Copyright 0 1973 AmericanSociety for Microbiology Printed in U.S.A.
Proteins
Specified by Herpes Simplex Virus
XI.
Identification and Relative Molar
Rates of
Synthesis of Structural and
Nonstructural Herpes Virus Polypeptides in the Infected Cell
ROBERT W. HONESS AND BERNARD ROIZMAN
Department ofMicrobiology, The University of Chicago, Chicago, Illinois60637
Received for publication 14August 1973
Analyses of polypeptides made in HEp-2 cells infected with herpes simplex
virus type 1 by high-resolution polyacrylamide gel electrophoresisrevealed the
synthesis ofat least 49 infected cell polypeptides (ICP) ranging in molecular
weight from 15,000 to280,000. Evidence for virusspecificity basedon increased
rates of synthesis postinfection, immunological specificity, and viral
control ofmobility andrateofsynthesiswasavailable for 47 of the ICP. These 47
polypeptides can account for 75% ofthe virus genetic information assuming a
DNAmolecular weight of10"andasymmetric transcription. On the basis of their
mobility relative tovirion proteins, the ICP were classified asstructural (S, 23
polypeptides), nonstructural (NS, 16 polypeptides), and unassigned (U, 10 polypeptides). Analysis of the synthesis of the ICP revealed the following. (i)
Rapidposttranslational cleavages of HSV proteinswerenotdetected; inparallel
experiments rapid posttranslational cleavages were readily demonstrated in
poliovirus-infected cells and thesewere blockedbyprotease inhibitors. (ii) Slow
posttranslational changes in the mobility of at least two polypeptides were
observed.(iii) Analysis of theratesofsynthesis of ICP examinedatfour intervals
postinfection revealed regulation of the patternandamountof ICPsynthesized.
ICP formed six classes (AtoF) differing in their kinetics ofsynthesis. S and NS
ICP were distributed nonrandomly amongthese classes. Thus, of thesum ofS
protein amino acid sequences apportioned among these kinetic classes, 47%,
constituting class A and comprising "late" structural proteins, were
character-izedby progressively increasing ratesofsynthesis until at least 12 h
postinfec-tion; whereas "early" structural proteins constituting class C, amountingto31%
of the totalamino acidsequences, weresynthesized with initially increasingrates
until 4 hpostinfection and with decliningratesthereafter. NS polypeptides and
remaining S polypeptides were distributedamongthe other kinetic classes-B,
D,E, and F. Control of protein abundance wasevident in that the polypeptides
werenotmade inequimolaramounts.However, Sand NS polypeptides couldnot
be differentiated onthe basis of their molar ratesofsynthesis. The bulk of the
detectedpolypeptides didnotdifferbymorethaneightfold in their molarratesof
synthesis.
Herpesvirus replication requires the
im-plementationof two programs,the transcription of viral DNAand the translation ofviral
mes-sages. Recentpapers (6, 7, 21) from our
labora-tory have documented the existence oftemporal
controlsofviral DNAtranscriptionand controls
of molar abundance of the RNA transcripts
accumulatingin theinfectedcell. In this paper
wereport the results of experiments designed to
outline the time ofsynthesisand abundance of
thetranslational productsofthe virus.
As aprefacetothis report, it seems desirable
tosummarize the backgroundand some of the
problems encountered inthis work.
Previous attempts to analyze the
polypep-tides madeininfected
cells,
definedasinfected cellpolypeptides
(ICP),
both inthislaboratory
(23) and in others (1, 10) arecurrentlyof limited
value for two main reasons. First, the
tech-niques of polyacrylamide gel electrophoresis employing the continuous buffer system (19), combined with the estimation ofradioactivity
within slices of fractionated gels, were
inade-quate to resolve the large number ofproteins
made in the
herpesvirus-infected
cell.Second,
the procedure then used for the purification of
1347
on November 10, 2019 by guest
http://jvi.asm.org/
herpesvirus particles was inadequate (14), and
hence no meaningful classification of proteins as either structural or nonstructural could be
made. The availability of techniques for the
purification of enveloped nucleocapsids
(vi-rions) (24), forhigh resolution electrophoresis of
proteins on polyacrylamidegels (3, 12), and for
quantitative assay of separated proteins (5) has
enabledus toundertake the present studies.
The major difficultyencountered in the
cur-rent studies of ICP is the differentiation
be-tween polypeptides specified by the virus and
the host, although the bulk of host proteins
ceases tobe made after infection (23).
Compel-lingevidence that a polypeptide is virus specific
is its synthesis in a cell-free amino acid
incor-porating system programmed by mRNA
com-plementary to viral DNA. Sincethis is not yet
feasible,wehavedefined ICPasvirusspecific if
they meet one or more ofthefollowingcriteria,
i.e., (i) stimulation in the rate of synthesis
postinfection in the face of general decline in
host proteinsynthesis; (ii) variations in
proper-ties of ICP as a function of the virus strain
infecting the cell, and (iii) immune
precipita-tion ofICP
by
antisera reactivesolely withvirusantigens.
In a similar manner, the differentiation
be-tween structural and nonstructural
polypep-tides
ideally
requires the rigorous demonstra-tion ofchemicalidentity between the structuralproteins ofthevirus, previouslydesignated(24)
as virion proteins (VP), and their putative
analogues amongtheICP.In practice we have temporarily defined ICP as structural or
non-structural solely onthe basis ofelectrophoretic mobility relative to VP, pending a more
com-plete characterization oftheseproteins.
MATERIALS AND METHODS Solutions and chemicals. Maintenance medium forinfected cellswasmixture199supplemented with
1%calfserum.Labelingmedium consisted of mixture
199 containing one-tenth the usual amountsof leu-cine, isoleucine, and valine and supplemented with
1% dialyzed calf serum. Labeled amino acids were
purchased fromSchwartz/Mann, Orangeburg, N. Y., andwere as follows: L-isoleucine-U-_4C,
L-leucine-U-14C, andL-valine-U-14C, all withspecific activities of
approximately 300mCi/mmol; L-isoleucine-3H,
L-leu-cine-3H, andL-valine-8Hwithspecific activities from
1to 30Ci/mmol;andamixture ofthirteen"4C-labeled
aminoacids(reconstituted proteinhydrolyzate,
com-posed of alanine, arginine, aspartic acid, glutamic acid, isoleucine, leucine, lysine, phenylalanine, pro-line, serine, threonine, tyrosine, and valine with
specific activities of about 150 to 450 mCi/mmol).
Tolylsulfonyl-lysyl chloromethyl ketone (TLCK), a
specific inhibitor of trypsin, and
tolylsulfonyl-phenylalanylchloromethylketone(TPCK),aspecific
inhibitor of chymotrypsin activities (20) were ob-tained fromNutritional Biochemical Co., Cleveland, Ohio.
Cells. Humanepidermoid carcinomano.2(HEp-2) cells were grown in Eagle minimal essential medium (EMEM) supplemented with 10%calf serum, 0.001% ferricnitrate, and 1%sodium pyruvate.
Viruses. Most of the experiments described in this paper were done with the F prototype of HSV-1 [HSV-1(F)]. Other strains analyzed insome ofthe studies were HSV-l(mP) and HSV-2(G) correspond-ing to a laboratory strain of HSV-1 and the G-prototype of HSV-2, respectively. The origin and some properties of these strains have been reported elsewhere (4). Procedures for production of HSV strains were asdescribedby Roizman and Spear (17) with the exception that HSV-1(F) and HSV-2(G) have beenpassaged a maximum of four times at low multiplicity priortotheir use inthehigh-multiplicity infectionsdescribed in this study. Titrations of infec-tious viruswere performedas describedby Roizman and Roane (15).
