JOURNALOFVIROLOGY,Apr. 1988, p. 1180-1185 0022-538X/88/041180-06$02.00/0
Copyright © 1988,AmericanSociety for Microbiology
Nucleotide Sequence of
the
Large Double-Stranded
RNA
Segment of
Bacteriophage +6:
Genes
Specifying
the
Viral
Replicase
and
Transcriptase
LEONARD MINDICH,1* IRIS NEMHAUSER,"2PAULGOTTLIEB,' MARTINROMANTSCHUK,3 JACOB CARTON,' STEPHEN FRUCHT,1JEFFREY STRASSMAN,1 DENNIS H. BAMFORD,3 ANDNISSE KALKKINEN4
Department ofMicrobiology, the Public Health ResearchInstitute ofthe City of New York, Inc.,' and Department of Microbiology, New York University School of Medicine,2 New York, New York 10016, and Department of Genetics3 and
Recombinant DNA Laboratory,3 University of Helsinki, SF-OO100 Helsinki, Finland
Received 19 October1987/Accepted22November1987
Thegenomeof thelipid-containing bacteriophage
+6
containsthreesegmentsofdouble-stranded RNA. Wedetermined the nucleotidesequenceof cDNAderived from thelargest RNA segment (L). Thissegment specifies theprocapsid proteinsnecessaryfortranscription andreplication of the
+6
genome. Thecodingsequencesof thefour proteinsonthissegmentwereidentifiedonthe basisof size and the correlation ofpredictedN-terminalamino acid sequences with those found through analysis of isolated proteins. This report completes the sfquenceanalysis of
+6.
This constitutes the firstcompletesequenceofadouble-stranded RNAgenomevirus. Bacteriophage +6 infects the plant pathogenPseudomo-nasphaseolicola HB1OY. Three separate pieces of
double-stranded RNA composeitsgenome,which is located inside apolyhedral nucleocapsid (26). The nucleocapsid has RNA
polymerase activity (20) and is covered byalipid-containing
membrane (26). The assembly pathway involvesthe forma-tion ofa polyhedral procapsid composed of four proteins,
P1,P2, P4, and P7, which is then filled withonecopyeach of thethreepieces of double-stranded RNApervirion (4, 16). These filled procapsids are then covered with a shell of
protein P8tobecomenucleocapsids, whicharesubsequently
enveloped within the lipid-containing membrane (15). This envelopment process is dependent upon the activity ofthe nonstructural protein P12 (17). The fourproteins that
con-stitutetheprocapsidarecoded for by the largest of the three
genomic segments,andtheirsynthesis begins early in infec-tion (25). Theprocapsid is responsible for genomic replica-tionandtranscription (24).
The study of the phage assembly processhas been facili-tatedby the cloning of cDNA of each genomicsegment(18). We recently determined the nucleotide sequence of the cDNA derived from the small RNA segment (12) and the middlesegment(P. Gottliebetal.,unpublished data). In this communication, we describe the nucleotide sequence
anal-ysis of cDNA derived from the large genomic segment designated L.
MATERIALSANDMETHODS
Bacterial strains, phage, and plasmids. Escherichia coli JM105 [A(lac pro) thi strA endA sbcB15 hsdR4 F' traD36 proABlacIq ZAM15]wasusedas ahost forM13cloning.E.
coliHB101 (hsd20,anrK- mK- strain) and MM294 (an rK-mK+ strain) were used as hosts for clones with plasmid
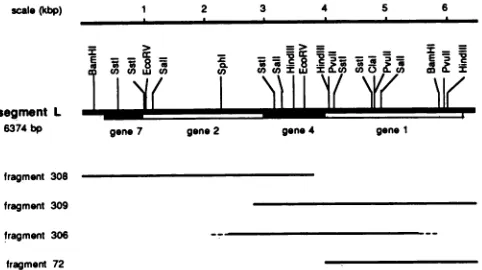
pBR322orpUC8 as avector. Phage M13 (mplO and mpll) replicative-form DNA (14) was used to clone cDNA frag-ments for the sequencing reaction. Plasmids pLMF72, pLMF306, pLMF308, and pLMF309arepBR322derivatives containing cDNA insertfragments ofsegment L(Fig. 1).
*Corresponding author.
Preparation of DNA. Purified single-stranded M13 phage DNA for thesequencing reaction templatewaspurifiedfrom
1.5-ml cultures by the procedure in the Amersham Corp. M13cloning andsequencing handbook. Largeplasmid
prep-arations(200-to500-ml overnight cultures)wereprepared by
the cleared-lysate method of Clewell (2), with subsequent CsClgradientcentrifugation. Restriction digests used endo-nucleasessupplied by Boehringer Mannheim Biochemicals, andthebuffer conditions werethose specified by this
man-ufacturer. Ligation reactions usedT4 DNA ligase supplied by Collaborative Research, Inc. Conditions were as
de-scribed in the M13 cloning and sequencing handbook. Procedures for 0.8% agarose and 5% polyacrylamide gel
electrophoresis have been previously described (13). Trans-formation of E. coli JM105 was as described in the M13
sequencing and cloning handbook. Transformation ofE.coli HB101 and MM294 has beenpreviously described (18).
Proteinpurification and amino acid sequencing. Unlabeled and[3H]leucine- and[3H]alanine-labeled 46h1sweregrown
and purified as described previously (1). Preparative and
analytical protein gel electrophoresis was performed as
previously described (1). Individual proteins electrophoreti-cally eluted from gel slices were free of contaminating materialasanalyzed by subsequent sodium dodecyl
sulfate-polyacrylamidegel electrophoresis.
