0022-538X/84/030997-05$02.00/0
Copyright © 1984,AmericanSociety forMicrobiology
Large-T-Antigen-p53 Complex Formation
Is Not
Cold Sensitive in
a
Cold-Sensitive Transformant Induced by Simian Virus 40
Mutant
tsA1499
NOEL BOUCK,* JOHN FIKES,AND M. KATHLEENRUNDELL
Department ofMicrobiologylImmunology and Cancer Center, Northwestern University Medical and DentalSchools,
Chicago, Illinois 60611
Received24August 1983/Accepted9November1983
Flll rat cells transformed by simian virus 40 mutant tsA1499 are cold sensitive for the
expression
oftransformation. Yet, unlike Flll cells transformed by
tsA58, they
do not lose theability
to stabilize thetransformation-associated hostcell
protein
p53atthe temperatureatwhichtransformation isextinguished.Although it has been clear for a number of years that the
maintenance of the neoplastic phenotype in a cell
trans-formedby simian virus40 (SV40) depends on the continual
functioning of the virus-encoded protein known as large T
antigen (2, 21, 30, 38, 46, 51), exactly how T antigen
performs this feat remainsobscure.
LargeTantigen isa708-amino-acidprotein which is found
tobeassociatedinhighconcentrations withvarious nuclear
fractions of the cell (45) and in low concentrations with the
plasma membrane(20).Multiple,genetically separable func-tions have been assigned to large T antigen (50), many of
which could conceivably be crucial in keeping a host cell
transformed. Some of these functions, however, can be
active in virusesthatareunable to maintain transformation
and, therefore, appear unlikely to be sufficient for the
maintenanceofmalignancy.Theseinclude the ability to bind to the SV40 origin of DNA replication (7), to initiate viral DNA synthesis (10), to regulate viral transcription (10), to conferimmortality (9), and tostimulate rRNAtranscription and DNAsynthesis in thehost cell genome (44).Some
large-T-antigen functionscanbe lostfromavirus without impair-ingitsabilitytotransformandtherefore appear unnecessary
formaintaining transformation. These functions includethe
ability to bind to the origin (49), to initiate viral DNA
synthesis (31, 48), and to help with adenovirus replication (1). Oftheknownfunctions oflarge Tantigen, only two have not been convincingly dissociatedfrom the maintenance of
transformation: an ATPase activitywhich copurifies with T
antigen (6, 15, 49) and the ability of T antigen to from a
complexwithand stabilizeahost cellprotein with a molecu-larweightof about 53,000 (p53)(24).
The most attractive current hypothesis suggesting the mechanismby whichTantigen might act to maintain
trans-formation focusesontheability of T antigen to complex with
p53. p53 is a highly conserved (23), very short-lived (36)
phosphoprotein with an associated protein kinase activity
(22) thatappears toplay a regulatory role in normal cellular growth control, as microinjected antibodies againstp53 can haltseruminductionof normal cellular DNA synthesis (33).
p53
levels are usually low in normal cells. In cellstrans-formedby SV40, virtuallyallcellularp53is complexed with aportion of T antigen (32), and the level and stability of the
p53 protein are increased manyfold (36, 37). It has been
* Correspondingauthor.
proposed that these high levels of p53 complexed with T
antigen act in a positive way to maintain the unregulated growthof theSV40-transformed cell(25).
This suggestion is enhanced by the finding that early
transformation-associated antigensoftwoother DNA virus-es,adenovirusandEpstein-Barr virus, also formcomplexes with andincrease the levels of p53 (28, 42). Inaddition, p53 levels are elevated in a wide array of variously induced
primarytumorsand tumorcelllines, especially inmice (13,
26,40, 41),butalso in humantumorcells(12, 14).However, thereare clearexceptions to these correlations ofhigh p53
levels and transformation. Some tumorcells of human (12) andmouseorigin (36) andsome primary mousetumors(40) do not showhigh p53levels. Inlymphocytes (34, 35), some cultured cells (14), and early embryos (5), high levels are seen in the absence oftransformation. As hasbeentrue of some earlier correlatesoftransformation, elevated levels of p53 could beafrequent consequence of theexpressionof the
rapid-growth phenotype ofthetransformed cell ratherthan
thefundamentalcause oftumorigenic growth deregulation.
