0022-538X/93/010150-11$02.00/0
Copyright ©1993,American SocietyforMicrobiology
Functional Role of the
Glycan Cluster
of the
Human
Immunodeficiency
Virus
Type
1
Transmembrane
Glycoprotein (gp4l) Ectodomain
E.
FENOUILLET,l* I. JONES,2 B. POWELL,2 D.SCHMITT,3
M. P. KIENY,3ANDJ. C. GLUCKMAN' Laboratoire de BiologieetGe6ne'tique desPathologies Immunitaires, CNRS URA 1463,83 boulevard de l'H6pital, 75651 Paris Cedex 13,1 and Transgene SA, 67000 Strasbourg,3 France, and NERC Institute ofVirology, Oxford OX1 3SR, Great Britain2
Received 8 May1992/Accepted 20 October 1992
To examine the role of the glycans of human immunodeficiencyvirustype 1 transmembraneglycoprotein gp4l, conserved glycosylation sites within theenv sequence(Asn-621, Asn-630,andAsn-642)weremutatedto Gln. The mutated and controlwild-typeenvgeneswereintroduced into recombinant vaccinia virus andused to infect BHK-21 or CD4+ CEM cells. Mutated gp4l appeared as a 35-kDa band in a Western blot (immunoblot), and it comigrated with the deglycosylatedform ofwild-type gp4l. Proteolytic cleavage of the recombinant wild-type and mutant forms of the gpl60 envelope glycoprotein precursor was analyzed by pulse-chase experiments and enzyme-linked immunosorbentassay:gpl60synthesiswassimilarwhether cells wereinfected with controlormutatedenv-expressing recombinant vaccinia virus,but about 10-fold less cleaved
gp120and gp4l wasproduced by the mutated constructthan the controlconstruct.The rates ofgpl20-gp41 cleavageateach ofthe twopotential sites appearedtobecomparableinthe twoconstructs. By usingapanel ofantibodies specific forgp4l andgpl20epitopes, itwasshown that the overall immunoreactivitiesof control and mutatedgp4l proteins were similar butthat reactivitytoepitopes atthe C and N termini ofgpl20, as presentongpl60producedby the mutated construct,wasenhanced. Thiswas nolonger observed for cleaved gpl20 in supernatants. Bothgpl20proteins, from control and mutatedenv, wereexpressedonthe cellsurface undera cleaved form and could bind to membraneCD4,asdeterminedby quantitativeimmunofluorescence
assay.Incontrast, anddespite sufficient expressionofenv productsat thecellmembrane, gp4l produced by
themutated constructwasunable toinduce membrane fusion.Therefore, while contradictoryresultsreported in the literature suggest that gp4l individual glycosylation sites are dispensable for the bioactivity and conformation ofenv products, it appears that such is not the case when the whole gp4l glycan cluster is removed.
Thematureenvelope glycoproteins of human immunode-ficiency virus type 1 (HIV-1)-outer membrane gpl20 and transmembrane gp4l-are proteolytic cleavage productsof
precursorgpl60, andtheyareresponsible for virusbinding
to CD4+ cells and fusion of virus and cell membranes, respectively (for reviews, seereferences 13 and 23). Previ-ous results indicate that afterbiosynthesis, glycans present on mature recombinant or viral gpl20 and gp4l are not directly involved in thebiological activity of these molecules (9, 12). Incontrast,by usinganinhibitor of a-glucosidases-1-deoxynojirimycin-which induces abnormal glycosyla-tion,we have recentlyshown thatat least anumber of the approximately 30 glycans (16, 26) ofgpl20contribute, during biosynthesis, to the folding, conformation (10, 19), and bioactivity (10) of nascent recombinant gpl60. This effect was due to the presence on gpl20 of 1-deoxynojirimycin-inducedabnormalhigh-mannose-type glycans with high
mo-lecularmassandnot tothat ofhigh-mannose glycansper se, inasmuchasbaculovirus-produced gpl20wasperfectly able tobindto CD4(10, 32).
The role ofgp4lglycosylation during biosynthesis could notbe studied by this approach. While 1-deoxynojirimycin affectsasufficientproportionof thenumerousgpl20glycans toinduce adetectable effect, its incomplete efficiency fora given glycoprotein (7) makes it inappropriate for
examina-* Corresponding author.
tion of the role of the limitednumberofglycansongp4l (4, 15): gp4l synthesis in the presence of 1-deoxynojirimycin would result in molecules of which ahigh proportion were normallyglycosylated, and the properties of these species would then mask those of the abnormally glycosylated species.
The ectodomain of gp4l of
HIV-1j_Aj
(41) presents five putative glycosylation sites (4), atleast three of which are glycosylated (12). A 30-amino-acid-long peptide (residues 615 to 645), glycosylation of which is conserved among HIV-1, HIV-2, and simian immunodeficiency virus (4),ap-pears to have features similar to those of transmembrane
glycoproteins of other lentiviruses (15). These sites are located in a region that corresponds to a variable, weak neutralizing epitope, and theyareclosetothegp4l immuno-dominant region and to a putative domain involved in the noncovalent binding of gp4l to gpl20 (for a review, see reference 15). Accordingto currentmodels (15), they might also be spatially close to the gp4l cleavage site and fusion peptide(14); therefore, the hydrophilicity of the correspond-ing glycansmaycontributetothe"hiding" of this hydropho-bicregionbeforeit isexposedtoinitiate membrane fusion.
Recently, apparently conflicting results have been ob-tainedbytwoindependentgroupswhich examinedthe role ofgp4l glycans by using single glycosylation site mutation. Dederaetal.(3)"demonstrate(d) that individual conserved potential N-glycosylation sites are dispensable for
produc-150
on November 9, 2019 by guest
http://jvi.asm.org/
ROLE OF gp41 GLYCANS 151
tion of infectious, cytopathic virus." Effectively, removal of individual glycosylation sites fromgp4i modified neither the cytopathic effect of an HXB2 clone nor reverse transcriptase activity. Nevertheless, of the three single potential glycos-ylation sites removed, mutation ofAsn-618 had the greatest effect on syncytium formation for CEM cells (fivefold reduc-tion) but not for SupTicells. In the same study, a vaccinia virus(VV)vectorharboring the same env mutant exhibited a reduced(16-fold with CEM, 9-fold with SupTicells) butstill patentability to induce syncytia and "no significant effects on envelope protein synthesis, conformation,
process-ing
.... or release into the culture medium" were observed. In contrast, by using the same virus clone and Sup Ticells, Lee et al. (25) showed that mutation of Asn-642 caused areduction in HXB2 infectivity (as determined by delayed reverse transcriptase production kinetics but not as a re-duced tissue culture infective dose), which contrasted with the absence of an effect of this mutation noted in the previous study. Surprisingly, in this work, removal of the glycosylation site corresponding to Asn-618 increased HXB2 infectivity. Therefore, the extent to which any single glycos-ylation site affects env bioactivity remains uncertain and appears tobehighly dependent on the experimental condi-tions. It therefore remains possible that single gp4i glycos-ylation sites are dispensable for HIV-1 infectivity. This is not
surprising when one considers the literature which shows that, in contrast to viralglycoprotein multiple glycosylation sitemutations, single glycosylation site removal usually does notaffect viral glycoprotein bioactivity (5, 24, 27, 31).
For these reasons, we mutated the
gp4i
cluster of the three glycosylation sites that are conserved among HIV-1, HIV-2, and simian immunodeficiency virus (4). Processing andcleavage of therecombinant mutated glycoprotein were examined andcompared with those of the wild-type recom-binantglycoprotein, both expressed by a VV vector. Con-formation, presentation at the cell surface, and biological activity of bothgp4i
and gp120, as wellastheir noncovalentassociation, wereevaluated.
MATERIALSANDMETHODS
Mutation of the gpl60-encoding gene and generation of recombinant VV. The
HIV-li_AI
env sequence used (clone VVTG9-1) (21) was from Transgene SA (Strasbourg, France).Agpl60 genelackingthe threeconservedcarbohy-drate additionsitesatpositions7663, 7690, and 7721 in gp41
(correspondingtoAsnresidues locatedatpositions 621, 630,
and642,respectively)wasconstructedbypolymerasechain reactiongeneration of twoenv fragments incorporating the
carbohydrate changes, followed by ligation to reconstruct the full gene.
