0022-538X/90/062457-10$02.00/0
Copyright © 1990, American Society for Microbiology
The Adenovirus ElI Early Promoter Has Multiple
EIA-Sensitive
Elements, Two of Which Function Cooperatively in
Basal and
Virus-Induced Transcription
CHITRAF. MANOHAR, JON KRATOCHVIL,t ANDBAYAR THIMMAPAYA*
Departmentof Microbiology and Immunology, Northwestern UniversityMedical School, 303 EastChicago Avenue, Chicago, Illinois 60611
Received 8 November 1989/Accepted 6 February 1990
Themechanism by which the adenovirus-encoded nuclearoncogene EIA activates transcriptionofseveral viral and hostpromotersisanimportantissue in the regulation of eucaryoticgeneexpression and virus-host
cell interactions. Identffication of cis-acting elements of the promoters and the cognate host transcription factors that are targets for EIA action is crucial for our understanding of the EIA-mediated control of coordinately regulatedgenes.The adenovirusEIIearlypromoterhasacomplex architecture and containstwo overlapping promoters with start sites at +1 (major promoter) and -26 (minor promoter). The major promoter responds strongly to virus-encoded trans activators EIA and EIV and contains four elements: a TAGA motif analogoustotheTATAbox,twoEIIFsitespresentinaninverted orientation, andanATF/CREB site. To determine precisely the roles played by these cis-acting elements in both basal and virus-induced transcriptionwhen thepromoter is situated in its natural context,weinvestigatedthephenotypeofaseries of linkerscanpromotersubstitutionmutantsinserted into theviral chromosome. Promoterconstructsharboring linkerscanmutationsineach elementwererebuiltintoanovel EIA- adenovirus vector,andtranscriptional activitywasmonitoredinvirus-infected cells.Inthe absenceof virus-encodedtransactivators, basalactivity in vivowasdependentonallfour cis-acting elements. Surprisingly, apromoter mutantwithonlyoneofthetwo ETTF sites intact could not promote transcription in vivo, suggesting that the two ETTF sites function cooperativelyeveninbasaltranscription. Promotersharboringmutations in either of these twoEIIF sites also failed tobindtoaninfection-specific form of EIIFingel shiftassaysandcompeted onlyveryweaklyfor EIIF bindingwith thewild-type promoterfragment. The dramatic cooperativity shown bythe twoinverted EIIF sites of the ETT promoter both in vivo and in vitro could reflect simultaneous contact of both sites by the transcription factorEIIF.Furthermore, promotermutantswith mutations in theTAGA motif,thetwoETTF sites, and the singleATF site all failed torespondtovirus-encodedtrans activators. Whereasrecent results demonstrate that EIIF activity can be modulated independently by EIV, leading to transactivation of this promoter, ourresultsand those published previously strongly indicatethat thethree different transcription
factors that bind toTAGA,EIIF,and ATFmotifs of the ETT earlypromoterareall targets forEIA regulation
invivo.Thus,strongtransactivation of the EII earlypromoterthrough these multiple EIA-sensitive elements andindependently bytherecently discovered EIV pathwaysuggeststhat theEIIearlypromoterisstringently regulated in virus-infected cells. Suchastringent regulation of thispromoteris consistent withthevitalroles played by the threegeneproductsof thistranscription unit inthevirallytic cycle.
Oneimportant step in theregulation of cellular andviral gene expression isat the levelof initiation oftranscription. Inmost cases, this is determinedby the type of cis-acting elements and the way they are organized in a given pro-moter, the host transcription factors that bind to these sequences, and the virus- orhost-encoded trans-acting fac-tors that stimulate
transcription
ofthese promoters. Early promoters of human adenovirus provide excellent model systemsin which to study thistype of transcriptionalregu-lation. Inhumancellsinfected with adenovirustype 2 (Ad2) orAdS, six early polymerase
IT
promoters arecoordinately regulated, and their efficienttranscription
is dependent on the289-amino-acid viral EIAprotein (reviewedin reference53). Most ifnotall upstream DNA sequence elements that are required for basal EIA-independent transcription of these promoters have been defined(reviewedinreference5), along with several host transcription factors that bind to
* Correspondingauthor.
tPresent address: Abbott Laboratories, North Chicago, IL 60064.
them (reviewed in reference 30). Yet despite intensive
ef-forts, no uniform mechanism to explain how either these
well-defined promoters ortheEIA-sensitive polymerase III promoters(25, 71) arestimulated hasemerged.
Forthe pastseveral years,ourlaboratoryhasfocusedon the structure and mechanism of activation of one of the
EIA-activated promoters, the ETT earlypromoter. The EII
unit is one ofthe most important of the early regions for successful completion ofvirus infection. It encodes a 72-kilodalton(kDa)DNA-binding protein (DBP),140-kDa DNA
polymerase, andthe 80-kDa precursorterminal
protein,
all of whicharevitalfor viral DNAreplication (66).Inaddition,several otherimportantmetabolic roles areassociated with DBP, such as mRNA stability (2), repression of EIV
tran-scription (54), host range function (33), assembly of virus
particles(55), andtransformation
(15).
Astringent
transcrip-tional control mechanism to ensuresynthesis
ofoptimum
amountsof these crucialproteins
duringthegrowth
cycle
isexpected. TheEIIearly promoteris also sensitiveto trans-activationby simian virus 40large-Tandsmall-t
antigens
in additiontoadenovirus EIAproteins (46-48).
This promoter 2457on November 10, 2019 by guest
http://jvi.asm.org/
is complex and contains two overlapping promoters with start sites at + 1 (major promoter) and -26 (minor promoter) (49). By linker scan (LS) mutagenesis studies, weand others (52, 72, 73) have shown that the major EII early promoter contains two cis-acting sequence elements: aTAGA motif between -22 and -30, analogous to theTATAbox, and an ATF-binding site located upstream between -68 and -77. This element is also found in the adenovirus EIII and EIV promoters and in a number of hostpromoters thatrespond to cyclic AMP (8, 11-13, 21, 26, 41, 42, 45). A host transcrip-tion factor, ATF, that binds to this sequence has been identified (28, 37, 41, 42, 63, 70). This factor has been purified (19, 29; C. F. Manohar et al., unpublished results), and its binding activity does not change aftervirus infection (28, 63, 70). Gel shift experiments have shown that a host protein designated EIIF binds to a third sequence element,
TTTCGCGC, duplicated between -35 and -68. A host 55-kDa polypeptide binds to this sequence and promotes transcription in vitro (69). The promoter-binding activity of this factor increases severalfold aftervirusinfection(34, 64, 70), and recent results show that this dramatic increase in binding activity is due to amodification ofEIIFinduced by one of the EIVpolypeptides (10, 20, 22, 25a, 60). Although the effect of EIV polypeptide on EIIF has recently been extensively analyzed at thebiochemicallevel, thefunctional
significance of these twocopiesof thesequence elementand genetic evidence fortheirrole in EII early promoter activa-tion havenot beenclearly established. Inaddition, transfec-tion studies have not provided unambiguous evidence for any sequenceelements in theElI earlypromoterthat can act as targets for EIA activation. In LS mutagenesis studies,
everymutantpromoter that was analyzed fortranscription
wasstimulated by EIAgeneproducts intransfection assays
(48,52, 72). Ontheother hand,certain deletionmutagenesis
studies haveimplicated, at least inpart, a role for the EIIF sites in EIA transactivation ofthe EII early promoter (27, 73), buttheroles ofother sequence elements in EIA trans-activation have notbeen resolved. Thisraises the question
as towhethertranscriptional regulation studied in transient
transfection assaysfaithfully reproduces the regulation that goes on in virus-infected cells when the promoters are present in their natural context.
Toaddress thisquestion, we havereturned a series of LS substitution mutants tothe viralchromosome, using anovel
EIA- adenovirus vector, and analyzed their transcription
with or without coinfection with a wild-type (WT) virus to provide EIA polypeptides. Results presentedhere andthose
publishedpreviously lead us to conclude that the EII early
promoter in virus-infected cells is regulated by three dif-ferent transcription factors interacting with four separate
DNAmotifs, all ofwhich appear to beinfluenced byEIA.
MATERIALS AND METHODS
Sources for HeLa suspension cultures, WT adenovirus
and thed1321 variant, plasmidsthatcontainthe WTAdS
EIl
early promoter fused to chloramphenicol acetyltransferase (CAT)-coding sequences(EIIA-early WTCAT),and the LS mutant derivatives of this plasmidwere as described previ-ously (52, 63,64).Aplasmidthatcontainedtheright terminal
portion from 78.5 (XbaI)to 100 map units of the adenovirus sequences (pA5-130), which was used to rebuild the pro-moter constructs into virus, was described in a previous report (6). Mutant d1321 contains the same deletionasdoes
d1312(6, 31).