An attenuated vaccine strain ofpoliovirus type 1 was employed as a positive control in experiments dealing with rapid posttranslationalcleavages of virus polypeptides. Media and procedures employed for infection and radiolabeling of poliovirus-infected HEp-2 cells were identical to those described for
HSV-infected cells.
Purification of isotopically labeled, enveloped virusparticles. Infected cellswerelabeledfrom 4 to 24 or 30 h postinfection with either 3H-leucine, isoleucine, andvaline (1 to1.5MCiofeachaminoacid per ml oflabelingmedium)or"4C-leucine, isoleucine, and valine (0.08gCiofeach aminoacid perml)in50
ml of labeling medium per 2 x 108 infected cells. Virionswerepurifiedfromlabeled cellsessentiallyas
describedbySpear and Roizman (24), except that the flotationthrough the discontinuoussucrosegradient, which followed thebanding inadextran-10gradient,
was replaced by a second dextran-10 gradient sedi-mentation. Particlespurifiedby thismodified proce-dure had a polypeptide profile similar to that ob-tainedusing theoriginalprocedure. Particlespurified
in this way consisted of approximately 95% of en-veloped particles (unpenetrated by neutral phos-photungstate negative stain) with particleto infectiv-ity ratios ofapproximately100.
Labelingofproteinssynthesized byinfected and
uninfected cells. ConfluentHEp-2cellmonolayersin
tissue culture flasks(approximately2 x 106 cellsper flask)wereexposedto20PFUof viruspercell(in20 ml ofmaintenance medium) or were mock infected with2.0mlofthismedium. Inoculated cultureswere
incubated withconstantagitation for1h at 37C,and thereaftervirus or mock inoculaweredecanted. The monolayerswerethenrinsed(5.0 mlof maintenance
medium perflask), replenished with5.0mlof mainte-nancemedium perflask,andreincubatedat 37C.For
labeling, the cultures were rinsed and then replen-ished with labeling medium containing "4C-leucine,
isoleucine, and valine (0.3to 1.7 gCi ofeachamino
acid perml).Atthe end ofthelabeling periodthecells were either: (i) rinsed with ice-cold phosphate-buf-fered saline(3 x 5.0ml/flask)toterminate
incorpora-J.
on November 10, 2019 by guest
http://jvi.asm.org/
PROTEINS SPECIFIED BY HERPES SIMPLEX VIRUS
tion ("pulse") and then stripped from the flask, denatured and solubilized by heating at 80 C with a small volume of 1% sodiumdodecyl sulfate (SDS) for subsequent trichloroacetic acid precipitation or 2% SDS, 5%
0-mercaptoethanol,
0.05M Tris-hydrochlo-ride (pH 7.0) for subsequent polyacrylamide gel electrophoresis, or (ii) rinsed (3x5.0 ml), replenished with maintenance medium (at 37 C), and reincubated in theabsence of exogenous labeled precursors prior to terminatingincorporation, asabove ("pulse chase"). The timesatwhichlabeled precursors were added and removed, etc., are stated as the times in hours after theaddition of the virus inoculum.Radio-immune precipitation. The radio-immune precipitation reactions for analysis of herpesvirus-specific polypeptides were performed as described by Honessand Watson (J.Gen. Virol., in press). Briefly, the supernatant fluid obtained after sedimentation (35,000 rpm, 1 h, in a Beckman SW40 rotor) of a homogenate of infected HEp-2 cells, which had been labeled with "C-amino acids from 4.5 to 6.5 h postinfection, was mixed with an excess of a rabbit hyperimmune serum specific for herpes simplex virus antigens and incubated at +4 C overnight. The re-sulting immune precipitate was sedimented (3,500 rpm for 15 min), washed thoroughly by resuspension and sedimentation in ice-cold phosphate-buffered saline, and finally resuspended and solubilized by heating in 2%SDS, 5% ,B-mercaptoethanol, and 0.5 M Tris-hydrochloride (pH 7.0) at 80Cfor 10min.The solubilized immune precipitate was then subjected to electrophoresison apolyacrylamide gel. The immune sera,kindly donated by P. Wildy and D. Watson, were hyperimmune rabbit sera prepared by inoculation with rabbitkidney cells infected with HSV-1,ashas been described in detail (27). These sera gave up to 12 precipitin bands in agargel radial immunodiffusion
testsagainst infectedHEp-2, RK13,orBHK-21cells, but hadnodemonstrable reaction against uninfected cells or componentsoftissueculturemedia, and did notprecipitate any significant radioactivity froman artificial mixture of labeled uninfected cells and unlabeled infected cell antigensamples (R. W.
Hon-essand D. H.Watson, J. Gen. Virol., in press). Analytical procedures. Incorporation of radioac-tive amino acids into acid-insoluble material was determined by precipitation with cold 5% trichlora-cetic acid. The precipitates were collected on mem-brane filters (Millipore Corp. type GA, 2.5 cm) and washed, and the radioactivity was estimated.
Polyacrylamide gel electrophoresis. The electro-phoretic, staining, and autoradiographic techniques were as described previously (W. Gibson and B. Roizman, J. Virol., submitted for publication, 24),
employing the discontinuous buffer system (2, 13)
modifiedbytheinclusion ofSDS (3, 12).
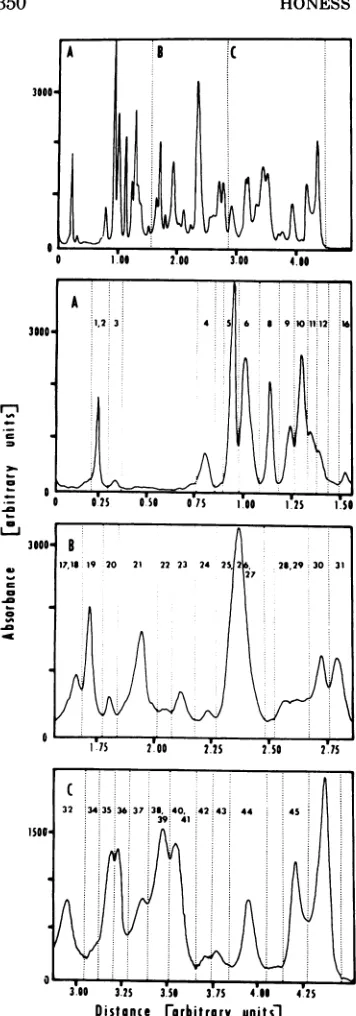
Densitometry of stained gels and autoradio-grams andcomputer-ai'dedanalysis of absorbance tracings. Absorbance tracings of stained gels and autoradiograms wereobtainedwith aGilford record-ing spectrophotometer equipped with a gel scanner
attachment. The voltage output from the photocell
was also recorded by a General Automation 16/45 digital computer anddisplayed as a plot of
absorb-ance (voltage) as a function of distance migrated
(time of scan) on a Tektronix oscilloscope. The computer wasprogrammed to expand regions of the initial profile as determined by the operator and to compute the areas under the tracing for individual bands defined by vertical lines set parallel to the absorbance (voltage) axis. The results of a typical analysis of proteins made in HSV-1(F)-infected HEp-2 cells are shown in Fig. 1; the upper panel shows thecomplete profile and the regions (A,B, C) selectedfor expansion are shown in the three lower panels. The lines intersecting theexpanded profilesin
the lower panels demarcate the areas ofthe bands calculatedby the computer. The bandsarenumbered inaccordance with the designationsofHSV-1(F) ICP which are described in Results. Thepeakof radioac-tivity migrating furthest towards the anode(the right side of all profiles shown in this paper) in this and subsequent gels represents material too small to be sieved at aparticular gel strength and which therefore migrates with the free-SDS front (3) close to the bromophenol blue (BPB) dye marker. In the 8.5% gel showninFig.1allICP smaller than ICP45migrate in this position.
