Vol. 65, No. 12 JOURNALOFVIROLOGY, Dec. 1991,p. 6645-6653
0022-538X/91/126645-09$02.00/0
CopyrightX 1991, American Society for Microbiology
Human T-Cell
Leukemia
Virus
Type II Rex Binding and Activity
Require
an
Intact Splice Donor Site and a Specific RNA
Secondary Structure
ALEXANDER C. BLACK,* CRISTINA T. RULAND, MICHELE T. YIP, JIE LUO, BICH TRAN,
ANUPAMAKALSI, EDDIE QUAN, MORDECAI ABOUD,t IRVIN S. Y. CHEN, AND JOSEPH D. ROSENBLATT
Division of Hematology-Oncology, Department of Medicine, UCLA School of Medicine, LosAngeles, California 90024-1678
Received 7 August1991/Accepted12September 1991
Thehuman T-cell leukemia virus type II (HTLV-II) regulatory protein Rex augments cytoplasmic levelsof unsplicedgag-pol mRNA by acting through a Rex-responsive element (RxRE) in the long terminal repeat. Purified Rex protein binds to long terminal repeat mRNA. Here, using an immunobinding assay to measure the binding of Rex protein to mutated RxRE RNAs, we show that efficient Rex binding requires astem-bulge-loop RNAsecondary structure(nucleotides [nt] 465 to 500) and specific sequences both within the stem-bulge (nt 470
to 476) and within a conserved upstream splice donor site (nt 449 to 455). Rex function in a transient transfection expression system correlates with Rex protein-RxRE RNA binding. The ability of HTLV-II Rex to interact directly with the HTLV-ll splice donor site suggests that HTLV-II Rex may increase expression of unspliced gag-pol mRNA, inpart, byinhibiting splicing.
HumanT-cellleukemiavirus type II (HTLV-II) has been
linked to several cases of chronic T-cell malignancy in humans (21, 31). Epidemiological studies over the last sev-eral years havefound ahigh prevalence ofHTLV-II
infec-tionamongintravenous drug abusers and seropositive blood donors in the United States and Europe (23, 29, 33). How-ever, unlike HTLV type I (HTLV-I), which causes both adult T-cell leukemia (19, 28, 36, 38, 39) and
HTLV-I-associated myelopathy(3, 14), HTLV-IIis not clearly
asso-ciated withhumandisease.
HTLV-Iand -II haveasimilar retroviralorganizationwith
gag, pol, and env structural genes and two trans-acting regulatory genes, tax and rex (8-13, 17, 32, 35). Tax is
required forefficient transcription from the 5' long terminal repeat(LTR) (8, 10-13,32), whereasRexregulates
process-ing of viralmRNAs(1, 2, 4,16, 18, 20, 22, 26, 27, 30, 34).In
HTLV-I, Rex increases cytoplasmic levels ofgag-pol and
envmRNAs(1,2,16,18,20, 27,34)andrequiresacis-acting Rex-responsive element(RxRE) in the 3' LTR that forms a complex RNA stem-loop structure (1, 2, 6, 16, 34). In HTLV-II, we have shown that Rex augments cytoplasmic
levels of gag-pol mRNA and that this function involves similar cis-acting sequences in the 5' LTR, the RxRE (4).
Othergroupshavedemonstratedthat Rexincreases
expres-sion ofunspliced RxRE-containingmRNAand havemapped
theHTLV-IIRxREtoboth the 5' LTR(26)and the 3' LTR
(22). Using HTLV-II Rex purified from a baculovirus expression system, we have demonstrated that HTLV-II Rex protein binds directly to HTLV-II LTR RNA (37). Othergroups have subsequently shown that
HTLV-I-puri-fiedRexprotein expressed in bacteriabinds directlyto the HTLV-I RxRE (2, 6, 15). Here, we delineate the specific requirements for Rex protein binding to RxRE RNA and correlate bindingwithRexfunction.
*Correspondingauthor.
tPermanent address: Faculty of Health Sciences, Ben Gurion Universityof theNegev,Beersheva,Israel 94105.
MATERIALS ANDMETHODS
Plasmid constructs. (i) RxRE deletion mutants for
RNA-binding studies.Segments ofthe HTLV-II LTRwereexcised
from preexisting LTR-chloramphenicol acetyltransferase (CAT)constructs (4)andsubcloned intoeitherpBluescript-1 (Stratagene)orpGEM-1 (Promega)for in vitro RNA synthe-sis (see Fig. 1). Thefollowing isalist ofconstructs witha
simplifieddescription ofthecloning procedure.
(a)
pGEM-1
LII361-786.pMl3+LTRIICAT,
aconstructlinking the bacterial CAT gene to the 5' LTR (4), was
digested withBamHI and EcoRI, anda425-nucleotide (nt)
BamHI (nt 361)-EcoRI (nt 786) fragment was isolated and subcloned into the pGEM-1 polylinker following digestion
with BamHI andEcoRI.
(b)pMl3+LII361-786.pGEM-1 LII361-786wasdigested
with HincIl and EcoRI, and the HincII-EcoRI fragment (containingnt361to786)wasisolatedandsubcloned intothe
EcoRV-EcoRIpBluescript SK
(pMl3+)
polylinker sites.(c)
pMl3+LH
361-520.pGEM-1LII361-786wasdigested
with BamHI (nt 361) and, after the overhang was filled in with Klenow fragment, digested with XmaI (nt 520). The
BamHI-XmaI fragmentwasisolatedand subclonedinto the EcoRV andXmaI sites in the
pMl3+
polylinker.(d)
pMl3+LII
405-630.Thepreexistingconstruct,pLA630CAT (4), was digested with AvaII (nt 405), filled in with Klenowfragment, and digestedwithXbaI. TheAvaII-XbaI fragment (nt405to630)wasisolated andsubcloned intothe SmaI and XbaI sites of
pMl3+ polylinker.
(e) pMl3+LII 405-520. A
preexisting
LTR deletion mu-tant,pLA520CAT
(4),wasdigestedwithAvaIl(nt405),filled in with Klenow fragment, and digested with XmaI. TheAvaII-XmaI
fragment (nt 405 to 520) was isolated andsubcloned into the EcoRV and XmaI sitesof
pMl3+
poly-linker.(f) pGEM-1 LII 483-786. pGEM-1 LII 361-786 was
di-gestedwithBsu36I (nt483),filled in with Klenow
fragment,
digested with
HindII,
which removes nt 361 to482,
andreligated.
(g)
pGEM-l
LII 460-520. Anoligonucleotide
thatcon-6645
on November 10, 2019 by guest
http://jvi.asm.org/
tained HTLV-II sequences from nt 460to 520, flanked by
PstIand XbaIrestriction sites (courtesy of W. Wachsman), was synthesizedand subclonedinto thePstIand XbaI sites of the pGEM-1 polylinker.
(h) pGEM-1 LII 460-503.pGEM-1 LII 460-520wascleaved with AvaI (nt 503 and twice in the pGEM-1 polylinker), filled in with Klenow fragment, and religated.
