0022-538X/92/041924-09$02.00/0
Copyright© 1992, American Society for
Microbiology
The Sequence Context of the Initiation Codon in the
Encephalomyocarditis Virus Leader Modulates
Efficiency of Internal Translation Initiation
MONIQUE V. DAVIES ANDRANDAL J. KAUFMAN*Genetics Institute, 87Cambridge ParkDrive, Cambridge, Massachusetts02140-2387 Received6September 1991/Accepted 19 December 1991
Translation initiation on poliovirus and encephalomyocarditis virus (EMCV) mRNAs occurs by a cap-independent mechanism utilizinganinternal ribosomalentrysite(IRES). However,nounifyingmechanismfor AUG initiation site selection has been proposed. Analysis of initiation of mRNAs translated in vitro has
suggested that initiation of poliovirus mRNA translation likely involves both internal binding of ribosomes and scanningtothe first AUG which is inafavorablecontextfor initiation. Incontrast,internal initiationonEMCV mRNAmaynotutilize scanning, since ribosomes binddirectlyor veryclosetotheinitiation codon AUG-li. We have studied in vivo thesequencerequirements for internal initiation around the EMCV initiation codon, both
inmonocistronic and in dicistronic mRNAs. Our studies show that theupstreamAUG-10 is normallynotused and that there isnospecificsequencerequirement for nucleotides between AUG-10 and AUG-11. However, the
sequence contextofAUG-11 does influence the efficiency of initiation at AUG-11. Efficient IRES-mediated internal initiation at AUG-11 exhibits a requirement for an adenine in the -3 position, similar to
cap-dependent initiation. These results support a model for internal initiation on EMCV mRNA in which scanning starts at ornearAUG-11. Although initiation primarily occurs at AUG-11, initiation at multiple downstream AUG codons can bedetected. In addition, a poor sequencecontext around AUG-11 results in increased initiation atone or moredownstreamAUG codons, indicative of leaky scanningorjumping by the ribosome fromAUG-11 mediated by the EMCV IRES.
Picornavirus mRNA translation initiates through direct, cap-independentinternalbindingof ribosomesmediatedbya complexinteraction betweenatleasttwodistinctregions in the5' untranslated regions (UTRs) of these mRNAs which
mayinvolve specificcellularproteins (9, 13, 28, 29, 33). On
the basis of sequence homology between putative internal
ribosomal entry sites (IRES) within their 5' UTRs, the
picornaviruses have been classified into three groups: (i) hepatitis Avirus, (ii) the cardioviruses and aphthoviruses,
and(iii) the enteroviruses andrhinoviruses.Cardiovirusand
aphthovirus mRNAs,asrepresented by
encephalomyocardi-tis virus (EMCV) and foot-and-mouth disease virus, are
translated with very high efficiency and accuracy in the rabbit reticulocyte lysate, while poliovirus and human
rhi-novirus mRNAsaretranslatedinefficientlyandinaccurately
in the rabbitreticulocytelysatebutefficientlyandaccurately
in HeLa cellextracts(5, 33).The IRESnucleotidesequence
is conserved within each class, but little conservation is found between the different classes. For cardioviruses, enteroviruses, and rhinoviruses, twodomains within the 5' UTR are essential for internal initiation: a stem-and-loop structure and a pyrimidine stretch in the vicinity of the initiationAUG(9, 12, 13, 33). The selection mechanism for the initiation AUG, however, may be different for each
class. Inpoliovirus,the ribosomes bindtotheIRES and then
scanfromapproximatelynucleotide(nt)600tothe initiation codonatnt 743. This mechanism issupported byadeletion (from nt 600to 727)which has noeffect onviralinfectivity (25). Furthermore,insertion ofa72-ntsequencewhich lacks AUG codons into the 5' UTR ofpoliovirusat nt 702 hasno effectoninfectivity,but insertion ofasequenceharboringan
*Correspondingauthor.
AUG codon with an unfavorable sequence context for initiation produces a small-plaque phenotype (24). In con-trast,studiesoncardiovirusessuggestthatribosomes initiate directlyatAUG-11withoutprior scanning (8, 14).Initiation
atAUG-10was not detectedupon introduction ofa
frame-shift mutationtoallow thedetection ofpolypeptidesinitiated
atAUG-10. AphthovirusmRNAsmayinitiateat eitherone
oftwo in-frame AUGswhich are 84nt apart and are each
preceded bytwoconservedpyrimidinestretches of
approx-imately 20 bases (35). If the first AUG codon is in an
unfavorable context for initiation (20, 21), it isvery rarely
used in vivo and asizableportion of the ribosomesscan to the next AUG codon which has a favorable context in all strains. In some serotypes, both AUGs are in a favorable contextfor initiation and bothareused in vivo and in vitro (26, 35).
Tostudythe mechanismof initiationonEMCV mRNA in
vivo,weintroduced theEMCVleader fromnt260to834 into
aeukaryotic expressionvectordirectly precedingthe marker
dihydrofolate reductase (DHFR) gene so that EMCV
AUG-11waseitherdirectly fusedorin frame witha
down-stream DHFR initiation codon. The efficiency of use of
EMCV AUG-11 as well as the occurrence of initiation at downstream AUGswasassessedbytransientexpression in
COS-1monkeycells transfectedwith the various constructs
and analysis of labeled cell extracts by reducing sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). We have evaluated the effect of specific base
substitutions around AUG-10 and AUG-11 which affect
utilization of AUG-11 as well as initiation at downstream
AUG codons. The results show significant initiation at codons downstream of AUG-11, and the efficiency of AUG-11 utilization is increased following nucleotide 1924
on November 9, 2019 by guest
http://jvi.asm.org/
changes which conserve the consensus sequence for the AUG initiation codon.
MATERIALS AND METHODS
Plasmidconstructions. pMT21, a derivative of pMT2 (17), and pED4 have been previously described (18). Changes in the EMCV leader region (nt 828 to 846) were introduced by using oligonucleotide-directed mutagenesis by the gapped heteroduplex procedure (30), with modifications as de-scribed previously (17) to yield the vectors pEDn (pED1 to pED5). All mutations were confirmed by DNA sequence analysis (36). A 1.6-kb eIF-2ao cDNA was obtained by digestion ofpeIF-2a VA+ (17) with EcoRI and ligated into the EcoRI site of pMT21 and pEDn, to yield dicistronic vectors pMT21-2cx and pEDn-2a.