The experiments reported here test whether the expres-sion ofthe transformed phenotype always correlates with thestabilization ofp53inanSV40-transformed cell line. The
results indicate that transformation can be lost without
significantreduction in the p53-T-antigen complexorinp53
stability andthat thep53 stabilizingfunction of Tantigenis notsufficientto maintainthe transformed state.
Amatched setof cell lines (38) wereusedwhich wereall selected and recloned in agar after virus infection of one parental rat fibroblast line, Flll (for culture methods, see reference 38). The lines were derived from infection with different T-antigen mutants and varied in the effect of temperature on their ability to express transformation. Transformationhas beenmeasuredasthe abilitytoclone in
softagar, anassay whichhas been showntobea strict and
appropriate in vitro correlate of neoplasia in
SV40-trans-formedratcells (50).
Table 1 illustrates the phenotypes of the cell lines as measured inafractionfrom each of the cultures used for the
pulse-chaseexperiments reportedbelow. Normal Flll does notclone in agar, whereas itswild-type-transformed
deriva-tive, F(wt)40,clonesefficientlywhether grown and testedat the high (39.3°C) or the low (31.5°C) temperature. Flll transformedby mutant tsA58 expressestransformation
effi-ciently onlyatthelow temperature,asis characteristic of the
majority of cells from various species transformed by this 997
on November 10, 2019 by guest
http://jvi.asm.org/
998 NOTES
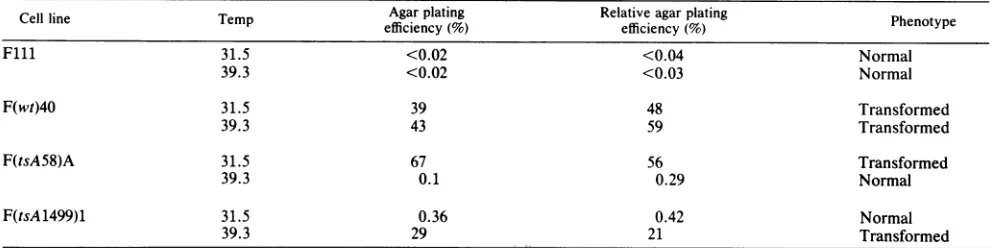
TABLE 1. Effect of temperature on expression of transformation
Cellline Temp Agar plating Relative agar plating Phenotype
efficiency(%) efficiency(%)
Flll 31.5 <0.02 <0.04 Normal
39.3 <0.02 <0.03 Normal
F(wt)40 31.5 39 48 Transformed
39.3 43 59 Transformed
F(tsA58)A 31.5 67 56 Transformed
39.3 0.1 0.29 Normal
F(tsA1499)1 31.5 0.36 0.42 Normal
39.3 29 21 Transformed
Fisher rat fibroblasts of cell line Flll and cloned derivatives transformed by wt 45-54, by tsA58 or by tsA1499 were grown at the temperatureindicated for3days and testedatthat
temperatuire
for theabilitytocloneonplasticand toclone in0.3% softagar (38). Relative agarplatingefficiency = (agarplatingefficiency/plastic plating efficiency) x 100.mutant(2, 21, 30, 38).Flll, transformed bymutanttsA1499
(39), expresses transformation efficiently only at the high
temperature. This cold-sensitive phenotype occurs in the
majority ofclonestransformed by the unusual mutant (38),
which contains adeletion at the carboxy-terminal end ofT
antigen, resulting in ashortenedprotein (dlT).