(i)pGEM-1 LII361-786Mst/K. pGEM-1 LII 361-786was cleaved with Bsu36I (previously MstII), filled in with Kle-nowfragment, whichrepeatsthe TTAsequenceimmediately after nt486 (nt 484 to486 [TTA]), and religated.
(j) pGEM-1LII361-786Mst/MB.pGEM-1 LII361-786was cleaved with Bsu36I (MstII), digested with mung bean nu-clease, whichremovessingle-stranded DNA overhangs (and thus deletesnt484to486, [TTA]), and religated.
(k) pGEM-1 LII361-786 A470-473. pGEM-1 LII 361-786 was cleaved withSstIand Klenowfragment, whichdigested the 3' overhang of SstI (and thus deleted nt 470 to 473,
[AGCT]), andreligated.
(1) pM13+LII 361-786 s449,450. A preexisting construct, pM13+LIIASDCAT, containinga2-ntsubstitutionatnt449 to450in thespliced donor site (GG--TT) (4) (see Fig. 1B), wasdigested withBamHI, filled in with Klenow fragment,
anddigested with EcoRI. TheBamHI-EcoRI fragment (with the 2-ntsubstitution at nt449and 450 indicated by s449,450) wasisolated and subcloned intopM13+cleaved withEcoRV and EcoRI.
(m) pM13+LII 451-520. pMl3+LIIASDCATwas cleaved withMseI(cutsatenzymesiteTTAA, nt449to452,created
by substitution at nt 448 to 449 [GG->TT]), filled in with Klenowfragment, and digested withXmaI (whichcuts atnt 520, 645, and 750 in HTLV-II and in the polylinker). The
70-nt MseI-Klenow-XmaI fragment was isolated (nt 451 to 520) and subcloned into pM13+ cleaved with EcoRV and
XmaI.
(ii)RxREnucleotidesubstitutionmutantsfor RNA-binding
studies. The following constructs were site-directed dele-tions or substitutions in pM13+LII 361-786. Mutagenesis was performed by the Amersham oligonucleotide-directed method (25) and confirmed by dideoxy sequencing. The constructs and oligonucleotides used to direct mutagenesis areas follows.
(a) pM13+LII 361-786 A465-501. 5'-CTAAAGACCCT TGAGAGGAGAC-3', complementary to nt 454to 464and
502to 512, deletes nt465to501.
(b) pMl3+LII 361-786 s447,448. 5'-AGACTTACCGCGG GAGGAGC-3mutates nt477 and 448 (AA-*GC). This sub-stitution creates a new restriction enzyme site, SacIl (5'-CCGCGG-3').
(c) pM13+LII 361-786 s453,454. 5'-GAGAGGAGAGGT ACCTTGGG-3' mutates nt 453 and 454 (AG->CC). This
substitution creates a new restriction enzyme site, KpnI
(5'-GGTACC-3').
(d) pM13+LII 361-786 s467,468. 5'-GCCGAGCTCTTC CTGAGAGG-3' mutates nt467 and468 (TC--AA).
(e) pM13+LII 361-786 s470,471. 5'-GCAGCCGAGGGC GACCTGAG-3' mutates nt470and471 (AG--CC).
(f) pM13+LII 361-786 s470,471, s495,496. 5'-CCCTCG GGGCCCGACTACCT-3 mutates nt495 and 496 (CT--GG) in pM13+LII 361-786 s470,471. This substitution creates a
newrestriction enzymesite, NlaIV (5'-GGNNCC-3'). (g) pMl3+LII 361-786 s473,474. 5'-GGGGCAGCCTTGC TCGACCT-3' mutates nt473 and 474(TC->AA).
(h) pM13+LII 361-786 s479. 5'-CCTAAGGGGAAGCC GAGCTCG-3' mutates nt479(G->T).
(i) pM13+LII 361-786 s495,496. 5'-CCCTCGGGGCCCG
ACTACCT-3' mutatesnt 495 and 496(CT--GG). This sub-stitution creates a new restriction enzyme site, NlaIV (5'-GGNNCC-3').
(iii)CMV-CAT constructs used in the transient transfection geneexpression assay. (a)pCMVLII 361-786 CAT. A preex-isting construct, pM13+LTRIICAT (4), was cleaved with
Hindlland Bsu36I(whichdeletesnt0to483). Apreexisting
construct containing genomic HTLV-II sequences from nt
361 to 8550 downstream of the cytomegalovirus (CMV) immediate-early gene promoter,BC-HTLV(5),wascleaved with Hindll and Bsu36I. The Hindll to Bsu36I fragment
containstheCMV immediateearlypromoter,80 nucleotides of leaderCMVsequences,andnt361to483 in theHTLV-II LTR. This fragment was subcloned into the pM13+LTRI
ICAT vector, withnt0to 483 deleted.
(b) pCMVLII 361-520 CAT. pCMVLII 361-786 CAT was
cleaved with SmaI (nt 520 and the pM13+ polylinker
up-streamof theCATgene) and religated(deletion ofnt520to
786).
(c)pCMVLII361-786Mst/K CAT.pCMVLII361-786 CAT was cleaved with Bsu36I (MstII), filled in with Klenow fragments, which duplicates nt 484 to486 (TTA), and
reli-gated.
(d) pCMVLII 361-786 Mst/MB CAT. pCMVLII 361-786
CATwas cleaved with Bsu36I (MstII), digested with mung bean nuclease, which deletes nt 484 to 486 (TTA), and religated.
(e)pCMV-CAT. A preexistingblunt-endedfragment
con-taining the CAT gene and simian virus 40 large T-antigen polyadenylation sequencesfrompM13+LTRIICAT (4) was
cloned into the Klenow fragment-filled-in SstI site of the pM13+ polylinker to generate pM13+CAT. A preexisting
construct, BC12 (courtesy of B. Cullen)(5), whichcontains
theCMVpromoter, was digestedwith HindII and HindIII. Theapproximately 800-nt HindII-HindIII fragment
contain-ing the CMV promoterwasisolated and subcloned into the Hindll andHindIll sites of the pM13+ polylinkerupstream of the CAT gene in pM13+CAT.
(f) pCMVLII
405-630 CAT. Apreexisting
construct, pM13+LIIA630CAT (4), was cleaved with Avall (nt 405),filled in with Klenow fragment, and digested with SpeI (downstreamofnt630in thepM13+ polylinker).The 240-nt
AvaII-Klenow-SpeI fragment was isolated and subcloned into pCMV-CATcleaved with EcoRV and SpeI.
(g)pCMVLII 361-786 A470-473 CAT.pGEM-1LII361-786 A470 to 473 wasdigested withEcoRI, filled in with Klenow
fragment, and cleaved with Hindlll. The HindIII-EcoRI fragment was isolated and subcloned into the HindIII and EcoRVsitesofpCMV-CATbetween theCMV promoter and the CAT gene.