The promoter mutants were first rebuilt into the EIII
region ofpA5-130. The viralportion oftheplasmidwasthen
ligated witha0- to78.5-map-unitXbaIfragmentof d1321 as described before (see Fig. 2; 6). Human 293 cells were
transfected with theligated DNA sample, and virus stocks werepropagated. Thevirusstocks were plaquepurified and
titeredon293 cells. The mutationswereconfirmedagainby analyzing mutantviralDNAas described previously (6).
Nuclear extracts fromvirus-infected cellswereprepared
byinfectingHeLacellsuspensionculturesat25PFU per cell
for 8 h in the presence of
1-p-D-arabinofuranosylcytosine
(araC) at 25
pg/ml
(9,63).
For CAT assays, virus infection wascarriedoutfortheperiods indicatedinthe figurelegends in the presence of araC (25 ,ug/ml). Protein concentrations were determined by the method of Bradford (7). Gel shift assays were performed as reported previously (64), with salmon sperm DNA as the nonspecific competitor. Thisnonspecific competitor DNA allowed detection of EIIF
factorspecifically (64, 70).
Primer extension was carried out according topublished protocols(1).Briefly, 15 pug ofpoly(A)+ RNAs was annealed with a 5'-end-labeled CATprimer (from +110 to +136)
(105
cpm)in30,ul ofabuffercontaining80%formamide, 40.0 mM piperazine-N,N'-bis(2-ethanesulfonic acid) (PIPES; pH6.4), 400.0 mM NaCl, and 1.0 mMEDTA overnight at 30°C. The nucleic acids were ethanol precipitated and suspended in abuffercontaining50 mM Trishydrochloride(pH 8.0), 40 mM NaCl, 0.5 mM EDTA, 5 mMMgCl2, 3 mM dithiothreitol, 40
,M deoxynucleoside triphosphates, and 1.25 [L of RNasin. Avian myeloblastosisvirus reverse transcriptase (20 to 40 U) was then added, and the sample was incubated for 3 h at
42°C. At the end of incubation, reaction was stopped by
addingRNase A(40pug/ml)and EDTA (20 mM). The nucleic acids were extracted with phenol, ethanol precipitated, and resolved on a6% DNA-sequencing gel. B-Actin mRNA was quantitated under identical conditions with 5.0 ,ug of
poly(A)+ RNA, using a 27-nucleotide (nt) primer comple-mentaryto +78 to +105 of the human,B-actin gene.
RESULTS
Strategy to analyze theEIlearly promoter inthe context of viral chromosome. Figure 1 shows the four transcriptional control elements that have been identified for the ElI early promoterwith respect to the major start site by this labora-tory and others (52, 63, 64, 70, 72). We previously con-structed and characterized intransfection assays a setof 15 LS substitution mutants of the EII early promoter fused to the CAT gene (52). Figure 1B shows the locations of these mutations with respect to different transcriptional control elements. We have now returned these LS mutants into the viral chromosome, using a novel EIA-deficient adenovirus vector system (6). In this vector system, the ElI promoter mutants were cloned into thenonessential EIIIregion of the virus, with the orientation of transcription of the newly introduced ElI promoter constructs identical to that of the resident ElI gene (Fig. 2). The original EII gene, which provides functions vital for viral DNAreplication, remained intact. Since the promoter sequences are fused to the re-porter CAT gene, the mutant promoters can be analyzed unambiguously in a sensitive CAT assay. The virus vector contained an EIA deletion identical of that ofd1312, so the virus could be propagated efficiently in the adenovirus-transformed human 293 cell line by trans complementation. Mutant viruses that harbor the LS substitution mutations in the ElI promoter can be analyzed for EIA-dependent and EIA-independent transcription by infecting HeLa cells with and without, respectively, the WT virus.
on November 10, 2019 by guest
http://jvi.asm.org/
-76 -69 -67 -60 -29 -23
TGACGTAG TTTCGCGC TTAAGA
-~---m
rAN EIIF -- 1 TG
-50 4'.
CGCGCTTT
-45 -36
ATF EIIF(D)
-74/-85 -52/-63 -55/-66 ATF/EIIF(D)
-63/-73 -65/-75
EIIF (P) -31/-41 -35/-46 -40/-50
TAGA -19/-29 -25/-34
FIG. 1. Transcriptional control elements of the Ad5 EII early promoter, theircognate transcription factors, and the LS substitution mutations that mutate these defined control elements. (A) ElI earlypromoterstructure.Locations ofasequenceanalogousto the TATA
sequence(TAGA motif), twoEIIFsitespresentinaninverted orientation, andasingle ATF site and thetranscription factors that bindto thesesequences areshown. Althoughnotidentified, it is presumed thatatranscriptionfactor bindstotheTTAAGAsequence.(B)Locations
of theLSsubstitutionmutations in relationtothedifferent cis-acting elements. EIIF-P, Proximal EIIF site; EIIF-D,distal EIIF site. Further details of thesemutantscanbeobtained from Murthyetal. (52).
Basaltranscriptional activity of themutant promotersand the cooperativity between the two EIIF sites for function in vivo. To determine the contribution of each cis-acting ele-mentofthe ElI earlypromoterfor basaltranscription, HeLa cells were infected with adenovirusmutants containing the WTandmutant derivatives ofthe EII-early CATconstruct (Ad5EII-EWT-CAT)at20 PFUpercellandinthepresence
ofaraC. Eight hours after infection, cell lysates were pre-pared and CAT activity present in the lysates was deter-mined as described previously (6, 17). The CAT activity
detected incellsinfectedwithmutantviruses harboring the EII-early CAT constructs was solely due to transcription driven from the EII early promoter. Because, as shown previously (6) andinthis study(Table 1; Fig. 3A, lane 1; data notshown), amutantvirus (Ad5 0-CAT) in whicha promot-erless CAT construct was cloned into the EIII region in a backgroundidentical to that ofthe adenovirus mutant car-rying the WT EII-early CAT construct did not show any
ClO I
100
pA5-130
Xboll
3
Xl78~5 83
Oto78-5 from virus, 78-5tolOOfromplasmid,
ligate, tronsfect HindHll BomHl EcoRI Xbal Xhol
o79,5g5 75-9 785 83 100
EIAA5
PIIA-F (%AT
-4 ^~-4 C,11A-C7k*A
EIIA
FIG. 2. StrategyforrebuildingtheEIIAearlypromotermutants
intoanEIA-Ad5 vector(see Materials and Methods for details). Restriction sites are positioned on the Ad5 map units. Symbols: X,chimericgene; _, deletionpresentin the EIAgeneof variant d1312(31). EIIA,TheoriginalElI geneof the virus.