The first polymerase chain reaction used the forward
primer
5'-CCGGGAGCTCATGAGAGTGAAGGAGAAA-3' and thereverseprimer 5'-GGTCATGGCATTCCAAATC
TGTTCCAGAGA'l''l'GACTCCAACTAGCGTTCCA-3'
togeneratea1,900-bpfragment from the 5'end of thegpi60
gene flanked by unique restriction sites Sacl at the 5' end and BsmIatthe 3' end. Thereverseprimeralsoincorporated
base changes to change the Asn of the most upstream
carbohydratesiteatposition7663toGln andremove asingle
internal BsmI sitebyathird basechange that didnotaffect the amino acid sequence.
In the second polymerase chain reaction, the forward primer
5'-AT'l-GGAATGCCATGACCTGGATGGAGTGG
GACAGAGAAATTAACCAATACAC-3' and the reverse
primer
5'-CCGGTCTAGATTATAGCAAAATCC'l'lTCCA
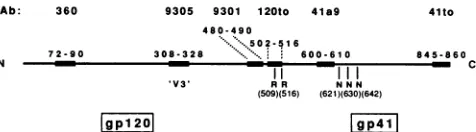
Ab: 360
N 72-90
9305 9301 120to 41a9
480-490 502-5 6
308-328 ***.... 600-61 0
[gpl 201
I I I I
RR NNN
(509)(516) (621)(630)(642)
[gp4l
41to
845-8 60 C
FIG. 1. Schematic drawing of gp160 indicating the epitopes recognized by the different Abs used in this study. Locations of the potential cleavage sites and conserved glycosylation sites are indi-cated. Numbersrefertoaminoacid residues.
A-3'wereused to generate a 720-bp fragment from the 3' end of thegpi60geneflanked by unique restriction sites BsmI at the 5' end andXbaI atthe 3' end. The forwardprimer also incorporated base changes to mutate Asn to Gln in the two downstream carbohydrate sites at positions 7690 and 7727.
Inboth cases, theamplification conditions were 10 ng of template DNA (a full-length env clone of
HIV-1LAI),
100 ng of eachprimer, and 25 cycles at94°C for 1min,
50°C for 1 min, and 72°C for 2 min. DNA fragments generated by polymerase chain reaction were gel eluted, digested with BsmI, and ligated for 2 h at room temperature. The ligation reaction was inactivated by heating, and the resulting frag-ment was recutwithSacI andXbaIand clonedinto pUC118 betweenthe Sacl and XbaI sites. The structure of the final genewasconfirmed by restriction and sequenceanalyses.The mutated env gene was introduced into plasmid pTG186POLY by allowing homologous recombination with the VV genome. Recombinant VV (rVV)wasgeneratedby using the protocol described in reference 20.
Productionof recombinantenvglycoproteins. BHK-21 cells wereinfectedasdescribed in references 10 and21, withrVV
expressing the wild-type (VVTG9-1 [21]; referred to as
VV9-1 thereafter) or mutant (VVO) env gene. Cells were cultured in Glasgow medium (J. Bio, Les Ullis, France),
which contained1% glutamine, 5% fetal calf serum, and5%
tryptose phosphate, as previously described (10, 21). They
wereinfected with eitherVV9-1orVVO (0.2PFU percell)in fetal calfserum-free mediumto allow easy further superna-tant concentration and analysis. After 2 days, cell pellets
werelysedwithphosphate-bufferedsaline(PBS)-2%Triton X-100 (Sigma, St. Louis, Mo.) andsupernatants were con-centrated fivefold byusing a 10-kDa cutoffCentriprep sys-tem (10) (Amicon, Danvers,Mass.).
Antibodies. Sheeppolyclonal immunoglobulinG antibody (Ab) D7324 (referred to thereafter asAbl20to; Aalto, Dub-lin, Ireland) recognizes a peptide encompassing the two
gpl60 potentialcleavage sites(aminoacids[aa]502to516)at the gpl20 C terminus; sheep Ab D7323
(Ab4lto;
Aalto) recognizes aC-terminal peptideofgp4l
(aa845 to860) (11) (Fig. 1).Monoclonal antibody (MAb) mAb360 (a gift from H. Holmes, MRC AIDS Reagent Project, Potters Bar, United
Kingdom) is a mouse MAb that
recognizes
thegpl20
N terminus(aa72 to90). mAb9305 andmAb9301(Du
Pont de Nemours, Dreieich,Germany) areMAbs directed to aa318 to328(inthe V3 loopofgpl20)
and 480to490(C
terminus ofgpl20),
respectively. mAbllO-4(Genetic Systems,
Seat-tle,Wash.)is another anti-V3loopAb.mAb41a9
(a gift
from F. Traincard, Hybridolab, Institut Pasteur,Paris,
France)
and mAb41-1 (Genetic Systems) are MAbs directed to the immunodominantregionof
gp4l
(aa602to610) (11;
Fig.
1).
Westernblot(immunoblot;WB)characterization of
recom-binantglycoproteins.Two days
postinfection
with VV9-1 or VOL. 67,1993on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.317.555.77.143.2]VVO,
celllysates
were nottreatedortreatedovernight
with 200 mU ofanendoglycosidase
FN-glycanase
mixture(endo
FN-glycanase;
Boehringer, Mannheim, Germany),
aspre-viously
described(10, 12),
and submitted to 10% sodiumdodecyl
sulfate(SDS)-polyacrylamide
gel electrophoresis
(PAGE).
Aftermigration, blotting
wasperformed
asprevi-ously
described(10, 11, 12)
andstaining
wasrealized with human anti-HIV-1 AbAbl20toorAb41to(diluted 1:1,000)
or mAb9305(diluted 1:100).
Afterovernight
incubationat20°C
(8), peroxidase-coupled anti-sheep,
-mouse, or-human Absdiluted
1:1,000
(Dakopatts, Glostrup, Denmark)
wereadded for 2 hand colorwasdeveloped by using
diaminobenzidine. HIV-1 viralpellets
(Diagnostics Pasteur,
Marne-la-Co-quette,France),
notdeglycosylated
ordeglycosylated
aspreviously
described(12),
were studied inparallel
under identicalconditions.Immunoreactivity
anddosage
ofrecombinantglycoproteins
by enzyme-linked
immunosorbent assay(ELISA).
Microtiterplates
(Nunc, Roskilde, Denmark)
were coated withAbl20to
orAb41to(200 ng/100 ,ul)
in 20 mM sodium bicar-bonatebuffer,
pH
8.After 2 h at20°C,
plates
wereblocked with5% casein.Increasing
amountsofcelllysates
or super-natantswereadded for 3 hat20°C
afterdilutioninPBS-1% Tween-1%casein, pH
7.4. Aftertwowashes,
mouse MAbsora
pool
ofhumananti-HIV-1Abs(diluted
1:100)
wasadded for 3 hat20°C
in SS buffer(supplemented
with10%normalsheep serum)
that was added to inhibitnonspecific
Abbinding
aspreviously
described(29).
After twowashes,
anti-mouse or anti-human Abscoupled
toperoxidase
(1:
1,000
dilutionin SSbuffer)
wasadded and incubated for 2 h at20°C.
Color wasdeveloped
with theappropriate
chro-mogen after five washes.Metabolic
labeling
andpulse-chase
experiments. Fifteenhours afterrVVinfectionof 10 x 106 BHK-21
cells,
3 ml oflow-Met-Cys
Glasgow
medium(J. Bio)
was addedasprevi-ously
described(21).
After 1h,
150,uCi
of radiolabeled[35S]Met-Cys (Trans
Label;
ICN,
CostaMesa,
Calif.)
permlwas added and incubated for 30 min. The supernatant was then
removed,
cells werewashed,
andMet-supplemented
(100 ,ug/ml) Glasgow
mediumwasadded and incubated for1,
4,
or 18h. Cellsand supernatantswereseparated
by
centrif-ugation immediately
after thepulse
or after each chaseperiod.
Cells werelysed
with 25 mM Tris-300 mMNaCl-0.25% SDS-0.5%Triton
X-100, pH
7. Recombinantglyco-proteins
in celllysates
orsupernatantswere immunoprecip-itatedby
anAIDSpatient's
serumdiluted 1:100 andproteinA-Sepharose
(Pharmacia,
Uppsala,
Sweden)
aspreviously
described(21).