DNAtransfections and analysis. COS-1 monkey cells were transfected by theDEAE-dextran procedure, with the addi-tion of achloroquine treatment as described previously (16). After 42 h, cells were labeled with [35S]methionine (100
,uCi/ml; 1,000 Ci/mmol; Amersham Corp., Arlington
Heights, Ill.) for 30 min in methionine-free minimal essential medium. Cell extracts were prepared by lysis in Nonidet P-40 asdescribed previously (17) and analyzed by reducing SDS-PAGE (27), either before or after immunoprecipitation withrabbit polyclonal anti-mouse DHFR kindly provided by Joseph Bertino(Sloan-Kettering Memorial Cancer Institute, New York, N.Y.). Gels were fixed in 40% methanol-10% acetic acid, prepared for fluorography by treatment with En3Hance (New England Nuclear Corp.), and dried. Dried
gels were autoradiographed with Kodak XAR-5 film with a Dupont Cronex Lightning-Plus screen. Protein levels were estimated by visual comparison of band intensities from multiple autoradiograms of different exposure times.
Total RNA was prepared by guanidine thiocyanate extrac-tion (3) and analyzed by Northern (RNA) blot hybridizaextrac-tion
(38) following electrophoresis on formaldehyde-formamide
denaturing agarose gels as described previously (4). Hybrid-ization was carried out by using a DHFR probe prepared by
[32P]dCTP
labeling, using random priming with oligonucleo-tides as described by the supplier (Pharmacia Inc.).Electroporation and selection of DHFR-deficient CHO cells (DUKX-B11) was performed as described previously (39), with modifications. Subconfluent cultures of cells were trypsinized, and 2 x 106cells wereresuspended in 0.9 ml of Dulbecco's minimal essential medium supplemented with 10 mM glutamine. DNA (100 ,ug) was linearized by digestion
withNdeI,resuspendedin 0.1 ml of sterile water, and added
tothe cells.This mixture was exposed to 200 V at 1,250 ,uF
(Cell ZapII; Andersen Electronics, Brookline, Mass.), and
the cells wereplated within 5 min into nonselective medium. Two days later, cells were subcultured 1:15 either into selective medium lacking the nucleosides or into the same medium containing increasing amounts of methotrexate
(MTX). After 10 days to 2 weeks, colonies were counted
afterstaining with methylene blue. RESULTS
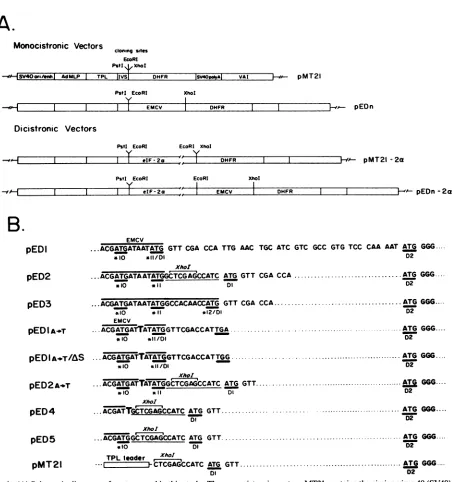
Constructionofexpression vectors incorporating the EMCV leader. To determine the sequence requirements around AUG-11 for efficient translation initiation mediated by the EMCV leader, the EMCV leader sequence (nt 260 to 834)
was inserted upstream of the reporter gene DHFR in the vectorpMT21 (18). To show that internal initiation mediated by the EMCV leader is independent of the expression of an
upstream open reading frame, dicistronic vectors were con-structed by inserting a 1.6-kb eIF-2a cDNAinto the unique EcoRI site preceding the EMCV leader. Schematic diagrams of monocistronic and dicistronic constructs used in this study are shown in Fig. 1A. Changes were introduced around the initiation sites by using gapped heteroduplex mutagenesis methods, generating the vectors pEDn. Se-quence junctions between the EMCV leader and DHFR coding regions in the different constructs are shown in Fig.
1B.
Utilization of EMCV AUG-11 and downstream initiation codons. The efficiency of use of the authentic EMCV initia-tion codon AUG-11 (designated *11) at nt 834 and the occurrence of initiation at downstream AUG codons was monitored by derivation of pED1, pED2, and pED3 (Fig. 1B). pEDi fuses *11 with the authentic DHFR initiation codon (designated Dl), resulting in a common initiation codon (*11/Dl). pED2 places Dl in frame with *11 by inserting a 12-bp oligonucleotide containing a XhoI site between *11 and Dl. pED3 retains the EMCV leader se-quence up to EMCV AUG-12 (designated *12), which is fused toDl, resulting in a common initiation codon (*12/Dl). pMT21, which encodes the authentic murine DHFR cDNA and does not contain the EMCV leader, was used as a control. Expression of DHFR from these four constructs was analyzed by transient transfection of COS-1 monkey kidney cells. EMCV vectors were transfected in duplicate, using independently derived clones from heteroduplex mu-tagenesis. DHFR synthesis was studied by
[35S]methionine
pulse-labeling of transfected cells and analysis of the total cell extracts by SDS-PAGE (Fig. 2A). Parallel plates of transfected cells were used to isolate total cellular RNA, which was analyzed by Northern blot hybridization to a DHFR-specific probe (Fig. 2B). In total cell extracts pre-pared from cells transfected with pMT21 (Fig. 2A, lane 2) as well as in cells transfected with pED1 (Fig. 2A, lanes 3 and 4), a major 21.5-kDa species which corresponds to DHFR initiated at Dl was detected. Extracts prepared from cells transfected with pED2 (Fig. 2A, lanes 5 and 6) and pED3 (Fig. 2A; lanes 7 and 8) showed, in addition to DHFR, a slightly more slowly migrating species. The slower mobility isconsistent with a DHFR species containing an additional five (Fig. 2A, lanes 5 and 6) or four (Fig. 2A, lanes 7 and 8) amino acids corresponding to initiation at *11. In all three vectors, the major site of initiation was *11, although in pED2 and pED3 a considerable amount of initiation also occurred at the downstream AUG,Dl. Initiation at both *11 and Dl was approximately fourfold greater in pED3 (Fig. 2A, lanes 7 and 8) than in pED2 (Fig. 2A, lanes 5 and 6). This finding suggests that nucleotide sequences 3' to the EMCV AUG-11 initiation codon can influence initiation at EMCV AUG-11.Northern blot analysis of RNA harvested from parallel transfected plates revealed similar levels of a single mRNA species of the expected size (Fig. 2B). Therefore, DHFR synthesis in these transfected cells reflected translation efficiency. Alteration of the sequence context of EMCV AUG-11 reduces its efficiency and increases downstream initiation.
Apurine (usually A) in position -3 of the initiation codon is the most conserved nucleotide in all eukaryotic mRNAs (20). In the literature, either an A (13, 14) or a U (31) was reported at the-3position (residue831) with respect to *11. Toevaluate the importance of this residue, the A at position -3(present in vectorspED1,pED2, and pED3) was mutated to T by site-directed mutagenesis in pED1 andpED2 (Fig.
on November 9, 2019 by guest
http://jvi.asm.org/
A.