Twosequencingstrategieswereusedtolocalize the amino terminus of the individual proteins upon the nucleotide sequence. First, unlabeled proteins (to which
14C
label hadbeen addedto facilitate protein localization within the
pre-parative sodium dodecyl sulfate-polyacrylamide gel) were
subjected to automated Edman degradation in a Beckman
890 Dsequencer and a0.1 MQuadril program. Theamino
acids were identified as their phenylthiohydantoin
deriva-tives by high-pressure liquid chromatography (Varian 5020 liquid chromatograph; UV-5 detector,269nm).
In the second method for radiosequence analysis, the eluted tritium-labeled proteins were degraded as described
above, together with 200 ,ug of apomyoglobin. Ninety
per-cent of each phenylthiohydantoin-derivatized sample was
counted by liquid scintillation, and the remaining 10% was
analyzed by high-pressure liquid chromatography, with the
1180
Vol. 62, No. 4
on November 10, 2019 by guest
http://jvi.asm.org/
GENOMIC SEGMENT L OF +6 1181
scale(kbp)
segment L
6374 bp
1 2 3 4 5 6
Ene 7 ; gene 2; 1 gene 4ZE gen X
gone 7 gone2 gne4 gone1
fragment308
fragment309
fragment 306
fragment72
FIG. 1. Restriction map of the entire large genomic segment. Sites were obtained from restriction nucleasedigestion of the cloned cDNA fragments indicated. The illustration is aligned with the direction of transcription to the right. cDNA fragments used for determination of the nucleotide sequence are shown below the map. Kbp, Kilobase pairs; bp, base pairs.
knownsequence ofcarrierapomyoglobin used as a control for sequencerperformance.
DNA sequencing. Nucleotide sequence analysis was per-formed by the dideoxy chain termination methodof Sanger etal. (23). The sequence was determined for both strands. Fragments ofcDNAfor sequencingwerefirst cloned into the unique restriction site region of the replicative form of bacteriophageM13 DNA. Forthiswork, M13 vectors mplO and mpll were used (14). Our strategy involved the use of specific restriction fragmentsaswell as the use of unselected collections of restriction fragments generated by treatment with TaqI. Sequences were determined on over 550 frag-ments.Single-strandedDNAisolatedfrom mature M13 virus servedas atemplatefor dideoxy sequencing. The polymer-ase reaction was primed with a 17-nucleotide primer pur-chased from P-L Biochemicals, Inc., and chain extension waswiththeKlenow fragment of DNA polymerase I (Boehr-inger Mannheim Biochemicals). Labeling was performed with
[a-35S]dATP,
>600Ci/mmol (Amersham Corp.).Dideoxy sequencing using dITP and reverse transcriptase. Some sections ofthe sequencedisplayed GC compressions. We resolved them by substituting dITP for dGTP in the reaction mixture (22). Chain extension reactionsusedavian myeloblastosis virus reverse transcriptase (Molecular Ge-netics Resources). In these reactions, the molar ratio of deoxy and dideoxy nucleotideswasaltered fromthat ofthe Klenowfragment-catalyzed reaction. Final reaction concen-trations were0.77 ,uM dATP to0.027 ,uM ddATP, 889 ,uM dCTP to819 ddCTP, 889 ,uM dTTPto 8.9 ,uM ddTTP, and 220 ,uM dITP to 0.35 ,uM ddGTP. The reaction buffer contained 33 mM Tris hydrochloride (pH 8.3), 6.7 mM
MgCl2,
46.7 mM KCI, and 20 mM mercaptoethanol. The17-merprimerwaspresent at 0.3 ng perreaction. Thechain termination reactions were
catalyzed by
2.5 U of avian myeloblastosis virus reverse transcriptase at42°C
for 20 min. These weresubsequentlychased withamixture of four deoxynucleotide triphosphatesat222 ,uMeach for an addi-tional 20min. Eachchain termination reaction contained2.5,uCi
of[a-355]dATP.
35S-labeled
DNAfragments
were dena-tured and analyzedby gel
electrophoresis
on 6 or 8% polyacrylamidegels
(42.5 cmlong)
containing
8Murea, 89 mM Tris-borate (pH 8.3),and 2 mM EDTA.Computer analysis. The computer
facility
of the Public Health Research InstituteoftheCity
ofNewYorkwasused.It consists ofthe Vax 11/750 computer equipped with the multiuser UNIX operating system. Nucleic acid analysis programs were provided by the sequence analysis package of the Biomathematics Computation Laboratory, Depart-ment of Biochemistry and Biophysics,University of Califor-nia at San Francisco.
RESULTS
cDNA cloning of the 416 genome was described by Min-dich et al. (19). Plasmids were constructed that bore frag-ments of the L segment, and the protein reading frame positionswere estimated by restriction site analysis in con-junction with in vivo complementation studies, in vitro coupled transcription translation of plasmid DNA, and ref-erence to the cloning work of Revel et al. (21). Ultimately, the readingframe assignments were made by the identifica-tion ofthe N-terminal amino acid sequence of the isolated proteinsand comparing these with thesequences predicted fromthe nucleotide sequence of the cDNA.
The large genomic segment of416 contains 6,374 nucleo-tides andit has55.5%guanine-cytosine. Four reading frames werefound that correspond to the four proteins known to be coded forby this segment (Fig. 1 and 2). As with the other two genomic segments, there were appreciable noncoding sequences at the termini. Thetermini of the segment agree with the sequence found by Iba et al. (9) by direct sequenc-ing of the genomic RNA. The order of the genes is in agreement with thatfoundby Revel et al. (21) on the basis of theprotein synthesis competence of fragments on plasmids containingT7promoters. Thesalientresultsof the sequence analysisare asfollows.