After fivewashes in2mlofbuffer,
recombi-nantglycoproteins
were elutedby
using
10% SDS-5%3-mercaptoethanol and heating (80°C, 5
min).
Eluateswereanalyzed by
SDS-PAGE andautofluorography
aspreviously
described(21).
Localizationof
gpl60
cleavagesites. WeutilizedanELISA toassesstherateofgpl60
proteolytic cleavage
ateither of its twopotential
cleavage
sites. Thiswas basedon thefinding
that aminority
ofgp4l
molecules react withAbl20to,
apolyclonal
antibody
whichpartly
recognizesthe amino acid sequence locatedbetween the twopotentialcleavage sites, an indication that these molecules are cleaved at Arg-509(11)
instead ofArg-516 (1,
11, 18, 28).The amount of such
gp4l
species
relative to the total amount ofgp4l
produced
was evaluated here as follows.Two
days
after rVV infection of BHK-21 cells, virus and membranepellets
obtained afterultracentrifugation (100,000 x g for 1h)
of the cleared supernatant were lysed by 2%Triton.
Only
gpl20
andgp4l,
andnogpl60,
weredetected inthelysate by WB and by ELISA with Ab4lto-coatedplates
andmAb9305staining (datanotshown). Increasingamounts oflysates diluted in PBS-1% Tween-1% casein,were incu-bated in Ab4lto- or Abl20to-coated plates as described above. Thegp4l boundtoAb41toorAbl20towasstainedas already described, with mAb41a9 and peroxidase-coupled
anti-mouse Ab in SS buffer.
Flowcytometryanalysis.(i) Binding ofgpl20toCD4+ cells. Humanlymphoid CD4+ CEM13 cellswerecultured in RPMI 1640 (J. Bio)-10% fetal calf serum-1% glutamine-1%
peni-cillin-streptomycin as previously described (9, 10, 12). In-creasing amountsof concentrated rVV-infected cell super-natants, which contained solublegpl20, were incubated with 106cells for 2 hat20°C in 100 ,ul of PBS-0.5% bovineserum albumin-0.05% NaN3 as previously described (9, 10, 12).
Labeling with a pool of anti-HIV-1 human sera or with mAb9305 diluted 1:100 was performed for 1 h at 20°C. Phycoerythrin-coupled sheep human or rabbit anti-mouse immunoglobulin G Abs diluted 1:100 (Becton Dick-inson, San Jose, Calif.) were added for 1 h at 20°C (9, 10,12).
Membrane fluorescence was measured with a FACScan apparatus(Becton-Dickinson).
(ii) Expression of gpl20 and gp4l on the surface of rVV-infected cells. At4, 8, or 24 h postinfection with VV9-1 or VVO (10 PFU percell), 106 CEM13 cells were incubated for 2 hat20°C withmAb9305, mAbllO-4,mAb41a9,ormAb41-1 diluted 1:100 to detect the presence of recombinant
glyco-proteins at the cell surface. Staining was performed as described above.
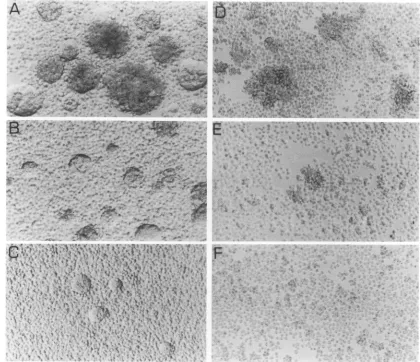
Syncytium formation assay. CEM13 cells (5 x 106) were infected withincreasing rVV doses (1, 10, and 100 PFU per cell) in RPMI 1640-1%glutamine-10% fetal calf serum. At 1 daypostinfection, cultureswerephotographed and syncytia oraggregateswerescored. The culturewasmonitored foran additional 2 days to observe the evolution of syncytium development in the different samples.
RESULTS
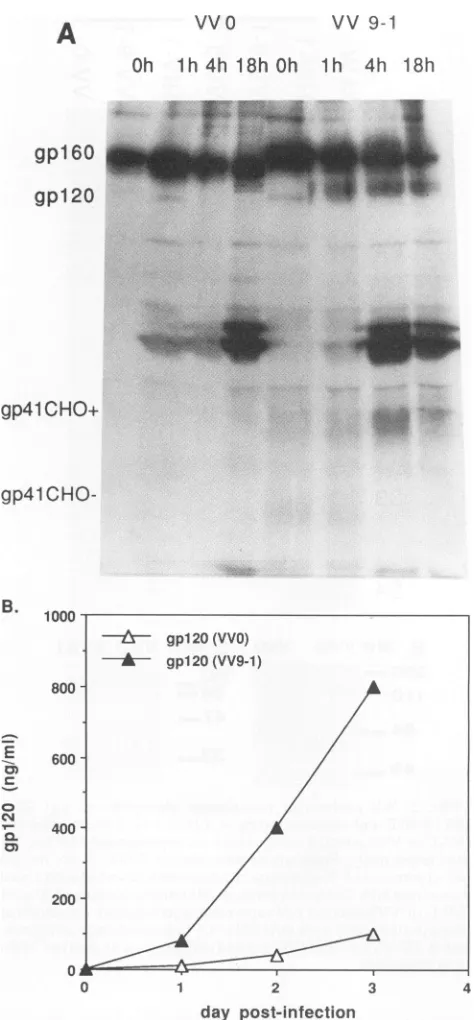
Apparent molecular weights (MWs) of recombinant env HIV-1 glycoproteins. First we used WB to determine the MWs of the recombinant molecules obtained following ex-pression in rVV, and then we examined whether gp4l
produced by mutated env (gp41CHO-) was glycosylated relative to its wild-type counterpart (gp41CHO+). (i) As expected (11), gp4l, gpl20, and lesser amounts of gpl60 weredetectable inVV9-1-infected BHK-21 celllysates (Fig.
2A), and secreted gpl20 was detected in the respective supernatants (Fig. 2B). (ii) When lysates of106 VV9-1- or VVO-infected cellswereimmunoblottedwithAb41to, similar amountsofgpl60 precursorweredetected while gp41CHO-was not (Fig. 2C); the latter was readily detected only by
usinglysatesfrom 10timesmorecells (Fig. 2A).
Under the latterconditions (Fig. 2A), ahigh-MW (about
140,000) glycoprotein was found in VVO-infected cell ly-sates. The decrease of its MW to 90,000 after endo F
N-glycanase(40) deglycosylation,aspreviously observed (9,
10, 12), indicated that this protein corresponded to the precursor envelope glycoprotein. No gpl20 species was
distinguishedhere, presumably because of superimposition of the 140,000-MWband.
A 35,000-MW band should correspond to gp41CHO-, because it also specifically reacted with Ab41to (data not
shown) and its MW agreed with that of both endo F
N-glycanase-treatedviralgp4l (12) andwild-type recombi-nantgp41CHO+. Moreover, endo F N-glycanase treatment
on November 9, 2019 by guest
http://jvi.asm.org/
ROLE OF gp4l GLYCANS 153
6
CD
A
z
*:
-M>>
~~~~~I
1~~~~.
_.10
i
J_
84
47
(5
z
I>>I
I >F >
10 100 1 10 100
supernatant added(pi)
FIG. 3. Determination of glycoproteins in rW-infected cells and supernatants by ELISA. (A) Comparison of cell-associated gp160 and gp4l. Ab41to-coated wells were incubated with increasing volumes of W9-1- orW0-infectedcelllysates.mAb41a9 was used todetect both capturedgpl60andgp4l,and mAb9305 was used to detectgpl60only. (B) Comparison of amounts ofgpl20in superna-tants.Abl20to-coatedwells were incubated with increasing volumes ofW9-1- orW0-infected cell supematants. The captured glyco-protein wasdetected with mAb9305. Control optical densities de-termined withnoninfected BHK-21 celllysatesor supematants were approximately0.02. All assays wereperformed three to five times with reproducible results.
vvo C MW VVO VV9.1
1o0
84- 47-
33-FIG. 2. WB analysis of recombinant glycoproteins. (A) SDS-(10%) PAGE and immunoblotting ofaHIV-1 viral preparation and
W9-1-orWO-infectedcelllysates from approximately 106 and107 cells, respectively. Preparationsweretreated (NG+)ornottreated (NG-)withendoFN-glycanase. Proteinsweredetected withapool ofserafrom HIV-1-infectedpatients. (B) Immunoblottingof 20Ilof W9-1-orWO-infectedcell supernatants (concentrated fivefold)at
3days postinfectionwith mAb9305.(C) Immunoblottingof
approx-imately106W9-1-andWO-infected cell lysates with Ab41to. MWs
areinthousands.
ofgp41CHO- did notresult inmodification of its MW, an indication that no N-linked glycans were present on the molecule.