RESULTS
Enumeration and differentiation between structural and nonstructural polypeptides synthesized in the infected cell. In the first
series of experiments the mobilities of labeled
ICP and VP were compared by electrophoresis
either as an artificial mixture
(coelectrophore-sis) or separately, but on the same polyacrylam-ide gel slab (parallel electrophoresis). Both
techniques (Fig. 2 and 3) showed that there
were moreICP than there were VP and that the
relative mobilities of ICP and VP were the same, whether obtained by a combination of coelectrophoresis and autoradiography (Fig. 2a
andb) or by autoradiography of parallel
separa-tion on slab gels (Fig. 3). Mobilities of ICP
relative to VP were independent of
polyacryl-amide gel concentration (Fig. 3).
However, as has been noted previously (24), autoradiography of whole gels resolved more
bands than measurements of radioactivity in
gel slices. For example, in Fig. 2a a maximum of
eight bands was defined by variation in the "C-radioactivity of fractions 15 to 40, and a
number of these peaks were defined by
eleva-tionofradioactivityinasingle slice.Incontrast,
12 well resolved bands were apparent in the
autoradiogramofthecorrespondingregion (Fig. 2b). On the basis of these results,
autoradiogra-phy of parallel separations was the method
chosen forsubsequent experiments.
Analyses such as those showninFig. 2and 3
have shown that cells infected withHSV-1(F)
synthesize at least 49 polypeptides, whereas
under the same conditions only 27 bandswere
resolved inseparations of proteins frompurified
virions. TheseICPwereassigned numbersfrom
1349 VOL.12,1973
on November 10, 2019 by guest
http://jvi.asm.org/
Distance Corbitrary
units]
FIG. 1. Illustration ofcomputerplanimetry of the proteins separated in polyacrylamide gels. The top panelshows the absorbancetracing ofan
autoradio-gram of HSV-1(F) ICPlabeled with'IC-amino acids from3.5to8.7hpostinfection.A,B, andCindicate
those regions of theprofile shown in the expanded forminthe lowerthreepanels.In eachpanel theareas
boundedbytheabscissa,absorbanceprofile, and the broken lines parallel to the ab )rbance axis were
1to 49 in order ofdescendingmolecularweight,
and independent of the established numbering
system for virionproteins (i.e., VP 1-24 [24]).
One objectiveofthisstudywas toclassifythe
ICP as structural and nonstructural and to
examinetheirsynthesis.For the purposes of this
paper the basis for the classification was as
follows. (i) All parallel-run comparisons on
whichassignments weremade included proteins
of purified virions in amounts sufficient to
permit the detection ofeventhose VP presentin
smallamounts (e.g., VP4, VP6,and VP
9).
(ii)AnICP with thesame electrophoretic
mobility
as a VPwasconsidered ananalogueofthis VP andwas
designated
a"structural"(S)
polypep-tide. (iii) ICP with electrophoretic mobilities differingfromthoseofVP,but migrating under
a broad VP band, were designated
"unas-signed" (U)
polypeptides.
(iv) ICP with electro-phoretic mobilities clearly different from those of VP were designated "nonstructural"(NS)
polypeptides.
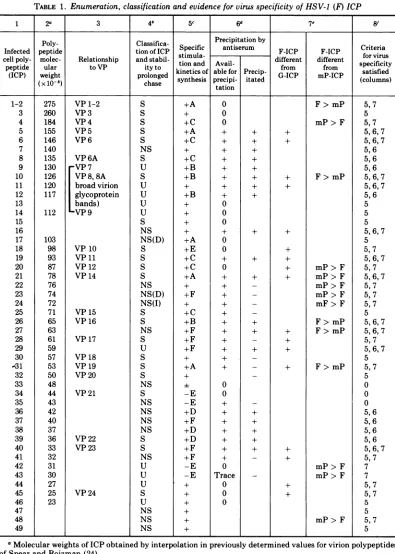
Table 1 (columns 1 to 4) summarizes the
enumeration and classification of ICP
by
the above criteria. Of the 49ICP,
10 were consid-eredunassigned,
16werenonstructural,
and the remainderwerestructural.Evidence for the virus
specificity
of ICP. Evidence that mostoftheICPwehave identi-fiedarespecified
by viral genetic information isbased on three types of experiments dealing with comparisons of
polypeptides
synthesized in infected and uninfected cells, the specific precipitation ofICPby
antiserareactingsolely withvirus-specific
antigens,andcomparisonsofHSV-1(F) ICP with ICP made in cells infected with other strains and variants ofherpes
sim-plex virus.
Comparison
ofpolypeptides synthesized
ininfected and uninfected cells.
Figure
4showsatypical
result ofacomparison
ofthepolypep-tides
synthesized
overashortperiod
ininfected and uninfectedHEp-2 cells. In this experiment replicate cultures of HEp-2 cells were labeled with"4C-amino
acids from 4.5 to 5.0 h afterinfection with HSV-1(F) or after mock
infec-tion.
Equivalent
samples of labeled infected anduninfected cellswere thenharvested, solu-bilized, and subjected to electrophoresis on 8.5% acrylamide gels. Visual examination ofgels stained with Coomassie brilliant blue re-calculatedbythecomputer. Thenumbers above the profile refertothe numericaldesignation of the ICP contained in each band. The arbitrary unitson the
ordinatearerelatedtoabsorbancebytheequation1.0
absorbance unit equals 1,139 arbitrary units. One arbitrary unit of distance(abscissae)wasequalto2.0
cm. r-i
1--
30-
6-F, .in LI
4D
1--.Jo
0
%A
.M
-W
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.497.61.239.58.566.2]c c
386;
02004~~~~~~92000
6~~~~~~~~~~3
b F
10 9 26-27
c 39
~~~~~~~~~~~~~35
36
0
31
41
21 4445
32
1629
4 ~~~23 37
4243 6
0 20 40 60 80 100
Fractions
FIG. 2. Distributionofradioactivity inasliced gel(a)andtheautoradiogram ofanidentical polyacrylamide
gel (b)containingelectrophcretically separated proteins from 3H-amino acids-labeledpurified virions andan
infected celllysatelabeled with14C-amino acids from4.25to6.5hpostinfection. In the radioactivity profileof
the slicedgel (upperpartofFig.2a),polypeptides ofthevirion (3H-amino acids,opencircles and broken line)
are indicated by thelarger numerals enclosed withinrectangular frames, andthe infected cell polypeptides
(ICP), labeledwith "4C-amino acids,areindicated by the closed circles, the solid line,and smaller numerals. The lower part of the figure (b) is an absorbance tracing of the autoradiographic image (i.e., "4C-amino
acids-ICP) ofanidenticalgel. Thesmallnumeralsonthis profilearethose of the ICP. Electrophoresison8.5%
cylindrical gels.