Gene 7 is the first significant open reading frame in the segment.The readingframe starts with an AUG at position 458 and terminatesat position 940 with a UGA. Thereis a Shine-Dalgarno sequence preceding the initiation codon (Fig. 2). The calculated molecular mass ofP7 is 17.3 kilo-daltons. The amino-terminal methionine is partially absent on the basis of amino acid sequencing. The sequence pre-dictsanabsence ofcysteine, andthiswas confirmedby the lack of labeling ofP7 in vivo when
+16-infected
cells were exposed to radioactive cysteine (results not shown). P7 is predicted to have a high net negative charge, and this is consistent with the behavior of the protein in isoelectric focusing gels (17).Gene 2 follows gene 7. The initiating codon begins at position943 with anAUG that overlaps the UGA termina-tion codon of gene 7. Gene 2 does not have a discernable Shine-Dalgarno sequence in the
vicinity
of the initiating codon. The reading frameends atposition 2937. The termi-nationcodonis UAA.ThecalculatedmolecularmassofP2 is 74.8 kilodaltons, and the N-terminal methionine is lost. The predicted amino acid composition indicates a basic protein, consistent with itsbehavior in isoelectricfocusing gels (17).Gene4followsgene 2. It hasaShine-Dalgarnosequence, and the
initiating
codon is an AUGbeginning
atposition
2943. The reading frame terminates at position 3938. The termination codon is UAA. The calculated molecular mass ofP4 is 35.0
kilodaltons,
and the N-terminal methionine is lost.Gene 1follows gene 4.Ithasa
Shine-Dalgarno
sequence, the initiating codon is an AUGbeginning
atposition 3951,
and thereadingframe terminatesat
position
6257 with UAA. Itisnotablethatgene1endsonly
117 basepairs
from the end ofthe segment.This is much closer thanthosefor the small VOL.62, 1988on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.62.303.76.211.2]II I~~~~~~~~~~~~~~~~~~~~36 07~~~7WITITMCC?71IMC?SSI71CCCT0TSCMlMY0101MyGMCCC?~~71OCee0001TeCC~CC00C?1WeCCTMGSITACTOCO eeCC1Warc 121
II
SOTC OMIITM Y0 OTWISSCOTO AOMG?SIOClCanMSOSAMITATICOT ?IClCGOTTooCACRIGA 7MCCTTTCA O T TcCTCGAO l 252 I
CG?CCC UTS ST? 1W1 SCIIIAPSMOCTI CC?SOCaCY SC?ICCGoo OCT 51a 11ICCCPA CCMAAoCeTSC G WCOCT? CMSGCoeCv 145
3WpIIWlee1m got kmLyspNoLyem1LyesGlyMLee L1ou Lyek A"jSlyValGlc alee.iLys.Ole OlySet3.lee51. @13Lee II 705CC?AC M ?AC10 Se OTCCCGTa IGT CCSOCOiCMTCCCMCe Sam amC CAT 5CACCoC CCCGCCCM IUCCRICooIC?CCOTCS 11 Baer PEG SmVll VteUpecSyl M1e L1. Vol lIe Lou G1leeI.tAe TIsAleeNileeL. .II*w Vlee Olee ProSeSGtProTIeLAmVle
TeeIC M 710 7CCMCGAIU M CCTI AM MA OMC OM C?S GUAM STAICG0CCG CMOSCT GMA MACCICCTSOCSC? GMCAmCIC 510
epGI.GIslAUS.lerLyeenetleeLyeSluSlyleeLeeII,Clee IpTyrGl Vly 1.SLyuSetVLy AzrPSeAISly II.51otlpee ACGAACGAGC07 ?TM ICOO51TAT CIO ICCCS?CC SOOTO AAC 1WT0CGCI SC?ICCGCCC OnSMAT GMAACOMCOn ICSIT 6CC 3211
Lse A3W Cee Vl* gty Ah spTyr LoueT.espmisLbee SlyIlaAsaTol k. Gl ITe lyAnValTy1Gl AsaOlyValACVI. IapVm
SI?TTGM A11TMCSC 0CAC CCACMSO 1TTSCCCRAGATCCCCCCMCMI?ACCT CCCIn MSC?kCMICC CT TCOAA GCCMIMCOCCGM 12014
1WCM CM 5CC7WCCT AM7CS?MCO 7CGCACACCGM501IT?fGMC1WC TAOTC CooCrCTCT?I ?C@ MCTCC?Ca771 Or?10 MCC 1104
II. AlaAs1, II.S.c LyeE, Icy ee0 AssLyp Sly 11. Il.Lem AleaAlaVale Sly Lee Le feseVtAsp WuIlu3ro Ieee Sec IOU
IM .I
AMQ MC C?CTC?G0UTITMCCCICCMT TAACM7CC1WC A C CCTATOWAlIT AAGACSWASITI 0MCSOTG&SIcT CS? GCCMCA 1144