Relative amounts ofgpl20and gp4l in rW-infected cells and supernatants. We verified that Ab41to, Abl20to, mAb41a9, mAb41-1, mAb9305, and mAbllO-4 recognized WO- or W9-1-synthesized glycoproteins to the same ex-tent. Results ofquantitativedotblotassays(datanotshown) performed as previously described (11), under conditions that abolishconformation-dependent antibody binding,were consistent with those of the ELISA described here,which was performed under liquid-phase conditions that allow
conformation-dependent binding. One may then conclude thattheaccessibilityof theepitopes recognized by these Abs was notaffected by mutations.
The respective amounts of recombinant gp4l and gpl20 that were recovered in cells and supernatants after rVV infection were then assessed by capture ELISA. (i) First, plates were coated with Ab41to (Ab directed against the gp4l C terminus). When the captured molecules were then labeled with mAb9305(anti-V3 loopMAb), which permitted detection of gpl60, there were approximately similar amountsofgpl60 inlysates of cells infected with either the controlorthemutant construct(Fig. 3A).In contrast,when the captured molecules were labeled with mAb4la9 (anti-gp4l MAb), 7 to 10 times more of the VVO-infected cell pelletwasnecessarytoobtain thesamesignalaswith VV9-1 (Fig. 3A). Although under the conditions used here, detec-tion of precursorgpl60, which is present in similaramounts in cells infected with both rVVs, maypotentially interfere with dosage of cleaved cell-stored gp4l, it nonetheless appears that approximately 10 times less gp41CHO- than
gp41CHO+waspresentin thecorrespondingcelllysates.(ii) Similarly, when Abl20to (directed against the gpl20 C
terminus) was used for capture, along with labeling with mAb9305, 10 times more supernatant from VVO-infected cellswas necessary to obtain the same signal as with the
VV9-1-infected cell supematant
(Fig.
3B). One may then assume that the amount of gpl20 in the VVO-infected cellsupernatantwas also about 10-fold lower than in the VV9-1-infected cell supematant.
These results should be takentogetherwith the WB data ofFig. 2,inwhich thestaining intensityoftheenvprecursor observed whenlysatesof 10 VVO-infected cellswere ana-lyzedwasapproximately10-fold stronger than that noted for
106W9-1-infectedcells,whereas under thesameconditions
gp41CHO+ stainingwas similartothat of
gp4lCHO-.
Thus, it appears that the amount of cleaved gp41 andgpl20 produced by VVO-infected cells was
approximately
10-fold lower than that in VV9-1 infectedcells,
while the amounts ofgpl60producedinthe two cases wereapproxi-matelysimilar.
Cleavage of gpl60 and kinetics of gpl20 release in cell
9 gpl2O (VV0) * 9pl2O(VV9-1)
33
24
B
Mw
VV9L
200-110- 4 *
84_
49-VOL.67, 1993
.01
la
:-I 1.,
M-WIP .. IF
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.318.557.74.189.2] [image:4.612.83.267.77.488.2]supernatants. (i) gpl60 cleavage. Approximately similar amounts of radioactivitywere incorporated into gp160 ob-tained fromW9-1- orWO-infected BHK-21 cells, but the
proteolytic cleavage kinetics of the two species differed
dramatically, as indicated by the respective amounts of
gp120 noted afterapulse-chase experiment (Fig. 4A). After a 1-h chase,
significant
amounts of radiolabeled gpl20 and lesseramountsof gp41 (theweaklabelingofwhichwasdue tothe fewMet andCysaminoacids presentonthe molecule[4])
weredetected inW9-1-infected cells, and thispersisted atleast for the 18-h chase. Far less and transient (only 1 h postpulse)
cell-associated gpl20was seenin WO-infected cells. A longer exposure time, however, showed a weakgpl20band aftera4-h chase (datanotshown). Thesegp120
species migrated
faster in SDS-PAGE (similarto thegp120species
observed at time zero and at 1 hin W9-1-infectedcells),
owing to a different glycosylation pattern, asdeter-mined by endoglycosidase H sensitivity (40). The W9-1
gp120
species with increased electrophoretic mobilitycorre-spond
to species the high mannose chains of which havebeen trimmed by glycosidases but not replaced by
high-er-MW
complex
type chains by glycosyltransferases (Fig. 4A, W9-1, 1 h,where bothspecies coexist).This is in line with acquisition ofendoglycosidase H resistance (data notshown)
andwiththe resultspreviously obtained with otherexpression
systems for HIV-1 and HIV-2 envelope glyco-proteins and discussed(2, 42).These results confirm the quantitative ELISA data(Fig.
3B)
regarding respectiveamountsof cleavedgp120andgp4l produced by the rWconstructs. Also, the apparently re-duced MW of gpl60 obtained after VVO infection was consistentwiththe WB data ofFig. 2, and part of this MW difference may be accounted forby lackofgp4lglycosyla-tion.
(ii) Quantitation ofglycoproteins produced. The
quantita-tive ELISAwasused inparallel todeterminegpl20release kinetics inthe supernatant ofrW-infected cells,takingas a referencetheplotobtained withasolublegp120batch(a gift
of the Medical Research Council AIDS Reagent Program)
withaknown concentration. This showed that after 3 days, 106 W9-1-infected cells secreted approximately 800 ng of
gp120,
compared with100 ng afterWOinfection (Fig. 4B).In
addition,
asemiquantitative
dot blot assay(11)allowed us to evaluate the amount of both cleaved and uncleaved cell-associatedgp41,with mAb41a9andrecombinantgpl60WTG-1163(11, 21)as areference. Thisapproachwastaken because gpl60 from VVTG-1163 does not express the
epitope recognized by
Ab41to(12a), whichmakes itimpos-sibletouseinacaptureELISA. We thus determinedthatat 2
days
post rWinfection, there were approximately 1 ,g and 200ngofW9-1- andWO-produced
gp41 and/orgpl60,respectively, per 107cells.
Rate ofgp160 cleavageateach ofits potential sites. Wehave
recently shown that, normally, about 10 to 20% of both recombinant and viral
gp4l
molecules are recognized byAbl20to in an ELISA and an immunoprecipitation assay. Thisindicates that these species express at least part of the
epitope recognized bythe Ab and, thus, that they should be cleaved atArg-509instead of Arg-516 for the rest
(approxi-mately 80 to 90%) of the gp4l species (11). Here we
investigated
byELISAwhether reduced cleavage of WO-produced gpl60is relatedtodifferent rates at either of these sites and whether cleavage at one site is suppressed or enhanced relativetothewild-typeproduct.In agreement with our previous data, after capture with
Ab41toand
subsequent
detectionwithmAb41a9,theamountA
VV
0
vv9-1
Oh
1h
4h18h
Oh1h
4h 18hgpl
60
gpl 20
gp4l
CHO+
gp4lCHO-B. 1000
800
E
0
erJ
a-cm
600
400
200
0 1 2 3 4
day post-infection
FIG. 4. Glycoprotein cleavage and kinetics of production. (A) Recombinantgpl60cleavage kinetics. Cells (3 x 106)were pulse-labeled(150 ,uCi/ml)at 18 h after infection with 0.2 PFU ofWOor W9-1 per cell. After 30 min, the cells werechased for the lengths of time indicated. After cell pellet lysis, glycoproteins were immunopu-rified and analyzed by SDS-8% PAGE. MWs are in thousands. (B) Kinetics of gpl20 appearance in cell supernatants. Cells were infected with WO or W9-1 as for panel A. At 1, 2, or 3 days postinfection, the amounts of gpl20 in the supematants were quantified byELISA.
'-I,
", M.7
i'O
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.320.557.93.603.2]ROLE OF gp4l GLYCANS 155 3
0) 2
C.