(h) pCMVLII 361-786 s449,450 CAT. A preexisting
con-struct, pMl3+LIIASDCAT (4), which has a nucleotide substitution at nt 449 to 450
(GG-*TT),
was cleaved with BamHI (nt 361), EcoRI (nt 786), and filled in with Klenowfragment. The 425-ntBamHI-EcoRI-Klenow fragment was
isolated and subcloned into pCMV-CAT, which had been cleaved at the SmaI (blunt) site. Only sense orientation
constructs were selected.
The following constructs were generated by cleaving pCMV-CATatXbaI andsubcloningtheXbaI-XbaIfragment
from thecorrespondingpMl3+LII 361-786 deletionor sub-stitution mutant as follows. Only sense orientation con-structs wereselected.
(i)pCMVLII361-786A465-501CAT.TheXbaI-XbaI
frag-ment wasfrom pM13+LII 361-786A465-501.
on November 10, 2019 by guest
http://jvi.asm.org/
HTLV-II Rex FUNCTION AND BINDING 6647
()
pCMVLII
361-786 s467,468 CAT. The XbaI-XbaIfragment was from
pM13+LII
361-786 s467,468.(k)
pCMVLII
361-786 s470,471 CAT. TheXbaI-XbaIfrag-ment was frompM13+ LII 361-786 s470,471.
(1) pCMVLII 361-786 s473,474 CAT. The XbaI-XbaI frag-ment was frompMl3+LII 361-786 s473,474.
(m) pCMVLII 361-786 s479 CAT. TheXbaI-XbaIfragment was from
pM13+LII
361-786 s479.Immunobinding assay for Rex protein binding to RxRE mutants.
a-CTP-radiolabeled
and -unlabeled RNA tran-scripts were synthesized in vitro (Promega reagents) by using the RNA polymerase promoter sites that flank the polylinker, T3 RNA polymerase, T7 RNA polymerase in pBluescript (pMl3+) constructs, and T7 and SP6 RNA polymerase in pGEM-1 constructs, as previously described (37). The immunobinding reactions were performed as pre-viously described (37). Briefly, after preincubation of Rex protein alone at37°C
for 15min, a 2 nM reaction concentra-tion of Rex protein was incubated with 50 pg of uniformly labeled RNA (approximately 15,000 cpm in a50-,u
reaction volume) at4°C
for 15 min. Immunoaffinity-purified Rex-specific antiserum (0.5R1)
was added, and the reaction was incubated for 20 min at4°C.
Protein A-Sepharose (15RIu
of 25% solution) in phosphate-buffered saline was added and roll mixed for 30minat4°C
(37). The Sepharose beads were washed twice with a mixture containing 40 mM Tris (pH 8.0), 2mM
MgCl2, 200 mMKCI,
and 0.05% Triton X-100, and bound-RNA counts per minute were measured by liquid scintillation.Transfections. 729-6 Bcellsor Jurkat T cells (5 x 106)were transfected by electroporation with a total of 25 ,ug of indicated plasmids, as described previously (4, 7). The cells were incubated at
37°C
and 5% Co2 in Iscove's medium containing 5% fetal calf serum, harvested at 24 h, and assayed for CAT activity, as described previously (4, 7).RESULTS
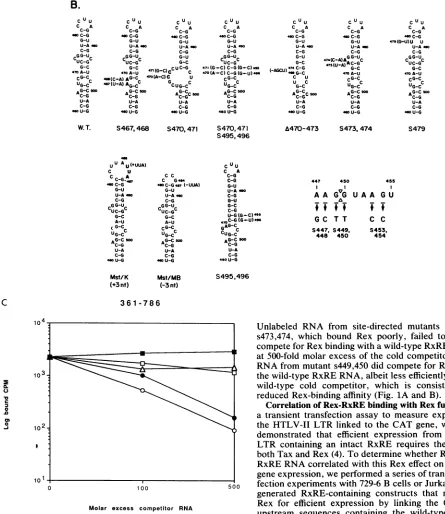
RNA sequence and secondary structure requirements for
HTLV-II
Rex protein binding to the RxRE. To identifyRex-RxRE-binding domains, we used a sensitive immu-nobinding assay to test for Rexbinding to a series of RxRE deletion and site-directed mutants (Fig. 1) (37). RNA from nt 361 to 786 inHTLV-II, which includes U5 and all but the 5' 46 nt of the R region, was employed as the positive control for Rex binding (37). The specificity of the Rex protein-RNA-probe binding was determined by competition with 2,000-fold molar excess of unlabeled nt 361 to786RNA (Fig.
1A, column labeled 2) or with excess nonspecific stem-loop-forming RNA from the human immunodeficiency virus type 1 (HIV-1) trans-activating response region, which does not bind Rex (Fig.1A, column labeled 3) (37).
The smallest segment of RNA to which Rex binds effi-ciently was from nt 405 to 520 (Fig.1A, column labeled 3). This is contained within the previously defined regions required for Rex binding (nt 361 to 520) (37) and for Rex regulatory effects on gene expression (nt 405 to 630) (4). This region contains both the HTLV splice donor consensus sequences (AAGGUAAGU, nt 447 to 455) (24) and a prom-inent stem-bulge-loop structure further downstream (nt 460 to 504) as predicted by the model of Zuker and Stiegler (40) (Fig. 1B). The deletion of the stem-bulge-loopstructure from nt 465 to 501 abrogates specific Rex binding(Fig.1A, probe
A465-501),
demonstrating that this RNA element is requiredfor Rex binding. Although necessary, this region is not sufficient for Rex binding, as is shown by the lack of binding
by
Rex to RNA from nt 460 to 503 and nt 460 to 520 (Fig.1A),
whichare also predictedto form the same stem-bulge-loop configuration. However, when sequences containingthe 3' portion of the LTR splice donor site are included
(UAAGU
ofthesplice site),
Rexbindsspecifically, althoughwith significantly lower affinity than to the positive control (Fig.
1A,
probe 451-520). Therefore, both thestem-bulge-loop structure and atleast the 3' portionof the splice donor site are required forefficient Rex binding.
Todetermine the precise Rex-binding domains within this
stem-bulge-loop structure, we tested a series of smaller
deletion and substitution mutants withinnt 465 to 501
(Fig.
1B).
Mutants that either add or delete a3-ntUUA sequence in the distal loop without altering the rest oftheproximal
stem-bulge-loop structure demonstrate efficientRex
binding
(Fig.
1A
and B, probes Mst/K and Mst/MB). Site-directedmutation ofnt479,which disrupts the distal stem, also binds
Rex (Fig. 1A and B,
probe
s479) twofold less avidly thandoesthe
wild-type.