[image:3.612.157.475.79.267.2]detectable CATactivityat8 h postinfection in the presence orabsenceofEIA geneproducts. This result indicatesthat the DNAsequences upstream of thenewEIIearly promoter didnotcontribute topromoteractivity fortuitously. Figure
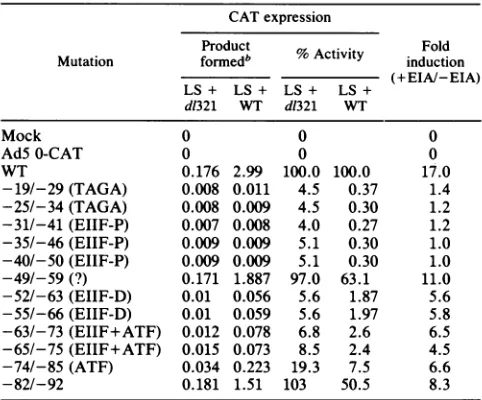
TABLE 1. CATexpression in adenovirus mutantsharboring ElI earlypromotermutationsa
CATexpression
Product Fold
Mutation formedb %
Activity
induction (+EIA/-EIA) LS+ LS + LS+ LS+d1321 WT d1321 WT
Mock 0 0 0
AdS0-CAT 0 0 0
WT 0.176 2.99 100.0 100.0 17.0
-19/-29(TAGA) 0.008 0.011 4.5 0.37 1.4 -25/-34(TAGA) 0.008 0.009 4.5 0.30 1.2 -31/-41 (EIIF-P) 0.007 0.008 4.0 0.27 1.2 -35/-46(EIIF-P) 0.009 0.009 5.1 0.30 1.0 -40/-50(EIIF-P) 0.009 0.009 5.1 0.30 1.0 -49/-59 (?) 0.171 1.887 97.0 63.1 11.0 -52/-63(EIIF-D) 0.01 0.056 5.6 1.87 5.6 -55/-66 (EIIF-D) 0.01 0.059 5.6 1.97 5.8 -63/-73(EIIF+ATF) 0.012 0.078 6.8 2.6 6.5 -65/-75(EIIF+ATF) 0.015 0.073 8.5 2.4 4.5 -74/-85(ATF) 0.034 0.223 19.3 7.5 6.6
-82/-92 0.181 1.51 103 50.5 8.3
aThecis-acting elements affected by the mutationsareshown in parenthe-ses.EIIF-P,ProximalEIIFsite; EIIF-D,distalEIIF site.Dataobtained from theexperiment shown inFig.3 arepresented.HeLa cells(60-mm-diameter dishes;2x 106cells)wereinfectedwithAdSmutantscontainingthevarious CAT constructs withWTvirus(+EIA)ord1321(-EIA)at20 PFUper cell in thepresence ofaraC(25p.g/ml)and harvestedat8 hpostinfection; thecell lysateswereprepared,and theirproteinconcentrationsweredeterminedby
the methodof Bradford(7). CAT activitywasassayedasdescribed previ-ously, (17),usingequalamounts ofprotein. Assayswere performedunder conditions in which theconversion of[54C]chloramphenicol toits products
was less than30% of thetotal substrate added. Whennecessary, the cell lysateswerediluted toobtainvalueswithin this range.
INanomolesofacetylatedchloramphenicolper106cells.SeelegendtoFig.
3forexperimental details.
on November 10, 2019 by guest
http://jvi.asm.org/
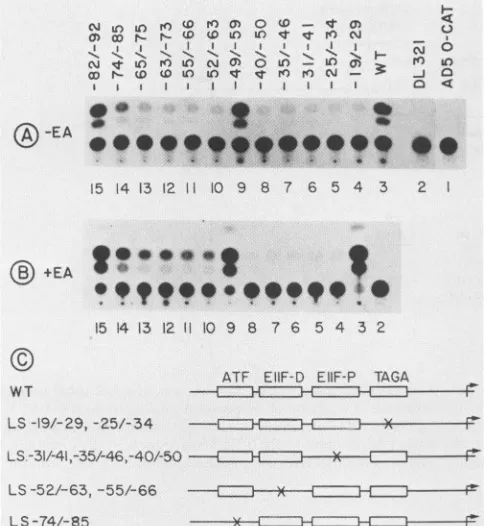
[image:3.612.318.559.423.623.2] [image:3.612.61.300.542.691.2]Cvo,£ t D O OQW- ¢
a)0rsiDa 0s ¢
\~ ~~~~r
Xoc]c
u>sS>Fr nui 4Q
(-) EA ***,,
15 14 13 12 11 109 8 7 6 5 4 3 2 1
®+EA t
15 14 13 12 11 10 9 8 7 6 5 4 3 2
WT
ATF EIIF-D EIIF-P TAGA
E5J
~LS -19/-29,-25/-34 -=C
LS.-31/-41,-351-46,-40/-50
LS-521-63, -551-66 X
[image:4.612.57.299.80.343.2]LS-74/-85
FIG. 3. Expression of the WT andthe LS derivatives of the ElI early promoter in the context of the viral chromosome in the absenceorpresenceofEIAgeneproducts. SeefootnoteatoTable 1forexperimental details. (A) CAT expressionof the WT and the LSmutants of the ElI earlypromoterin the absence of EIA. Ad5 O-CAT isan EIA- AdS variant that contains a promoterless CAT constructintheEIII region (6). (B)CATexpressionin thepresence of EIAproducts. Mutant viruseswerecoinfectedwithaWT virus. In EIA- experiments, mutant viruses were coinfected with the EIA- virus dl321. However, no difference in the pattern of CAT expressionwasobserved whetherinfectionwascarriedoutaloneor
with EIA-virus d1321. The autoradiograms shown in panelsAand Bwereobtainedby exposing the thin-layer chromatography plates for different periods of time. For quantitation, see Table 1. (C) Schematic representation ofthe mutations in the various LS
mu-tantswithrespecttodifferenttranscriptionalcontrol elements. 3Ashows CAT activities for the WT and LS mutantsat8 h postinfection in HeLa cells infected with mutant viruses; Fig. 3C showsaschematicrepresentationof the LSmutants withrespecttothecis-actingelements of thepromoter.CAT activitywasquantitated by directly countingtheradioactive spots inthe thin-layer chromatography plate. Inourearlier study, we found that the levels of hexon mRNA (internal control)didnotchangebetweenmutantinfections inasingle experiment (6). Therefore, when using infections it is not
necessary to normalize the CAT activity to an internal
control. Relative levels of CAT activity of the different promoter mutants are shown in Table 1. A 20- to 25-fold-reduced CAT activity was observed for mutants that alter the TAGAmotif(LS mutants -19/-29 and -25/-34) and the proximal (LS mutants -31/-41, -35/-46, and -40/-50) and distal (LS mutants -52/-63 and -55/-66) EIIFsites.MutantsLS-74/-85, whichmutatesexclusively the ATF site, was fivefold defective. Two LS mutants (-63/-73and -65/-75)that abutboth distalEIIFand ATF sites were also severely defective (20-fold). Thus, efficient basaltranscriptionof themajorElI earlypromoterinvivois dependent on all four identified cis-actingelements. Unex-pectedly, there was a dramatic cooperativity between the
twoEIIFsites forpromoter function.
Integrity
of bothEIIF siteswasvital fortranscription.
Forexample,
inmutantsLS-31/-41,
-35/-46,
and-40/-50, only
theproximal
EIIFsite was
mutated;
the distal EIIF site wasintact,
yet themutants were defective
by
20-fold.Similarly,
LS mutants-52/-63 and -55/-66 exhibited a 20-fold-reduced
activity
although
theproximal
EIIFsite in thesemutantswasintact.Thus,
mutations in either of the two EIIF sites reducedtranscription
dramatically,
andonesite did notcompensateforthe other. This result
strongly
suggestsahigh degree
ofcooperativity
between thetwoEIIF sitesinbasaltranscrip-tion.
Simultaneous
requirement
of both EIIF sites for efficientEIIF
binding
invitro.TheobservationthattheElI promoterrequired
synergistic
action ofthe two EIIF sites for basaltranscription
prompted
ustoinvestigate
theformation of theEIIF
complexes
in vitro moreclosely.
We showed earlierthat the nuclearextracts
prepared
from virus-infected cells at7 hpostinfection
formedDNA-protein
complexes
specific
for thetwoEIIFsites located between -35and -68
(64).
It wasalsoclearin thesestudiesthatEIIF failedtobindtoLS mutants that mutatedspecifically
either theproximal
ordistal EIIF
site, indicating
that asingle
EIIF siteprobably
was notfunctional. These
experiments
wererepeated
with twoLSmutantsthatmutated thetwoEIIFsitesindividually.
A DNA
fragment
from-17to-96wasendlabeledandused ingel
shift assays with nuclear extractsprepared
from virus-infected cells. As shownpreviously,
when the WT promoterDNAfragment
wasusedas aprobe,
three slower-moving bandsrepresenting
three different DNAprotein
complexes
were observed(Fig. 4A,
lanes 2 and12).
Themajor
band(band
I)
wasduetoEIIF,
sinceitwascompeted
against by
anoligonucleotide carrying
theEIIF
site of theEIA promoter andanunlabeledWTpromoter
fragment
but notby
anoligonucleotide
carrying
an ATF site(Fig. 4A,
lanes 3to
6;
64). By
methylation
interferencestudiesandby
useofaseriesofLSmutantsacrossthepromotersequence, weshowedearlierthatband IIIwas
nonspecific (64)
whereas band I madespecific DNA-protein
contacts with the twoTTTCGCGC motifsbetween -35 and -68. Inourextensive
study,
the appearance or theintensity
ofband II was notconsistent,
and in manyexperiments
it could notbeabol-ished with
oligonucleotides
that carry theEIA-EIIF
or the ATF sequence(Fig. 4A,
lanes 9to11).
Theintensity
ofthisband also remained
unchanged
whengel
shiftexperiments
werecarriedoutwithaWTpromoter
fragment
andincreas-ing
concentrations of nuclearextract(C.