Cu co
0
(0
0
co
n1
1 10
3
0
2-C)
C.
Cu
co:
co
.0
0
C: <
1-100 1 10 100
[image:6.612.56.298.76.289.2]supernatant added (p1)
FIG. 5. Determinationof gp160 proteolytic cleavage rate at each potentialsite.Abl2Oto-orAb4lto-coatedwells were incubated with increasingvolumes of pellet lysatesobtained after ultracentrifuga-tion of VV9-1- or VVO-infected cell supernatants. The captured glycoproteinwasdetectedwithmAb4la9.Forexample,gp4l(VVO)/ 41to stands for theVVO-producedgp4l speciescaptured by Ab41to.
[image:6.612.314.555.76.289.2]supernatant added (p1)
FIG. 6. Comparison of mAb9301 or mAb360 binding to recom-binantglycoproteins as determined by ELISA.Ab4lto-coatedwells wereincubated withincreasing volumes ofVV9-1-orVVO-infected cell lysates. Thecaptured glycoproteinwasdetectedwith mAb360 or mAb9301. For example,gp160(VV0)/9301 stands for the VVO-producedgp160speciesdetectedby mAb9301.
ofgp41CHO- detected in cell lysateswasapproximately 10 times lower than that ofgp41CHO+ (Fig. 5). Nosignificant amount of precursor gpl60 was detected with mAb9305 in membrane and virus pellets of the cleared supernatants. When the molecules were captured withAbl20to, approxi-mately 10 to 20% of gp41CHO+, as well as gp4lCHO-,
boundtoAb4lto, aspreviously described (11) (Fig. 5). Thus, the relative percentages of molecules cleaved at eachpotentialsiteweresimilar forthe twoconstructs.
Immunoreactivityof recombinantgpl60, gpl20, andgp4l.
Keeping in mind that the avidities of binding of Abl20to, Ab41to, mAb9305, and mAb41a9 to VV9-1- and
VVO-pro-ducedglycoproteins were similarand, thus, that the
corre-sponding epitopeswereexposedto similarextentsonboth, wefurther evaluated theimmunoreactivityoftwoadditional MAbs by ELISA: mAb360 (directed against the gpl20 N
terminus)andmAb9301(directed against thegp120C termi-nus 10 aaupstream of thepeptiderecognized byAbl20to). Capture Abswere eitherAb41to (which allowedfor detec-tion ofonlyprecursorgp160)orAb12Oto(acondition under which bothgp160andgp120weredetectable).BothmAb360 and mAb9301 displayed increased reactivityto cell-associ-ated mutcell-associ-ated VVO glycoproteins, by more than 10-fold relative to VV9-1 (Fig. 6), whereas they recognized gpl20
present insupernatantsof cellsinfected with eitherconstruct after captureby Abl20totothesamelimitedextent(datanot
shown).
This indicates that thesegpl20N-andC-terminalepitopes
are particularlywell exposed when expressed on mutated uncleaved precursor gpl60 but that the reactivities of the mutant and the wild type were similar once cleavage of
gpl20hadoccurred.
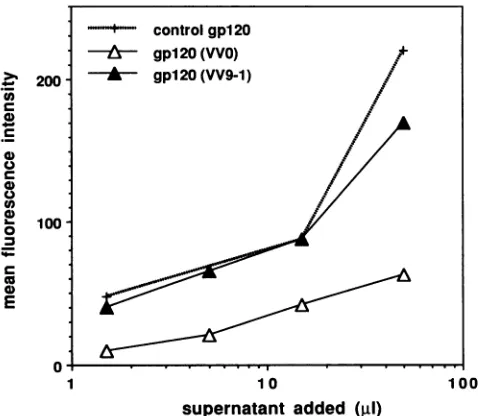
Bindingof recombinantgpl20toCD4+cells. Thecapacity
ofgpl20produced bythe mutated and controlconstructs to bind to membrane CD4 was evaluated by immunofluores-cence analysis. Various amounts of concentrated
rVV-in-fected cell supernatants wereincubated with CD4+ CEM13 cells. Bound gpl20was then labeled with mAb9305. gpl20 from the reference batchwas used as acontrol toquantify bioactive rVV-produced gpl20(Fig. 7).
Taking into account the solublegp120ELISAquantitation
described inFig. 4B, thesamequantity ofVV9-1,VVO,and
U.
C
u)
4"
._
(0
c
ID
-o
0
E
200
-100
-1 10 100
supernatant added (p1)
FIG. 7. Bindingofrecombinant secretedgpl20tothemembrane ofCD4+ CEM13 cells as determined by flow cytometry analysis afterlabeling with mAb9305. Cells (5 x 105) wereincubated with increasingvolumes of fivefold-concentrated VV9-1-orVVO-infected
cell supernatants. Results are expressed as mean fluorescence
intensities. The reference batch (control gpl20)contained 2 ,ugof gpl20perml.
---} gpl60 (VVO)/9301 0 gp160 (VV9-1)/9301
-- gpl60(VVO)/360 A gp160 (VV9-1)/360
4.... controlgp120 -A- gpl20(VVO)
A gpl20(VV9-1) VOL. 67,1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.316.555.448.656.2]m
m
0
m
z C m
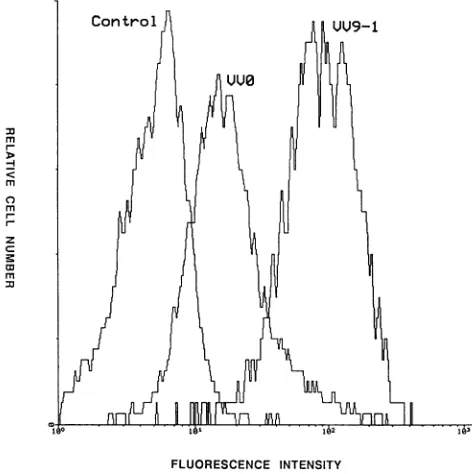
[image:7.612.62.298.76.313.2]FLUORESCENCE INTENSITY
FIG. 8. Evaluation ofgp120 expression onthe surfaceof
rVV-infected CEM13 cells. Cells (106)wereinfected with VV9-1orVVO
at 10PFUpercell. At 24 h postinfection, cellswereanalyzed by
flowcytometryaftermAb9305 labeling.Uninfected cells servedas a
control. Resultsareexpressed as cellnumbers relative to
fluores-cenceintensity.
gpl20 from the reference batch yielded the same labeling
signal, indicating that the three recombinantgpl20 proteins had the same bioactivity inasmuchas they bound to mem-brane CD4 to the same extent, even though 10-fold more
VVO than VV9-1 supernatant was necessary. Comparable results were obtained when human Abs were used for
labeling (datanotshown).
Expressionofgpl20onthe surface ofrW-infectedcells. At 2dayspostrW infection, CEM13 cellswereincubated with mAb9305 (or with mAbllO-4; data not shown) to detect gpl20 present on the cell surface. Both W9-1 and WO-infected cells expressed gpl20, but the latter in a lesser amount (Fig. 8). Reduced labeling observed after WO infection, compared with W9-1 infection, was consistent withanapproximately 10-foldreductionofgpl20 cell surface expression. Because nogpl20wasdetected in the superna-tantundertheseconditions,onemayexclude labelingdueto gpl2O capturebymembrane CD4. In agreement withwhat has beenreported (39), neither of the anti-gp4l MAbs used (mAb4la9 and mAb41-1) allowed us to detect cell surface recombinant gp4l. The same results were obtained with
rW-infectedBHK-21 cells (datanotshown).