vealed a large number of bands, too closely vealed any significant difference between the
spaced to be well resolved by densitometry. stained profiles ofelectrophoretically separated
Neither visual inspection nordensitometry re- proteins of total infected or uninfected cells
on November 10, 2019 by guest
http://jvi.asm.org/
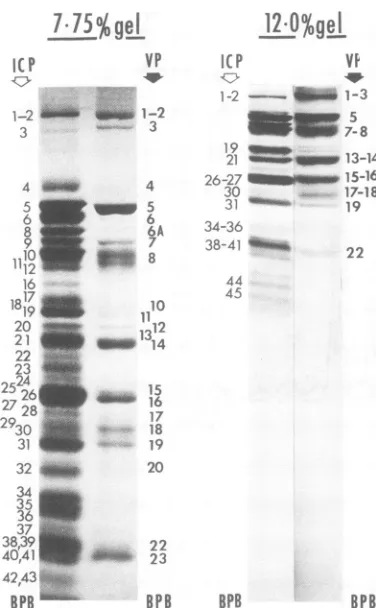
[image:5.497.104.396.61.527.2]7l7lS%gej
IC P1-2
"ii
3
4 5 62
278 930 1110 r
312 16
19_
2
22
4243 25
26_4
2728
°2930
32_
potnecin Sou 34_ 36_
38i3e
40'41__
BPB
p eG. 3. Autoradio,
phoreticallyseparatt
polypeptides (purifie
20hpostinfection)a
labeled withefC-a
postinfection.Solubi
infected cells were parallelon7.75%an
processed forautorao
ductions of photogr
gram,andtherefore,,
darkerareaatthetol
illumination during printedwithdifferen
twosamples.
(Fig. 4, middletrao
ographic images o
differentiated(Fig
The distribution
uninfected cell pr(
total protein (mic
Fig. 4), with the i
grating with
polyl
molecular weight
lysates contained
and their distribu
the distribution of proportion of "C-f
12.0%ge
l
into components with molecular weights greater than 50,000.VP ICP VI In view of thevery
large
number ofpolypep--W - tide
species
synthesized
in uninfected cells for1-2- m 1-3 nearly every ICP there is a component in the
1-2 58 uninfected cell characterized
by
the same or19
7mm-8
similarelectrophoretic mobility.
Attempts
to21 U M 13-14 differentiate between
polypeptides synthesized
4
3026-2
-1516 in infected and uninfected cells solely on the5
31
019-1
basis of directcomparisons
oftheirelectropho-6A 34-36 retic mobilities therefore seemed unprofitable.
7 38-41 2
However,
the fact that theprofiles
ofproteins
8 22 synthesized are different may be used as
evi-44 dence for selective inhibition or stimulation
10
(induction) to account for these differences.12
Quantitativecomparisons
of amino acidincor-1314
poration over similarregions
of infected anduninfected cell
profiles (top
and bottomtrac-15 ings of Fig. 4) indicated a higher rate of
synthe-16 sis in the infected cell over regions of ICP other
17 than those indicated
by "h"
inFig.
4(and
the18
19
components travelling
at thedye
front inFig.
20 4). Thus, incorporation into ICP 34, 35, and 42,
43 was
quantitatively comparable
to that ob-served over the same region ofthe uninfected cell.22 This
type
ofcomparison
may be termed the23 criterion of induction; it provides some evidence
for virus specificity of ICP other than those
BPB BPB
BPB
comigratingwith prominentcomponents ofthegraphic comparisons of electro uninfected cells. These conclusions are
rein-ed14C-amino acids-labeledvirion
forced
by
the results of analyses of thekinetics
'd
fromcells and labeledfrom 3to of synthesis of ICP considered later in the text.ndpolypeptides of infected cells Precipitation of ICP by antiserum specific
mino acids from 3.5 to 8.7 h for virusantigens.In thisexperiment, infected
ilized labeled virions andlabeled cells were labeled with "4C-amino acids from
subjected to electrophoresis in 4.25 to 6.25 h afterinfection withHSV-1(F). A
yd12% slabgels. These were then sample of the infected cell homogenate was
diography. The figures are repro- sedimented (35,000 rpm, 1 h, in a Beckman
,aphs of the original autoradio-
SW40
rotor), and the supernatant fluid, whichsuffer fromlossofresolution. The contained 35% of the acid-insoluble radioactivity
pofthe7.75%gelis dueto uneven of the total cell
homogenate,
was mixed withaphotography. The
12%o
gel was o h oa elhmgnt,wsmxdwtt
exposure
periods for each of thesufficient
volume
of a hyperimmune rabbitserum
specific
forherpes simplex
virus anti-gens togive
maximalprecipitation
oflabeledcomponents (R. W. Honess and D. H. Watson,
cing). However,the autoradi- J. Gen.
Virol.,
inpress).
Theresulting
immunefthese
gels
could bereadilyprecipitate
contained 25 to27% oftheradioac-4, topand bottomtracings).
tivity
presentin thesupernatantfluid.Samples
of
"4C-amino
acids-labeled of thetotal infected cellhomogenate,
thesuper-oteins coincided with that of natant
fluid,
and the immuneprecipitate
wereidle
and bottom tracings of thensubjected
toelectrophoresis
on 8.5%poly-majority ofradioactivity mi-
acrylamide gel cylinders. Autoradiograms
ofpeptides less than 50,000 in these
gels
are shown inFig.
