of several of these helicase motifs
a... ..0
OIYGPDWSWYOSLGVRTLRKCE1LI
within UL9 inactivated the function of UL9 in transient 170motif
VI origin-dependent replication assays (13). The presence of thesemotifs in thegene51proteinsuggeststhat, like UL9, the430 440 450 gene 51 protein has DNA-dependent ATPase and helicase
IS.-*ffXS"U
l---- mRcssactivity.
A leucinezipper
noted near the amino terminus ofrSLG0laRI-YNArRYI
Y UL9 was shownby
insertionmutagenesis
to berequired
for450
1460
470 cooperative binding ofUL9to the origin (8, 11). The leucine zipper motif in the amino-terminal portion of the gene 51 510 520 530 protein has a64%
homology
with thecorresponding
UL9BSLI
DNLFGI
IT motif and differsprimarily
at two leucine residues which have been replaced by isoleucine (Fig. 9).530 I540 The
sequence-specific DNA-binding activity
of UL9 waslocalized to amino acids 564 to 832 (6). A pseudo-leucine
590 600 610
zipper
has been noted within thisdomain,
and insertionalmutagenesis
has shown that it isimportant
for DNAbinding
PIYTHYASPVLDTGEFSTATIT
(1, 6).
A helix-turn-helix motifwas also identified near the
8IYD'tDAT;WPTINVTGELELVAL-
carboxyl terminus, but its relevance to the DNA binding610 620 630 activity of UL9 remains unknown (6). The presence of these two motifs in the predicted primary sequence of gene 51 660 670 680 protein, however, is not apparent. The boundaries of the
dmi h
DSLQLIFEVM AKIILNIPV origin-binding domain of the gene 51
protein
arevery similarMDRYOLVFELGHQcTDGLL-L--SE
to those determined for UL9, but the homology of the'680
1690
origin-binding domains is actually less (33% identityand44%similarity) than the overall homology of the two molecules. 740 -750 760 The sequence requirements for binding of both OBPs, how-0IK0LET
DSINVIRSARTY
ever, are virtually identical, and each OBP binds to bothRMILTOTIV0AHOIDH
SACRTlIR
origins
(23
andunpublished observation).
There are two SB 4 770 780 regions of high homology in the origin-binding domain. The region of homology referred to here as CSB 4 was recently shown to be important for, but not sufficient for, UL9 DNA 820 830 binding (12).Whether theseconserved amino acids are directly:LSPGSWPRINAVNESLZ
'LSPG..
...AVNFE.. or indirectly involved in sequence recognition remains to be:LPTEAWPNI4AVNFSTLZI
determined. CSB 5atthecarboxyl terminus isnotrequired
for CSB 5 850 DNA binding of either protein (Fig. 2) (1, 6). This suggests that the origin-binding domain is involved in an as-yet-unde-acid sequences of the terminedfunction common to bothOBPs.
p line shows the amino HSV
oris
andVZVo,is
have very similarOBP
recognition bottom line shows thesequences,
but an apparent difference in the arrangement ofindicate the positions of the binding sites relative to their AT-rich regions (Fig. 1).
13AMAtifkI IT ITT TV t
10).MlVltJl; 1, 11, 111,IV,
tifs.The N-terminal and ne51 proteinsequence
DNA-binding domain. dDNA-binding activity,
Theboxes marked CSB
1 through CSB 5 indicate conserved sequence blocks which are arbitrarily marked regions of high(.60%) homology.
tSAE
J.VIROL.
c .4
*I@-__USE
.
on November 9, 2019 by guest
http://jvi.asm.org/
WhereasHSV
ons
has ahigh-affinitybindingsiteoneachside of the AT-rich region, the VZV origin contains three origin-binding sites which are all on the same side of the AT-rich region.The VZV OBP was shown to bind with equal affinity to site Aand siteBandwithaslightlyloweraffinitytositeC.The similar bindingaffinity of all three sites is interesting given that plasmid replication assays indicated that site A alone is suffi-cient for plasmid replication in VZV-infected cells (23). This structural differencebetween the HSV and VZV origins is also interesting in light of the observation that plasmids containing the VZVorigin are replicated in HSV-infected cells (22). This suggests that the apparent difference in arrangement of the binding sites is not critical to the formation of a functional initiation complex. Alternatively, it is possible that there areas-yet-undetected binding sites to the right of the AT-rich region inVZV
oris.
By using an oligonucleotide probederived from sequences to the right of the AT-rich region, which contain 7 of the10 bp of the recognition sequence (see dashed line, Fig. 1C), no binding was observed (data not shown). A very low affinity binding site, however, could be missed in binding assays using DNA probes which contain a single binding site or in DNase I footprint assays using gene 51 protein lackingthe amino terminus (23). The amino terminus of UL9 has beenshown to be required for cooperative binding, and binding to a low-affinity site (siteIII) has been observedonly in DNase I footprints of full-length UL9 on DNA fragments which also contain siteI (7, 8, 10). Identification of low-affinity gene 51 protein binding sites on VZV
ons
may, therefore, depend upon binding studies with full-length gene 51 protein.ACKNOWLEDGMENTS
We thank L. Gelb forproviding the VZVBamHI D fragment, B. Moss for providing plasmid pTM1, and M. Horgan for providing plasmid pMH4. We also thank Erik Stabell, David Leib, and Skip Virginforhelpfulcomments onthemanuscript.
REFERENCES
1. Arbuckle, M. I., and N. D. Stow.1993. A mutational analysis of the DNA-binding domain of the herpes simplex virus type 1 UL9 protein.J. Gen.Virol. 74:1349-1355.