LyeleeAleLysSelIySyt Lee 7hr IspSieproTyr Me.leorA V1 HopSIyTy SLysIcy, VGluy SlyAleeIsrSly VeloI luhlee
OCCITT GU ICCCM TCM110MCeeITTCCION C ICC CCC 1WG1COMA CC C?COCTICCMC CITOMSOM CS?SCCOTCOCTTS In2d 3D,GIeSly1 Se IYSlesa Notle SoPe3lly Lse lIeAW ASPAzlea3WrGLeeZANle z Mee lee lye ALyeITyrTyrIsp Vl Typ
II
C?IOCWITSOCACC?GLWIC GMCM SICMTC C?CTC?C7W OMAWTIC OM1C5 7Wa11 ?TAVOCATC MTGCCCGA ACC C?M 12744
A"e MA.CIASly 3WcVe" Sesa Sle Ic sp Lee."PrLee 31.IcyLyeII. Val IcrLee.A"t 3A"nez VapLeAleAarVpeGI TyWLe AMICACM MI001G CCRSaCCI CCC ICCCM WI TMCCMMCST ICC 001C 1T AMIhCCS ICC1WOMCMA Sec? CC? GMcMI 1744 VlyehIl.yGlyOeS eec ec o7hly Cye11W lycI, LyseecAspAl Set SlyLazLyelIAI T1.Ily1 lieAspVaI proLeeSleye 1WSMCSMQC?SOCO MIT CCC ICCCCCMoSananCC 51COM1WoACCMCCCACMCM ICCSg OM?1WA7?gM GCM ACC&I 1114
1no51 OleGigeSly lee L TSe Lee 51.Sly LyeproIsp Isp Louayc@1., ISo Se01 SetSeVlySly GIslycyValU VsmLys
CMT CC CM ICTMC517 AC? ICC1WChCCS CC?Amh ICCe #0?71M 50aIC AM511 CMIWASICSCTMICCMTgM15CC 1644
Iy11.TO .e 5.,TYzIsp II. Il.PbzEleeOtep3W Lye7hr Sly Lye G1.V.1ee LyeIspIcysSet V.1 set IspCyeSale UysI
II
071 Am SO STT GMaCMSCOCICChCCCG7 SC?OC?g TAMO 111WGC CC?WICC0 AMCAaMQCM CAM50011TN7 aCCCC SIC SOS 174
ValThzSLyuSlySleSI SlySocLeeL~ ProI.I SezLyea IspAlea.IcGAlaPr OlyeGISly61. TyzSl LIe IspVolyA"eeIs Sly
AMMICATCTC GUS GCM0CM CSC? ICC OMSOCPA ON711 710MCC SC? CC? ICC ICC 1WCTCa1 MA Cr76WCAMMa T 10)4
ASO PeeCye LOUIcyIAL IcY hzU lIeSetSly S1ly3 3U.II.AISO lee11lyII. Se lieVeOUNt 11.01 3zTyrIcy Vle Lye
1C MITGTCACAM MSrC?CI MCC CMC?I?CMCCMAMA AMA@CAM Q AOCaAMOMT611 = CGMISOIC 71TTMCMSC?IC? 1914
Il. ?yc eec Lye Iyz AlapyoalzLeoale seg hz 7hz LysAsplee LyeSoeAle Lye AVlLyeSLeu lop SozIx CyeV.111,*7hzgi SM 071AICCSOC CMaAiSIC 1W ICC 100CC?50700 C?S IT 7W ICC 101AAA a AlCCCCCC MC ICCSOTa CAT MM e IS 00 2004
Isp01Vol SecAXI AtI 10SO7hza31.sp Ysp?LpssWlo lely cylpLep rILye lypAS leLeeoIuVle SlySly Niyck113 ?oPb tlp
MAMOCIO A CCGMG ICACICAC7 AM?CACC? ?A ATOAM00SOCSC?CC?SAC?CCAGMCWGC SOTC CM T an7WWACMSI?CCC7,CC 204
Ho1Lye LeeGluSly.7zeeLysa es peleeg rTyW notLysVol Slylet3Wye3WuNi Gl 1lylly 7hzleel eeSlyIspIl 3Wuo Syc M&C ICQC?T COTGWAO 7C SOTCC ICCCAC T01CMOST1WTIACCScACCAICSCGA1W ACCCCICCM ICMCMCC C 010 21044
lSeeArWI Le1eg le olSlyleer SecSleSly ValSlyAsa7hz lop lee Set Sly 7A" Lee LeeSet,seclIeGiz Tyclee Val Sle
CMSCT 011CMCICC SC? C?CM7a MCMMACGCCWeICA T SCCaCTOCCTCIC 5CCOMVWCAlC MCT?A=ICC1MT ISOVCM00 ACM 2274
61*ale IspAre Ahz11. PctLSe LeeVl eec IcyCV ProLye Isp SetPee. ec Ice lyeG Icys3AaeIspTee alycAlaop01.Slyie SMCAMICCCM C ICC7CC?AM OC?SI?STM C?1WIenSC 75OMMOm 01T CMSLkC?AM0 7C551CMTTN G CM7WICC GM Sol4
Sle 5 LIe Ic SLeu IerSecLyeSec IspIotIceAlelee SlyAlop7hzLyeSlyGIGLouLeeVrol1Sly SlyeleI aLee31.SI ICCCC ACAMGMSC CAMTAT AN CCAC CC?OMICCAM ICC ICCO PTMGMCACMA50SC W ICCCITSGCOTS ICCCgmCT?Tm 2041
SetLeeAsLye LouSlyLyeVLyleo Tsc 3WllycAset Vly IeA"clyc OleAle SlySalyLo notP is eSlyIspIet Vlee Lelys
S4
SICaCCCM CMSMCC?SOC CC?1WO ICCAGO7 S ATSICCOATMCCCIACC?MMCAOMCCAT MCCaIrMTCA510CCCC 2544 lop eec Icy Icy Sle Pc.SlSecValn PIeAVolpSlyhS le lI lo Sec SetVle leo leoSlePIeAsec UWSealy Sly VolA Ale
G1WSTOCCCACSIC WeIC? aMTC MWewCTCM C CMrCM7A OCT A0C1W M1WMICCAMSITMCTICAa 1WeTCkc? we CC 2043 Sec klyVIcyw IspIzyee Vly IcetLye IcyVaW Wt 3Wly Lee.PrIce Top Ie SecSetVlyeaIs7hzLysTSlTcyerIIug".