F.Manohar and B.Thimmapaya,
unpublished results). Thus,
we believe thatthis band is a
nonspecific
band. Consistent with earlierresults,
mutantsthat alter theproximal (LS -35/-46)
orthedistal EIIF
(LS -55/-66)
site(Fig. 4B)
failed to form theEIIF
complex,
similartoresults for the WT promoter(Fig.
4A,
lanes 7 and8; 64). However,
it ispossible
that thesepromoter
mutants withsingle
EIIF sites may bind to EIIFwithamuchreduced
efficiency.
Thesesingle-site
complexes
may
migrate along
with one of thenonspecific bands;
thishas notbeen examined further.
A
simple
competition experiment
wasperformed
todeter-minewhetherpromotermutantsinwhich
only
oneof thetwoEIIF siteswasmutatedwas
biologically
active withrespecttoin vitro factor
binding.
Gel shift assays were carriedout withlabeled WTprobe
in the presence ofdifferentconcen-trations ofunlabeled DNA
fragments
corresponding
to LS mutants -35/-46 or -55/-66. LS mutants -35/-46 and-55/-66containmutations in the
proximal
anddistalEIIF
sites, respectively
(Fig.
4B).
These unlabeled promoteron November 10, 2019 by guest
http://jvi.asm.org/
H-(0 U.)
(D
(0
to,
_J 'LO9 LL wL _LJ
00MW< ~~~~LC) X~ X XX X
r!) x
O)2O
X >)C
romp.'. ('.j toNiC PO O0JI 0x0U) f "-U
WT , -551-66 WT WT
2 3 4 5 6 7 8 9 10
11A
46
8p.
5
0
z
80
CD
EIlF-D EIIF-P
WT _
LS-35/-46
LS-55/-66 X i z
z hJ
LU
a-4
sort
Q ~
40 80
FOLD EXCESS
FIG. 4. Simultaneous requirement ofthe two EIIFsitesforEIIFbindingfrom infectedcell extracts.Gel shiftexperimentswereperformed withsalmon sperm DNA (2.0 ,ug per assay) as the competitor and with 88-base-pair end-labeled DNAfragments (fromBssHII at -17to Hindlllat-96; 64)from theWT promoter orits LS derivativesasprobes. About 0.2 ngofthe3'-end-labeledprobes withorwithoutvarious specific competitor DNAs were incubated with 10
jig
of protein obtained from nuclear extracts preparedfrom virus-infected cells (see MaterialsandMethods). I, II,and III denote the three different DNA-protein complexes detectedinour assays and arediscussed in thetext. Inexperiments shown inpanelC,complexIIwasdetected as a faint band, and therefore itsposition isnotindicated. (A) EIIFcomplexes formed in gel shiftassayswith theWTand thetwoLSmutantprobes -35/-46 and -55/-66.Competitor oligonucleotides: EIIF,5'-GATC GTTTGGCCATTTTCGCGGGAAAACTG-3' (nucleotidesequencefrom-234 to -217ofthe EIApromoter); ATF,5'-GATCCTAAAAAA TGACGTAACGGTTAAAGTC-3'(nucleotide sequence from -57 to -31 of the EIV promoter).Oligonucleotideswereusedasduplexeswith theappropriate complementary sequence. Lanes: 1,probe withoutprotein;2 to 6, assayswithWTprobeand withEIIF or ATF sequences ascompetitors; 7 and 8, assays with LS -35/-46 and -55/-66 probes, respectively, and with no specific competitor DNAs; 9 to 11, competition byEIIF or ATF sequencesforthe DNA protein complexes detected when theLS -55/-66probewasused; 12, control. (B) Diagram showing the mutationspresentin probes LS -35/-46andLS -55/-66 withrespectto two EIIFsites.(C) Competition byWTand mutantpromoterfragments derived from LS -35/-46andLS -55/-66on EIIFbinding.Assayconditionswere asdescribed above. Lane 12, Competition by a 90-base-pair pBR322 DNA fragment. (D) Graph showing the competition effects ofWT(0) and LS -35/-46 (0) promotersforEIIFbinding. EIIFcomplexes detectedinthe gel showninpanelCwereexcisedand counted and thenplottedaspercentage WTprobebindingversusfold molarexcessof the unlabeled DNAfragments.fragments contained one of the two EIIF sites intact yet
failedtocompeteefficiently withthe WT promoter (Fig. 4C, lanes 5 to 11;Fig.4D). Forexample, at a 5-fold molar excess
ofthe unlabeled WT promoter fragment EIIF binding was
nearly abolished,whereasat a7.5-foldmolar excessofDNA
fragments derived from mutant LS -35/-46 or -55/-66, EIIFbinding was notsignificantly affected (Fig. 4C,lanes 5 and 9;Fig.4D). (For reasons that we cannotexplain,the WT promoter also competed with the nonspecific band in this experiment [Fig. 4C, lanes 3 and 4].) Competition was detected only when the mutant promoters were used at a 35-fold molarexcess(Fig. 4C,lanes 6 and10;Fig.4D). EIIF
bindingwasnotabolishedeven at a105-fold molarexcessof mutant DNA fragments; 25% ofWT binding was still ob-served at this concentration (Fig. 4C, lane 8; Fig. 4D),
indicatingthat asingle EIIF site binds to EIIF witha very low affinity. This observation combined with the fact that these mutants inwhich only one of the EIIF sites is intact retained less than5%of the basaltranscription (Fig. 3A and
Table 1) indicate that the ElI early promoter shows a simultaneous requirement for both the sites to function in
vivoandin vitro. These resultsareconsistentwith therecent results ofHardy et al., who showed the dramatic
cooper-ativity ofthetwo EIIF sites for EIIF
binding
in vitro(22).
Roleofcis-actingelements inthe induction oftranscriptionby virus-encoded trans activators. One ofthe aims of this
studywas to determine
precisely
the contributions ofeachcis-acting element oftheElI
early
promoterwhen the EIA protein was provided in cells through the natural infection process rather than the transfection approach. Since our mutantslack EIAcoding sequences, we could evaluate the effect of EIAby coinfecting the mutants with a WT virus. This analysis has become somewhatcomplicated by
the recent finding that in virus-infectedcells,
thecooperative
interactionof EIIF with two EIIF sites is facilitatedby
the EIVE6/7polypeptide(25a).Ourvector wasdesigned
before this newobservation and contains an intact EIV.Coinfec-tion with WTAd5 or
d1356,
a viable variant that lacks theIi
.1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.145.473.79.376.2]E6/7 polypeptide, will influence the EIT early promoter activity through EIIF. However, this effect is confined to only EIIF. This and other previously published results (see Discussion) have allowed us to evaluate the EIA effect on the different cis elements of thepromoter.
[image:6.612.321.553.77.509.2]HeLa cells were infected in the presence of araC with mutantviruses harboring CAT constructs alongwith WT or d1321, the parental virus used to construct these mutants (6). At 8 h postinfection, cell lysates were prepared and CAT activity was determined. Figure 3B shows CAT activities detected for various mutants in the presence of WT virus. Table 1shows a comparison of promoteractivities ofvarious mutants for basal and virus-induced transcription. As ex-pected, theWT promoter was induced 17-fold in the exper-iment shown in Fig. 3B. Surprisingly, mutant promoters with mutations in the TAGA box (LS mutants -19/-29 and -25/-34) and the proximal EIIF site (LS mutants -31/-41, -35/-46, and -40/-50) were not induced by virus-encoded transactivators in any of ourexperiments(Fig. 3B andTable 1). Transcription of mutants with mutations in the distal EIIF site (LS mutants -52/-63 and -55/-66) and the ATF site (LS -74/-85) were induced five- to sixfold. These data were highly reproducible in numerous experiments. There was no difference in the pattern of expression when WT virus wasreplaced by anAdS mutant that synthesized only the largeEIA protein(50) (data not shown). One mutant with a substitution mutation located between the two EIIF sites showed normal phenotype with respect to both basal and virus-induced transcription (Fig. 3B and Table 1). Wedid not see an overexpression of CAT in this mutant as we did in transfection assays (52).