SyncytiumformationofrW-infected CD4+lymphoid cells. Todetermine whether gp41CHO- couldmediate membrane fusion ofrW-infected and noninfected cells following gpl20 binding to CD4, CEM13 cells were infected with different
concentrations-spanning3orders of magnitude-ofWOor
W9-1. All doses of W9-1 resulted in formation of cell aggregates, detectable at 4 to 5 h postinfection, which indicatedproperbindingcapacity ofgpl20onthe surface of infected cells with membraneCD4onadjacent cells. These
aggregates developed into gp4l-mediated syncytia within a fewadditional hours. Incontrast,whileaggregateswerealso
observed after VVOinfection, nofusion occurredhere,even at the highest rVV concentration used (Fig. 9), and no further syncytiaweredetected when the cultureperiodwas prolonged by 3 days(datanotshown).There was,however,
a remote possibility that a threshold level of envelope
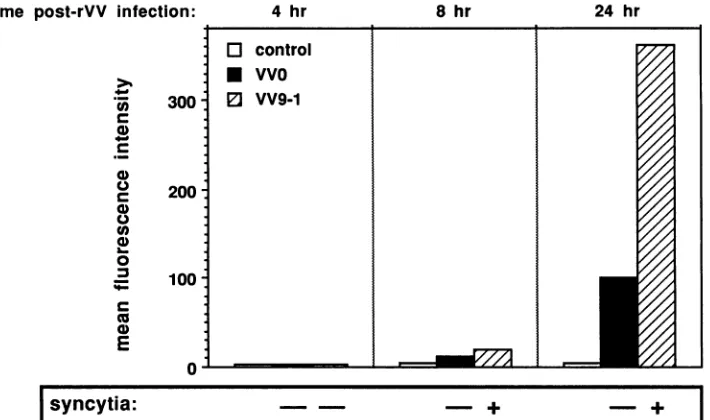
glycoprotein per cell was actually required for fusion and thatthiscouldnotbeattained withVV0, evenwith 100 PFU per cell. That such was not the case is implied by the experiment described in Fig. 10, in which the kinetics of
gp120expressiononthe membraneand theabilitytomediate syncytia of rVV-infected cells (100 PFU per cell) were compared. At 8 h after infection, W9-1-infected cells al-ready mediated syncytia at a time when membrane gpl20 detectionwaslimited,whereasby24h stillnosyncytiawere noted withVVO-infected cells, despite a greateramountof envproductexpressionat the cell membrane.
These data argue for the total inability ofgp41CHO- to mediate membranefusion.
DISCUSSION
The aim of thisstudywas toexamine whether the cluster of N-linked glycans corresponding to the three conserved glycosylation sites of the ectodomain of HIV-1 transmem-branegp41 (4, 15)wereinvolved in itsbiological property, i.e., essentially mediation of membrane fusion. Because
gp4landgp120arecleavageproductsofacommon precur-sorandbecauseit hasrecentlybeenshown that distant parts ofgp120contributetomoleculefolding(30, 36),thepossible
effects of the absence of theseglycans ongpl60proteolytic
cleavage and on gpl20 characteristics were also investi-gated. Intracellular routing or oligomerization (6, 34, 38),
which are currently under investigation, were out of the scope ofthis study.
The three conserved glycosylation sites of the env se-quenceweremutated,expressed byusing rVV, and usedto infect either CD4- BHK-21orCD4+ CEM cellstoproduce amutatedenvelopeglycoprotein.
First, we showed by WB analysis that the mutations deletedglycans from thegp4lmoleculesproduced.The MW of mutated recombinant gp41CHO- was 35,000, which in agreementwithapreviousreport(12),correspondedtothat of viral gp4l or its wild-type recombinant (gp41CHO+)
counterpart aftercompletedeglycosylationby endoF
N-gly-canase, an enzyme that cleaves all N-linked glycans (40).
This reduced MW may beaccounted for by the absence of three approximately 2,000-MWglycans (22). That the MW did not change after N-glycanase treatment indicates that
gp41CHO- wasnotglycosylatedatall. Itis thereforelikely
that the three conserved potential
glycosylation
sites re-moved (Asn-621, Asn-630, and Asn-642) are the only ones thatareactuallyglycosylatedongp41CHO+.This stands in contrast tothe resultsof Dederaetal.(3)andLee etal.(25),whosuggestedthatAsn-616 may beglycosylated.However, it ispossiblethat the lackofglycosylation at this site in the VVO construct is due toimpairment of
gp4l's
normal con-formationby themutation, resultingin steric hindrance and inaccessibility of this regiontoglycosylation enzymes(22).Therateofsynthesisofmutant gpl60wascomparableto that ofcontrol
gpl60
inpulse-chaseexperiments performed after BHK-21 cell infection with eitherVV9-1orVVO.Lack of gp41 glycosylation strongly affected the gpl60 cleavage rate,whichappeared to decrease by approximately90% as determined by quantitative ELISA. In pulse-chaseexperi-ments performed with VV9-1-infected cells, large amounts ofgpl20werenoted aftera1-hchaseand even though it was
on November 9, 2019 by guest
http://jvi.asm.org/
ROLE OFgp4l GLYCANS 157
rk~~~< j _ r-'ww &
S: '%. ^ _ t" E . e;
z~~~
.p'1 ( *-4,*~~,,,,~ _
t
_;ma
_<.~~~~~*1...
S;l ..-- s
V-4c
Ir
2 b < x?Fi.:e,"¢i. C
*~~~$g'fS;f *TrX.1;'at.b
4~~~~~~~~~~
XSw
f4,
'Y. *'
la
F.
J.;
%.+
I i. ,FIG. 9. AbilityofVV9-1andVVOtomediatesyncytium formation. CD4+CEM13 cells(106)wereinfected withVV9-1(A, B,and C) or VVO(D and E)at100(AandD),10(B andE),or1(C)PFUpercell. At 1 day postinfection, plates were photographed. Uninfected cells are showninpanelF.
secretedathighlevels in culture supematants, asdetermined by ELISA, itwasstill detected incells after 18 h. Thus, the
remaining radiolabeled gpl60 poolcontinued to be cleaved foralongtime aftersynthesis. Incontrast, somegpl20with
immature glycan speciesresultinginareduced apparent MW
appeared in VVO-infected cells during the first hour of the
chase, and it was not detectable later. This presumably
resulted fromsheddingof themolecules (6), asdetermined byaweak level of detection in the supematant byELISA. One may then assume that
only
asmallproportion of VV0gpl60 was able to be cleaved normally; transported to the cell surface,asdeterminedbyflow cytometry; and
progres-sively shed (6, 38). In contrast with what was noted with
VV9-1,
lack of cleavage of the remaining gpl60 species might be due to the fact that they were directed to a cell compartment where proteolytic activitywas not present, apossibility which is under investigation. This has already
been described for gpl60 after amino acid point mutations
(44).
However,gpl60 synthesizedbythe mutatedconstructdid not accumulate in cells, as indicated by the equivalent
amountsof intracellulargpl60detectedbyELISA in VV9-1-andVVO-infectedcelllysates,which agrees with results that
suggestthat uncleaved gpl60is cleared in cells (43). Some otherposttranslational modifications, such as, inparticular, changes in glycan processing(22),may occur onuncleaved mutatedgpl60, asindicatedbyitsdecreased MW observed in the WB and pulse-chase experiments. This was further
suggestedbythe increasedendoglycosidaseHsensitivity of
mutated gpl60 species relative to their wild-type counter-parts(datanotshown).
Lackofgp4lglycosylation apparently did not modify the
accessibility of the gp120 and gp4l central andC-terminal regions to Abs such asmAb9305, mAbllO-4, and Abl20to with respectto gpl20and mAb41a9, mAb41-1, and Ab41to forgp4lwhen thecorresponding epitopeswereexpressedon intracellular uncleavedgpl60. Surprisingly, MAbs directed to anN-terminalgpl20 epitope (mAb360,whichrecognizes
the sequence ofaa 72 to 90) or to a C-terminal epitope of
gp120 (mAb9301), a few residues upstream of that recog-nizedby polyclonalAbl20to,boundtomutated recombinant
gpl60to a much greaterextent thantoitscontrol counter-part, indicating greater accessibility of the
corresponding
regions of gpl20 under these conditions. Because of their distance from the mutatedglycosylationsites, this suggests thatdistantregionsofgpl60 act inconcert toallow normal VOL. 67,1993
A 11
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.97.517.77.439.2]time post-rVV infection:
._
cn
.1
c Co 0 0 C.) L.) 0
Cu
E
[image:9.612.136.491.74.284.2]4 hr 8 hr 24 hr
|syncytia:
--
-+
- +I
FIG. 10. Comparative evaluationof the kinetics ofgpl20expressiononthe membrane ofVVO-andVV9-1-infectedcells,asdetermined byfluorescence-activated cell sorteranalysiswith mAb9305 andexpressedas meanfluorescenceintensities,and of the abilities ofVVOand VV9-1tomediatesyncytium formation (+, syncytia observed; -,nosyncytia).
conformation of the molecule,which is in line with recent data (30, 36) indicating that distinct regions combine for properconformation ofgpl20.Suchamechanismmightalso explain the observed impaired precursor cleavage due to lack ofaccessibility to proteases and/or inappropriate con-formation ofthe relevant region (28). However, bothtypes of bioactive gpl20 species produced after cleavage and released in the supematant exhibited similar, very low avidities for mAb360 and mAb9301. As a consequence, it seems that some reshaping might occur after cleavage,
resulting in gpl20 molecules that are correctly folded for
bindingto membrane CD4.