Thestability ofp53 and ofthep53-T-antigen complexwas tested inpulse-chaseexperiments under conditions inwhich transformationwaseitheron oroff(Fig. 1).Brotherdishes of
cells growing at high or lowtemperature were pulsed with
[35S]methionine
and chased for variousperiods oftime. An identical amount ofextractfromeach dishwas immunopre-cipitated withmonoclonalantibodiestoeitherp53
orlargeTantigen, and amountsofp53 and Tantigen were visualized on fluorograms of sodium dodecyl sulfate gels. The
wild-typetransformantbehaved asexpected (32). LargeTantigen
was stable, the
mnajority
of itwas not complexed with p53, and, as reported previously (3, 4), it appeared to complex with p53 slowly as the amount ofpulse-labeled
T antigen precipitated byanti-p53
increased with time,especially
at high temperature. p53 was also stable in thewild-type
transformant and ran on thesegels as a doublet with label appearing first in the
slower-moving
band and thenequili-brating betweenthetwo. Thisapparent increasein
mobility
ofp53has beenseen
by
othersaftershortpulses
(3)and may be due todifferences inphosphorylation
(3) or tomodifica-tions in primary structure (29, 47). In these rat cells, the
faster-moving
band labels moreheavily
with32p
(data not shown), butbothbandscancomplex withTantigen,
asbothareprecipitated by
anti-T-antigen
antibodies.The tsA58-transformed ratcellsbehave in the same man-ner as mouse cellstransformed
by
this same mutant(4, 17, 27)(Fig. 1).
At low temperature, where transformation isexpressed, the cells are
indistinguishable
fromwild-type
transformants, except for the
slightly
increasedlability
of largeTantigen.Athightemperature, where thetransformed phenotypeisextinguished,Tantigenisquitelabile, andp53is extremely unstable. Little
p53-T-antigen
complex is formed in these cellsathightemperature,for,althoughbothp53 and T antigen are clearly present, precipitation of one
protein withthemonoclonalantibodiesspecificfor the other was seen onlyin veryoverexposed gels.
RatcellstransformedbytsA1499arecold sensitiveforthe
expression of transformation to approximately the same
extent that tsA58 transformants areheat sensitive (Table 1). Yet, the amountofp53-T-antigen complexand the stability ofp53 do not appear to change withgrowth temperature, although under the same conditions, a dramatic change is observed in F(tsA58)A (Fig. 1A and B). In F(tsA1499)1 at low temperature, as well as at high temperature, p53 is
clearly complexed with T antigen and is reasonably stable.
Despite the fact that these cells are as isogenic as we can
make them,there are severalconsistentdifferencesbetween thisline and the wild-type and tsA58transformants. (i) The
slower-migrating component ofp53 remainsheavily labeled and does not equilibrate with the faster component in the tsA1499 line. (ii) A larger proportion oftotal T antigen is
complexed with p53 in the tsA1499 transformant. This is
due,in part, to thefactthat thereisless total Tantigen inthis
line than in the other lines. (iii) p53 is less stable in the tsA1499transformants,morenoticeably soafteraveryshort pulse (Fig. 1A)than after alonger pulse (Fig. 1B).However, although these differences may limitsomewhat direct com-parisonsamongdifferentlines,noneof theunique character-istics ofthe tsA1499 line appeartobecausallyrelatedtothe
expression oftransformation,astheyallpersistatbothhigh
and low temperatures, regardless of whether transformation is expressed or extinguished. For example, densitometry tracings ofthegeldepicted in Fig.1B, withanexposurethat
brings the relevant bands well within the experimentally
determined linear range ofgel
sensitivity,
show that28%oftheinitial p53 remained in the tsA1499transformantaftera 22-hchaseat31.5°C and48%remainedafterthe same chase at 39.3°C. In the tsAS8 transformant, 24% of initial p53 remainedafter 22 hat31.5°C, whereas nonewasdetectable at 39.3°C. Thus, in the tsA1499 lineexpressing the normal
phenotypeatlow temperature, p53was asstableasitwasin the tsA58-transformed line, whichwas fully transformed at this temperature.