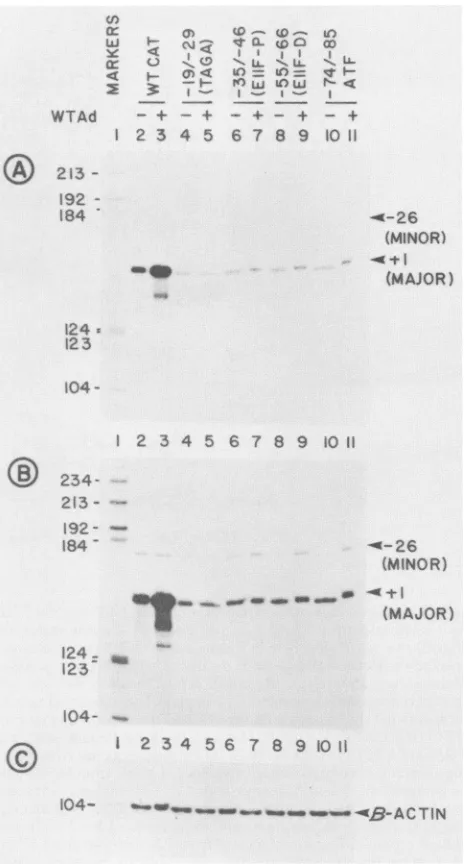
As stated earlier, the ElI early promoter contains dual overlapping promoters with a minor start site at -26. The promoteractivity in this study was quantitated onthe basis
of expression of a reporter gene. It was therefore possible that a five-to sixfold stimulation oftranscription by the ATF and thedistal EIIF site mutantsin WTcoinfectionswasdue to CAT activity resulting from RNAs transcribed from the -26 startsite; the distalEIIF site and the ATF site could be as important as theTAGA and theproximal EIIF motifs for themajor promoter in virus-inducedtranscription. Toclarify this point, we carried out primer extension analysis of CAT mRNAstranscribed from Ad5 mutants that contained muta-tions in the TAGA motif, the two EIIF sites, and the ATF site. HeLa cells were infected with mutant CAT viruses containing mutations in individual control elements and coinfected with WTAdS or the EIA- parental virus, d1321. Poly(A)+ RNAs were prepared and annealed with a 27-nt-longprimer complementary to +110 to +136(with respect to the major start site) of the CAT gene. Theprimers were then extended with reverse transcriptase as described in Materials and Methods. This strategy can distinguish be-tween the mRNAs transcribed from the two overlapping promoters. The mRNAs transcribed from the major pro-moter should generate a 136-nt-long product, whereas the mRNAstranscribed from the minor promoter should gener-ate a product 162 nt long. As an internal control,
P-actin
mRNA was also quantitated from the same RNA samples,using an appropriate primer (Fig. SC) (see Materials and Methods for details). Basal transcription from the major promoterwas dramatically reduced (7- to 10-fold, as judged from densitometric analysis of the autoradiogram) for the four mutants that individually perturb the TAGA motif, the two inverted EIIF sites, and the single ATF site (Fig. 5A). When coinfected with WT Ad5, transcription of the major promoter of the WT CAT gene was induced by 10-fold,
cr-LAJ F
< I-.
WT Ad - +
123
C}
-A_<
(-
-I-4 5 CtDL
IeI
6r)=
6 7
D
-to.=
I
8 9
Il) 02
I0lI}
10 11 213
192 -184
(MINOR)
(MAJOR) 4m
4p
124: 123
104-2 3 4 5 6 78 9 1011
234-213-
-192 184
-- 4p --. -- --.
'Am
124 _
123 *,
104- ..
©
104-<-z26 (MINOR)
.MA+R (MAJOR)
i 234 56 7 810119
""00"loom0-W- "
¶B-A
CTINFIG. 5. Quantitationof CATmRNAstranscribedfrom themajor
andminor ElI early promoters in the presence orabsence ofWT AdS infections. HeLa cellswereinfected withAdSmutants(20PFU per cell) containing aWT promoter-CATconstruct orthemutant
derivativeswith WTAd5ord1321(20PFU percell)inthe presence of araC. Poly(A)+ RNAs were prepared at 8 h postinfection and annealed to aCATprimer(from +110 to +136)or ,-actinprimer
(from +78to+105).Theprimerswerethenextended withreversed transcriptase and electrophoresed as described in Materials and Methods. A5'-end-labeledHaeIIIdigestofpBR322DNAwasused as markers.(A)Effect of LS mutations onbasaland virus-induced transcription from the major start site; (B) longerexposure of the samegeltoshow theeffect of mutationsonthe minorstartsite; (C)
quantitationofP-actinmRNAs.
whereas themutantpromoters
uniformly
failedtorespond
to the virus-encoded trans activator EIA or EIV. Identical results were obtained in twoindependent
experiments;
the stimulation oftranscription
of ATF and the distal EIIF mutants for themajor
promoter was not more than 1.5-fold(quantitation
basedondensitometerscanning).
We thereforeconclude that afive- to sixfold increase in CAT
activity
for thesemutantsinthepresenceofWTAdS(Fig.
3B and Tableon November 10, 2019 by guest
http://jvi.asm.org/
1)most
likely
wasduetomRNAstranscribed from the minorpromoter.Indeed, although basal transcription from the -26
site was somewhat reduced for these mutants, the virus-induced transcriptionwasunaffected (Fig. 5B). The
P-actin
mRNA levels remained unchanged in all ofthese samples.
These results suggest that the TAGAmotifand the ATFsites
areequally sensitivetoEIAregulation.WithregardtoEIIF,
although
ourresultscannotclearly distinguish between theEIAandEIVeffectsontheEIIFsites, previously published
results clearly indicate thatthese sites are sensitive to EIA
regulation (see Discussion). Thus, we conclude that all
cis-acting
elements ofthe ElI early promoterand thecog-natehosttranscriptionfactorsaretargetsforEIAregulation.
DISCUSSION
This is thefirst time that the control elements ofthe ElI
early
promoterhave beenanalyzed
inthecontextoftheviralchromosome. This
analysis
has shed newlight onthetran-scriptional
regulation of this widely used promoter. Byevaluating
the contributions ofeachtranscriptional controlelement in both basal and EIA-mediated activation when
these elementsare
organized
in the milieu ofnucleoprotein
complex
andintheirnaturalenvironment,
wehavebeen ableto
(i) provide genetic
evidence fortherequirement
of thetwoinverted EIIF sites for both basal and virus-induced
tran-scription
and(ii)
show that the threepreviously
identifiedtranscription
factorsthatinteractwith theTAGAmotif,
thetwo EIIF
sites,
and the ATF site are all targets for EIAaction.
Ourresults
clearly
demonstrate that theintegrity
of both EIIFsites is critical forthe promotertofunctionwhether it is in the presenceorabsence of virus-encoded trans activa-tors. Promoters with mutations in either the proximal or distal EIIF site fail to function in basal and virus-inducedtranscription (Fig.
3 and5;Table 1). The EIIFcomplexcanbarely
be detected with mutant promoterscarrying
muta-tionsineither the
proximal
orthedistalEIIFsite,
and such mutants are very inefficient incompeting
for EIIFbinding
with the WT promoter (Fig. 4). The requirement for both sitesin theElI promotercanbe
explained by
thecooperative
interaction ofEIIF with the two EIIF sites. The two EIIF
sites
individually
may bind to EIIF only very weakly,whereas the two sites in an inverted orientation cooperate
dramatically, resulting
inefficient factorbinding.
Therecentdemonstration
by Hardy
and Shenk (22) that EIIF bindscooperatively
tothe EIIF sites of the EIIpromoterin vitrosupports this contention. Our results
complement
thesebiochemical observations and
provide
genetic evidence for therole ofcooperative binding
of EIIFforpromoterfunctionin vivo.
Recently,
it has been shown that the virus-encoded EIVpolypeptide
canfacilitatethecooperative
interaction of EIIFwiththetwoEIIF sites oftheElI earlypromoter(10, 20, 21).
Whether EIA also can promotethe
cooperative
interactionofEIIF with thepromoteris notknownatpresent. InadditiontotheAdSElI earlypromoter,EIIFhasbeen
implicated
intranscription
of AdSEIA (35) and c-myc (23,67)
promoters. Two EIIF sitesare present in eachofthese promoters. It seemslikely
that the two EIIF sites may cooperatefortranscription
in thesecases too. The distance between the two EIIFsites,
their sequence context, and their orientationprobably
allcontributetothisphenomenon.
The
cooperation
of the two EIIF sites intranscriptional
activation of the ElI promoter in basal or virus-induced
transcription
that we observed has some similarities withthatreported for the bovine papillomavirus ElI-dependent enhancer elements: twocopies of the bovine papillomavirus enhancer element cooperate in EII-dependent transcrip-tionalactivation;asingle sitecanfunction only very weakly
(65).