?MYNIC?90LMSGI?A07TOCCCG AGMWIC&TACGMaCM"?CCC100~p15CI5GMC1 CWeelAGOMACTIC?CM1WI CM?001SI?ICOC7 AM CC 32724
lIzSecIspVolLee 610 lb II. Ole Icy lye lopToPlee11MAh.spSleSebly lpycIcy Valy Iee le.IspJSe leeLysIro
GMT 1CCCACCOMC?CCI CM ICWMrA 7C?CICC SC?OCM ACMW 000 CCC C? 011 AM C?TM It?01aTMGMC610 7 Am 2014 lop 7hz LeeAlelee Sc Iy0ycVolY11, SYEcSetIsIy1*1.GlyVl ra Louno 11.Ole.lee 7hzSeW lIeIyspletIe T1VoleleeIc SMIspWAMLys MCAMT CC7W CMC*IA?IC?AM75 M S 1W SI? 0C 1W 1W MTICCCMTGM611GAAMO110 A?SC07C 1WSC 011WAMG 2043
los Lye Lee. 61.Vly Lyeo?cp7hz Ole 11*IspVa1Alz 11.AleeTyrA"s IsVlM lee SetAl'eSaly o e o ly
IC?GMCTOMAW7 C0MIC? 011ICC WeMS10 CTCIOCTC AIT?OM 071a MTCM1aW C? CT?011C CMTC COC ICC1WM1 23923
AspGzOle IcytPI leeICyS te,VolySLow$Ic Ala Low Sey Pc.aIeVolAeVol tozsletMIVlelIepr IspIcLolSlylIaemLy
7WICCOGM Om CCCAMT CMSTa OT 1Wan M'CSOT CM 01 171 CM, CC? 1WkW CC CCC AMSM MCM CC7WMC 17
Voly SetAleLee.AlIeS lyllyieV lyAlVac lee Ic lyVlIso o SekI Iy3o i ecVl8S lee LeeLe.Is 7hIce _I.P_o7hz362
1182
on November 10, 2019 by guest
http://jvi.asm.org/
VOL.62, 1988 GENOMIC SEGMENT L OF ct6 1183
weanmWam KW1AMGC tanO SMC6?"a"aa aMGTIMtcranIIC66 CMGOTCATrammaTmGACTamCaM amCI"TSOT 3713
VOLNetV.1 V.1"As Pt. Met ValAapAla61.LysI1.61.TyrValPh.6ly6inValmetAlaseat 1hsVal61yAla It.LoaCya
ml 61?66CAmeOt.maMA ammmMvcmanAMamAAA ?COTA??TICM&CGOTmaGam cc?Caam amSC WVCmCAte M3
Al.Asp 61yAm.Va1setAUV1zEatPMaAre1hzA"Is. aLyeyAUVI. Ph.A."61yAl.AlaProLouAlaAt.Asp1hzlugNot amIMC£1 MITcaC61 AmCAmCA AMamCICcmaTCA?M Icam6 mC McrmC GMcogc1oem mca aem21 noGIL6OAC 3663 Pt. EazEa8tASPAn~IP.1hsSet MtLyeAla,LosAMp3s.hzSatIteAla,gatValAl.ft.OLa. Gla AtyStySatValAsp1h
81:Pt II
hc61? ccC M?m m CCCcCCcmca661 WIamC?WCTMaCia UaMO1A?AMA mC MC CIcAMLa"AM 1?mieaC661 3942 AmA"pA"Ass SWAla,proAtga" lymeame.no.S.zL.. IP1I tm" nLGD "A mA l ma am Coemcan mamCTcm mMcamWCm0GMma"a a cmChoCMaI= cGMaO Twoc1M 6 CCCOM CMmTCma 4673 SatIiAtyStybM1z 61.M&PMm& II. 6ly61.if LyeAs.61n,inseatVa1SlyW&aIn63Ai Pa. M6MAVWh UK
AOleh Ph.UazGIsAS,tMF1hzAl See61 V6616A sp61.oV.1alyLyeGIs Am II. AsTyrUt.V1i,S tb AU.rAsp IaPh.Ph. WC m cAMmaam emG TheMCmamaGm2aT=6A maacONMmcmawe TA?W an acma ma mc am Amcom 611 4363
IleTyp AlV61e LAl. TMy Ala,T 166 StrVlyAspOlm 166AV.. 61.PleLye 61.A" z 61. 61.ElLhstVal Cy. AProh 61.e
I
CImOMmCI aM maC maCCOCTam aMemA maC mAG aMmc? ma am AMCammcC aCTIC OTCAMC 616RTon a maamet 43433
lie GIp GAs LyeMm6 Urh Alay P1aoh6wGIsRSt A..LAsp ale.lIl.e Al. AspAlaibValSlyLye V.1Pt Pos 1hz Ala It
I
emmICam maecmamcam cTMCenTMCmc manaec cAm mAMamcmc A"mcTMC6?CT!ama6?TGmmanC 4623 SlyPh.Il1016 Pt. AspAl.,AlaWyeV.1Tyrasp V.16lyAsp1hzAl..1hzTy: Pt.AmPh.Tyr"1.166ValAspCye0V1Asp ON A mCmamgfOM at maGAM OMaCFOmaMaOTCGL TMa amAateC CAOMAMaTMAMGcAMemOCTC?OM 4613
AlaStAspLmAsp AspNot1661h Al.166 Iaz S. ValAsp atLyeHet166 61. Ala 1hzPh.LyeAl.LyeGlyMAl 16MAl
II
1mOTTmaOAMma CTMTCaT CaamaMGmCCTmC AMCmaAm mca amacmicc amammca T11 mCC MC mlC mC 66 mAC 4103 PotVal 166TkI.eS 61.Etla.AiVL.wAmpAl.Alt Shz1hz Al. Ph.61.LoAspSatAs SeyA.. Ph. AspAlaAmThAsaVoValyIle
I
maTa amcamCcmTemCCmgcMCteamaACmGMaTCChoGAmAOMC aMG CmA amMaCoe ma amAmc am eM am 4103
SpatIlal6h l.La.S"lyuAspla.JspSat Pt.AltThrPtGyeAaaaAspPt.VloU SAtl ArlAleaAIspVal II, AmBaryAtar1
6tmamaOmcCG AC maCmaCmaTmcamCmaC& OMaAM61 6cAaMG mCOG 'emAAA mmOTC MCAmC mcG mAC mcAem 4663 Asp 61.se. Ayset Ams1eIleCUVfl.16 h 1 aleAla lTyt61 Asp Eat VO1LybarI.ZAsp&wSly Al.UV le. AsaPh.lbSot Aspet