In contrast, mutationswithinoradjacent
tothe bulge formed
by
nt472 to476 (CUCGG)significantly
reduce Rex binding. Deletion of nt 470 to 473markedly
reduces Rexbinding, despite the preserved distal stem-loopRNAstructureformedbynt476 to 492(Fig.1A and B,probe
A470-473). Substitution at nt470 and 471 (AG--CC), which
disrupts and enlarges the bulge but preserves the distal stem-loop structure, abrogates Rexbinding (Fig. 1A and B,
probe s470,471). A compensatory dinucleotide substitution, which regenerates the wild-type RNA secondary structure
(Fig. 1A and B, probe s470,471; s495,496) but contains the
substituted nt470 and 471, increases bindingto only21% of thatofthewild type, suggestingaspecific sequence
require-ment. The s495,496dinucleotidesubstitution in thewild-type
RxRE, which eliminates the wild-type RNA bulge from nt 472to 476 anddisruptsthe nt 469 to 472 stem butleaves the wild-type sequences at nt 470 and 471 unchanged (Fig. 1A and B, probe s495,496), also reduced binding to a degree
similar to that of the mutant, s470,471; s495,496. Substitu-tions at nt 473 and 474 (UC--AA), which change the sequence within the bulge (nt 472 to 476,
CUCGG--CAAGG) without altering the predicted RNA secondary structure, reduce specific Rex binding five- to sixfold
com-pared with that of the wild type (Fig. 1, probe s473,474).
These results suggest that both the stem-bulge RNA
struc-ture from nt 470 to 476 and specific sequences within that structure are
required
forefficient Rex binding. In contrast, substitution of nt 467 and 468 (UC-+AA) changes these-quenceof a smallerproximal bulge (nt 466 to 469,
GUCG--GAAG) but leaves the secondary structure intact and has onlyatwofoldeffectonRexbinding(Fig. 1,probes467,468).
In summary, efficient Rex-RxRE binding depends on an intact stem-bulge-loop RNA structurefrom nt 465 to 501, in particular, the sequences (nt 471 to 474, GCUC) and
struc-ture of thestem-bulgeformed by nt 470 to 476and 492 to 496. Since maximal Rex binding was seen with RNAs that extended upstreamof the splice donor site, we tested several dinucleotide substitutions within the splice donor consensus sequences (nt 447 to 455, 5'-AAGGUAAGU-3', with the splice site between nt 449 and 450 [GG]). Substitution of the 5' end of those sequences (Fig.1A and B, probe s447,448
[AA-GC]) only marginally affected Rex binding. In con-trast, substitution at either the splice donor cleavage site
(s449,450 [GG--TT]) or at the 3' end of the consensus sequences (s453,454 [AG--CC]) reduced Rex binding five-fold. None of these substitutions significantly disrupt the predicted RNA secondary structure (results not shown). VOL.65, 1991
on November 10, 2019 by guest
http://jvi.asm.org/
314 ... 763
I I . I I
HTLV-H LTR
(NucleotideBoundaries)
361-786
405-520
451-520
460-520
460-503
361,786, s447,448 361-786, s449, 450
361-786, s453, 454
361-786, A465-501
361-786, s467,468
361-786, A470-473 361-786, s470, 471 361-786, s470, 471; s495, 496
361-786, s473, 474 361-786. s479
361-786, MstIMB
361-786. MstIK 361-786,s495-496
520 Sense RNAP
449 562 RexPrc
SD ColdProbe ColdNonspecific(TAR)
361 786
405 520
6i
451 520
460 520
460 503
361 S 786
+
361 s 786
361 s 786
361
_I
465 501 786I
361 S 786
361
16
A361 s
4
786
786
361 s s 786
36 78
361 s 786
361 s 786
361 -3 nt 786
361 +3 nt 786
361 s 786
_
4,
Results
1 2 3
'robe + + +
otein + +
RNA- +
)RNA - - +
0.02 0.04 1.00
0.02 0.03 0.76
0.02 0.03 0.25
0.04 0.02 0.07
0.01 0.01 0.01
0.02 0.04 0.47*
0.02 0.02 0.19/0.27* 0.02 0.04 0.20* 0.02 0.02 0.06 0.02 0.03 0.52 0.03 0.02 0.11 0.01 0.01 0.05/0.15* 0.06 0.02 0.21* 0.03 0.03 0.14/0.22*
0.03 0.04 0.47
0.01 0.01 0.41
0.01 0.04 0.85
0.02 0.03 0.21*
FIG. 1. Immunobindingassayfor Rex protein bindingtoRxREmutants. (A) The radiolabeled RNA transcripts testedarerepresented schematically onthe left (37), and the mutations are described in Materials and Methods. The results of the immunobindingassay (see Materials and Methods)arerepresentedonthe rightas apercentageof theamountof radiolabeledmutantprobe bound by Rex proteinineach binding reactiontothepercentage of radiolabeled positive control probe (nt 361to786) bound by Rex in thepresenceofa2,000-foldmolar
excessofnonspecific competitor RNA (column 3) in thesameexperiment. Each columnrepresentsthe standardizedbinding conditions (see
Materials andMethods) withsenseRNAradiolabeled probe alone (column 1);senseprobe, 2 nM Rex protein, and 2,000-fold molarexcess ofcoldnt361to786 RNA(specific competitor) (column 2); andsenseprobe, 2 nM Rex protein, and 2,000-fold molarexcesscoldHIV-1 trans-activatorresponse(TAR) region RNA from HIV-1nt435to508(nonspecific competitor) (37) (column 3). These resultsrepresentpooled data fromatleast three differentexperiments with each probe. The standard deviation for eachprobe in each columnwasless thanorequal
to ±0.01 for columns 1 and 2 and +0.07 for column 3,exceptfor probe 405-520 (+0.17, column 3). The difference in relative immunobinding values betweenanytwoprobeswasstatistically significant (P<0.05,Student'sttest)when the differencewasgreaterthan 0.12.The asterisks denoteasecond preparation of heparin agarose-purified HTLV-II Rex protein (37) in which 5 nMRexwasused in binding reactions, dueto
theslightly lower binding activity of this preparation. (B) The predicted RNA secondarystructureofanessential stem-bulge-loop within the
HTLV-IIRxRE (nt 460to504)is depicted for the wild-typesequences(RxRE)and for the different deletion and site-directedmutantswithin this stem-bulge-loop. The specific dinucleotide substitutions with the consensus splice donor site (nt 447 to455 [AAGGUAAGU]) are
represented schematically. RNA secondarystructuresforallmutantstestedweregenerated bycomputermodeling by the methodof Zuker andStiegler (40). The complete predicted RNA secondarystructuresof the transcribedpartof theHTLV-II 5' LTR (nt 314to763,R/U5) and the 3' LTR (nt8190to8751,U3/R)have been published elsewhere(4, 34). (C)The resultsof cross-competition immunobinding experiment involvingapositive control probe(361-786)witha100- and 500-fold molarexcessof coldmutantRxRE RNA (s449,450[0];s470,471[A]; s473,474 [O], wild-type RxRERNA(361-786 [0]), and nonspecificcompetitor RNA (HIV-1 TAR[-])areshown. Theyaxisrepresentsthe log of radiolabeled probe bound byRex,andthexaxisrepresentsnocompetitor (origin) and 100- and 500-fold molarexcessofcompetitor
RNAs,asindicated. All immunobinding reactions involved2 nM Rexprotein andthebinding conditionsaredescribed above.