Todetermine whether some difference could be detected
by sedimentationin thep53-T-antigen complex whichcould account for the shift in transformed phenotype in
F(tsA1499)1, extracts from the three cell lines growing at high and low temperatures were run on 5 to 20% sucrose
gradients, and individual fractions across thegradient
pre-cipitated withanti-T-antigenantibodieswere run onsodium
dodecyl sulfategels (Fig. 2).
Inthewild-type transformant, sedimentation profilesforT J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
A
F II
F(wt)40
p p
1
2
3
r-
____-
-F(ts A58)A
p
1
2
3
- -_
31*50
_ _-_
T-
_- -_
-mF(tsA
1499)l
F(tsA58)A
31.50 39*30 39 30
dl
T-p
53=
p 1 2 p 1 2 p 1 2
FIG. 1. (A) Stability of p53. Cells described in Table 1werelabeled inquadruplicate with [35S]methionine for 50 minat31.50Corfor20min
at39.30C andeither extractedatonce(lanep)orchasedin media containingafivefoldexcessof cold methionine for1h(lane1), 4h (lane 2),or
24 h(lane 3). Extracts made in buffer containing 0.15MNaCl, 1%sodium deoxycholate, 1% Nonidet P-40, 0.1% sodium dodecylsulfate, 10 mMphosphate buffer (pH 7.4), and 1% Trasylolwereimmunoprecipitated with monoclonal antibodiestop53,PAb 122 (18)(uppertworowsof
panels),orwithmonoclonal antibodiestolarge T antigen, PAb 419 (11, 19) (lowertworowsof panels). Monoclonal antibodieswerepurified
bya50%ammoniumsulfatecutfrom culturesupernatantsof hybridomasgrownin staphylococcus-adsorbedfetal calfserum. Immunoprecipi-tates were run onOrtec 20% gels and fluorographed asdescribed previously (38). Control precipitations ofeach extractwith ammonium sulfatecutsof media showednobands.Sequential immunoprecipitations showed that antibody concentrations weresufficienttoprecipitate allbuttrace amountsof recognized proteins. (B) Stability of p53 afteralong pulse.Methodswere asdescribed for(A),exceptthat cellswere
labeledfor 1 hat39.3°C and for 2 h and 15min at31.50C. Chase timeswere6h(lane 1)or22h (lane 2).
antigenandp53didnotvarywithtemperature. NoTantigen
was seen at the top of the gradient, where monomers and
dimers would be expected in these standardgradients (32). There was a peak of T-antigen oligomers just below the
middleof thegradient, and heavy p53-T-antigen complexes could be seen infractions nearthe bottom.
Inthe tsA58 transformant grownat lowtemperature, p53 sedimented as in the wild-type transformant, although T
antigen banded more broadly withmore label in the
mono-mer-dimer region. At high temperature, as reported
previ-ously in other tsA58-transformed cells (4, 17), T antigen sedimented largely in the monomer-dimer region, and p53
was notdetectableoverthe background.
InthetsA1499 transformant, Tantigensedimentedovera
broaderrangethan in the other cells tested, butp53wasseen
in the same fractionsas in the parallel wild-type gradients.
There was nostriking difference in patternbetween cellsat
different temperatures where the transformed phenotype clearly varied. When thesegradients wereprecipitatedwith
anti-p53insteadof anti-T-antigenserum,similarresultswere
obtainedfor the distribution of the p53-T-antigencomplex, and no p53 was detectable sedimenting outside of the complex region (datanot shown).
In both the tsA58- and tsA1499-transformed cells, the maintenance of transformation appears to depend on T
antigen,as transformation is sensitiveto temperature shifts
dl
T-p
53=
F(tsA 1499)1
p
12 3
p
53=-anti-p53
39.30
p 53=
_
-
-p
53=-31
.50
anti-T
B F I
Il
39.30
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.138.483.70.483.2]1000 NOTES
tsA1499 transformants expressing the normal phenotype is functional. It is possible that the p53-T-antigen complex is
50
functionally
cold sensitive withoutdisplaying
metabolicla-31@5
bility.