5. Thesamples
ofIn contrast, infected cell the total infected cell
homogenate
and thesu-fewer labeled polypeptides pernatant fraction were derived from a
com-tion in thegel didnotreflect parable number ofcells; thesample ofthe
im-thetotal cellprotein.Alarge muneprecipitatewasobtained fromavolume of
amino acid
incorporation
was supernatant fluid sixtoeight
timesgreater than J.on November 10, 2019 by guest
http://jvi.asm.org/
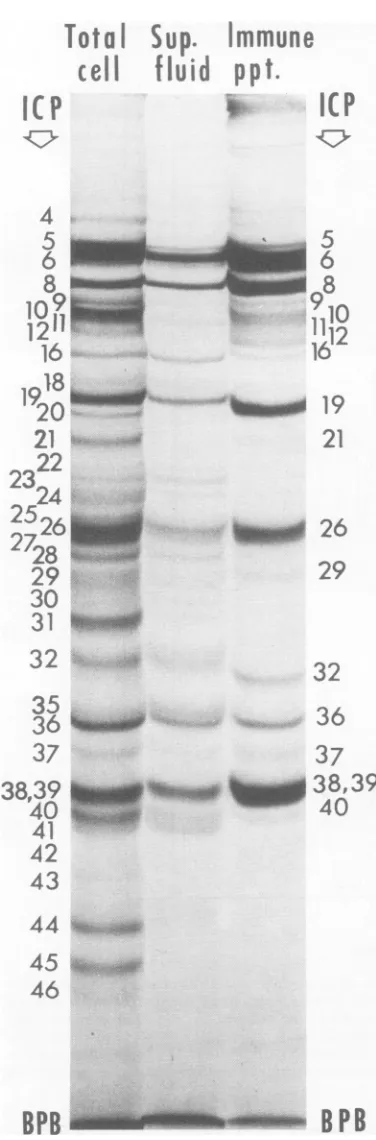
[image:6.497.58.246.71.375.2]PROTEINS SPECIFIED BY HERPES SIMPLEX VIRUS
1353
TABLE 1. Enumeration, classification and evidenceforvirusspecificity ofHSV-1 (F) ICP
1 2a 3 4b 5C 6 7e- 8'
Poly- Classifica- Precipitation by
Infected peptide tionofICP
Specific
antiserum F-ICP F-ICP Criteriacellpoly- molec- Relationship and stabil- stimula- different different for virus
peptide ular toVP ityto tionand Avail- from from specificity (ICP) weight prolonged kinetics of able for Precip- G-ICP mP-ICP
satisfied
(x10-8) chase synthesis precipi- itated (columns)
tation 275 260 184 155 146 140 135 130 126 120 117 112 103 98 93 87 78 76 74 72 71 65 63 61 59 57 53 50 48 44 43 42 40 37 36 33 32 31 30 27 25 23 VP 1-2 VP 3 VP4 VP 5 VP 6 VP 6A VP 7 VP8, 8A broad virion glycoprotein bands) VP9 VP 10 VP11 VP 12 VP 14 VP 15 VP 16 VP17 VP18 VP19 VP20 VP21 VP 22 VP23 VP 24 S S S S S NS S U S U U U U S NS NS(D) S S S S NS NS(D) NS(I) S S NS S U S S S NS S NS NS NS NS S S NS U U U S U NS NS NS +A + +A +C +C + +C +B +A + +C +E +E +E +E +A +F +F +F +A + +F +D +D +F +F -E -E 0 0 0 + + + + + + + + 0 0 0 + 0 0 + + + + + + + + + + + + 0rc 0 + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + F> mP
mP> F
F> mP
mP> F mP> F mP> F mP>F mF> F F> mP F>mP + F>mP + + mP> F mP> F mP> F 5,7 5 5,7 5,6,7 5,6,7 5,6 5,6 5,6 5,6,7 5,6,7 5,6 5 5 5 5,6,7 5 5,7 5,6,7 5,7 5,6,7 5,7 5,7 5,7 5 5,6,7 5,6,7 5,7 5,6,7 5 5,7 5 0 0 0 5,6 5,6 5,6 5,6 5,6,7 5,7 7 7 5, 7 5,7 5 5 5,7 5 VOL. 12, 1973
1-2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 -31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49
aMolecularweightsofICP obtainedby interpolationinpreviouslydetermined values for virionpolypeptides
ofSpearand Roizman(24).
S, structural; NS, nonstructural; U, unassigned. See text for criteria
applied
in the classification. Inadditiontothe 24 polypeptidesdescribedby Spearand Roizman(24), theseanalysestookcognizanceoftwo additional polypeptidesfoundin purifiedvirions (Heine, Honess, andRoizman, manuscriptinpreparation).
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1.-Continued
These were VP 6A, a minor virion polypeptide which co-electrophoresed with ICP 8, and a glycoprotein designatedVP 8A.Duetopoor resolutionofVP7, 8,and 8A, themobility analogueofVP 8Aamong the ICP is uncertain and alloftheICP included within this mobility rangearethereforeclassified as S (ICP 10 which co-electrophoreses with VP 8) or as U. (D), Components declining or disappearingon prolonged chase. (I), Components increasingorappearingon prolonged chase.
c+, Aminoacid incorporation into components migratingin these regionswas higherin infectedthan in
uninfectedcells. -,Aminoacidincorporation into componentsmigratingintheseregionsininfectedcellswas
comparable to,orlowerthan, that observed in thesameregions of uninfected cells. See Fig. 3 and relatedtext.
LettersA toF refertothe patternsofsynthesisasshown inFig.11.Absence ofaletterdesignation indicates that the patternofsynthesis isuncertain.
dInthe left column("Available forprecipitation"), 0 and + indicate, respectively, undetectable amounts and the presenceof aparticular ICPinthesupernatant fluid. In theright column, + indicates precipitated and - indicatesnotprecipitated, by antiserum. See Fig.5and relatedtext.
eIn the comparisonofHSV-1(F) andHSV-2(G) (left column), + indicates any HSV-1(F) ICPforwhich there
was no exactcounterpart in mobilityoramount in HSV-2(G)-infected cells. Comparisons of HSV-1(F) and HSV-1(mP) (right column) indicate significantquantitativedifferences between ICP with identical mobilities producedincellsinfected with thetwoviruses, i.e., F > mP referstoapolypeptide producedinlargeramounts inFthaninmP-infected cells.
fNumbers refer to the columnssummarizing those criteria forvirusspecificity which were satisfiedby a
particular ICPofHSV-1(F). Forexample, ICP5satisfiedthe criterionofinduction(column 5), possessed viral antigenic specificity (column 6), and its mobility was affected by virus type-specific variation (column 7). Symbol 0 indicates thatnoneofthese criteria werefulfilled.
that subjected to electrophoresis on the
com-paniongel.
By inspection of Fig. 5 it may be seen that
certain polypeptides of the homogenate
parti-tionedselectivelywith thepelletafter
sedimen-tation and were in relativelylowconcentration
(ICP9 to 12, 20, 21,and31)or werenotdetected
(ICP44andICP 45)inthesupernatant fluid.Of
the polypeptides present in the supernatant
fluid,some,i.e., ICP 5, 6, 7, 8, 9 to 12, 19, 26, 32,
36, 38, and 39 were
precipitated
efficiently,whereas others, i.e., ICP 16, 21, 28, 29, 37, and
40 were
precipitated
less efficiently, and some,ICP22 to 24and41, were notprecipitatedatall.
Although precipitation
by
such antisera isadditive evidence for virus specificity, the
fail-uretoprecipitatea
particular
componentisnotevidence that thiscomponent is host
specified.
Virus type and intratype
(strain)
specific
differences inICP. In theseexperiments
repli-catemonolayersofHEp-2cellswereexposedto 20 PFU of either
HSV-1(F), HSV-l(mP),
orHSV-2(G) per cell and labeled with "C-amino
acids from 3.5 to 8.7 h postinfection. The cells
were then harvested and solubilized, and the
proteins were subjected to electrophoresis in
parallel. Theresults ofautoradiographic
analy-ses (not shown) may be summarizedbriefly as
follows. (i) ICP
synthesized
in cells infectedwith HSV-1(F) and HSV-l(mP) were largely
congruent in both number and electrophoretic
mobilitybutdifferedinthe relative amounts of
a small number of polypeptides which were
made within the same time interval. Thus,
significantly moreofICP1-2, 10, 26, 27,and31
were present in
HSV-1(F)-infected
cells.Con-versely,HSV-1 (mP) -infected cells accumulated
moreofICP4, 21, 22, 23, 24, 42, 43,and 48than
did HSV-1(F)-infected cells. (ii) Although the general appearance of profiles ofproteins
syn-thesized in HSV-1(F)- and HSV-2(G)-infected cellsweresimilar, thereweredifferences bothin
mobility and molarconcentrations of polypep-tides migratingin corresponding regions ofthe twoelectrophoretic separations.For ourpresent
purpose ofestablishing the virus specificity of
the ICP in HSV-1(F)-infected cells, it is
suffi-cient to indicate thoseHSV-1(F) ICPforwhich
an exact counterpart, either in mobility or
molarconcentration,was absent fromlysatesof
HSV-2(G)-infected
cells. Thus, HSV-2(G)-infected cells lacked exact counterparts toHSV-1(F), ICP5, 6, 10, 11, 16, 18, 19, 21, 27, 28,
29, 40, 41, 44, and45.