6648
I I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.135.479.50.513.2]HTLV-I1 Rex FUNCTION AND BINDING 6649
C U U C A C-G 4 C-G G-U U-A4-0 C-G GG-U UC-G
471(G-C)C-G(G-C)495
470(A-C)C-G(G-U)4"
CG-C UG_c AG-Csoo C-G U-A C-G 4.0U-G
C U U
C A C-6 4.0C-G G-U U-A4oo C-G G-U G-C
(-AGCU)474460G-CC-6
C U U C G-C AG-C S00W C-6 U-A C-6 40U-G C A C-G 460C-G G-U U-A4.0 C-G 4-4(C-A)A C
4n(U-A)AC-GG-C
470A-U CG-C UG-C AG-C500 C-G U-A C-G 40U-G
W. T. S467, 468 S470, 471
4M
UU AU(+UUA)
C U C A 40C-G G-U U-A*0o C-6 GG-UC UC-6 G-C A-U CG-C UG6c AG-C500 C-G U-A C-G 4.0U-G Mst/K (+3nt) c c
C G6m
440C-G4. (-UUA)
G-U U-A4o C-G cGG-U UC-6 G-C A-U CG-C UG-Cc AG-C5co C-G U-A C-G 40U-G Mst/MB (-3 nt) S470, 471 S495, 496 cUu C A C-G C-G 6-U U-A4.0 C-6 G-U G-C C-G
U-G(G-C)4.0
470C-G (G-U) 4.
AG-CC CUG-C A6-Cgoo C-6 U-A C-6 460U-6
A470-473 S473, 474 S479
447 450 455
AA GVG U AA GU
GC TT C C
S447, S449, S453,
448 450 454
S495,496
3 61-78 6
Molar excess competitor RNA
FIG. 1--Continued.
These results are consistent with the decreased binding of Rex to theRxRE deletion mutant containing the 3' part of the splicedonorsite(Fig. 1AandB, probe 451-520).
There-fore,efficient Rex-RxREbinding requiresnt449to455 ofthe splice donor (nt 447 to 455) and, in particular, appears to requirethespecific sequencesatnt449 and450 and 453 and 454 (GGNNAG).
The relativeaffinityof Rexbindingto mutantRxRE RNA wasfurthertestedincross-competition-binding assayswith excessunlabeled wild-typeormutantRxRERNA(Fig.1C).
Unlabeled RNA from site-directed mutants s470,471 and s473,474, which bound Rex poorly, failed to significantly competeforRexbindingwithawild-typeRxREprobe,even at 500-fold molar excess of the coldcompetitor. Unlabeled RNA frommutant s449,450 did compete forRex bindingto thewild-type RxRERNA,albeit lessefficientlythandid the wild-type cold competitor, which is consistent with the reduced Rex-binding affinity (Fig. 1Aand B).
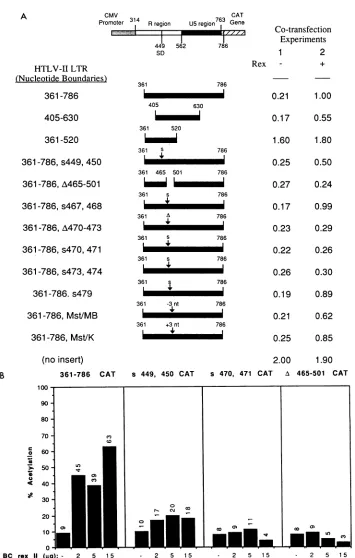
Correlation of Rex-RxREbindingwith Rex function. Using a transient transfection assay to measure expression from the HTLV-II LTR linked to the CAT gene, we previously demonstrated that efficient expression from an HTLV-II LTR containing an intact RxRE requires the presence of both Tax and Rex(4). Todetermine whether Rexbindingto RxRE RNAcorrelated with this Rex effecton RxRE-linked
geneexpression, weperformedaseries of transient cotrans-fection experimentswith729-6 B cellsorJurkatT cells. We generated RxRE-containing constructs that required only Rex for efficient expression by linking the CAT gene to upstream sequences containing the wild-type or mutated RxRE.Transcription of RxRE-CATmRNAwasinitiatedby
the CMV immediate early promoter (Fig. 2). The deletion and substitution RxREmutantsweresubcloned downstream from the CMV promoter in sense orientation in the CMV-CAT vector and cotransfected with the HTLV-II Rex expressionvectorortheplasmid control. Since RxRECAT
mRNA isconstitutivelytranscribedfrom the CMVpromoter
in all CMV-RxRE CAT constructs, differences in CAT expressionin these cotransfections aredue to the posttran-scriptional effect of Rex onthe wild-typeor mutated RxRE
leader sequences.
Gene expression from a CMV-CAT -construct without
HTLV-II LTR sequencesis unaffectedby Rex (Fig. 2A, no VOL. 65, 1991
B.
c u u C A C-G 460 C-G G-U U-Aao C-G GG-U UC-G G-C 4*0A-U CG-C UG-C AG-C s00 C-G U-A C-G 460U-G
C U U
C A C-G 460C-G G-U U-A4o C-G GG-U UC-G G-C 470A-U 4*(C-A)AG-C 4.(U-A)AG-C AG-Cso C-G U-A C-G 40U-G
C U U
C A C-6 460C-G G-U U-A4.0 C-G G-U G-C
471G-C)cCUC-6
40(A-C) C U
CCUG-C AG-CCsoo C-G U-A C-G 460U-G
C U U
C A
C-G
40C-G
47(G-U)U U U-A460 C-G GG-U UC-G G-C 470 A-U CG-C UG-cC AG-CSo C-G U-A C-G 4.0U-G C 2 0. 0 00 .0 -J
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.67.512.84.598.2]A
HTLV-LI LTR (Nucleotide Boundaries)
361-786
CMV CAT
c 314
763CA
Promoter Rregion U5 region I Gene
s:-..,:i-.-l.'~.*: A -77A
449 562
SD
786
361 786
405 630
Co-transfection
Experiments
1 2
Rex - +
0.21 1.00
405-630
361-520 361-786, s449, 450
361-786,A465-501
361-786,s467, 468 361-786, A470-473
361-786,s470, 471 361-786, s473, 474
361-786. s479 361-786, Mst/MB
361-786, Mst/K (no insert)
361-786 CAT
361 520
361 s
14
0.17 0.55
1.60 1.80 786
361 465 501 786
361 s 786
361 a 786
361 s 786
361 s 786
_
361 s 7861
361 -3nt 786
361 +3nt 786
_
+
s 449, 450 CAT s 470, 471 CAI
0.25 0.50
0.27 0.24
0.17 0.99 0.23 0.29 0.22 0.26 0.26 0.30 0.19 0.89 0.21 0.62 0.25 0.85 2.00 1.90
T A 465-501 CAT
90
80
70
o 60
a
. 50 < 40 30 20 10
-(D
LcA
0r)
0
0
r-. CMJNi
o
l
- 2 5 15
GiECD
"Itw
Mom
[image:6.612.129.484.74.632.2]- 2 5 15
FIG. 2. GeneexpressionassayofRxREmutants.(A) The CATexpressionvectorslinkedtoleadersequencescontainingthewild-typeor mutated RxRE are represented schematically on the left. Each construct contains an upstream CMV immediate early promoter that
constitutively drivestranscriptionofthe RxRE leadersequencesand the CATgene.Thepolyadenylationsignalisprovided bysimianvirus
40large T-antigensequences(4). 729-6 B cells (5 x106)weretransfectedbyelectroporationwithatotal of 25 ,ugofplasmid (seeMaterials andMethods). Transfected cellswereharvestedat24 h andassayed for CAT activity (see MaterialsandMethods).On theright,theresults oftransient transfectionexperimentsarerepresentedas aratio of thepercentacetylation ofthepositivecontrolplasmid (RxREinsert361-786) withcotransfectedHTLV-II Rexexpressionvector(pBCrexII) (5)tothepercentacetylation of themutantwithorwithout cotransfected Rex