Alternatively, the p53-T-antigen complex could beperfectly functionalatlow temperature, butits activity isnot involved in themaintenance of transformation. Although the latterexplanation is most appealing,anassayfor the activity
39.30
ofp53 orp53-Tantigen or both will probably be needed to distinguish rigorously between these alternatives.FtsA1499)1
dT- * 31.50
p53-dl T- 3930
p53-F(tsA58)A
T- *
-31.5
p53-
T-39.30
p53-FIG. 2. T-antigen-p53 complexes. Cells described in Table 1
werelabeled with32pfor120minateither 31.5or39.3°C,with twice
as much label atthe lowertemperature. Tomaintain temperature
controls, all media were prewarmed, and plates were left undis-turbed inincubatorsfor the pulse period. Extracts prepared in Tris-bufferedsaline (pH 7.4) containing 0.5% Nonidet P-40were
centri-fuged in parallel through 5 to20% linear sucrosegradients by the
procedure of McCormick and Harlow(32). Fractions were
collect-ed,immunoprecipitated with monoclonal anti-T antigen, andrun on
gelsasinFig. 1,exceptwithout fluorography. Sedimentation is from righttoleft. dl Tmarks the shortenedTantigen produced bymutant
tsA1499.
only in the mutant transformants and not in the wild-type transformants (Table 1) (38). Yet, only in the tsA58
trans-formant is the loss of transformation accompanied by a
dramatic decrease in p53 stability and loss of the p53-T-antigen complex. In the tsA1499 transformant, p53 is quite stable whether transformation is being expressed or not.
These results indicate that p53 stabilization alone is not sufficient to maintain the transformed state. The possibility that p53 stabilization, in additionto being insufficient, may
sometimesalsobeunnecessarytomaintaintransformation is raisedbyobservations that (i) in SV40, twoplasmid-cloned deletedgenomes are ableto transform, although their
trun-catedlargeT antigens failto stabilizep53 (8, 43); and (ii) in adenovirus, fully transformed cells can be derived from
transfections with DNA fragments unable to make a
full-sized p58, the adenovirus tumorantigen known tocomplex with and stabilize p53 (16, 41a).
The data presented in this paper do not address the question of whether the persistent p53-T-antigen complex in
Wewould liketothank Ed Harlow for the hybridoma producing PAb 419, John Carinofor help with early experiments, and Mary Headand Karen Mayerfor excellent technical assistance.
LITERATURE CITED
1. Bouck, N., N. Beales, T. Shenk, P. Berg, and G. di Mayorca. 1978. New region of the simian virus 40 genome required for
efficient viral transformation. Proc. Natl. Acad. Sci. U.S.A. 75:2473-2477.
2. Brockman, W. W. 1978. Transformation of BALB/c-3T3 cells by tsAmutantsof simian virus 40:temperaturesensitivity of the transformed phenotype and retransformation by wild-type
vi-rus.J. Virol.25:860-870.
3. Carroll, R. B., A. S. Blum, and D. S. Greenspan. 1982. Host
nuclearphosphoproteins that complex with simian virus 40 large Tantigen. Adv. Viral Oncol. 2:103-136.
4. Carroll,R.B.,and E.G.Gurney. 1982.Time-dependent
matu-ration of the simian virus 40large T antigen-p53 complex studied by using monoclonal antibodies. J. Virol. 44:565-573.