Monocistronic Vectors cloning
sates
EcoRI
Pst!.4 XhoI
-VSV4Oon/nIiAMLP §TPL |IVS DHFR ISV40p*Al VAI- pMT21 Pst! EcoRI XhoI
-v.~~ ~ ~ ~~~~I EMCV
-I
DHFR I i'-e pEDnDicistronic Vectors
Pstl EcoRI EcoRI Xhol
Y EIIC f |YHR. _* pO
Xr E .I{ I
-~fI elF-2a DHFR -F- pMTZI-7a
Pstl EcoRI EcoRI Xhol
elF-2a EMCV ONFR l pEDn-2a
EMCV
pEDI ...ACGATGATAATATG GTT CGA CCA TTG AAC TGC ATC GTC GCC GTG TCC CAA AAT ATG GGG
*10 *II/D1 02
Xhor
pED2
...ACGATGATAATATGGCTCGAGCCATC ATG GTT CGA CCA.ATG GGG.*l10 *11 Dl D2
pED3 ...ACGATGATAATATGGCCACAAOCATG GTT CGA cca.ATGGGG
0lo *11 *12/01 D2
EMCV
pEDIA-T ...ACGATGATTATATGGTTCGACCATTGA ATG GGG....
si10 *11/DI D2
pEDIAOT/tAS ..ACGATGATTATATGGTTCGACCATTGG .ATG GGG....
*10 /11/D2
XhoI
pED2Ar2T ACGATGATTATATGGCTCGAGCCATC ATG GTT.ATGG6GG.
*10 *1I DI D2
Xhor
pED4 ... A CGATTGCGAGCCATC ATG GT... ATG GGG..
XhoI
pED5
...ACGATGGCTCGAGCCATC
ATG GTT ....*10 DI 02
TPL leader xhoI
pMT21 .. J-CC TCGAGCCATC ATG Ga GGG....
DI D2
FIG. 1. (A)Schematicdiagramsofvectorsusedinthisstudy.ThemonocistronicvectorpMT21containsthesimianvirus 40
(SV40) origin
ofreplicationandenhancerelement(HindIII-PvuIIfragment), theadenovirusmajorlate promoter(AdMLP)fromtheXhoI site
(15.83
mapunits) to the 5' cap site, the tripartite leader (TPL; 180 bp of thefirst twoand two-thirds of the third leader from adenovirus
major
late mRNAs),aninterveningsequencecomposed ofthe 5' splicesite from the first leader of adenovirusmajor
latemRNAsanda3'splice
site fromanimmunoglobulingene(IVS),threeuniquecloningsites(PstI,EcoRI,andXhoI),
amurine DHFRcodingregion
(DHFR),
thesimian virus40earlypolyadenylationsignal(SV40polyA),and theadenovirusVAI genefromHpaI(28.02mapunits)toBalI(29.62mapunits)
(VAI).
The backbone of thisvectorispUC18containingthe,-lactamasegeneand theEscherichiacolioriginofreplication.pEDn
vectorsarederivedfrompMT21 byinsertion of the EMCV leader(fromnt260to834)upstream of DHFR in theunique
cloning
sitesEcoRIandXhoI.Changes
were introduced aroundthe EMCVinitiationcodonby using heteroduplexmutagenesismethods. Fordicistronic vectors,eIF-2a cDNA,a 1.6-kbfragmentexcised frompeIF-2aVA+ (17),wasinsertedinto theEcoRI site ofpMT21andpEDnto
yield pMT21-2a
andpEDn-2a.
InthevectorspEDn-2a, the EMCV leader ispositionedbetween the eIF-2aandDHFR genes, while in
pMT21-2a,
theDHFRgeneisdirectly
adjacenttoeIF-2a. (B) Sequencejunctionsbetween the EMCVleaderand DHFRcDNA. The EMCVleader sequence ishighlighted
by
a thickblack line above the sequence. The EMCV AUGsareunderlinedandlabeled*10, *11,and *12. Theauthentic DHFR initiationcodonis underlinedand labeledDl.The second DHFR AUG is alsounderlinedandlabeled D2. WhentwoAUGsarefused,thecommonAUG is underlined and labeled*11/D1 forthe fusionof EMCVAUG-11and DHFRAUG-1or*12/Di for the fusion of EMCVAUG-12 andDHFR AUG-1. TheXhoIlinker sequence is underlinedbyathinline. The mutation of AtoTis indicatedbyabold letter in
pED1
A-*T andpED2
A--T. The TGA stop codon in frame with EMCV AUG-10 is underlined in pED1A--T and changedto aTGG in
pED1
A-*TAS. The sequencefor the DHFR cDNA is identical in allvectors. ThecontrolexpressionvectorpMT21doesnotcontain the EMCV leader.. -^Al.,I -e-'%
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.89.542.76.558.2]DHFR- * _
- N to
oi a 0
w bJ W
ca cL a.
- - t
*11/Di- -_
A
i N4 U
o a
2
Kd
69-46- _ 4
DHFR _ _
3 4 5 6 7 8
B CN
06 06 X 0E
I r
---9 10 If 12 13 14 15
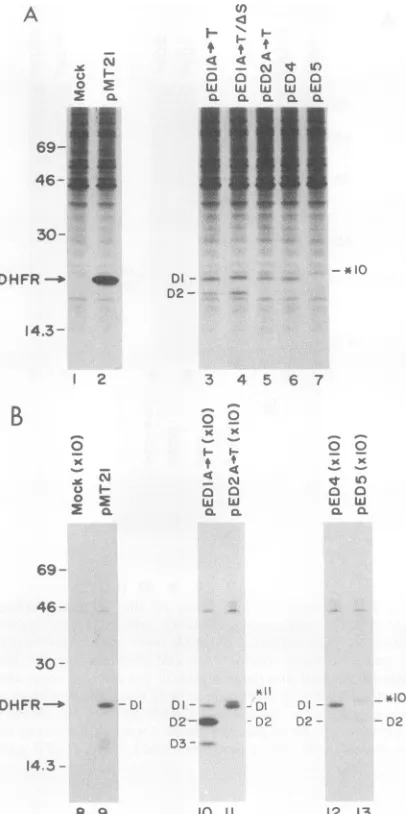
FIG. 2. Utilizationof EMCVAUG-11and downstream initiation codons. The indicatedplasmidsweretransfected into COS-1
mon-key cells.pED1,pED2,andpED3weretransfected induplicate.(A)
At 42 hposttransfection, cells werepulse-labeled for 30minwith
[35S]methionine and total cell extracts prepared for analysis by SDS-PAGE as described in Materials and Methods. Equivalents amount of cell extracts were loaded per lane (on the basis of trichloroacetic-precipitablecountsperminute). Migrationof DHFR
isindicatedbyanarrow.All DHFRspeciesarelabeledaccordingto their initiationcodon. Molecularweightmarkers(in kilodaltons)are
indicatedonthe left.(B)Fromparallel plates,at42 h posttransfec-tion,total cell RNAwasharvested andanalyzed byNorthern blot
hybridization to a DHFR probe as described in Materials and Methods.