emTmama maAmaamamAcm166mcXam e maCGAtaMCA? 616mancmTC AMGmamc am amm=AaGAGMcAm 4573 61.Ila.SlyEat 1hzGle1.P.teIlEaatoHt4Alah31 I W Eat IS.VtI So Pt. Ph. LyeLaAspTPtGU n Ams 61. 1hz 1hz Tst
TTamc aemma: amm maCama"TCMatemTCmanmaam catA=mamca m emMA Ca IaChomcm amneamamcA 1663
AyaIt.6110y63.1zSMmAlaVal.s ESeEtUsl 61.Pal SmOAss V.1Us.1.1 Tyr 61GApIspua 61. Ph. AlaLye61. 31. 1m AMAMcma OMmcamo Ama mCCam ammaIancMT mcma Gwmcemcm emamtmacOCT aTC OMa amC manma 6163
amAssP 1hz Pt.ly V.1LyeLouAlAmYAmPrSm W61. Asp Ph.LAnAsp V.1 61. Pt. Asly tAS Asp SmHoSmls Ala1hzAlaAl
Al.PTyrt 6y~1Ala ValAlaRiAsPrGIVah St Ph.Val. 10alyeA asp1laV.1al lAl1l .VolSmy61. AspSlyr 1 Val?T Am I
amCamcACTmAGem ohAe"eaa amcThema m= mTcWAmc amc mc amAcm mcCO ham ccTMaGmAtemcmaa amTam 5"33
Sml AmlyAl.61.oMMt11hzrV.alSlyPhTyrSmA.1I.1 61. Asp Asp Tyt A.166 Asp AspAsPtoG.IEM V Al.MetIl Al. Alr
a AMc CCa emCCammCC TMt WA MCmGema=mcUAm maOG TMcmaAmmamAammA mMArCh MC TMaCOG616 Ate 6433
366AysPr IleGIyP1. AspuValSSet La.61.AlaAysp Ala Sm Amsp 166AlyeAspSmo Eat Ph.AmtTye Tyt Al.T.Pro EMI
catMT TM Am Gm CMMtam AM mammcGmaeeMOCmA= emCcA mc c?ACcAam ma cmarcmAa TICGUamC 531
SOleyAl.a V.1Al. be.AmaTyPt. 61.AV.1V.1 B.1Sm61.l61.1.SlyV.1y iVTrl Al1 1 GlyaSm LaIys Gla.V1 Tsp Amu V.1Asp hz61yLIa. AspIrt.Pt.AV1 SalyAyt11Am Al.Ea6.1ytyStI.asy C l I 1hz Pt. 61n Pot. Ia. 61. At. T. Ala TyVa GACam Cm AmCma GmaeWA aTe amAC C maCmCGA 1maCG mW AMCmT ma AmC maOMCmf am mTGCA tAw caT 5603 VamhLyePt. 31.61 Pt.IIt61. VatLa 1.Aly LyeVh.1l IleGlAlpLa. AmGtIshleVlRiAla 1* etuls 1. tsps PtAspGS.
I
SoaAlaSatyArt IalPhAl.iy61AspAl.SegyoatV1thAspLogiVaGlIet AmsLyAsg tyt1hzisAt1161Ala.1Le 1 Ph. 61, IalY
II 64
37
FIG.
2.Sequence
of cDN ApAsAp1tLaAlAl.geoAt.At.enLoz E6Thz t.AepuAspeAla thayet6Atfthep t Aspratd. ITteAlanAslat. o rdcso tefu n wormiddle
genomic
segVmen 1tsL.L..1hheSlcalaulyateadatoAlaVutlar,Ly 16.Et 1 h La.sA.infsctithanVas spmAsp 61.ml hirlpocpi
ofPlis85.0 kildalt 1nsThemN-terminmamlmeticmca tinremcatins ma c mc mmca lm m mm c54EaAalyAsSy. 3. spAs St e Ap a.s atSl taAmA16 S A.tAse,pro
apTspilaS
isaAloablohVh.1a srpts n withthebasicityfou
La. 61.iseMlectriy166Lacetusin 1t76.Alp1.AarochzLe. apsidsmrAmtA166re EinthV.eAla o a p n o eah doFIG.2Sequnceo cDNA f genmic sgmentL of 46 Th sequnce i thatof8thAnstand.siThe
transltoionpoductsy
of thefourknrownwithth
basictyfouDbyISoelectIcN
focsig17.uroapidstedcattureintefr
fcasi
compoen
andodecahedrone
Thefourproteinscoded for by the largegenomicsegment cules each of P1 and P4, about 20 molecules of P2, and are
P1,
P2, P4, and P7. Theseproteinsaresynthesizedearly between 80 and 100 molecules ofP7. Although the results ofon November 10, 2019 by guest
http://jvi.asm.org/
1184 MINDICH ET AL.
this report change the predicted molecular weights for the proteins, the changes are not great enough to alter the expected stoichiometry oftheparticle significantly. Recent studies (10, 19) suggest thatP1 itselfcan forma dodecahe-dral structure. Itisnot knownwhether the active site(s)for polymerase activity is shared by different proteins or whether it is located within one ofthe four proteins ofthe procapsid; however, no RNApolymerase activity hasbeen demonstrated in any structure simpler than an RNA-filled procapsid.