5. Chandrasekaran, K., V. W. McFarland, D. T. Simmons, M. Dziadek, E. G.Gurney, and P. T. Mora. 1981. Quantitation and characterization ofa species-specific andembryostage depen-dent 55-kilodalton phosphoprotein also present in cells
trans-formedby SV-40. Proc.Natl.Acad. Sci. U.S.A. 78:6953-6957. 6. Clark, R., D. P.Lane, and R.Tjian. 1981. Useof monoclonal
antibodies as probes of simian virus 40 T antigen ATPase
activity. J. Biol. Chem. 256:11854-11858.
7. Clark, R., K. Peden, J. M. Pipas, D. Nathans, and R. Tjian. 1983. Biochemical activities ofT-antigen proteins encoded by simianvirus 40Agenedeletionmutants.Mol. Cell. Biol. 3:220-228.
8. Clayton, C. E.,D.Murphy,M.Lovett,and P. W.J. Rigby.1982.
A fragment of the SV-40 large T-antigen gene transforms. Nature(London) 299:59-61.
9. Colby, W. W., and T. Shenk. 1982. Fragments of the simian
virus 40transforminggenefacilitatetransformation ofrat
em-bryocells. Proc. Natl. Acad. Sci. U.S.A. 79:5189-5193. 10. Cosman,D.J.,and M.J.Tevethia. 1981. Characterization ofa
temperature-sensitive, DNA-positive, non-transformingmutant
of simian virus 40. Virology 112:605-624.
11. Crawford, L., K. Leppard, D. Lane, and E. Harlow. 1982.
Cellularproteinsreactive withmonoclonal antibodies directed
against simianvirus 40T-antigen. J. Virol.42:612-620.
12. Crawford,L.V.,D. C.Pim,E.G.Gurney,P.Goodfellow,andJ. Taylor-Papadimitriou. 1981. Detection ofacommonfeature in several humantumorcell lines-a53,000-dalton protein. Proc. Natl. Acad. Sci. U.S.A. 78:41-45.
13. DeLeo,A.B.,G.Jay,E.Appella,G. C.Dubois,L. W.Law,and L.J.Old. 1979. Detection ofatransformation-relatedantigenin
chemicallyinducedsarcomasand other transformed cells ofthe
mouse. Proc. Natl. Acad. Sci. U.S.A. 76:2420-2424.
14. Dippold,W.G., G.Jay,A. B.DeLeo,G.Khoury,and L.J.Old. 1981.p53transformation-relatedprotein:detectionby monoclo-nalantibodyinmouseand human cells. Proc.Natl. Acad. Sci.
U.S.A. 78:1695-1699.
15. Giacherio, D., and L. P. Hager. 1979. A Poly(dt)-stimulated ATPaseactivityassociated with simian virus 40largeTantigen. J.Biol. Chem. 254:8113-8116.
16. Graham, F. L., P. J. Abrahams, C. Mulder, H. L.Heyneker,
S. 0.Warnaar,F. A.J.deVries,W.Fiers,andA.J.vanderEb. 1975. Studies on in vitro transformation by DNA and DNA
p53-J. VIROL.
F(w4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.58.299.73.390.2]fragments of human adenoviruses and simian virus 40. Cold Spring HarborSymp. Quant. Biol. 39:637-650.
17. Greenspan, D.S., andR. B. Carroll. 1981. Complex of simian virus 40 large tumor antigen and 48,000 dalton host tumor antigen. Proc. Natl. Acad. Sci. U.S.A. 78:105-109.
18. Gurney,E.G.,R.0.Harrison,andJ.Fenno.1980.Monoclonal antibodiesagainst simian virus40antigens: evidence for distinct subclasses of largeTantigen and for similarities among nonviral Tantigens. J. Virol. 34:752-763.
19. Harlow, E.,L.V.Crawford,D.C. Pim, andN.M.Williamson. 1981.Monoclonalantibodies specific for simian virus 40tumor antigens.J. Virol. 39:861-869.
20. Henning, R., J.Lange-Mutschler,and W.Deppert. 1981. SV-40 transformed cells express SV-40 Tantigen-related antigens on cellsurface. Virology 108:325-337.