1B).The relative useof *11 and the downstreamAUG in all fourconstructswastestedbytransienttransfection inCOS-1 cells(Fig. 3A).The utilization of *11 fusedtoDl (*11/Dl)in
pED1 A-*T (Fig. 3A, lane 5) was approximately fourfold lower than in pED1 (Fig. 3A, lane 3). In addition, a new polypeptideof 19.5kDawasdetected. Themigrationofthis
newpolypeptide species corresponds toinitiation at DHFR AUG-2 (D2), 42 nt downstream from Dl. Identification of this newpolypeptide asDHFRwasconfirmed by
immuno-precipitation with apolyclonal rabbit anti-DHFR (Fig. 4B,
lane 10). Initiation at D2 was also detected from pED1, although at a lower level (Fig. SA, lanes 14 and 24). The utilization of *11 in pED2A- T (Fig. 3A, lane 6)was also decreased relativetotheuseof *11 inpED2 (comparebands
*11 in Fig. 3A, lanes 4 and 6), while the use of the next
downstreamAUG,Dl,wasincreased. Inaddition,whereas utilization of D2 could be detected in pD2A--T (Fig. 5B,
lane 21), itwas considerably less than that ofpED1 A-*T
(Fig. SB, lane 20). These results are consistent with a scanning hypothesis in which the presence ofan upstream
AUG inafavorablecontextreducesinitiationatdownstream
AUG codons. mRNA levelsfrom allplasmidswere similar, indicatingthatdifferences in DHFRsynthesis resulted from
1 2
B
3 4
N NCQ N
a a
ai CL CoaCSLf
5 6
7 8 9 10 11
FIG. 3. Evidence that alteration of the sequence context of
EMCVAUG-11 reduces its efficiency and increases downstream
initiation. (A)Theindicatedplasmidsweretransfected intoCOS-1 cells, and equivalent amounts of cell extracts (on the basis of trichloroacetic acid-precipitable countsperminute)were analyzed asdescribedforFig. 2. Migrationof DHFRis indicatedbyan arrow.
Initiation codonsfor the various DHFR species are identified for eachplasmid.(B)Fromparallel plates, total cell RNAwasisolated and analyzed byNorthern blothybridizationtoaDHFRprobe.
translationefficiencyof the mRNAs(Fig. 3B).These results
indicate thattheadenineatposition -3 of *11,whichmeets
the consensussequence requirement for efficient initiation,
enhancesinitiation at*11. Mutationof -3 AtoTresulted in
a greater percentage of ribosomes bypassing AUG-11 to
initiate at a downstream AUG having a more favorable
context (Fig. 1B).
EMCV AUG-10 is not required for internal initiation. DHFR translation from pED1A-*Tyielded detectable ini-tiationatD2(Fig. 3A,lane5; Fig. 5A,lane6).Thesequence
between EMCVAUG-10(*10)and D2 containsastop codon TGA in frame with *10 but notwith *11/D1. To determine whether the initiation at D2resulted from initiation at *10, termination, and reinitiation, the stopcodon TGA in pED1
A-*T was mutated to a TGG in pED1 A-*TAS. Cells transfected with pED1 A-*T (Fig. 4A, lane 3) and pED1
A-*TAS (Fig. 4A, lane4) exhibitedsimilar levels ofDHFR
polypeptideinitiationatD2. Therefore,mutation of thestop
codon in frame with *10 did not alterinitiation at D2. This result indicates that initiation at D2 did not result from initiation atAUG-10, termination, and reinitiation butmore likelyresultedfrom ribosomes bypassing *11. This conclu-sion is consistent with conclusions drawn from in vitro
experiments showing thatAUG-10 isnot required for initi-ation (14).
A
X
&o X
a CL
Kd
69-
30-_-N
v C-i a0
w _
a. a
I6i6
*Am .i
14.3-*11 -XW ~ NDI
t t _ N
DO
ffi
we
ii
am1 2
30-*11/01 --_ - o,IDI
14.3-
,*I-*11- :- 'DI
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.83.284.76.356.2] [image:4.612.340.541.78.371.2]U)
< 4 c'l q LC
l
lJhJ WW
CL
..i.
-*10
14.3
--1 2 3 4 5 6 7
B
x
v
2 CL
-a
2x x
t
1 ci
w w
C. CL
K
0 0
a c)
W CL
69-
46-30
-, l D2
DHFR- - -Dl DI
-D2-_ 03
-14.3
-8 9 10 II
Di-_1*10 D2I -.D
02- -D2
12 13
FIG. 4. Evidence that EMCVAUG-10 isnotrequired for inter-nal initiation. (A) The indicated plasmids were transfected into
COS-1cells, andequivalentamountsof cellextracts(on the basis of trichloroacetic acid-precipitable countsper minute)were analyzed asdescribed for Fig. 2. MigrationofDHFRis indicatedbyanarrow.
Initiation codonsfor the various DHFR species are identified. (B)
Cell extracts were immunoprecipitated with a rabbit polyclonal
anti-mouse DHFR. Equivalentamounts ofcellextracts (trichloro-acetic acid-precipitable counts per minute) were used except for
pMT21-transfected cells, forwhich only 1/10 ofthat amount was
used. DHFR species resulting from initiation at either EMCV or
DHFR AUGsarelabeled bytheirstartcodons *10, *11, Dl,D2, and
D3.The antibody does notquantitatively precipitate DHFR, since
thesmallerDHFRspeciesare moreimmunoreactive and therefore areoverrepresented. Sizesareindicated in kilodaltons.
Todeterminewhetherthereisauniquesequence
require-ment between AUG-10 and AUG-11 for internal initiation,
AUG *11 and its surrounding sequence was deleted and
replaced by a 15-bp oligonucleotide containing aXhoI site
which keeps Dlinframewith *10 (pED5).Analysis oftotal cellextractsincellstransfected withpED5 (Fig.4A, lane 7) revealed a very faint band migrating slightly more slowly
than DHFR. Immunoprecipitation of this cell extract (Fig.
4B, lane 13) revealed two bands, one corresponding to initiation at (*10) and the other corresponding to initiation at D2. Surprisingly, initiation at *10 and not at Dl was ob-served. These results showed that *10 could be used, al-though very inefficiently. However, *10 was used preferen-tially toDl. When *10 was mutated to an ATT to yield pED4 (Fig. 4A, lane 6), the DHFR polypeptide resulting from initiation at *10 was not detected and the use of Dl was increased considerably (compare Fig. 4A, lanes 6 and 7, and Fig. 4B, lanes 12 and 13). The increase inDl utilization was unexpected and is discussed below.