2. Challberg, M. D. 1986. Amethod for identifying the viral genes required for DNA herpes replication. Proc. Natl. Acad. Sci. USA 83:9094-9098.
3. Challberg, M. D., and T. J. Kelly. 1989. Animal virus DNA replication. Annu. Rev. Biochem. 58:671-717.
4. Davison, A. J., and J.E.Scott. 1985. DNA sequence of the major invertedrepeat in thevaricella zoster virus genome. J. Gen. Virol. 66:207-220.
5. Davison,A. J., and J. E.Scott.1986.The complete DNA sequence ofvaricella zoster virus. J. Gen. Virol. 67:1759-1816.
6. Deb, S., and S. P.Deb. 1991. A269-amino-acid segment with a pseudo-leucine zipper and ahelix-turn-helix motif codes for the sequence-specific DNA-binding domain of herpes simplex virus type 1origin-bindingprotein. J. Virol. 65:2829-2838.
7. Elias, P., C. M.Gustafsson,and0.Hammarsten. 1990. The origin binding-protein ofherpes-simplex virus-I binds cooperatively to
the viral origin of replication oriS. J. Biol. Chem. 265:17167. 8. Elias, P., C. M. Gustafsson, 0. Hammarsten, and N. D. Stow.
1992. Structural elements required for the cooperative binding of the herpes simplex virus origin binding protein to oriS resideinthe N-terminal part of the protein. J. Biol. Chem. 267:17424-17429. 9. Elias, P., M. E. O'Donnell, E. S. Mocarski, and
I.
R.Lehman.
1986. A DNA binding protein specific for an origin of replication of herpes simplex virus type 1. Proc.
Natl.
Acad. Sci. USA 83:6322-6326.10. Hazuda, D. F., H. C. Perry, and W. L. McClements. 1992. Cooperative interactions between replication origin-bound mole-cules of herpes simplexvirus origin-binding proteinare mediated via the amino terminus of the protein. J. Biol. Chem.
267:14309-14315.
11. Hazuda, D. J., H. C.Perry,A. M. Naylor, and W.
L.
McClements. 1991. Characterization of the herpes simplex virus origin binding protein interaction withoris.J. Biol. Chem. 266:24621-24626. 12. Martin, D. W., R. M. Munoz, D. Oliver, M. A. Subler, and S. Deb.1994. Analysis of the DNA-binding domain of the HSV-1 origin-binding protein. Virology 198:71-80.
13. Martinez, R., L. Shao, and S. K. Weller. 1992. The conserved helicase motifs of the herpes simplex virus type 1 origin-binding protein UL9 are important for function. J. Virol. 66:6735-6746. 14. Matthews, R. E. F. 1982. Classification and nomenclature of
viruses. Fourth report of the International Committee on Taxon-omyofViruses. Intervirology 17:1-199.
15. McGeoch, D. J., M. A. Dalrymple, A. Dolan, D. McNab, L. J.
Perry,P. Taylor, and M. D. Challberg. 1988. Structures of herpes simplex virus type 1 genesrequired for replication ofvirus DNA. J. Virol.62:444-453.
16. Moss, B. 1990. New mammalian expression vectors. Nature (Lon-don) 348:91-92.
17. Olivo, P. D., N. J. Nelson, and M. D. Challberg. 1988. Herpes simplex virus DNA replication: the UL9 gene encodes an origin binding protein. Proc. Natl. Acad. Sci. USA 85:5414-5418. 18. Olivo, P. D., N. J. Nelson, and M. D.
Challberg.
1989. Herpessimplex virus type 1 gene products required for DNA replication: identification and overexpression. J. Virol. 63:196-204.
19. Spaete, R. R., and N. Frenkel. 1982. The herpes simplex virus amplicon: a new eukaryotic defective-virus cloning-amplifying vector. Cell 30:295-304.
20. Stillman, B. 1989. Initiation of eukaryotic DNA replication in vitro. Annu. Rev. Cell Biol. 5:197-245.
21. Stow, N. D. 1982. Localization of an origin of DNA replication within the TRs/IRs repeated region of the herpes simplex virus type 1 genome. EMBO J. 1:863-867.
22. Stow, N. D., and A. J. Davison. 1986. Identification of a varicella zoster virus origin of DNA replication and its activation by herpes simplex type 1 gene products. J. Gen. Virol. 67:1613-1623. 23. Stow, N. D., H. M. Weir, and E. C. Stow. 1990. Analysis of the
binding sites for the varicella-zoster virus gene 51 product within the viral origin of DNA replication. Virology177:570-577. 24. Vlasny,D.A.,and N. Frenkel. 1982. Replication of herpes simplex
virus DNA: localization of replication recognition signals within defective virus genomes. Proc. Natl. Acad. Sci. USA 78:742-746. 25. Weller, S. K., A. Spadaro, J. E. Schafer, A. W. Murray, A. M. Maxam, and P. A. Schaffer. 1985. Cloning, sequencing, and functional analysisofo"L,a herpes simplex virus type 1originof DNA synthesis. Mol. Cell. Biol.5:930-942.
26. Wu, C. A., N. J. Nelson, D. J. McGeoch, and M. D.
Challberg.
1988. Identification of herpes simplex virus type 1 genes required for origin-dependent DNA synthesis. J.Virol.62:435-443.
![FIG. 5.beledproduct;51(515-XI9);lane(2tons)the [l) SDS-polyacrylamide gel analysis of L-[35S]methionine-la- products of gene 51 deletion mutants synthesized in vitro](https://thumb-us.123doks.com/thumbv2/123dok_us/1293389.82290/4.612.94.265.454.617/beledproduct-polyacrylamide-analysis-methionine-products-deletion-mutants-synthesized.webp)