The evidencefor virusspecificityofHSV-1(F) ICP which we have adduced in the preceding section is summarizedincolumns5, 6, 7, and8 ofTable 1.
Evidence that
HSV-l(F)
ICP are thepri-mary products of translation: comparison of
HSV-1(F)
ICP labeled duringshortand long pulses and after a pulse chase. In these experiments replicate cultures of cells wereexposed to "C-amino acids at 4.5 and 6.5 h
postinfection forperiods of 10, 30, and 60 min.
Some of the additional cultures labeled for 10
min wereincubated for anadditional40min in
maintenance medium without labeled
precur-sors. The proteins oftheharvested, solubilized
cells were then subjected to electrophoresis in
parallel on an 8.5% acrylamide gel slab. Com-parisons of the autoradiograms (not shown)
on November 10, 2019 by guest
http://jvi.asm.org/
PROTEINS SPECIFIED BY HERPES SIMPLEX VIRUS
=
Cl -M -Ic
0 10 30 50 70 90
Distance
(m
m)
FIG. 4. Absorbance profiles of autoradiographic images of electrophoretically separated proteins from infected (toptracing)oruninfectedcells(bottomtracing) pulselabeled from 4.5to5.0hafter infectionormock infection. Theprofile ofthetotalelectrophoretically separated infectedcellproteins determinedby scanning the Coomassie brilliant blue stainedgel is alsoshown.Theproteinswereseparatedon 8.5%polyacrylamide gels. Numbersonupperprofileareof selected ICPtofacilitatecomparison withprofiles. The letter "h" identifies proteins in the infected cell which were synthesizedat rates comparable toproteins of similar mobilityin uninfected cells.
revealednosignificant differences inthe
mobil-ityorrelativequantity ofpolypeptides
accumu-lating in the cells during 10- to60-min pulses
administered at a given time postinfection.
Neither were there any significant differences
between the autoradiograms ofa 10-min
pulse
1355 VOL.12,1973
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.497.110.387.67.548.2]Total
Sup.
cell
flui
'(P
4.
5ji.
16
0
21
16
...
21
sm.;22
234
25
..
.......
2526
2728
29
30
31
32
...i
35
37
1
39
:
41
42
43
,44
46
I
mmune anda10-min
pulse
followedby
a40-min
chase. Asdiscussedbelow and shown in Fig. 6, similar d p pt. pulse and pulse-chase experimentsclearlydem-onstrated rapid posttranslational processing of
4
|(I
Ppolypeptides
inpoliovirus-infected
HEp-2
cells. §2' Moreover, a prolonged chase resulted in rela-tively minorchangesintheprofileofHSV-1(F)ICP (compare the 5.0- to 5.25-h pulse with the
5.0- to5.5-h pulse chased to 9.0 h; Fig. 9).
Failure tomodify the pattern of synthesis
of HSV-1(F) ICP by exposure to specific
6 inhibitors of trypsin (TLCK) and
chymo-6 trypsin (TPCK) proteases. Chloromethyl
ke-8 tone derivatives inhibiting proteolysis by
tryp-sin and
chymotrypsin
have been shown to1112 preventthe rapid posttranslationalcleavages of
16
poliovirus proteins inanumber ofcell lines (11,26) and to cause the accumulation of
"poly-protein" precursors. The effect of these
inhibi-19 tors on the synthesis of HSV-1(F) ICP was
21
therefore examined in conjunction withexperi-ments on poliovirus-infected HEp-2 cells as a
positive control system.
Replicate HEp-2 cellcultures were infected at
high multiplicityeither withHSV-1(F) or
polio-26
virustype 1, labeledwith "C-aminoacids for a15-mininterval beginning at 5.0 h postinfection,
29
either in the absence of inhibitors or in thepresence of 10' M TPCK or of TLCK, and
thereafter removed and processed for
electro-phoresis. An additional culture of cells infected
with poliovirus was pulse labeled for a similar
32
15-minperiod
in the absence of inhibitors andthereafter incubated for 40 min with medium
36 lacking labeled precursors. At the end of this chaseperiod these cellswere also processed for
37 electrophoresis. All samples were subjected to
8,
39
electrophoresis on the same (9.0% acrylamide)40
_lJ^-7 gel slab. The autoradiograms of these samples
40v (Fig. 6)showed the following. (i) Control
experi-ments with poliovirus-infected cells readily
demonstratedrapid
posttranslational
cleavages which were effectively blockedby inhibitors ofprotease activity. Thus, comparisonsoflysates
fromcellsgiven apulse and
pulse
chase(Fig. 6)showthat, in the absence ofinhibitors,
radioac-tivityrapidly disappeared frombands NCVP0
and NCVP11/2 andappearedinbands VP 0, VP
1 (VP 2), and VP 3. In thepresence of 10' M
FIG.5. Autoradiograms of8.5%polyacrylamide gels containing electrophoretically separated (i) total
in-fected cell lysatelabeled with 14C-amino acids from 4.25to6.5hpostinfection (total cell), (ii)the
superna-tantfluidobtainedafterhigh-speed sedimentation of
thelysate (sup.fluid), and (iii) animmune
precipi-tateformed by the addition of antiserumspecific for
virus antigens (immune ppt), to the supernatant fluid.Numbers to theleftof the three gels refer to ICP in the total celllysate, those to the right indicate the
ICP precipitated bythevirus-specificantiserum.
38
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.497.57.245.64.634.2]PROTEINSSPECIFIED BY HERPES SIMPLEX VIRUS
Polio-I
HSV-1
Pulse Pulse, Pulse Pulse Polio-.I Pus Ple Ple
proteins
TPCK
TLCK
Chase TLCK TPCK
IC P
NCVP 00
7
NCVP 0 L' 8
11149-12
-7~~~~~~~~51
-_NCVP
1I
:'
15-17;;;
FI~CVP
172
" ww 1921
_ _ _
~~~~NCVP
2 r-~2323_
NCVP3Ow:
~~~~~25-27
30 31
32
NCVP
X VPOVPI
-?P VP 2
VP3
4_IW
34-36_jW
38-39AMM 40-41
42-43
1*0v 44
45 46
FIG. 6. Autoradiogram ofa9.0%polyacrylamide slab gel containing electrophoreticallyseparated polypep-tidessynthesizedinHSV-1i(F)andpoliovirus type 1-infected HEp-2 cellslabeled with "4C-amino acids inthe presence and absence ofspecific inhibitors of trypsin (TLCK) and chymotrypsin (TPCK) proteases. Left, proteins synthesizedinpoliovirus-infected cells, pulsed for15min in the absence of inhibitors (Pulse), pulsed for15 minand chasedfor30 mininthe absenceofinhibitors (Pulse, Chase), andpulsed for15 min in the presenceof10-4MTLCK(Pulse TLCK) and TPCK (PulseTPCK). Samplesontherightareof HSV-1(F)ICP synthesizedinthe absence(Pulse)orpresenceofinhibitors(TPCK, TLCK).Thepoliovirustype1proteinsare designated accordingtoJacobson,Asso,and Baltimore(9).Thenumberstotheright of HSV-1i(F) samples refer to selectedHSV-1(F) ICP. Theoriginal autoradiogram was cut for thepreparation of this figure, and the
exposure interval employed for theprinting ofthe autoradiograms of HSV-1(F)-Pulse TPCK samplewas longerthan thatemployed for other samples.The molecularweightestimatesof poliovirus proteins agreed well with those of virion proteins subjected to electrophoresis inperipheral chambers of thesamegel slab but removedtosimplifythisfigure.