The behavior of the EIIF-binding sites of the EIA pro-moter is considerably different from that of the ElI early promoter. The oligonucleotides that carry either of these
sites cancompete efficientlywiththe WT ElI promoter for EIIF binding, whereas the single EIIF site of the ElI promoteris veryinefficient(Fig. 4A; 64). A single EIIF site from EIA promoter is sufficient to activate transcription
fromaheterologouspromoter(34). The different affinities of thetwoclasses ofEIIFsites for EIIFbindingmaybe dueto
their sequence context. The flanking sequences of a
cis-actingelement can have aremarkable effect withregard to
factorbinding. Forexample,the 17 EIIprotein-bindingsites of the bovine papillomavirusgenome showawide range of
binding affinitiestoElI protein, dependingon the sequence context(43).
The strategy used here to investigate the EII promoter
mutations involved insertion ofmutantpromotersas
dupli-cate copies in the viral genome. There is a possibility that thesemutantpromoters competewiththe residentpromoter
forthe
limiting transcription
factors. Ourresults show thatthe WTCAT construct is
efficiently
transcribed andtrans-activatedintheEIIIregion.We haveanalyzedthe WT and mutant promoters under identical genetic backgrounds.
Therefore,
itisunlikely
that such a competition willinflu-encethe finalnatureof the results obtained and the interpre-tation.
Asecondmajorfinding of this study isthat three
promot-er-specific
motifs participate in EIA enhancement. Thisconclusion is based on the
following
observations. Our results clearly show that promoters with mutations in theTAGAsequence,thetwoEIIFsites,and the ATFsitefailto
respond to virus-encoded trans activators. Previously, in
transfection assays
using
cloned genes, it was shown thatonly the EIIF sites were involved in the EIV-mediated
transactivation(16). Recentbiochemical studies confirmthis observation and show that(i) EIVmodifiesEIIFtogenerate
an
infection-specific
form ofEIIF(20)and(ii)
thismodifica-tion is mediated by direct physical association ofthe EIV
polypeptide
(Huang andHearing,
in press). Given theseobservations,
it is reasonable to conclude that mutantscontainingmutations in the TAGA and ATFmotifof theEII
early promoter failto respond to virus-encoded trans acti-vators because these sites are targets for EIA regulation. Ourresultsdonot
distinguish
between theeffectsofEIAand EIVonEIIF. However,Zajchowskietal. (73)showed that intransfection assays,adeletionmutantof theElI promoter in which only the two EIIF sites were deleted failed torespond to EIA. Inother
studies,
5' deletion mutants withdeletions
extending
into the distal EIIF sites failed tore-spond to EIA,
suggesting
that EIIF at least in part isinvolved in EIA transactivation
(27, 52).
Moreover, in an adenovirus-transformed cell line that containsaconditional lethal mutationaffecting
the EIA largeprotein,
at nonper-missive temperature, EIVexpression
was notaffected butalarge decrease in EIIF
activity
wasobserved,
strongly
suggestingaroleforEIAinEIIFactivation
independent
of EIV (3). Thus, it is almost certain thatimmediately
afterinfection,before the EIV
polypeptide
ismade,
theElIearly
promoteris activated by EIAthrough
thesemultiple
EIA-sensitive elements.
In the promoter mutant in which the TAGA motif is
on November 10, 2019 by guest
http://jvi.asm.org/
mutated, the otherEIA/EIV-responsive elements, EIIFand ATF, are intact. Therefore, the promoter should have re-sponded to EIA stimulation at least on a small scale. Similarly, proximal EIIF site mutants contain TAGA and ATF sites intactyetarevirtually nonresponsivetoEIA.This showsthatotherEIA-responsiveelements failto exerttheir EIA-related phenotype either cumulatively or individually when any one of the EIA-responsive elements is unable to function. One element alonecannotconferEIA responsive-nesstothepromoterin thiscase.These resultsarestrikingly
different from those obtained in stable and transient trans-fection assays. Mutations in these elements reduced basal transcription but did not affect EIA-induced transcription (48, 52, 72). Although we do notknow the specific reasons
for these differences, transfectionassays,whether stableor transient, do not reproduce in the infected cells the regula-tory pattern that is in effect in the viral chromosome. In stable transfection assays, a mutant with only the TAGA
sequence was sufficient forbasal and EIA-stimulated tran-scription (32), and LS mutations that affected basal tran-scription didnotaffectEIA-mediatedtransactivation. When measured after transfection, mutations in the TAGA motif, EIIF sites,orthe ATF sitewere all inducedby EIAtonear normal levels (48, 52, 72). Similarresults werealso obtained for theEIA induction of the AdS EIII promoter(12, 38).
Although a number of laboratories have attempted to define the mechanismbywhichEIAstimulatestranscription of RNApolymeraseIIorIIIpromoters,sofarnoconsensus
has developed. It is, however, clear that not all transcrip-tional control elementsare sensitivetoEIA regulation (57). In virusexperiments,the TATA motif has beenimplicatedin EIA transactivation of the AdS EIB promoter (56, 57, 68). TATA box was also implicated in the case of the rabbit
P-globin
(18) and human HSP70 (62) promoters in EIA stimulation. For theEIVpromoter, theCREB/ATFfactor is believedto be the target for EIA (13, 14, 36). Our studies suggestthat all threetranscriptionfactorsregulatingtheEII earlypromoteraretargetsforEIA. It isnotclearatpresent whether the recently postulated EIA-sensitive TFIID that interacts withEIA-sensitiveTATA elements(39, 40) isalso responsible for transcriptional activation of promoters con-taining analogous sequences.Whatever factor interacts with the TAGAsequence,ourresults indicatethat it isatargetfor EIAaction.What kind of mechanismcanbeenvisagedfor the activa-tion of theEII earlypromoterby EIA?Twohypotheseshave beenputforwardto definethe mechanism ofEIA transacti-vation. One is based on thephosphorylation of EIA-sensi-tivetranscriptionfactors leading toan enhanced bindingof theseproteinstotheirtarget promoters. Recent results show that in virus-infected cells, several transcription factors, includingEIIF, arelikely tobe modified atthe posttransla-tional level (4, 24, 58). It is conceivable that the three transcription factors that are specific to the EIl early pro-moterareallmodifiedby EIAinvirus-infectedcells. Each of thesemodifiedfactors mayenhance the transcriptionalrate by binding directly either to DNA or to the multiprotein transcriptional complexes. The other mechanism postulates that EIA protein functions directly in the vicinity of the promoterandincreases transcription by bindingtothe DNA directly or by binding to the DNA-transcription complex (44). If this were the mechanism, the EIA protein would facilitate the interaction of themultiple EIA-sensitive tran-scription factors with their target sites on the EIl early
promoter. Our results are consistent with either of these
mechanisms.
This is the first instance in which
multiple
transcriptional
control elements andtranscriptionfactors have been impli-cated in asingle
promoter in EIA transactivation.Excep-tionally stringent
control of this promoter isperhaps
to beexpected considering
theimportance
of the geneproducts
that it controls. The 72-kDa
DBP,
the 140-kDa DNA poly-merase,and the80-kDaterminalprotein
areall vital for viralDNA
replication
(66).
The 72-kDaDBP alone hasmultiple
functions in virus
replication (2, 15, 33, 54, 55).
Transacti-vation of the ElI
early
promoterby
the differentEIA-sensitive
transcription
factors combined witha noveladdi-tionalmechanism of transactivation
by
the EIV gene(16, 20,
22, 25a,
60)
guarantees an efficienttranscription
of thispromoter when EIA becomes available in the cell and ensures an
adequate
supply
of theseproteins
in thelytic
cycle
of the virus.ACKNOWLEDGMENTS
WethankNoel Bouck and Richard Scarpullafor criticalreading
ofthemanuscript,ManoharR. Furtado forhelpin virus construc-tionexperiments, and Prithi Rajan forproofreading. The
oligonu-cleotides wereprovided by theNorthwesternUniversity
Biotech-nologyFacility.Thisworkwassupported byPublic HealthServicegrant A120156 from the National Institutes of Health. B.T. was an established
investigatoroftheAmericanHeartAssociation duringthe
perfor-manceof this work.
LITERATURE CITED
1. Ausubel, F. M.,R. Brent, R. E.Kingston, D. D. Moore,J. A. Smith,J.G.Seidman,and K.Struhl. 1987.Current
protocols
in molecular biology. Greene Publishing Associates and John Wiley&Sons,NewYork.2. Babich, A.,andJ.R. Nevins.1981. ThestabilityofearlymRNA is controlled by the viral 72 kD DNA
binding protein.