The identification of the faster-migrating polypeptides in cell extracts from COS-1 transfected cells was performed by immunoprecipitation with anti-DHFR polyclonal antibody. Internal initiationmediated by the EMCV leader resulted in the synthesis of multiple DHFR-related polypeptides in addition to authentic DHFR (Fig. 4B, lanes 10 to 13). This was especially apparent in cellstransfected withpED1 A--T (Fig. 4B, lane 10; Fig. 5B, lane 20). The sizes of these polypeptides were 19.5, 16.7, and 14.5 kDa and correlated to initiation at the next three downstream AUG codons in frame with DHFR at nt 42, 111, and 156. Although the immunoprecipitation was not quantitative due to higher immunoreactivity of truncated DHFR species (compare the levels ofinitiation atDl, D2, and D3in Fig. 4A, lane 3, and Fig. 4B, lane 10) and did not accurately represent the proportion of the different DHFR species, comparison of DHFR polypeptides derived from pMT21 (Fig. 4B, lane 9) with those translated from the EMCV IRES (Fig. 4B, lanes 10 to 13) showed that internal initiation mediated by the EMCV leader is more prone to leakiness. Multiple down-stream initiation mediated by the EMCV IRES was also observed with another cDNA insert encoding adenosine deaminase inserted into the pED4 vector (29a).
Introduction of an open reading frame upstream of the
EMCV leader does not alter the pattern ofinitiation events. To determine whether the initiation events in the experiments described above result frominternal initiation, wemonitored
the effect of inserting an upstream open reading frame. Human eukaryotic initiation factor eIF-2(x was cloned into the unique EcoRI site immediately upstream of the EMCV leader in all of the EMCV expression vectors as well as in the controleukaryotic expression vector pMT21. All
mono-cistronic and dicistronic constructs were introduced into COS-1 cells. Synthesis of the two proteins, eIF-2a and DHFR, was analyzed in total cell extracts by SDS-PAGE (Fig. 5A). eIF-2a synthesis is readily detected as a major
band migrating at 35kDa. Allexpressionvectorscontaining
eIF-2a expressed this protein with the same
efficiency,
demonstrating that the EMCV leader did not interfere witheIF-2a translation or mRNAexpression. DHFRexpression
inpMT21-2ao-transfected cells(Fig. 5A, lane3)could notbe detected above background level. In contrast, DHFR expression in allEMCV dicistronicvectors wasdetected and
was either notaffectedby the presence of the upstream gene eIF-2a (lanes 5, 9, and13)orminimallyaffected (lanes 7, 11,
and 15). The pattern ofinitiation atdownstream AUGswas unchanged indicistronic vectors, and the
efficiency
ofuseofthe various AUGs was also conserved.
Immunoprecipitation of extracts from cells transfected
withdicistronic vectors (Fig. SB) detected the same DHFR specieswhich were observed inFig.4B. Note that DHFRin
lane 17 was immunoprecipitated from 10-fold-less cell ex-tract thanin lanes 18through 24.DHFR
expression
from thedicistronic vector pMT21-2cc was at least 100-fold reduced
from that of the monocistronicvectorpMT21,which didnot
A
CU
oE :ECL
69-l
46-f
30-DHFR- 4 DI-s
D2
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.79.282.76.483.2]A.
N N~~~~~~~~~~a
Cv N N N
v No St < < CU n C
CQJ
WX
7'- t "0jw 0F00i**~4I*-eIF-2ca
DHFR- qU
-B.
°
N2^°
4 5 6 7n_ *Q+* N N N
¢
N N r< N
-a
CL CL CL CL C a CL a
DHFR--* M -__
8 9 10 11 12 13 14 15
C.
NtN N
6 CL
N\ N\ N N
- ,NN r) -)
a a a a a a a a
CL :L CL C-L CL CL Ck OL
monoc
L~.
16 17 18 19 20 21 22 23 24
2 3 4 5 6 7 8 9 10
FIG. 5. Evidence thatintroductionof an openreading frameupstreamof theEMCV leaderdoes notalterinitiation at DHFR AUG codons.
(A)AllplasmidDNAsweretransfectedintoCOS-1cells foranalysisasdescribedearlier. ThepositionsofeIF-2a and DHFR areindicated by arrows. (B) Cell extracts from dicistronic constructs and control vector pMT21 were immunoprecipitated with a rabbit polyclonal anti-mouse DHFR. Equivalent amounts (trichloroacetic acid-precipitable counts per minute) of cell extracts were used except for pMT21-transfected cells,for which1/10 ofthatamount wasused.Molecularweight markersat30 and 14.3kDa are indicatedby two marks
ontheright. (C)Total cell RNAswerepreparedfrom transfectedcells and analyzed byNorthernblotandhybridizationtoaDHFR probe. Positions of 18S and 28S rRNAs are indicated by squares.
contain the EMCV leader. In contrast, DHFR expression
from theEMCVleader-containingvectorsisnotaffectedby
insertionof the eIF-2ao cDNA.
NorthernanalysesofmRNAs(Fig. 4C),inwhichselected
monocistronic and dicistronicvectorsweretransfected side by side, show thatDHFR expression obtained from dicis-tronicvectorsdidnotresultfromcap-dependent translation
fromtruncatedmRNAspecies, sinceonlyfull-length mRNA
hybridizing to a DHFR probe was detected (Fig. 4C). A longer exposure of the same blot did not reveal any trun-cated transcripts, and by comparison of short and long exposures, we are confident that truncated transcripts at a level of 1% could be detected. Dicistronic mRNA levels
wereconsistently three-tofourfold lower thanthatobtained
from monocistronic vectors. Therefore, the translational
efficiency of the second cistron in EMCV dicistronic
con-structsis eitherequaltoorgreaterthan that inits monocis-tronic counterpart. Thisfindingis consistent with previously
described observations(11, 18).
Stable expression of dicistronic vectors inCHO cells. The effect of the EMCV leader in promoting internal initiation
was alsostudied in adifferent cell and transfection system.
The different constructs were electroporated into DHFR-deficient CHO cells. After 48 h, cells were split into a
selection medium lacking nucleosides, in the absence and presenceofincreasing concentrations of MTX. Electropora-tion efficiencies were determined by counting colonies of
surviving cellswhich resulted from their ability to express
DHFR (Table 1).