TLCK, thepoliovirus protein profilewas
modi-fied in a rather minor way (compare sample TLCK with the controlpulse), leadingto detec-tion ofa novel polypeptide slightly larger than NCVP 0 with a molecular weight of
approxi-mately 175,000andtoaslightreduction in label
in the regionof bandsNCVP 3a and NCVP 3b
in comparison to the untreated control.
How-ever, 10' M TPCK drastically reduced
incor-porationinto totalprotein,almostabolishedthe
appearance ofsecondary products, and led to
the accumulation ofpolypeptides of molecular
weight235,000(NCVP 00?) and160,000(NCVP
0). In general our results concur with those of
Summerset al. (26).
(ii) In contrast, at 5.0 h postinfection, no
rapid posttranslational cleavagesweredetected
in herpesvirus-infected HEp-2 cells. Thus, in
*::bqx
E._A%,
*bc ^
1357 VOL.12,1973
4110mo 11--Aww.'
low
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.497.114.394.72.432.2]samples infected with HSV-1 no difference either in the overallrateof protein synthesisor
theelectrophoretic profilesofpolypeptides
syn-thesizedwasdetectedbetween thecontrol
sam-ple (Fig. 6, pulse) and that labeled in the
presence of 10-4 MTLCK. Although overall
in-corporation was reduced in the culture labeled
in the presenceof10-4 M TPCK, there was no
clear difference between the mobilities ofICP
madeinthe presenceandabsenceofthe
inhibi-tor. With the exception of glycosylated ICP,
which were made in disproportionately lower
amounts, the remaining polypeptides were
la-beled inthe same relative proportions asthose
in the untreated control culture. More signifi-cantly, inviewof the
objectives
oftheseexperi-ments, therewas noaccumulation ofany poly-peptide ofhigher molecular weight than those detected during the control pulse. We exclude the possibility that a polyprotein too large to
enter the separation gel was produced under
these conditions since the radioactivity at the
interface between 3.5% "stacker" gel and 9.0%
"separation" gelwas aslow, relativeto radioac-tivity within the9.0%gel, asthatof the control
gel. Moreover, the exclusion limitof 9.0%gels is
> 335,000, basedontheentry ofthethyroglobin
polypeptide into this gel strength.
Changes
inthesynthesis of ICP during thevirus growth cycle. Three series of
experi-ments were done with the object ofexamining
changes
in overall proteinsynthesis and inthe synthesis ofindividual groups ofICPthrough-outthevirus growthcycle. Inthe firstseries we
measured the overall rate ofprotein synthesis
by labeling
cells for 2-h intervalsthroughout the growth cycle ofthevirus. The incorporationof"C-amino acids into acid-insoluble material
per 2 x 108 cellsper 2hinonesuch experiment
is shown in Fig. 7, along with the titers ofthe
infectious virus accumulated intracellularly
throughout
the same experiment. The overallrate of synthesis first declined rather rapidly, reachingaminimumat 2 to 3hpostinfection, it
then recoveredto a maximum ratecomparable
tothat ofmock-infected control cultures at 5 to
6 h, and thereafter declined in a nonuniform manner.
In the preceding experiments and in those
which follow, ICP were labeled with leucine,
isoleucine,andvaline.Quantitative estimations
ofthe relative rateoramountofsynthesisofICP
could therefore be in error iftheir content of
these three amino acids was not uniform. To
examine this
possibility,
in thesecond series ofexperiments two sets of
replicate
cultures ofcells infected with
HSV-1(F)
werelabeledwithmixtures of
'4C-leucine,
isoleucine,
and valine=
1-
4-Z._
l4
12
In
10
o
x 8 .C
E
4
.0
.s,
12
'Time
(hrs)
FIG. 7. Accumulation of infectious virus (A) and changes in therateof amino acidincorporation into acid-insoluble material in infected (0) and mock-infected cells (0). Amino acid incorporation was measuredasacid-insoluble radioactivity at the end of 2-h labeling intervalsduring infection or mock infec-tion and plotted at the midpoint of the interval. Pointsforinfectedcultures are the mean of duplicate determinations (2 x 106 cells perdetermination) and thosefor mock-infected cells are results of a single determination. Accumulation of infectious virus was measuredby plaque titrations of duplicate samplesof homogenatescontaining 2 x10 infected cells for each timeinterval.
and with a mixture of thirteen
"C-labeled
amino acids (reconstituted protein
hydroly-zate), respectively. Labeling medium in
experi-ments with
leucine,
isoleucine,
and valine wasas described in Materials and
Methods;
the medium used for labeling with the "C-protein hydrolysate was EMEM with one-tenth thenormal concentrations of amino acids supple-mented with 1% dialyzed calfserum. Cultures
were labeled for 15-min intervals
beginning
at7.25 and at 22.25 h postinfection. The labeled
samples were subjected to
electrophoresis
in parallelon apolyacrylamide gel
slab.Compari-son ofabsorbance tracingsofthe derived
auto-radiograms of the 7.25-h (Fig. 8) and of the 22.25-h (notshown)
labeling
intervalsindicated that, although the population ofpolypeptides
synthesizedat the twotime periodswas
mark-edly different, at neither time was there any
significant difference between estimates ofthe
relative amounts of ICP synthesized based on
incorporation ofleucine, isoleucine, and valine
or of the mixture of thirteen labeled amino
acids.
It should be emphasized that these
findings
do not indicate that the amino acid
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.497.257.447.61.257.2]PROTEINSSPECIFIEDBY HERPES SIMPLEX VIRUS
tions ofICP are uniform. In the same
experi-mentthereweresignificant differencesbetween the profiles of ICP labeled with the thirteen
amino acids and those labeled with single
amino acids (35S-methionine or "C-tyrosine).
However, in the context of this paper, the
pertinent observation is that
HSV-1(F)
ICP donot differ significantly in the sum of their
contents of the three amino acids which we
routinely employedaslabeled precursors.
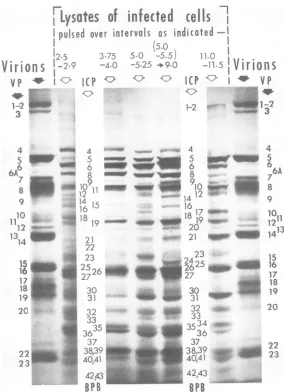
In the third series of experiments, replicate culturesofinfected cells were labeled from 2.5
to2.9, 3.75 to 4.0, 5.0 to 5.25,and11.0 to 11.5h
postinfection, respectively. These times were
chosenonthebasis of Fig.7andothertemporal features ofthevirus growth
cycle.
The intervalfrom 2.5 to 2.9h coincided with the end ofthe
initial decline in protein synthesis and before viral DNA synthesis is normally detected (14, 16). The next two intervals were
just
before (3.75to4.0h) andat (5.0to5.25h) the
timeof maximal overall proteinsynthesis
andhigh
rates ofviral DNA synthesis. The last interval
(11.0to 11.5 h) was at atime atwhich rates of
protein and viralDNA
syntheses
weredeclining
Distance [arbitrary units]
FIG. 8. Absorbance tracings of the autoradio-graphic images of polyacrylamide gelslabscontaining electrophoretically separated polypeptides from HSV-1(F)-infectedcells labeled with "4C-leucine,
iso-leucine,and valine(top panel)andwithamixtureof 13 "4C-amino acids (bottom panel) for a 15-min
interval from 7.25 h post-infection.
and the intracellular rate of accumulation of
infectious virus was near maximal.