vector.pCMV-RxRE-CATvector(15,ug)wascotransfected witheither 10 ,ug of BC12 (no Rex)or10,ug of BCrexII (Rexexpressed).The results representtheaverageof threeor moreseparatetransfectionexperiments in whichapositive controlwasalsopresent. The standard deviationofrelativebindingvalues foreachprobeineachcolumnwaslessthanorequalto+0.04 forcolumn 1and +0.07 for column 2. The
B
BC rex 11 (9g):- 2 5 15
0Da)
No
_o
2 5 15
on November 10, 2019 by guest
http://jvi.asm.org/
HTLV-II Rex FUNCTION AND BINDING 6651 insertconstruct). In addition,aspredictedfrom our previous
functional studies of Rex and the HTLV-II LTR (4), Rex effects on gene expression wereobserved only inCMV-CAT constructs containing the cis-acting repressive sequences (CRS) from nt 520 to 630, which inhibit nuclear-to-cytoplas-mic transport of CRS-containing mRNA unless both an intact RxRE and Rex are present (Fig. 2A; compare pCM-VLII 361-786 CAT,which contains theCRS, with pCMVLII 361-520 CAT, which lacks the CRS). CMV-RxRE-CAT constructs containing RxRE mutants in which Rex binding wasconserved (361-786, 405-630, Mst/K, Mst/MB, s467,468 and s479) demonstrated a consistent three- to fivefold in-crease inexpressionwithcotransfected Rex compared with the cotransfected vector control (Fig. 2A). CMV-RxRE-CAT constructs containing RxRE mutants in which Rex binding was markedly decreased (A465-501, A470-473, s470,471, and s473,474) showed little or no increase in
expression with cotransfected Rex (Fig. 2A). These results suggestthat inthis transient transfection assay, efficient Rex binding to RxRE RNA is required for Rex function. In
titrationexperiments with increasing amounts of Rex vector,
nearly maximal expression wasseen with the positive con-trol (Fig. 2B, RxRE insert 361-786) at a 5:1 ratio of CMV-CAT vector to Rex expression vector, whereas CMV-CMV-CAT vectors containing an RxRE mutant that fails to bind Rex
(Fig.2B, RxRE inserts A465-501 and s470,471) demonstrate no significant Rex responsiveness for any amount of Rex vector tested. In addition, an RxRE mutant that has a
fivefoldlower levelof specific binding demonstrates a mea-surablebut blunted response to Rex in the CMV-RxRE-CAT assay system(Fig. 1A and2Aand B, RxREinsert s449,450). Therefore, the affinity ofRexbinding appears to correlate with the degree of Rex responsiveness in a functional assay.
DISCUSSION
Ourresults represent the first detailedmapping of both the RNA sequence- and secondary structure-specific Rex
pro-tein-binding domains within RxRE and the correlation of binding with Rex regulatory effects on gene expression in HTLV-II. We have shown that efficient in vitro HTLV-II Rex proteinbinding to RxRE RNA requires a stable
stem-bulge-loopRNAsecondary structurefromnt465 to 501 and
specificsequenceswithin thestem-bulgefromnt470 to 476. Inaddition, Rexprotein-RxRERNAbinding and Rex
func-tion requirent449 to 455of the consensus sequences (nt 447
to 455, 5'-AAGGUAAGU-3') in the HTLV-II LTR splice
donor, suggesting that Rex may interact directly with the
splice donor site in vivo. One possibility is that Rex may competewith Ul small nuclearribonucleoprotein (snRNP) bindingtothesplicedonor site in the LTRand, thus, inhibit
splicingandfacilitate nuclear-to-cytoplasmic exportof gag-pol mRNA.
Using an HTLV-I Rex functional assay, several groups
identifiedanHTLV-I RxRE in U3/Rofthe 3' LTR (1, 2, 16,
34). In previous functional studies ofthe HTLV-II RxRE,
ourgroupaswellas Kim and coworkersdemonstrated that the HTLV-II RxRErequires sequences only withinR(4, 22).
Recently, Ballaun and coworkers have shown that the HTLV-I Rex function through the HTLV-I RxRE deletion mutants correlates with the ability of the HTLV-I Rex protein to bind to mutated RxRE RNA in vitro (2). A critical RNAsecondary structure within theHTLV-I RxRE (nt 457 to 545 of HTLV-I), which is required for HTLV-I Rex
function(1, 2, 16, 34) andHTLV-I Rexprotein-RxRE RNA binding (2, 6), appears to be similar in both structure and
specific nucleotide sequences to the stem-bulge-loop ele-ment that we have mapped within the HTLV-II RxRE (nt 465to501). Usingasensitive immunobindingassay todetect therelative affinity ofHTLV-II Rexbinding to RxRE RNA, we more precisely mapped the HTLV-II RxRE RNA sec-ondary structure and specific sequence requirements for efficient Rex binding and function and demonstrated the importance of both the stem-bulge-loop and splice donor sites.
Several investigators have also speculated, without pre-senting mutagenesis data, that the GUCG bulge sequences maybeimportantforHTLV-I Rexbinding andfunction(2, 34). Otherinvestigatorshave shown thatwhen the stringof nucleotides from nt 506 to 515 in HTLV-I, which contains
the GUCG element, is mutated without changing the
pre-dictedRNAsecondarystructure, bothRexfunction (1) and
binding(6) to the HTLV-I RxRE RNAarelost. In contrast, wehavedemonstrated thatadinucleotide substitutionin the analogous HTLV-II RxRE bulge sequences fromnt 466 to
469(GUCG, s467,468) did not significantly affect HTLV-II Rexbinding orfunction. Mutagenesis ofthe adjacent
stem-bulge in the HTLV-II RxRE (mutants s470,471, s470,471,
s495,496,
A470-473,
and s473,474) demonstrates that these sequencesare criticalfor Rex-RxREbinding andRex func-tion. In aggregate, these results suggest that HTLV-I and HTLV-II Rexbindingto andfunctionthrough their respec-tive RxREs require similar RNA secondary structures but differentspecific sequences.Our mapping ofthe critical regions for Rex
binding
andfunction in the HTLV-II RxRE suggests a
possible
mecha-nism for the enhancement ofgag-pol expression by Rex.