Cell 26:371-379.3. Babiss, L. E. 1989. The cellulartranscription factor E2F re-quires viral ElA and E4 gene products for increased DNA
bindingactivityand functionstostimulateadenovirusE2A gene
expression. J. Virol. 63:2709-2717.
4. Bagchi,S., P.Raychaudhuri,andJ.R. Nevins. 1989.
Phospho-rylation-dependentactivation oftheadenovirus inducibleE2F
transcriptionfactorinacell-freesystem.Proc.Natl. Acad.Sci. USA86:4352-4356.
5. Berk, A. J. 1986. Adenovirus promotersandEIA transactiva-tion. Annu. Rev.Genet. 20:45-79.
6. Bhat, G.,L. SivaRaman,S. Murthy, P. Domer, andB. Thim-mappaya. 1987. In vivo identification of multiple promoter domains of adenovirus EIIA-late promoter. EMBOJ. 6:2045-2052.
7. Bradford, M. M. 1976. A rapid and sensitive method forthe quantitation of microgram quantities of protein utilizing the principleofprotein-dye binding.Anal. Biochem. 72:248-254. 8. Deutsch,P.J.,J.P.
Hoeffier,
J.L.Jameson,andJ.F.Habener.1988. Cyclic AMP and phorbol ester-stimulated transcription
mediatedby similarDNAelementsthatbinddistinct
proteins.
Proc.Natl. Acad. Sci. USA85:7922-7926.
9. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder. 1983. Accurate transcription initiation by RNA polymerase II in a
soluble extractfrom isolated mammaliannuclei. Nucleic Acids Res. 11:1475-1489.
10. Durr-Jansen,P.,H.Boeuf,andC.Kedinger. 1989.Cooperative bindingoftwoE2Fmoleculesto an
Ela-responsive
promoteristriggered by the adenovirus Ela, but not by a cellular Ela
activity. EMBO J. 8:3365-3370.
11. Engel, D. A., S. Hardy, and T. Shenk. 1988. cAMP acts in synergy withEIAproteintoactivatetranscriptionofthe aden-ovirusearlygenes E4 andElA.Genes Dev. 2:1517-1528. 12. Garcia, J.,F. Wu,and R. Gaynor. 1987. Upstream regulatory
regionsrequiredtostabilizebindingtothe TATA sequence inan
on November 10, 2019 by guest
http://jvi.asm.org/
adenovirusearlypromoter.Nucleic AcidsRes.15:8367-9385. 13. Gilardi, P., and M. Perricaudet. 1984. The E4 transcriptional
unitof Ad2: farupstream sequences arerequired for its trans-activation byEIA.Nucleic Acids. Res. 12:7877-7888. 14. Gilardi, P., and M. Perricaudet. 1986. The E4 promoter of
adenovirus type 2 contains an ElAdependent cis-acting ele-ment. NucleicAcids Res. 14:9035-9049.
15. Ginsberg, H.S.,M.J. Ensinger, R. S. Kaufman, A. J. Mayer, andU. Lundholm.1974. Cell transformation:astudy of regula-tion with types 5 and 12 adenovirus temperature-sensitive mutants. ColdSpring Harbor Symp. Quant. Biol. 39:419-426. 16. Goding, C., P.Jalinot, D.Zajchowski, H. Boeuf, andC.
Ked-inger. 1985. Sequence-specific trans-activation of the adenovi-rus Ella early promoter by the viral EIV transcription unit. EMBOJ.6:1523-1528.
17. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinantgenomes whichexpress chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051. 18. Green,M. R.,R.Treisman,and T. Maniatis. 1983.
Transcrip-tional activation of cloned human beta-globin genes by viral immediateearlygeneproducts. Cell 35:137-148.
19. Hai, T.,F. Liu,E. A. Allergretto,M.Karin,and M. R. Green. 1988. Afamilyofimmunologicallyrelatedtranscription factors that includes multiple forms of ATF and AP-1. Genes Dev. 2:1216-1226.
20. Hardy,S.,D.A.Engel,and T.Shenk. 1989.Anadenovirusearly region4 geneproduct is required for induction of the infection-specificform of cellularE2Factivity. GenesDev. 3:1062-1074. 21. Hardy, S., and T. Shenk. 1988. Adenoviral control regions activated byEIAandthe cAMPresponseelementbind thesame factor. Proc. Natl. Acad. Sci. USA85:4171-4175.
22. Hardy, S.,and T.Shenk. 1989. E2Ffrom adenovirus infected cells binds cooperatively to DNA containing two properly oriented andspaced recognition sites. Mol. Cell. Biol. 9:4495-4506.
23. Hiebert, S. W.,M.Lipp,andJ.R.Nevins.1989.ElA dependent trans-activation of the human MYCpromoteris mediatedbythe E2Ffactor. Proc. Natl. Acad. Sci. USA 86:3594-3598. 24. Hoeffler,W.K.,R.Kovelman,and R.G. Roeder.1988.
Activa-tion oftranscription factor IIIC by the adenovirus EIAprotein. Cell 53:907-920.
25. Hoeffler,W.K.,and R.G. Roeder. 1985. EnhancmentofRNA polymeraseIIItranscriptionbythe EIA geneproduct of aden-ovirus. Cell 41:955-963.
25a.Huang,M.,and P.Hearing. 1989.The adenovirusearly region4 open reading frame 6/7 protein regulates the DNA binding activity of the cellular transcription factor, E2F, through a directcomplex. Genes Dev.3:1399-1710.
26. Hurst,H.C.,and N.C.Jones.1987. Theidentification of factors that interact with the EIA-inducible adenovirus E3 promoter. Genes Dev. 1:1132-1146.
27. Imperiale, M. J., R. P. Hart, and J. R. Nevins. 1985. An enhancer element in the adenovirus E2 promoter contains sequences essential forthe uninduced and ElA-induced
tran-scription. Proc. Natl. Acad. Sci. USA82:381-385.
28. Jalinot, P.,B. Devaux, and C. Kedinger. 1987. The abundance and in vitrobinding ofthreecellularproteins interacting with the adenovirus ElIa early promoter are not modified by the Ela geneproducts. Mol. Cell. Biol. 7:3806-3817.
29. Jalinot, P.,M.Wintzerith, M.Gaire,C.Hauss, J.M.Egly, and C.Kedinger. 1988. Purification andfunctional characterization of a cellular transcription factor that binds to an enhancer elementwithin the adenovirusearly Ellapromoter. Proc.Natl. Acad.Sci. USA85:2484-2488.
30. Jones, N.C.,P.W.J. Rigby,and E. B. Ziff.1988.Trans-acting protein factors and theregulation of eukaryotic transcription: lessons from studies on DNA tumor viruses. Genes Dev. 2:267-281.
31. Jones, N. C., and T. Shenk. 1979. An adenovirus type 5 early gene function regulates expression ofotherearly viral genes. Proc. Natl. Acad. Sci. USA76:3665-3669.
32. Kingston, R. E., R. J. Kaufman, and P. A. Sharp. 1984. Regulation oftranscription of the adenovirusElI promoterby
Elageneproducts: absence ofsequencespecificity.Mol. Cell. Biol. 4:1970-1977.
33. Klessig, D.F., and T. Grodzicker. 1979. Mutations thatallow human Ad2 andAdStoexpresslategenesinmonkeycellsmap in theviralgeneencodingthe72K DNAbindingprotein. Cell 17:957-966.
34. Kovesdi,I., R.Reichel,andJ.R.Nevins.1986.Identification of acellulartranscriptionfactor involved inEIAtrans-activation. Cell 45:219-228.
35. Kovesdi, I.,R.Reichel,andJ.R.Nevins. 1987. Role of adeno-virus E2 promoterbindingfactor in EIA-mediated coordinate genecontrol.Proc. Natl.Acad.Sci. USA 84:2180-2184. 36. Lee,K. A.W.,and M. R.Green.1987. Acellulartranscription
factorE4F1interacts with Ela-inducible enhancerandmediates constitutive enhancer function in vitro. EMBO J. 6:1345-1353. 37. Lee,K. A.W.,T.-Y.Hai,L.SivaRaman,B.Thimmappaya,H. C.Hurst,N.C.Jones,and M. R.Green.1987. A cellularprotein, activating transcription factor, activates transcription of multi-ple EIA-inducible adenovirus early promoters. Proc. Natl. Acad. Sci. USA 84:8355-8359.