Acomparison of the amino acidsequencesofproteins P1, P2, P4, andP7againstthe Dayhoff proteinsequencelibrary as of April 1987 showed no significant similarity to other proteins. However, a search for sequences suggestive of nucleotide-binding sites revealedthat P4containssequences highly similar to those found for a class of proteins that includesthemultiple drug resistance proteins oftumorcells, transportproteins ofbacteria, and UVrepair proteins (Fig. 3) (7,8, 27). Themeaning of this site inP4isnotclear. Since the
procapsid
is involved in RNApolymerization and pack-aging andmustitself be assembled, wecouldspeculate that the P4 site is involved in one of these processes. This sequence is, however, not characteristic of polymerases. Recentunpublished work inourlaboratory indicatesthat P4 has nucleotide triphosphatephosphatase activity.
Iba et al. (9)showedthat thereisidentity among the ends ofthe threegenomicsegmentsfor17basesatthe3'end and for18basesatthe5'end,with theexception ofadifference in the secondposition. We have shown that the identity at the
3'
endismoreextensivebetweenthesmalland medium segments(Gottliebetal., unpublished data). We nowshow thatthis istrueforthe largesegment as well. Although the exact identity at the 3' end stops at nucleotide 18, there is overallsimilarity that continues until 80 nucleotides fromthe 3' end (Fig. 4). It appears reasonable that the regions of similarity should be involved in the regulation and mecha-nisms of transcription, replication, andgenome packaging.During infection, production of proteins P1, P4,and P7is almostidenticalon amolar basis. ProteinP2 isproducedat about10% ofthe rateoftheothers (25).Nonsensemutations ingene 7 arecompletely polarontheproduction ofP2(11). Itappears that the mechanism of control ofthesynthesis of P2 is translational coupling. The same motif has been de-scribed fortwoothergenepairs in4)6. Production ofP12is dependentonribosome loadingongene8,whichis immedi-ately upstream, and production of P5 is dependent on ribosomeloadingongene 9 (12). Inbothofthesecases, the downstreamproductis synthesizedat about10% of the rate ofthe upstream product, and nonsense mutations in the upstream genes are completely polar on the production of the downstreamgene product. In the three cases, the
ribo-some-binding
site upstream from the amino acid initiatingRWPSEGIYSGVTALMGATGSGKSITLNE 11 III GLNLKVKSGQTVALVGNSGCGKSTTVQL NINLSIKQGEVIGIVGRSGSGKSTLTKL DLNFTLRAGETLGIVGESGSGKSQSRLR DINLDIHEGEFVVFVGPSGCGKSTLLRM GVSLQARAGDVISIIGSSGSGKSTFLRC NINLDIAKNQVTAFIGPSGCGKSTLLRT NINLVIPRDKLIVVTGLSGSGKSSLAFD P4 111-138 Mdr HylB OppD MalK HisP PstB UvrA-1 411-438 488-505 40-557 21--48 24--51 28--5 5 16--43
FIG. 3. Comparison ofaportion ofthe amino acidsequence of
protein P4 with those ofa groupof proteins withnucleotide-binding
sites. The sequences are derived from thegenes for the multiple
drug resistance protein (Mdr), hemolysintransport protein (HylB),
oligopeptide permease (OppD), andtransportproteins for maltose
(MalK), histidine (HisP), and phosphate(PstB) (7). Asequencefrom
anATP-dependent UV repair protein(UvrA-1) (5)is also shown.
A
L GUAAAAAAACUUUAUAUAG 1111111111111111 M GGAAAAAAACUUUAUAUAU 1I1111111111111111 s GGAAAAAAACUUUAUAUAAB
L M S 3UCUUUUACCUGGAUUCUCUGUG UUUUCUACGUUGAGCUCCGUAUAUCCAUAAGUCCUWAGAUWUCUAAGGCGAGACUCGCUWUGCGAGCGUCCAAUAGGACGGCCCCCUCGGGGGCUC UCUCUCU I11111 111111111 111111111111111111I II 1111111111 111111 11111111111111111
UAAAUAAGUCCUUAGAUUUCUAACGCGAGACUCGCUUUGCGAGCAUCCAAUAGGAUGGCCCCUUCGGGGGCUCUCUCUCU
1111 11111111 111111111111 1111 1111111111
UAAACAAGUCCUGUAUAAC-AAGGCGAGACUCACUAUGUGAGCGUCCAAUAGGACGGCCCCWCGGGGGCUCUCUCUCU
FIG. 4. Comparison ofthe 5' (A) and 3' (B) ends of the three genomic segments. Inallcases,the 5' endisattheleft.
codon is either absent or
spaced
far away. The4)6
system also uses other means of control ofgeneexpression.
The production ofproteins codedon the middle segmentdiffers among the genes, although eachgene hasitsown ribosome-binding site. It is likelythat the strength of the interaction between ribosomes and some of the messages determines the level of production of individualproteins.
Inaddition,
there is a dramaticchange in thepatternofsingle-stranded RNA synthesis during infection (3). This is certainly in-volved in the turn onof translation ofthelateproteins.
Anumber of viruses contain segmentedgenomes of dou-ble-stranded RNA. These include reovirus, rotavirus, blue tonguevirus,and
4)6.
Inaddition, anumberofparticles
that replicate double-stranded RNA have beendescribed, nota-bly, the killer particle in Saccharomyces cerevisiae (6). Although the replication of4)6
RNA differs from most of these in that transcription of4)6 is by strand displacement, whereas the others use a conservative mechanism, we anticipatethat thereis considerablehomologyin theproteins ofthese particles. In particular, we expect to find thatthe replicaseproteinsarerelated.The complete nucleotide sequence of the three genomic segments has beenentered into the GenBank database. The accession numbers are M17461 for segment L, M17462 for segmentM, andM12921for segmentS.
ACKNOWLEDGMENTS
Partsof this workweresupported by Public Health Servicegrant GM 34352 fromthe National Institutes of HealthtoL.M. I.N. was supported in part by National Research Service award 5 T32 AI-07180 from the National Institute of Allergy and Infectious Diseases. M.R. was supported in partby the Academy of Finland during histenureatthePublic Health Research Institute.