Since HTLV-II Rex requires sequences within the splice
donor site in the LTR for efficient binding toRxRE RNA,
Rex may directly compete with Ul snRNP for binding. Competitionfor binding tothe 5' LTR RxRE could poten-tially inhibit splicing ofunspliced viral mRNA and lead to
increased expression of unsplicedorgag-polviral mRNAs.
Differential recognition of env mRNA may proceed by similar recognition of alternate response elements outside
the 5' or 3' LTRs.
ACKNOWLEDGMENTS
WethankPatrick Green fortechnical assistance,William Wachs-manforproviding oligonucleotides, and Wendy Aft forher invalu-ableassistance inpreparationofthe manuscript.
A.C.B. is supported by grants from the Leukemia Society of America, the California Institute for Cancer Research, and the CancerResearch Coordinating Committee and by National
Insti-tutesofHealthgrantCA01566-01. J.D.R. issupported byNational Institutes ofHealth grants CA01314,CA 53632,and CA 52410-01
differenceinrelative CATactivitybetween anytwoprobeswasstatisticallysignificant(P<0.05,Student'sttest)whenthedifferencewas
greater than 0.20.(B)Theresultsofrepresentative Rextitrationexperimentaredepicted.CMV-CATconstructs(10,ug)werecotransfected into5 x 106Jurkat T cells withdifferentamountsofRexexpressionvector(BCrexII),asindicatedbeloweachcolumn, andacorresponding amountofBC12(control)vector wasaddedtobringthetotaltransfectedplasmidto25 ,ug. Note thatnosignificantRexresponseisseenwith theA465-501ands470,471CMV-RxRE-CATvectors;this lack ofresponsecorrespondstothe lackofsignificantRexprotein bindingtothe correspondingmutantRxRE RNA(Fig. 1A andC).
VOL. 65,1991
on November 10, 2019 by guest
http://jvi.asm.org/
andby the U.S. ArmyMedical Research and Development Com-mand.I.S.Y.C. is supported by the LeukemiaSociety of America, UniversitywideTask Force onAIDSgrantR87LA057,and National Institutesof Health grantsCA 38597,CA 01811,CA32737,and CA 43370.
REFERENCES
1. Ahmed, Y. F., S. M. Hanly, M. H. Malim, B. R. Cullen, and W.C. Greene.1990.Structure-functionanalyses oftheHTLV-I Rexand HIV-1 Rev RNA response elements: insights intothe mechanism ofRexandRevaction. GenesDev. 4:1014-1022. 2. Ballaun, C., G. K. Farrington, M. Dobrovnik, J. Rusche, J.
Hauber, and E. Bohnlein. 1991. Functionalanalysis ofhuman T-cellleukemiavirus type I rex-responseelement: directRNA binding ofRexproteincorrelates with invivoactivity.J.Virol. 65:4408-4413.
3. Bhagavati, S., G. Ehrlich, R. W. Kula, S. Kwok, J.Sninsky,V. Udani, and B. J. Poiesz. 1988. Detection of human T-cell lymphoma/leukemiavirus type I DNA andantigeninspinalfluid andblood ofpatients withchronicprogressivemyelopathy.N. Engl.J. Med. 318:1141-1147.
4. Black,A.C.,I.S. Y.Chen, S. J. Arrigo, C. T.Ruland,E.Chin, T.Allogiamento,andJ. D. Rosenblatt. 1991. Differentcis-acting regions ofthe HTLV-II 5' LTR are involved in regulation of geneexpression byRex. Virology 181:433-444.
5. Black, A. C., S. D. Nimer, I. S. Y. Chen, and J. D. Rosenblatt. 1990. Theeffects oftaxandrexproteins ofHTLV-I/-II onviral and cellulargeneregulation,p. 33-47. In J.Groopman,I. S. Y. Chen,M.Essex, and R. Weiss(ed.), UCLAsymposia: human retroviruses, vol. 119. AlanR. Liss, Inc.,NewYork. 6. Bogerd,H.P., G. L. Huckaby,Y. F.Ahmed, S. M.Hanly,and
W. C. Greene. 1991. The type 1 humanT-cell leukemia virus (HTLV-I)Rextrans-activatorbindsdirectlytotheHTLV-IRex and the type 1 human immunodeficiency virus Rev RNA re-sponseelements. Proc. Natl. Acad.Sci. USA88:5704-5708. 7. Cann, A. J., Y. Koyanagi, and I. S. Y. Chen. 1988. High
efficiency transfection of primary human lymphocytesand stud-iesofgeneexpression. Oncogene3:123-128.
8. Cann,A. J., J. D.Rosenblatt,W.Wachsman, N. P.Shah,and I. S. Y. Chen. 1985. Identification ofthe gene responsible for humanT-cell leukemia virustranscriptional regulation.Nature (London)318:571-574.
9. Chen, I. S. Y., J.McLaughlin, and D. W. Golde. 1984. Long terminal repeats of human T-cell leukemia virus II genome determinetargetcellspecificity.Nature(London)309:276-279. 10. Chen,I.S. Y.,D.J.Slamon, J. D. Rosenblatt, N.P.Shah, S. G. Quan, and W. Wachsman. 1985. The x gene is essential for HTLVreplication. Science 229:54-58.
11. Felber, B. K., H. Paskalis, C. Kleinman-Ewing, F.Wong-Staal, and G. N. Pavlakis. 1985. The pX protein of HTLV-I is a
transcriptional activator of its long terminal repeats. Science 229:675-679.
12. Fujisawa, J., M. Seiki, T. Kiyokawa, and M. Yoshida. 1985. Functional activation of the long terminal repeat of human T-cellleukemia virustype Ibyatrans-acting factor.Proc. Natl. Acad. Sci. USA82:2277-2281.
13. Gelmann,E., R. Gallo, and W. A. Haseltine. 1984. Repetitive
structurein thelong-terminal-repeatelement ofatype IIhuman T-cell leukemia virus. Proc. Natl. Acad. Sci. USA 81:4617-4621.
14. Gessain, A., F. Barin, J. C. Vernant, 0. Gout, L. Maurs, A. Calender,andG. De The.1985.Antibodiestohuman T-lympho-tropic virustype Iinpatientswith tropical spastic paraparesis. Lancet ii:407-409.
15. Grassman, R., S. Berchtold, C. Aepinus, C. Ballaun, E. Boehnlein, and B. Fleckenstein. 1991. In vitro binding of human T-cellleukemia virusrexproteinstothe rex-response element of viraltranscripts. J. Virol. 65:3721-3727.
16. Hanly, S. M., L. T. Rimsky, M. H. Malim, J. H. Kim, J. Hauber, M. D.Dodon, S.-Y. Le, J. V. Maizel, B. R. Cullen, and W. C. Greene. 1989. Comparative analysis of the HTLV-I Rex and HIV-1 Rev trans-regulatory proteins and their RNA response elements. Genes Dev. 3:1534-1544.