We have recently shown that gpl60 cleavage occurs
predominantlybutnotexclusivelyatArg-516,asreported by others (1, 11, 18, 28), and that approximately 10to 20% of cleavedsubunits result fromcleavageatArg-509 (11). Here,
we evaluated the cleavage rate at either of these sites to
examine whether the reduced cleavage of mutated gpl60 could be accounted for by impairment of cleavage at the preferential site (Arg-516).Infact, althoughoverallcleavage
wasgreatly reduced, cleavage of mutatedgpl60 occurredat
both sites in the same proportion as for wild-type gpl60.
Transportoftheresultingsubunitstothe cellsurfacewasnot abolished,andVVO-producedgpl20could bedetectedboth at the membrane and in the supernatant of VVO-infected cells,albeitatreduced levels(approximately 10%) relativeto
VV9-1-infected cells.
Ourfindingsonthe influenceof lack ofgp4l glycosylation
on gpl60 cleavage are in line with findings that abnormal
glycosylation resulting from theuseofglucosidase inhibitors isassociated with human T-cellleukemia virustype1(35),as
wellaswithHIV-1 (33; reviewedin reference 37), impaired env-encoded glycoprotein cleavage.
We also investigated the ability of mutated env-encoded glycoproteins to mediate fusion of rVV-infected and unin-fected CD4+ cells, since membrane glycoprotein gpl20 of VV9-1-infected cells isalready knowntobindtomembrane CD4 ofadjacent cells and to initiate gp4l-triggered mem-branefusion(21).Relativetothiscontrol, VVO-infected cells
displayed agreaterthan 3logunit-reducedabilitytoinduce
syncytia. This effectwasmuch inexcess of what would be
expected fromgpl60 cleavageimpairment, aknown
requi-site for membrane fusion (21, 28). VVO env products ex-pressed on thecell surface are actually cleaved: nosignifi-cant amount ofgpl60 was detectedby ELISA and WB in membrane and viruspellet lysates after ultracentrifugation
of the cleared supernatant from VV9-1- or VVO-infected cells, which is in line with the absence of precursorgpl60at the cell membrane and in HIV-1 particles (8, 11, 34).
Moreover, expression of bioactivegpl20 on the surface of these cellswasreducedbynomorethan10-fold,asufficient amounttoallow for itsinteraction withCD4+ cells. One may then reasonably hypothesize that lack ofsyncytium forma-tion capacityis dueto the absence ofgp4l glycans and that theresultingreducedhydrophilicity maymodifythecorrect
presentationof the hydrophobicfusionpeptide (14).
These resultsareincontrastwith those of Dederaetal.(3)
andLee etal. (25),whoshowed thatsingle N-glycosylation
sitesare dispensable forproduction of infectious virus and that precursor cleavage and
gp4l
fusion ability were notsignificantly affected, while we found that these properties
weredramatically affected. Suchadiscrepancycouldsignify
that single glycosylation site removals perse are not suffi-cienttoabolishgp4lbioactivity, while mutation of clustered
glycosylationsites and theresultinglack ofgp4l glycanscan diminish cleavage of gpl60 and completely suppress the
fusogenicpropertiesof
gp4l.
This effectmightbeexplainedby possible complementation of one site by the others, resultinginonly limited consequences ofsingle-siteremoval. The same type of mechanism has also been suggested for HIV-1 and HIV-2gpl20, in which single glycosylation site removal showedonly limited consequences relativeto
mul-tiple-siteremoval(5, 24, 31).
In conclusion, our results confirm and extend previous findingsthatwhileglycanspresenton mature
gp4l
andgpl20afterbiosynthesis are notofparamount importance for the
biological propertiesof theproteins (9, 12), N-glycosylation
on November 9, 2019 by guest
http://jvi.asm.org/
ROLE OF gp4l GLYCANS 159 during biosynthesis of both gp120 and gp4l is essential for
properly foldedbioactive molecules (10, 17, 33).
ACKNOWLEDGMENTS
We are indebted to John P. Moore (Aaron Diamond AIDS Research Center, New York, N.Y.) for helpful suggestions regard-ing the ELISA experiments. Bruno Guy (Trangene SA) and M. Moulard are thanked for advice relative to pulse-chase experiments. We acknowledge the help of Micael Yagello and Catherine Pioche for FACS analysis and Western blot analysis. The MRC AIDS ReagentProgramme and F. Traincard areacknowledgedfor the kind giftsofmAb360 andmAb41a9, respectively.
This work was supported by the French Agence Nationale de Recherchesurle SIDA(ANRS)andbythe U.K. Medical Research Council AIDS DirectedProgramme.
REFERENCES
1. Bosch, V., and M. Pawlita. 1990. Mutational analysis of the humanimmunodeficiency virustype 1 env geneproduct proteo-lytic cleavage site.J.Virol. 64:2337-2344.
2. Chakrabarti, S., T. Mizukami, G. Franchini, and B. Moss. 1990. Synthesis, oligomerisationandbiological activityof the human immunodeficiencyvirus type 2envelope glycoprotein expressed byarecombinantvaccinia virus.Virology 178:134-142. 3. Dedera, D., R. Gu, and L. Ratner. 1992. Role of
aspargine-linkedglycosylation in human immunodeficiencyvirus type 1 transmembraneenvelope function. Virology 187:377-382. 4. Desrosiers, R. C., P. Tiollais, and P. Sonigo. 1987. Sequence of
simianimmunodeficiencyvirus from macaque and its relation-shiptoother human and simianimmunodeficiencyretroviruses. Nature(London) 328:543-547.
5. Dirckx,L., R. Lindemann, R. Ette,C.Manzoni, D. Moritz, and J. Mous. 1990. Mutation of conserved N-glycosylation around theCD4-binding site of humanimmunodeficiency virustype 1 gpl20affects viralinfectivity. VirusRes. 18:9-20.
6. Earl, P. L., B.Moss,and R. W. Doms. 1991.Folding, interaction withGRP78-BiP, assembly, andtransportof thehuman immu-nodeficiencyvirus type 1 envelope protein. J. Virol. 65:2047-2055.
7. Elbein, A. D. 1987.Inhibitors ofthebiosynthesisandprocessing of N-linked oligosaccharide chains. Annu. Rev. Biochem. 56: 497-534.
8. Fenouillet, E.,N. Blanes,andJ.C. Gluckman. 1991. gpl60of commercial Western blot isnotgpl60env. Shouldcriteria for seropositivityberevised? AIDS 5:707.
9. Fenouillet, E., B.Clerget-Raslain,J. C.Gluckman,D.Guetard, L.Montagnier, and E.Bahraoui.1989.RoleofN-linkedglycans inthe interaction betweentheenvelope glycoproteinof human immunodeficiencyvirus and its CD4 cellular receptor. J. Exp. Med. 169:807-822.
10. Fenouillet, E.,andJ.C.Gluckman.1991. Effect ofaglucosidase inhibitor on the bioactivity and immunoreactivity of human immunodeficiencyvirus type 1envelope glycoprotein. J. Gen. Virol. 72:1919-1926.
11. Fenouillet, E.,andJ.C. Gluckman. 1992.Immunological anal-ysisof humanimmunodeficiency virus type 1 envelope glyco-protein proteolyticcleavage. Virology187:825-828.
12. Fenouillet, E., J. C.Gluckman,and E. Bahraoui. 1990.Role of N-linked glycans of envelope glycoproteins in infectivity of humanimmunodeficiencyvirus type 1. J. Virol. 64:2841-2848. 12a.Fenouillet, E.,and M. P.Kieny.Unpublished data.
13. Freed, E., and R.Risser. 1990. The role of the HIV envelope glycoproteinincell fusion and thepathogenesisofAIDS. Bull. Inst. Pasteur 88:73-110.