21. Hiscott, J. B.,and V. Defendi. 1979.Viral and cellularcontrol of the SV-40 transformed phenotype. Cold Spring Harbor Symp. Quant. Biol. 44:343-352.
22. Jay,G.,G. Khoury,A.B.DeLeo, W.G. Dippold,and L.J. Old. 1981.p53 transformation-related protein: detection of an associ-atedphosphotransferase activity.Proc.Natl. Acad.Sci. U.S.A. 78:2932-2936.
23. Jornvall, H., J. Luka, G. Klein, and E. Appella. 1982. A 53-kilodalton protein common to chemically and virally trans-formed cells shows extensive sequence similarities between species. Proc. Natl. Acad.Sci. U.S.A. 79:287-291.
24. Klein,G.(ed.).1982.Thetransformation-associated cellularp53 protein.RavenPress, NewYork.
25. Levine, A. J., M. Oren, N. Reich, and P. Sarnow. 1982. The mechanisms regulating levels of cellular p53 tumor antigen in transformed cells. Adv. Viral Onco:. 2:81-102.
26. Linzer,D. I. H.,and A.J. Levine. 1979.Characterization ofa 54K dalton cellular SV-40 tumor antigen present in SV-40 transformed cells and uninfected embryonal carcinoma cells. Cell 17:43-52.
27. Linzer,D.I.H.,W.Maltzmon,and A.J.Levine.1979.The SV-40 Agene product is required for the production ofa 54,000 molecularweight cellulartumorantigen. Virology 98:308-318. 28. Luka, J., H. Jornvall, and G. Klein. 1980. Purification and
biochemical characterization ofthe Epstein-Barr virus-deter-mined nuclearantigen andanassociated protein witha 53,000-daltonsubunit. J. Virol. 35:592-602.
29. Maltzman, W., M. Oren,and A.J.Levine.1981. Thestructural relationships between 54,000 molecular weight cellulartumor antigens detected in viral and nonviral-transformed cells. Virol-ogy 112:145-156.
30. Martin,R.G.,V. P.Setlow,andC. A. F. Edwards. 1979. Roles of the simian virus 40 tumor antigen in transformation of Chinese hamsterlung cells: studies with simian virus40double mutants.J. Virol. 31:596-607.
31. May, E.,C.Lasne, C. Prives, J. Borde,and P.May. 1983.Study of the functional activities concomitantly retained by the 115,000 Mr superTantigen, an evolutionary variant of simian virus40largeTantigen expressed in transformedratcells. J. Virol. 45:901-913.
32. McCormick, F., and E. Harlow. 1980. Association ofamurine 53,000-daltonphosphoproteinwith simian virus40large-T anti-genin transformed cells. J. Virol. 34:213-224.
33. Mercer, W. E., D. Nelson, A. B. DeLeo, L. J. Old, and R. Baserga. 1982.Microinjection of monoclonal antibodytoprotein p53 inhibits serum-induced DNA synthesis in 3T3 cells. Proc. Natl. Acad. Sci. U.S.A. 79:6309-6312.
34. Milner, J., and F.McCormick.1980. Lymphocyte stimulation: concanavalin Ainduces expression of 53Kprotein. Cell Biol.
(Int. Rep.) 4:663-667.
35. Milner, J., and S. Milner. 1981. SV-40-53Kantigen: apossible rolefor 53K in normal cells. Virology 112:785-788.
36. Mora, P. T., K. Chandrasekaran, J. C. Hoffman, and V. W. McFarland. 1982.Quantitation ofa55Kcellularprotein: similar amount and instability in normal and malignant mouse cells. Mol.Cell. Biol. 2:763-771.
37. Oren, M., W. Maltzman,and A. J.Levine. 1981. Post-transla-tional regulation of the54Kcellulartumorantigen in normaland transformed cells. Mol. Cell. Biol. 1:101-110.