17. Haseltine,W.A., J.Sodroski,R.Patarca,D.Briggs,D.Perkins, and F.Wong-Staal. 1984.Structure of3'terminalregionoftype IIhuman Tlymphotropic virus: evidence ofnewcodingregion.
Science225:419-421.
18. Hikada, M., J. Inoue, M. Yoshida, and M. Seiki. 1988. Post-transcriptional regulator (rex) ofHTLV-Iinitiatesexpressionof viralstructuralproteinsbut suppressesexpressionofregulatory
proteins. EMBOJ.7:519-523.
19. Hinuma,Y., K.Nagata,M.Hanaoka,M.Nakai,T.Matsumoto, K.-I.Kinoshita,S.Shirakawa,andI.Miyoshi. 1981. Adult T-cell leukemia:antigeninanATLcell lineanddetectionofantibodies
to the antigen in human sera. Proc. Natl. Acad. Sci. USA 78:6476-6480.
20. Inoue, J.-I., M. Yoshida, andM. Seiki. 1987. Transcriptional
(p40x) andpost-transcriptional (p27xIII)regulators arerequired
for the expression and replication of human T-cell leukemia virustype I genes. Proc. Natl. Acad. Sci. USA84:3653-3657. 21. Kalyanaraman, V.S.,M.G.Sarnagadharan,M.Robert-Guroff,
I.Miyoshi,D.Blayney,D.Golde,andR.C. Gallo.1982. Anew
subtype of human T-cell leukemia virus(HTLV-II) associated with aT-cell variant ofhairy cell leukemia. Science 218:571-573.
22. Kim, J. H., P. A. Kaufman, S. M. Hanly, L. T. Rimsky, and W. C. Greene. 1991. Rex transregulation ofhuman T-cell leu-kemia virus type II geneexpression. J. Virol.65:405-414. 23. Lee, H., P. Swanson, V. S. Shorty, J. A. Zack, J. D.Rosenblatt,
and I. S. Y. Chen. 1989. High rate of HTLV-II infection in seropositive IVdrug abusersfrom New Orleans. Science 244: 471-475.
24. Mount, S. M. 1982. Acatalogue ofsplicejunction sequences. Nucleic Acids Res. 10:459-472.
25. Nakamaye, K., and F. Eckstein. 1986. Inhibition ofrestriction endonuclease NciIcleavageby phosphorothioategroupsandits applicationtooligonucleotide-derived mutagenesis.Nucleic Ac-ids Res. 14:9679-9698.
26. Ohta, M.,H.Nyunoya, H. Tanako, T. Okamoto, T.Akagi, and K. Shimotohno. 1988. Identification ofacis-regulatoryelement involved inaccumulationofhumanT-cell leukemia virustypeII genomic mRNA. J. Virol. 62:4445-4449.
27. Ohtani, K., M. Nakamura, S. Saito, T. Noda, Y. Ito, K.
Sugamura, and Y. Hinuma. 1987. Identification oftwo distinct elements in thelong terminalrepeatofHTLV-Iresponsiblefor maximum geneexpression. EMBOJ. 6:389-395.
28. Poiesz, B. J., F. W. Ruscetti, A. F. Gazdar, P. A. Bunn, J. D. Minna, andR.C. Gallo. 1980.Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes ofa patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. USA 77:7415-7419.
29. Robert-Guroff, M., S. H. Weiss, J. A. Giron, A. M. Jennings, H. M.Ginzburg,I.B.Margolis,W.A.Blattner, and R. C.Gallo. 1986. Prevalence of antibodies to HTLV-I, -II, and -III in intravenousdrugabusersfrom an AIDS endemic region. JAMA 255:3133-3137.
30. Rosenblatt, J. D., A. J. Cann, D. J. Slamon, I. S. Smalberg, N. P. Shah, J. Fujii, W. Wachsman, and I. S. Y. Chen. 1988. HTLV-II trans-activation is regulatedby two overlapping
non-structural genes. Science 240:916-919.
31. Rosenblatt, J. D., D. W. Golde, W. Wachsman, A. Jacobs, G. Schmidt, S. Quan, J. C. Gasson, and I. S. Y. Chen. 1986. A second HTLV-II isolate associated with atypical hairy-cell leukemia. N. Engl. J. Med. 315:372-375.
32. Sodroski, J. G., C. A. Rosen, and W. A. Haseltine. 1984. Trans-acting transcriptional activation of the long terminal re-peat ofhuman Tlymphotropic viruses in infected cells. Science 225:381-385.
33. Tedder, R. S., D. C. Shanson, D. J. Jeffries, R. Cheingsong-Popov, A. Dalgleish, P. Clapham, K. Nagy, and R. A. Weiss. 1984. Low prevalence in the UK of HTLV-I and HTLV-II infection insubjects withAIDS, with extended lymphadenopa-thy, andat riskofAIDS. Lancet ii:125-128.
34. Toyoshima, H., M. Itoh, J.-I. Inoue, M. Seiki, F. Takaku, and M. Yoshida. 1990. Secondary structure of the human T-cell leukemia virus type I Rex-responsive element is essential for
on November 10, 2019 by guest
http://jvi.asm.org/
HTLV-II Rex FUNCTION AND BINDING 6653 Rex regulation of RNA processing and transport ofunspliced
RNAs. J. Virol. 64:2825-2832.
35. Wachsman, W., K. Shimotohno, S. C. Clark, D.W.Golde, and
I. S. Y. Chen. 1984. Expression of the 3' terminal region of
human T-cell leukemia viruses. Science 226:177-179.
36. Wong-Staal, F., B. Hahn, V. Manzari,S. Colombini, G. Fran-chini, E. P. Gelmann, and R. C. Gallo. 1983. Asurveyof human leukemias forsequences ofahuman retrovirus. Nature (Lon-don)302:626-628.
37. Yip, M. T., W. S. Dynan, P. L. Green,A. C. Black,S.J. Arrigo, A.Torbati,S. Heaphy, C. Ruland,J. D. Rosenblatt, andI.S.Y.
Chen. 1991. Human T-cell leukemia virus(HTLV) typeIIRex
protein binds specificallytoRNAsequencesof the HTLV long terminalrepeatbut poorlytothe human immunodeficiency virus
type1Rev-responsive element. J. Virol. 65:2261-2272. 38. Yoshida, M., I. Miyoshi, and Y. Hinuma. 1982. Isolation and
characterization ofretrovirus from cell lines of human adult T-cell leukemia and its implication in the disease. Proc. Natl. Acad. Sci. USA79:2031-2035.
39. Yoshida, M., M.Seiki, K. Yamaguchi, and K. Takatsuki. 1984. Monoclonal integration of human T-cell leukemia provirus in all primarytumorsof adult T-cell leukemiasuggestscausative role of human T-cell leukemia virus in the disease. Proc.Natl. Acad. Sci.USA 81:2534-2537.
40. Zuker, M.,and P. Stiegler. 1981. Optimalcomputerfoldingof large RNA sequences using thermodynamics and auxiliary information. Nucleic Acids Res. 9:133-148.
VOL. 65, 1991