Theelectroporation efficiency of all ofthe
on November 9, 2019 by guest
http://jvi.asm.org/
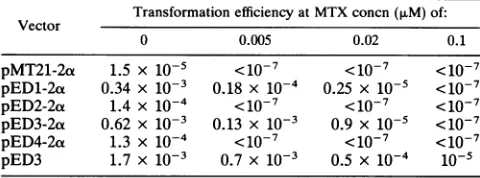
[image:6.612.145.478.77.495.2]TABLE 1. Transfectionefficiency of dicistronic vectors into CHO cells
Transformationefficiencyat MTX concn (,uM) of: Vector
0 0.005 0.02 0.1
pMT21-2a 1.5 x 10-5 <10-7 <10-7 <10-7
pED1-2a 0.34 x 10-3 0.18 x 10-4 0.25 x 10-5 <10-7 pED2-2ox 1.4 x 10-4 <10-7 <O1-7 <10-7 pED3-2oa 0.62 x 10-3 0.13 x 10-3 0.9 X 10-5 <10-7 pED4-2ot 1.3 x 10-4 <10-7 <10-7 <1o-7 pED3 1.7 x 10-3 0.7 x 10-3 0.5 x 10-4 10-5
aThe indicated plasmidswereelectroporated into DHFR-deficient CHO cells as described in Materialsand Methods.Forty-eight hours later, cells
were subculturedinto selective mediumcontaining0, 0.005, 0.02or0.1 ,uM MTX.Transformationefficienciesweredetermined by staining colonies with methyleneblue and areexpressedastheratio of coloniesto thenumber of cellselectroporated.
ing dicistronicvectors was from 10- to 50-fold higher than
that ofpMT21-2ao. This increased efficiency likely resulted
froman increased level of DHFRexpression in the EMCV-containing vectors(19). Whereas no colonies were observed when cells electroporated with pMT21-2a were selected in the presence of0.005or0.02 ,uMMTX, some cells electro-porated with pED3-2a or pED1-2a survived growth in
.02,uMMTX.TheincreasedMTXresistance reflectedhigher
DHFR expression in these cells. The electroporation
effi-cienciesof allexpressionvectorsin CHO cells reflected their
levels of DHFR expression observed in the COS-1 transient assay system.The transfection efficiency forpED3-2a was
one-third that of pED3 and correlated with the level of DHFRsynthesis observed in COS-1 cells(Fig. 5A,lanes 10 and11). Ifwe assume thataDHFRpolypeptide containing fourorfiveextraaminoacids is
active,
the DHFRsynthesisfrom thesevectorsinCOS-1 cells decreasedinthefollowing
order: pED3-2ao, pED1-2ot, pED2-2ao, and pED4-2a. Trans-fection efficiencies in CHO cells decreased in the same
order.
DISCUSSION
Phylogeneticand biochemical analysis of the 5' UTRs of
picornavirus RNAs have elucidated a minimum of two
structural elements that appear to be required to promote
internal translation initiation. A57-kDaprotein was
identi-fiedforwhich
binding
to stem-loopEatthe5' border of theIRES correlated with IRES function (13, 28). The second structural element, an oligopyrimidine
(Yn)
tract near the initiation AUGcodon,wasfirst observed in foot-and-mouthdisease virus(2)andmorerecently showntobe
required
forIRES function (13, 33).Inthe latterarticle, itwasproposed
that the Yn tract isseparatedfrom theAUG-11 codonbya number of nucleotides (Xm) and that the combined unit
(Yn-Xm-AUG-11) may act as an essential signal to
ribo-somal entry. This study was undertaken to evaluate the sequence requirements around the initiation codon of the EMCV leader in promoting cap-independent translation in
vivo. Our results support amechanism in which ribosomes
start scanning at or immediately upstream of AUG-11.
Scanning initiated at AUG-11 was
leaky,
and a poorse-quence context for initiation around AUG-11 resulted in increased initiation at one or more downstream AUG codons.
Fusion of thewild-typeEMCV leader sequenceat AUG-11tothe DHFRcoding
region
atAUG-1(pED1)
resulted inan mRNA in which themajor site of initiation was EMCV AUG-11; however, initiation at the downstream DHFR AUG-2also occurred (Fig. SA, lanes 14 and 15). Similarly, initiation at two neighboring AUGs occurred in two
con-structs in which AUG-11 was linked to DHFR AUG-1 by either a 12-bp XhoI linker (pED2) or 9 bp of the EMCV leader sequence between AUG-11 and AUG-12 (pED3). The occurrenceofinitiation at more than one AUG could be due
totheproximity of the IRES (atorvery nearAUG-11) to the
initiation codon AUG-11. In cap-dependent translation,
ini-tiation at downstream AUG codons may occur when there
are fewer than 10 nt between the 5' cap structure and the initiation codon (23). Forexample, initiation at two neigh-boring codons occurs in eukaryotic mRNAs, such as the Ia antigen-associated invariant chain (37) and the BA3/A1 crystallin polypeptide (34), in which the 5' UTRs are very short, less than 7 bp. Initiation on mRNA from pED3, in which the EMCV AUG-12 was fused to DHFR AUG-1, occurred at both EMCV AUG-11 and EMCV AUG-12/
DHFR-1. Mutation of the EMCV AUG-11 (in pED3) to an AUU resulted in a unique initiation event at EMCV AUG-12/DHFR AUG-1 (equivalent tothe level of initiation from EMCV AUG-11). In addition, initiation at downstream AUGs was not detected (data not shown). These results showthatAUG-11 isnotrequired for internalinitiationand suggest that AUG-11 may be too close to the putative ribosomal entry site, of which one part may be the Yn tract. Increasing the distance from the Yn tract to the first AUG codon resulted in initiation at a unique site. Thisfinding is consistent with results from the poliovirus UTR in which a 72-bp insertion between the Yn tract and the first AUG codon didnotreduce viralinfectivityaslongastheinserted sequencelacked anAUG codon(24).
The utilization of downstream AUGs became enhanced when the sequencecontextupstreamof the EMCVAUG-11
was made less favorable by mutation ofA to T in the -3
position. In the presence of the A-to-Tmutation,ribosomes
appearedto bypassthe EMCV AUG-11 and initiated at the
next downstream AUG, making that AUG the preferred
initiationsite. Ourexperimentsdonotdistinguish betweena
mechanism involving leaky scanning (23) andone in which the ribosome binds to the EMCV leader and jumps to a
downstream AUG codon. However, thedetection of
poly-peptides initiated at three downstream AUG codons even
though all of these AUGs are in favorable context for
initiation (21) is suggestive of amechanism involving
ribo-some jumping. Initiation at several AUG codons occurs
frequently in viral mRNAs, allowing the synthesis of two
proteins from different initiation codons. Seventeen viral
mRNAs (22) have each been shown to produce two or
sometimes three different
proteins
throughscanning.
It isinteresting to note that direct sequence analysisofisolated
EMCV RNA identified the sequence around EMCVAUG-11
asAUUAUAUG (31). SubsequentDNA sequence analysis
of all infectious clonesyielded the sequenceAUAAUAUG (6). Thus it appears that the A in
position
-3might
beimportant for virusviabilityand supports theimportanceof
afavorablecontextfor EMCVAUG-11 (30a).