LITERATURECITED
1. Bamford, D. H., M.Romantschuk, and P. J. Somerharju. 1987. Membrane fusion inprocaryotes: bacteriophage 4)6 membrane fuses with the Pseudomonassyringae outer membrane. EMBO J. 6:1467-1473.
2. Clewell, D. B. 1972. Nature of Col El plasmid replication in Escherichiacoli in the presenceof chloramphenicol. J. Bacte-riol. 110:\667-676.
3. Coplin,D.L., J.L. VanEtten, R. K. Koski, and A. K. Vidaver. 1975. Intermediates in thebiosynthesisof double-stranded ribo-nucleic acidsofbacteriophage4+6. Proc. Natl. Acad. Sci. USA 72:849-853.
4. Day, L. A., and L. Mindich. 1980. The molecular weight of bacteriophage
4+6
anditsnucleocapsid. Virology103:376-385. 5. Doolittle, R. F., M. S. Johnison, I. Husain, B. Van Houten, D. C.Thomas, and A. Sancar. 1986.Domainal evolution of a prokary-otic DNArepair proteinandits relationshiptoactive-transport proteins.Nature(London)323:451-453.
6. Fujimura,T., R. Esteban, and R. B.Wickner. 1986. In vitro L-A J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.75.276.614.666.2]GENOMIC SEGMENT L OF 4)6 1185 double-stranded RNA synthesis in virus-like particles from
Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 83: 4433-4437.
7. Gros, P., J. Croop, and D. Housman. 1986. Mammalian multi-drug resistance gene: complete cDNA sequence indicates strong homology to bacterialtransport proteins. Cell 47:371-380. 8. Higgins, C. F., I. D. Hiles, J. A. Downie, I. J. Evans, I. B.
Holland, L. Gray, S. D.Buckel, A. W. Bell, and M. A. Hermod-son. 1986. Afamily of related ATP-binding subunits coupled to manydistinct biological processes in bacteria. Nature (London) 323:448-450.
9. Iba, H., T. Watanabe, Y. Emori, and Y. Okada. 1982. Three double-stranded RNA genome segments of bacteriophage 4)6 havehomologousterminal sequences. FEBS Lett. 141:111-115. 10. Ktistakis, N. T., and D. Lang. 1987. The dodecahedral frame-work of the bacteriophage 4)6 nucleocapsid is composed of proteinP1. J. Virol. 61:2621-2623.
11. Lehman, J. F., and L. Mindich. 1979. The isolation of new mutantsof bacteriophage4)6.Virology97:164-170.
12. McGraw, T., L. Mindich, and B. Frangione. 1986. Nucleotide sequenceof the small double-stranded RNA segment of bacte-riophage4)6: novel mechanism of natural translational control. J.Virol. 58:142-151.
13. McGraw, T., H. Yang, and L. Mindich. 1983. Establishmentof aphysical and geneticmapforbacteriophage PRD1. Mol. Gen. Genet. 190:237-244.
14. Messing, J. 1983. New M13 vectors for cloning. Methods Enzymol. 101:20-80.
15. Mindich, L.1978. Bacteriophages that containlipid, p. 271-335. In H.Fraenkel-Conrat, and R. R. Wagner (ed.),Comprehensive virology, vol. 12. Plenum Publishing Corp., New York. 16. Mindich, L., and R.Davidoff Abelson. 1980. The
characteriza-tion ofa 120S particle formed during 4)6 infection. Virology 103:386-391.
17. Mindich, L., and J. Lehman. 1983. Characterization of 4)6 mutants that are temperature sensitive in the morphogenetic
protein P12. Virology 127:438-445.
18. Mindich, L., G. MacKenzie, J. Strassman, T. McGraw, S. Metzger,M.Romantschuk, andD.Bamford.1985. cDNA clon-ing of portions of the bacteriophage4)6genome. J. Bacteriol. 162:992-999.
19. Olkkonen,V.M.,andD. H. Bamford. 1987. Thenucleocapsid of the lipid-containing double-stranded RNA bacteriophage 4)6 contains a protein skeleton consisting of a single polypeptide species. J.Virology 61:2362-2367.
20. Partridge, J. E., J. L. Van Etten, D. E. Burbank, andA. K. Vidaver. 1979.RNApolymeraseactivity associated with bacte-riophage4)6nucleocapsid. J. Gen. Virol. 43:299-307.
21. Revel, H. R.,M. E. Ewen, J. Busslan, and N. Pagratis. 1986. Generation of cDNA clones of the bacteriophage4)6 segmented dsRNAgenome:characterization andexpression ofLsegment clones. Virology 155:402-417.
22. Sanger, F., A. R. Coulson, G. F. Hong, D. F. Hill, and G. B. Petersen. 1982. Nucleotide sequence ofbacteriophage A DNA. J. Mol. Biol. 162:729-773.
23. Sanger, F.,S.Nicklen,and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
24. Sinclair, J. F., and L. Mindich. 1976. RNA synthesis during infection with bacteriophage4)6.Virology 75:209-217. 25. Sinclair, J. F., A. Tzagoloff, D.Levine,and L. Mindich. 1975.
Proteins of bacteriophage4)6. J.Virol. 16:685-695.
26. Vidaver, A. K., R. K. Koski, and J. L. Van Etten. 1973. Bacteriophage 4)6: a lipid-containing virus of Pseudomonas phaseolicola.J.Virol. 11:799-805.
27. Walker, J. E., M. Saraste, M. J. Runswick, and N. J. Gray. 1982.Distantly related sequences in the a and a subunits ofATP synthetase,myosin, kinases and otherATP-requiring enzymes anda commonnucleotidebinding fold. EMBO J. 1:945-951. 28. Yang, Y.,and D.Lang. 1984.Electronmicroscopy of
bacterio-phage 4)6 nucleocapsid: three-dimensional image analysis. J. Virol. 51:484-488.
VOL.62, 1988