38. Leff, T., J. Corden, R. Elkaim, and P. Sassone-Corsi. 1985. Transcriptional analysis of the adenovirus-5 EIII promoter: absence ofsequence specificity for stimulation by Ela gene products. Nucleic AcidsRes. 13:1209-1221.
39. Leong, K.,and A. J. Berk. 1986. Adenovirus early region 1A proteinincreasesthe numberoftemplatemolecules transcribed incell-freeextracts.Proc. Natl. Acad. Sci. USA 83:5844-5848. 40. Leong, K.,L.Brunet,and A.J.Berk.1988. Factorsresponsible forthehigher transcriptional activityofextractsof adenovirus-infected cells fractionate with the TATA-boxtranscription fac-tor.Mol.Cell. Biol.8:1765-1774.
41. Leza, M. A., and P. Hearing. 1988. A cellular transcription factor bindstoadenovirusearly regionpromoters andto acyclic AMP responseelement.J. Virol.62:3003-3013.
42. Leza,M.A.,and P.Hearing. 1989.Independent cyclicAMPand EIAinduction of adenovirusearlyregion4expression.J. Virol. 63:3057-3064.
43. Li, R., J. Knight,G.Bream,A.Stenlund,and M. Botchan.1989. Specific recognition nucleotidesandtheirDNAcontext deter-mine theaffinityofE2proteinfor17bindingsites in the BPV-1 genome.Genes Dev.3:510-526.
44. Lillie, J. W.,and M. R.Green. 1989.Transcriptionactivationby theadenovirusElaprotein. Nature(London)338:39-44. 45. Lin, Y.-S., and M. R. Green. 1988. Interaction ofa common
cellulartranscriptionfactor, ATF, withregulatoryelementsin both Ela- and cyclic AMP-inducible promoters. Proc. Natl. Acad. Sci. USA 85:3396-3400.
46. Loeken, M., I. Bikel, D. M. Livingston, and J. Brady. 1988. Trans activation ofRNA polymerase II and III promotersby SV40 smalltantigen.Cell55:1171-1177.
47. Loeken, M.,G.Khoury,andJ. Brady.1986. Stimulation of the adenovirus E2 promoter by simian virus T antigen or EIA occursbydifferentmechanisms. Mol. Cell.Biol. 6:2020-2026. 48. Loeken,M. R.,andJ. Brady. 1989. The adenovirusenhancer. Analysisofregulatorysequences andchangesinbinding activ-ity ofATFand EIIF following adenovirus infection. J. Biol. Chem.264:6572-6579.
49. Mathis,D.J.,R.Elkaim, C.Kedinger,P.Sassone-Corsi,and P. Chambon.1981.Specific bindingoftheadenovirustype-2early and late E2 transcription units. Proc. Natl. Acad. Sci. USA 6:2684-2694.
50. Montel,C.,G.Courtois,C.Eng,and A.J.Berk.1984.Complete transformationbyadenovirus 2requiresboth EIAproteins.Cell 36:951-961.
51. Montminy, M. R., and L. M. Bilezikjian. 1987. Binding ofa nuclear protein to the cyclic AMP response element of the somatostatingene. Nature(London)328:175-178.
52. Murthy, S. C. S., G. P. Bhat, and B. Thimmappaya. 1985. Adenovirus EIIA early promoter: transcriptional control ele-ments and induction by the viral pre-early EIA gene, which appears to be sequence independent. Proc. Natl. Acad. Sci. USA82:2230-2234.
53. Nevins, J.R.1987.Regulationofearlyadenovirusgene
on November 10, 2019 by guest
http://jvi.asm.org/
sion.Microbiol. Rev. 51:419-430.
54. Nevins, J. R., and J. J. Jensen-Winkler. 1980. Regulation of earlyadenovirus transcription: a protein product of early region 2specificallyrepressesregion4transcription.Proc.Natl. Acad. Sci. USA77:1893-1897.
55. Nicolas, J. C., P. Sarnow, M. Girard, and A. J. Levine. 1983. Host range temperature conditionalmutants in the adenovirus DNAbinding protein are defective in the assembly of infectious virus. Virology 126:228-239.
56. Parks, C. L., S. Banerjee, and D. J. Spector. 1988.Organization ofthe transcriptionalcontrol region oftheEIb gene of adeno-virus type 5. J. Virol. 62:54-67.
57. Pei, R., and A. J. Berk. 1989. Multiple transcription factor binding sites mediate adenovirusEIAtransactivation. J.Virol. 63:3499-3506.
58. Raychaudhuri, P., 5. Bagchi, and J. R. Nevins. 1989. DNA binding activity of the adenovirus-induced E4F transcription factor is regulatedbyphosphorylation. GenesDev. 3:620-627. 59. Reichel, R., I. Kovesdi, and J. R. Nevins. 1988. Activation ofa
pre-existing cellular factor as a basis for adenovirus E1A-mediated transcription control. Proc. Natl. Acad. Sci. USA 85:387-390.
60. Reichel, R., S. D. Neill, I.Kovesdi, M. C. Simon, P. Raychaud-huri, and J. R. Nevins. 1989. The adenovirus E4 gene, in additiontothe ElA gene, is importantfor trans-activationof E2 transcription andfor E2F activation. J. Virol. 63:3646-3650. 61. Sassone-Corsi, P. 1988. Cyclic-AMP inductionofearly
adenovi-rus promoters involves sequences required for EIA trans-activation. Proc. Natl. Acad. Sci.USA85:7192-7196. 62. Simon, M. C., T. M. Fisch, B. J. Benecke, J. R. Nevins, and N.
Heintz. 1988. Definition ofmultiple, functionally distinctTATA elements,oneofwhich isatargetinthehsp70promoterforEIA regulation. Cell 52:723-729.
63. SivaRaman, L., S. Subramanian, and B.Thimmappaya. 1987. Identification ofafactorinHeLacells specificfor an upstream transcriptionalcontrol sequenceofanEIA-inducible adenovirus promoterand itsrelative abundanceininfected and uninfected
cells.Proc.Natl. Acad. Sci. USA83:5914-5918.
64. SivaRaman L., and B. Thimmappaya. 1987. Two promoter specific host factors interact with adjacent sequences in an EIA-inducible adenovirus promoter. Proc. Natl. Acad. Sci. USA 84:6112-6116.
65. Spalholz, B. A., J. C. Byrne, and P. M. Howley. 1988. Evidence forcooperativity between E2bindingsites in E2 trans-regula-tion ofbovinepapillomavirustype 1. J.Virol. 62:3143-3150. 66. Stillman B. W., J. B. Lewis, L. T. Chow, M. B. Mathews, and
J. E. Smart. 1981.Identification of thegene and mRNA for the adenovirus terminalproteinprecursor.Cell 23:497-508. 67. Thalmeier, K., H. Synovzik, R.Mertz., E.-L. Winnacker, and
M.Lipp.1989.NuclearfactorE2Fmediates basictranscription andtrans-activationbyEIA of the human myc promoter. Genes Dev. 3:527-536.
68. Wu, L., D. S. E. Rosser, M. C. Schmidt, and A. J. Berk.1987. A TATAboximplicatedin EIAtranscriptional activation of a simple adenovirus2promoter. Nature(London)326:512-515. 69. Yee, A. S.,P. Raychaudhuri,L.Jakoi,andJ.R.Nevins. 1989.
The adenovirus-inducible factor E2F stimulates transcription afterspecificDNAbinding.Mol. Cell. Biol. 9:578-585. 70. Yee, A. S., R. Reichel, I. Kovesdi, and J. R. Nevins. 1987.
Promoterinteraction of the ElA-inducibleE2Fanditspotential rolein the formationofamulti-component complex.EMBO J. 6:2061-2068.
71. Yoshinaga, S., N. Dean, M. Han, and A. J. Berk. 1986. Adeno-virus stimulation of transcription by RNA polymerase III: evidencefor anElA-dependentincrease intranscriptionfactor IIIC concentration. EMBOJ. 5:343-354.
72. Zajchowski, D. A., H. Boeuf, and C. Kedinger. 1985. The adenovirus-2early Ellatranscription unitpossessestwo over-lapping promoters with different sequence requirements for ETa-dependent stimulation. EMBOJ. 4:1293-1300.
73. Zajchowski, D. A., H. Boeuf, and C. Kedinger. 1987. Ela inducibility of the adenoviralearlyE2apromoterisdetermined by specific combinations ofsequenceelements. Gene
58:243-256.