The sequence downstream of EMCV AUG-11 was also found to influence the utilization ofAUG-11aswell asthe downstream AUG. Thiswas mostobviousbycomparingthe
useof EMCVAUG-11 andDHFRAUG-1 inpED3,inwhich
12 bp coding for the first four amino acids of the EMCV
polyprotein precedes DHFR AUG-1, with the use of the
same codons in pED2, in which the 12-bp sequence was
replacedby a 15-bp linker separating EMCV AUG-11 and
on November 9, 2019 by guest
http://jvi.asm.org/
DHFRAUG-1 (Fig. 2). BothAUGs are used fourfoldmore efficiently in pED3. We do not know whether the RNA
sequence, secondary structure,orthe particular codonsare important in affecting the increased AUG utilization
ob-served. Our data suggestthatEMCV AUG-10is notused in
the intact viral RNA, consistent with in vitro studies of Kaminski et al. (14). Our mutagenesis studies show that EMCVAUG-10 and thenucleotide sequencebetweenitand EMCV AUG-11 are not required for internal initiation. A 12-bp linker replacement of nucleotides after EMCV
AUG-10 (pED5) allowed detectable initiation at EMCV AUG-10 and DHFR AUG-2, whereas no detectable
initia-tion occurred at DHFR AUG-1. Mutation of AUG-10 to
ATTyielded a large increase in initiation atDHFR AUG-1
with lessinitiationatDHFR AUG-2, althoughthesequences between AUG-10 and DHFRAUG-2 were identical in the
two constructs. Analysis of the secondary structure which
could be formed within the 40 bp extending from the pyrimidine stretch to DHFR AUG-1 (using the FOLD pro-gram[10])revealed thatthelinkerreplacementafter AUG-10
generateda stem-and-loopstructure of AG = -6.5 kcal (ca.
-27.2 U). The mutation ofAUG toAUU destabilizes this stem-and-loop structure toAG = -1.0kcal (ca. -4.2 U). It
is difficult to know how this small change may affect the structureofthe EMCVIRES. This stem-and-loopstructure
maypreclude theuseofDHFR AUG-1 andmayaccountfor thedetectable useofEMCVAUG-10. Since substitution of the nucleotides between AUG-10 and AUG-11 did not re-duce internal initiation, the specific sequence between
AUG-10 andAUG-11 is notnecessaryfor internal initiation as long as secondary structure is absent and a favorable
context forAUG initiation is present (21).
Theintroduction ofanopenreading frameupstreamof the EMCV leader did not affect DHFR expression. However, translation ofthesecond cistronfollowing the EMCVleader
was not as efficient as the translation of the first cistron following the tripartite leader. We do not know the reason and speculate that the two leaders are competing for
com-moninitiation factors. EMCV dicistronicmRNA expression
vectors encoding an mRNAwith DHFR in the 3' position
andeithercontainingorlacking the EMCV leaderupstream
of DHFR were introduced into CHO cells. DHFR
expres-sion,asmeasuredby the numberof cell coloniessurviving in nucleoside-free medium and with increasing amounts of
MTX, was 10- to 50-fold higher for the EMCV-containing vectors. The level of DHFR expression from these vectors
followed the same order as in the COS-1 cell transient expression system. These findings indicate that cellular factors important for internal initiation from the EMCV leaderarealso presentin CHO cells.
PolycistronicvectorsutilizinganEMCV leader-selectable
marker gene cassette downstream of the gene of interest have been used to derive stable cell lines expressing high levelsof thedesiredgeneproduct (15, 18,40). The
introduc-tion of the EMCV leader in T7 expression vectors has allowedmuchhigher level oftranslationof thetargetgenein the vaccinia virus/bacteriophage hybrid expression system
(7). Retroviral dicistronic vectors incorporating either the poliovirus leader or theEMCV leaderbetween two coding
regions havebeenshown toallow coordinate expressionof
thetwogeneproductsfromasinglemRNA(1).Astheuseof
theEMCVleaderinexpressionvectorsis likelytoincrease,
ourstudiespointouttheimportance of designingafavorable
junction between the EMCV leader and the downstream
gene in order to maximize expression of that gene and
minimizedownstream initiation.
ACKNOWLEDGMENTS
We thank Ann C. Palmenberg for use of the EMCV leader and for providing its complete sequence prior to publication. We are grate-ful to Andrew J. Dorner and Clive R. Wood for valuable comments onthemanuscript and to Joanne Janeiro for help in its preparation.
REFERENCES
1. Adam, M. A., N. Ramesh, A. D. Miller, and W. R. A. Osborne. 1991. Internal initiation of translation in retroviral vectors carrying picornavirus 5' nontranslated regions. J. Virol. 65: 4985-4990.
2. Beck, E., S. Forss, K. Strebel, R. Cattaneo, and G. Feil. 1983. Structure of the FMDV translation initiation site and of the structural proteins. Nucleic Acids Res. 11:7873-7885. 3. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W. J.
Rutter. 1979. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18:5294-5299.
4. Derman, E., K. Krauter, L. Walling, C. Weinberger, M. Ray, and J. R. Darnell. 1981. Transcriptional control in the produc-tion of liver-specific mRNAs. Cell 23:731-739.
5. Dorner, A. J., B. L. Semler, R. J. Jackson, R. Hanecak, E. Duprey,andE. Wimmer. 1984. In vitro translation of poliovirus RNA:utilization of internal initiation sites in reticulocytelysate. J. Virol. 50:507-514.
6. Duke, G. M., M. A. Hoffman, and A. C. Palmenberg. 1992. Sequence and structural elements that contribute to efficient encephalomyocarditis virus RNA translation. J. Virol. 66:1602-1609.
7. Elroy-Stein,O., T. R. Fuerst, and B. Moss. 1989. Cap-indepen-dent translation of mRNA conferred by encephalomyocarditis virus 5' sequence improves the performance of the vaccinia virus/bacteriophage T7 hybrid expression system. Proc. Natl. Acad. Sci. USA 86:6126-6130.
8. Howell, M. T., A. Kaminski, and R. J. Jackson. 1990. Unique
featuresofinitiation ofpicornavirus RNA translation, p. 144-151. In M. A. Brinton (ed.), New aspects of positive-strand RNAviruses.American Society for Microbiology, Washington,
D.C.
9. Jackson,R.J.,M.T.Howell,and A. Kaminski. 1990. The novel
mechanism of initiation of picornavirus RNA translation.
TrendsBiotechnol. 15:477-483.
10. Jacobson, A. B., L. Good, J. Simonetti, and M. Zuker. 1984. Somesimplecomputational methods to improve the folding of
large RNAs.NucleicAcidsRes. 12:45-52.