14. Freed,E.O., D.J. Myers,andR. Risser.1990.Characterization of thefusion domain of the humanimmunodeficiencyvirus type 1 envelope glycoprotein gp4l. Proc. Natl. Acad. Sci. USA 87:4650-4654.
15. Gallaher, W. R., J.M. Ball,R. F. Garry,M. C. Griffin,and R.C. Montelaro. 1989. Ageneralmodel for the transmembrane proteins of HIV and other retroviruses. AIDS Res. Hum. Retroviruses5:431-437.
16. Geyer, H., C. Holschbach, G. Hunsmann, and J. Schneider. 1988.Carbohydrates of human immunodeficiencyvirus. J. Biol. Chem. 263:11760-11767.
17. Gruters, R. A., J. J.Neetfes,M.Tersmette, R. E. V. De Goede, G.Huisman, R. Miedema, and H. L. Ploegh. 1987. Interference with HIV-inducedsyncytiumformation and viralinfectivity by inhibitors of trimming glucosidase.Nature(London)330:74-77. 18. Guo,H.G.,F.Veronese, E. Tschachler, R. Pal, V. S. Kalyanara-man, R. C. Gallo, and M. Reitz. 1990. Characterizationofan HIV-1pointmutantblocked inenvelope glycoprotein cleavage. Virology174:217-224.
19. Jones, I., and G. S. Jacob. 1991. Anti-HIVdrug mechanism. Nature(London)352:198.
20. Kieny, M. P., R.Lathe, and R.Drilliens. 1984. Expression of rabies virus glycoprotein from a recombinant vaccinia virus. Nature(London)312:163-166.
21. Kieny, M. P., R. Lathe, Y.Riviere, K. Dott, D. Schmitt, M. Girard, L. Montagnier, and J. P. Lecocq. 1988. Improved antigenicity of the HIVenvprotein by cleavage siteremoval. ProteinEng. 2:219-225.
22. Kornfeld, R., and S. Kornfeld. 1985. Assemblyof asparagine-linkedoligosaccharides.Annu. Rev.Biochem. 5:631-664. 23. Kowalski, M., J. Potz, L.Basiripour,T.Dorfman,W.C.Goh,E.
Terwilliger, A. Dayton, C.Rosen,W.Haseltine,andJ.Sodroski. 1987. Functional regions of the envelope glycoproteins of HIV-1.Science 237:1351-1355.
24. Lee, W. R., W.Syu, B.Du, M.Matsuda,S.Tan,A.Wolf,M. Essex,and T. H. Lee. 1992. Non random distribution ofgpl20 N-linkedglycosylationsitesimportant forinfectivityof human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 89:2213-2217.
25. Lee, W.R.,X.F.Yu,W.J.Syu,M.Essex,and T. H. Lee.1992. Mutationalanalysis of conserved N-linked glycosylation sites of humanimmunodeficiencyvirus type 1 gp4l.J.Virol. 66:1799-1803.
26. Leonard,C.K.,M. W.Spellman,L.Riddle,R.J.Harris,J.N. Thomas, and T. J. Gregory. 1990. Assignment of intrachain disulfide bonds and characterization ofpotential glycosylation sites of the type 1 recombinant humanimmunodeficiencyvirus envelope glycoprotein (gpl20) expressed in Chinese hamster ovarycells. J. Biol. Chem. 265:10373-10382.
27. Machamer, C.,R. Z. Florkiewicz, andJ.Rose. 1985. A single N-linked oligosaccharide at either ofthe two normal sites is sufficient for transport of vesicular stomatitis virus Gproteinto the cell surface. Mol. Cell. Biol. 5:3074-3085.
28. McCune, J.,L. B.Rabin,M. B.Feinberg,M.Lieberman, J.C. Kosek, G. R. Reyes, and I. L. Weissman. 1989.Endoproteolytic cleavage of gpl60 is required for the activation of human immunodeficiencyvirus. Cell 53:55-67.
29. Moore, J. P., J.A.McKeating,I. M.Jones,P. E. Stephens,G. Clements,S.Thomson,and R. A.Weiss. 1990.Characterization of recombinantgpl20andgpl60 fromHIV-1:bindingto mono-clonal antibodiesand solubleCD4.AIDS 4:307-315.
30. Morikawa, Y., J.P.Moore,E.Fenouillet,andI. M.Jones.1992. Complementation of HIVglycoprotein mutations in trans. J. Gen. Virol. 73:1907-1913.
31. Morikawa, Y., J.P.M.Moore,A.J.Wilkinson,andI. M.Jones. 1991. Reduction in CD4bindingaffinityassociated with removal of a single glycosylation site in the external glycoprotein of HIV-2.Virology 180:853-856.
32. Morikawa, Y., H.A. Overston, J.P. Moore,A. J. Wilkinson, R. L. Brady, S. Lewis,and I. M. Jones. 1990. Expression of HIV-1 gpl20and human soluble CD4by recombinant baculo-viruses and their interaction in vitro. AIDS Res. Hum. Retro-viruses6:765-773.
33. Pal, R., G.Hoke,and M. G.Sarngadharan. 1989. Roleof the oligosaccharides in theprocessingand maturation ofenvelope glycoproteinsof human immunodeficiencyvirus type 1. Proc. Natl.Acad. Sci. USA 86:3386-3388.
34. Pinter, A.,W.J.Honnen, S. A.Tilley,C.Bona,H. Zaghouhi, M. K.Gorny,andS. Zolla-Pazner. 1989.Oligomericstructureof gp4l, thetransmembrane proteinofHIV-1. J. Virol. 63:2674-2679.
VOL. 67,1993
on November 9, 2019 by guest
http://jvi.asm.org/
35. Pique, C., D. Pham, T. Tursz, and M. C. Dokhelar. 1992. Human T-cell leukemiavirustype1 envelope protein matura-tion process: requirement for syncytium formation. J. Virol. 66:906-913.
36. Pollard, S. R.,M. D.Rosa, J. J. Rosa, andD. C. Wiley.1992. Truncated variants of gpl20 bind CD4 with high affinity and
suggestaminimum CD4 binding region. EMBOJ. 11:585-591.
37. Ratner, L.1992. Glucosidase inhibitors fortreatmentofHIV-1 infection. AIDS Res. Hum. Retroviruses 8:165-173.
38. Stein, B. S.,and E.G.Engleman.1990.Intracellular processing of the gpl60 HIV-1 envelope precursor. J. Biol. Chem. 265: 2640-2649.
39. Stevenson, M., C. Meier, A. M. Mann, N. Chapman, and A. Wasiak. 1988. Envelope glycoprotein of HIV induces
interfer-ence and cytolysis resistance in CD4+ cells: mechanism for
persistence in AIDS. Cell 53:483-496.
40. Tarentino, A. L., R. B. Trimble, and T. H. Plummer. 1989. Enzymatic approaches for studying the structure, synthesis,
andprocessing of glycoproteins. Methods Cell Biol. 32:111-139. 41. Wain-Hobson, S.,J. P.Vartanian, M. Henry, N.Chenciner, R. Cheynier, S. Delassus, L. Pedraza-Martin, M. Sala, M. T. Nugeyre, D. Guetard, D. Klatzmann, J. C. Gluckman, W. Rozenbaum, F.Barre-Sinoussi,and L.Montagnier. 1991. LAV revisited: origins of the early HIV-1 isolates from Institut Pasteur.Science 252:961-965.
42. Wells, D. E., and R. W. Compans. 1990. Expression and characterisation ofafunctional humanimmunodeficiency virus
envelope glycoprotein in insect cells. Virology 176:575-586. 43. Willey, R. L., J. S. Bonifacino, B. J. Potts, M. A. Martin, and
R.0.Klausner.1988.Biosynthesis, cleavage and degradation of
the humanimmunodeficiency virus type-1 envelope glycopro-tein gpl60. Proc.Natl. Acad. Sci. USA 86:9580-9584. 44. Willey, R. L.,T.Klimkait, D. M. Frucht, J. S. Bonifacino, and
D. M.Martin.1991. Mutations within the human immunodefi-ciency virus type-1 gpl60 envelope glycoprotein alter its intra-cellulartransportandprocessing. Virology 184:326-329.