38. Pintel, D., N. Bouck, and G.di Mayorca. 1981. Separation of lytic andtransforming functions of the simian virus40 Aregion: two mutantswhich are temperature sensitivefor lytic functions haveopposite effectsontransformation. J. Virol.38:518-528. 39. Pintel, D., N. Bouck, G. di Mayorca, B. Thimmappaya, B.
Swerdlow, and T. Shenk. 1980. SV-40 mutanttsA1499 is heat sensitive forlytic growth but generates cold-sensitive rat-cell transformants. ColdSpringHarbor Symp. Quant. Biol. 44:305-309.
40. Rotter, V.1983. p53atransformation-related cellular-encoded protein can be used as abiochemical marker for the detection of primary mouse tumor cells. Proc. Natl. Acad. Sci. U.S.A. 80:2613-2617.
41. Rotter, V.,0. W. Witte, R. Coffman, and D. Baltimore. 1980. Abelsonmurine leukemia viral-induced tumors elicit antibodies againstahostcell protein,P50.J. Virol.36:547-555.
41a.Rowe,D.T., and F. L.Graham. 1983.Transformation of rodent cellsby DNA extracted fromtransformation-defective adenovi-rusmutants. J. Virol.46:1039-1044.
42. Sarnow, P., Y. S. Ho, J. Williams, and A. J. Levine. 1982. Adenovirus Elb-58kd tumor antigen and SV-40 large tumor antigenare physically associated with the same 54kd cellular protein in transformed cells. Cell 28:387-394.
43. Sompayrac, L. M., E. G. Gurney, and K. J. Danna. 1983. Stabilization of the 53,000-daltonnonviraltumorantigen isnot required for transformation by simian virus 40. Mol. Cell. Biol. 3:290-296.
44. Soprano, K. J., N. Galanti, G. J. Jonak, S. McKercher, J. M. Pipas,K.W. C. Peden,and R.Baserga. 1983. Mutational analy-sis of simian virus 40 Tantigen: stimulation of cellularDNA synthesis and activation ofrRNAgenesby mutants with dele-tions in theT-antigen gene. Mol. Cell. Biol. 3:214-219. 45. Staufenbiel, M., and W. Deppert. 1983. Different structural
systems of the nucleus are targets for SV-40large T antigen. Cell 33:173-181.
46. Steinberg, B., R. Pollack, W. Topp, and M. Botchan. 1978. Isolation and characterization ofTantigen negativerevertants fromaline oftransformedratcellscontainingonecopyof the SV40genome. Cell 13:19-32.
47. Stitt,D. T.,and W. F. Mangel. 1981.Analysis ofthenonviral antigens immunoprecipitable by SV-40Tantibody from SV-40-transformed human/mouse hybrid cell lines. Virology 114:149-160.
48. Stringer, J.R. 1982. Mutantof simian virus40large T-antigen that is defective for viral DNA synthesis, but competent for transformation ofculturedratcells. J. Virol.42:854-864. 49. Tjian,R., A.Robbins,andR. Clark. 1979.Catalyticproperties
of the SV-40largeTantigen. Cold Spring Harbor Symp. Quant. Biol. 44:103-111.
50. Tooze, J. (ed.). 1980. DNAtumorviruses. ColdSpring Harbor Laboratory, ColdSpring Harbor, N.Y.
51. Yamaguchi, N., and T. Kuchino. 1975. Temperature-sensitive mutantsof simian virus40selectedby transforming ability. J. Virol. 15:1297-1301.
![FIG.1.atall24panels),tatesbylabeledmMsulfate 39.30C h but (A) Stability of p53. Cells described in Table 1 were labeled in quadruplicate with [35S]methionine for 50 min at 31.50C or for 20 min and either extracted at once (lane p) or chased in media contai](https://thumb-us.123doks.com/thumbv2/123dok_us/1435363.96027/3.612.138.483.70.483/tatesbylabeledmmsulfate-stability-described-labeled-quadruplicate-methionine-extracted-chased.webp)