11. Jang, S. K.,M. V. Davies, R. J. Kaufman, and E. Wimmer. 1989. Initiation ofprotein synthesis by internal entry of
ribo-somes into the5'nontranslated region ofencephalomyocarditis
virus RNA invivo.J. Virol.63:1651-1660.
12. Jang, S.K., T.V.Pestova, C.U. T.Hellen,G. W. Witherell, and
E.Wimmer. 1990. Cap-independenttranslationof picornaviral
mRNAs:structure andfunctionof the internal ribosomal entry
site. Enzyme44:292-309.
13. Jang, S. K., and E.Wimmer.1990.Cap-independenttranslation
of encephalomyocarditis virus RNA: structural elements of
internal ribosomal entry site and involvement of a cellular
58-kDaRNA-bindingprotein.GenesDev.4:1560-1572. 14. Kaminski,A.,M.T.Howell,and R.J. Jackson. 1990. Initiation
ofencephalomyocarditisvirus RNAtranslation: the authentic
initiation site is notselectedbyascanningmechanism. EMBO
J. 9:3753-3759.
15. Kaufman, R. J.1990. Strategies forobtaininghigh level
expres-sion inmammaliancells.Technique 2:221-236.
16. Kaufman, R. J. 1990.Vectorsusedforexpressionin mammalian
cells. Methods Enzymol. 185:487-511.
17. Kaufman, R. J., M. V. Davies, V. K. Pathak, and J. W. B.
Hershey. 1989. The phosphorylation stateofeucaryotic
initia-tion factor 2a alterstranslational efficiencyofspecificmRNAs.
Mol.Cell. Biol.9:946-958.
18. Kaufman, R. J., M. V. Davies, L. C.Wasley,and D.Michnick.
1991.Improvedvectorsforstableexpression offoreign genes in
mammalian cells by use of the untranslated leader sequence
from EMC virus. Nucleic Acids Res. 19:4485-4490.
on November 9, 2019 by guest
http://jvi.asm.org/
19. Kaufman, R. J., and P. A. Sharp. 1982. Construction of a modulardihydrofolatereductase cDNA gene: analysis of signals
utilized forefficient expression. Mol. Cell. Biol. 2:1304-1319. 20. Kozak, M. 1984. Compilation and analysis of sequences
up-streamfromthe translational start siteineukaryotic mRNAs.
Nucleic Acids Res. 12:857-872.
21. Kozak, M.1986.Point mutationsdefinea sequenceflanking the AUG initiatorcodon thatmodulates translation byeukaryotic
ribosomes. Cell44:283-292.
22. Kozak, M. 1986. Bifunctional messenger RNAs ineukaryotes. Cell 47:481-483.
23. Kozak, M.1989. Thescanning modelfortranslation:Anupdate. J.Cell Biol. 108:229-241.
24. Kuge, S., N. Nawamura, and A. Nomoto. 1989.Genetic variation occurring on the genome of an in vitro insertion mutant of poliovirustype1.J. Virol.63:1069-1075.
25. Kuge, S., and A. Nomoto. 1987. Construction of viable deletion and insertion mutants of the Sabin strain type 1 poliovirus:
function ofthe 5' noncoding sequence inviral replication. J.
Virol. 61:1478-1487.
26. Kuhn, R., N. Luz, and E.Beck.1990.Functionalanalysis of the internal translation initiation site of foot-and-mouth disease virus.J.Virol. 64:4625-4631.
27. Laemmli, U. K. 1970.Cleavage of structural proteins duringthe
assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
28. Luz, N., and E. Beck. 1991. Interaction of a cellular
57-kilodalton proteinwith theinternaltranslation initiationsite of
foot-and-mouth disease virus.J. Virol. 65:6486-6494.
29. Meerovitch, K., J. Pelletier, and N.Sonenberg. 1989. A cellular
protein that bindstothe5'-noncoding regionofpoliovirusRNA:
implications for internal translation initiation. Genes Dev.
3:1026-1034.
29a.Michnick,D.Unpublished data.
30. Morinaga,Y., T.Franceschini,S.Inouye,and M.Inouye.1984.
Improvement ofoligonucleotide-directed site-specific
mutagen-esis using double-stranded plasmid DNA. Bio/Technology 2:636-639.
30a.Palmenberg,A. Personal communication.
31. Palmenberg, A. C., E. M. Kirby, M. R. Janda, N. L. Drake, G. M. Duke, K. F. Potratz, and M. S. Collett. 1984. The nucleotide and deduced aminoacid sequencesof the encephalo-myocarditisviralpolyprotein codingregion.NucleicAcidsRes. 12:2969-2985.
32. Pelletier, J., G. Kaplan, V. R. Racaniello, and N. Sonenberg.
1988. Translational efficiency of poliovirus mRNA: mapping inhibitorycis-acting elementswithin the 5' noncoding region. J.
Virol. 62:2219-2227.
33. Pestova, T. V., C. U. T.Hellen,andE.Wimmer. 1991.
Trans-lation of poliovirusRNA:role of an essential cis-acting oligopy-rimidineelement within the5'nontranslatedregionand
involve-mentof acellular57-kilodaltonprotein.J.Virol.65:6194-6204. 34. Peterson, C. A., and J.Piatigorsky. 1986. Preferential
conserva-tion oftheglobular domainsof the,BA3/A1-crystallin
polypep-tide of the chickeneyelens. Gene 45:139-147.
35. Sangar,D.V.,S. E.Newton, D. J. Rowlands, and B. E. Clarke.
1987.Allfootand mouth diseasevirusserotypesinitiate protein
synthesis attwoseparateAUGs. Nucleic Acids Res.
15:3305-3315.
36. Sanger,F., S.Nicklen, and A. R.Coulson. 1977.DNA
sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.
USA 74:5463-5467.
37. Strubin,M., E.0. Long, and B. Mach. 1986. Two formsofthe Ia antigen-associated invariant chain result from alternative
initiations attwoin-phase AUGs.Cell 47:619-625.
38. Thomas, P. 1980. Hybridizationofdenatured RNA and small DNAfragmentstransferredtonitrocellulose.Proc. Natl.Acad. Sci. USA77:5201-5205.
39. Wood, C. R., A. J. Dorner, G. E. Morris, E. M. Alderman, D. Wilson, R. M.O'Hara, Jr., and R. J. Kaufman. 1990.Highlevel
synthesisofimmunoglobulinsinChinese hamster ovary cells. J.
Immunol. 145:3011-3016.
40. Wood, C. R., G. E. Morris, E. M.Alderman, L. Fouser, and R. J. Kaufman.1991.Aninternal ribosomebinding sitecanbe usedtoselect forhomologous recombinantsat an
immunoglob-ulin heavy-chain locus. Proc. Natl. Acad. Sci. USA 88:8006-8010.