Figure9showsanautoradiogram of a polyac-rylamide gel slab on which these samples were
subjected to electrophoresis. To facilitate the
presentation and interpretation of the data
available from this figure it is convenient to
express the amount of each ICP synthesized
during the various intervals as a "relative molar
amount synthesized," or a "relative molar rate
ofsynthesis" (RM). This datum was obtained
as follows: the autoradiograms of the gel slab
(Fig. 9) were scanned withthe aid of a Gilford spectrophotometer connected to a GA 16/45
computer. The latterdisplayed the absorbance
tracing, the absorbance ofeach band
(A,.
An)
and of the sum of the bands(A,
... An)*Since electrophoresis on more than one gel
strength wasnecessary toresolve certain of the 49ICP, the absorbanceofthosebands known to
containmorethan onepolypeptidewas divided
by
the number of known components, on theformalassumption that theband was composed ofequal masses of its knownconstituents. The
relativemolar amount,RM, of protein i
synthe-sized was then obtainedfrom the relation:
RMi=
107Kt.
At
Mi
(ZAi
..An)
where
Kt
istheoverallrateofprotein synthesisatthetime ofthe determinationexpressed as a
fraction of thehighestobservedrate,
Ai
andMt
arethe absorbanceofband i andits molecular weight, respectively, and107 is a constant.
The RM
profiles
of thepolypeptides
detectedintheautoradiogramsofFig.9 areshowninFig.
10.
Figure11shows themannerinwhich theRM values of synthesis of polypeptides vary
throughout infection.
The results
presented
in Fig. 9 to 11 may be summarized as follows. (i)During
any onepulse-labeling
interval,the RM formost S,NS,
and U polypeptides were within an eightfold
range; S and NS polypeptidesweredistributed
throughout this range. The most significant exceptions were the RM values ofthe S
poly-peptides, ICP1-2andICP 31, which differed
by
130-fold during the 11.0- to 11.5-h labeling
interval (Fig. 10) and formed the extremes of theobservedrange of values.
(ii) The variation in the relative proportions
ofS, NS, and U classes of
polypeptides
synthe-sized throughout the interval of the growth
cycle analyzed in this
study
was less than thevariation between polypeptides within the S
grouping. Thus, of thetotal amount ofprotein
VOL.12,1973 1359
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.497.55.241.335.590.2]HONESSAND ROIZMAN
Lysates
of infected cells
7
ouised
over
intervals
as
indicated
-I
i5.0
Vi
r
i
o n
s
V
i
r
i
o n s
Vp r
p
C P I V P1-2
1l2
3
Q
.... 1
14 l
1
^Q
...
'a!g_- 21
j22
. 23
2
3
23~~~~~~3
15 256
_
2625
16l17 1
19 ^ * 31 l 31
-32 20
3
3534
st
3635
*356
6423 j
42343
BPB BPB
FIG. 9. Autoradiogram of a part of a 7.75%polyacrylamide gel slab containing electrophoretically separated proteins pulse labeled in HSV-1(F)-infected cells during intervals shown on top of each sample. The legend on top of the third gel from the right indicates that cells were labeledfrom 5.0 to 5.5 h and then incubated in medium lacking labeled precursors until 9 hpostinfection. Peripheral samples are virion proteins, identified by largenumbers to the extreme right and left of the figure. The autoradiogram was cut in the preparation of this figure to permit convenient location of numbering for the ICP (smaller numbers between sample positions 2 and 3 and 5 and 6 of thisfigure).
synthesized, the S polypeptide fraction
(ex-pressed in moles percent) increasedonly
two-fold fromearly(2.5to2.9h)tolate(11.0to11.5 h) times in infection, whereas duringthesame
intervalthe molar ratios of SpolypeptideICP 5
to that of S polypeptide ICP 4 increased more than 30-fold.
(iii) Analysis of the kinetics ofsynthesis of
ICP (Fig. 11) showed sixpatterns ofsynthesis,
designated by the letters A toF. The
segrega-tion of ICP among these six patterns was
independentof the maximum rate ofsynthesis
of the polypeptides. However, the distribution
of S, NS, and U polypeptides among the six
groups was nonrandom. All but one of the
polypeptides presently included in group Aare
S. The exception, ICP 17, may bea long-lived
precursor since it disappeared on prolonged
chase (compare 5.0-to 5.25-hpulse and5.0- to
5.5-h pulse chased to 9.0 h shown in Fig. 9). Group C contained S polypeptides only. The S
ICP included in the analysis presented in Fig. 11 had an aggregate molecularweight of 2.3 x 106, and this S protein molecular weight was
4
5
6
7
8 9
11 ,
z
14
_mw _"w 8
_9
_w10
4 12 16
r::: .V
18 17
-19
:,~ _4 19 .;iw_ 20
21 M
4
5 6
6A 7 8 9
1°11
1211
1413
1360 J. VIROL.
fE% n
on November 10, 2019 by guest
http://jvi.asm.org/
[image:14.497.107.391.69.461.2]PROTEINS SPECIFIED BY HERPES SIMPLEXVIRUS
2.5-2.9hr 2.7 32 7
23
4 i I
0~~~~~2
3.75
-4.0Ohr389
6 ~~~~~~~~2627 6
8 32
19 23
20 129 137~
25hr
~~~~~38-31
26
6~~~~~~~~~~3
I 1371
1~~~~~30i1
10~~~I~33
II i2I9
I
110-1155hr
26
21 32
4 S~~~~~~~~~~~83
36
1719 3
2
1L..,:1i
~~~~~J
250 ISO
10-
SO - 60 40Molecular weight X
10-3
FIG. 10. Relative molar amounts of in} polypeptides synthesized duringshort labe
distributed in groups A, C, B, F, E, and D in
descending
order in amounts of 47, 31,8.1, 8.0,4.2, and1.6%, respectively.
(iv) The rates of synthesis of polypeptides
0-43
comprising groupsA, B, C, and D showeitheragradual increase (A), or an initial increase followed bya levelingoff(group B) or adecline
(groups C and D). As we noted earlier in the
text,
the initial stimulation in their rate ofsynthesis
isadditionalevidence thatthesepoly-peptides
are virusspecific.
However,
polypep-tides of group E could be "very early"
virus-specific products or host polypeptides with
gradually diminishing rates of synthesis. It is
noteworthythat several polypeptides which do notfulfill the generalcriterion ofinduction, and
which are not precipitated by the antiserum
(e.g.,ICP34, 35 andICP42, 43;see Fig. 4 and
Table 1, columns 5and6), fall into groupE.
Patterns
F of Fig. 11 were probably conse-quences ofsuperimpositions
ofthe patterns ofsynthesis
oftwoormorepolypeptides,
ofwhichL.
one or morebelongedtoclass Eand the otherstoclass
A, B, C,
orD. This artifactualpattern
wasobserved for
polypeptides
migrating
ingels
of a
particular
strength inmultiple,
incom-pletelyresolved bands. In the electropherogram illustrating this analysis (Fig. 9, 7.75% gel) the
groupsofICP34 to 36and40 to 43maybe cited
asexamplesof
poorly
resolved specieswhich,
in40-43 the absenceofother
data,
would beincludedincategory
F.Analyses
of the same and similarsamples
on othergel
strengths
(7 and 9%)..
permitted
classification
ofICP34,35,42,and43asgroup Epolypeptides (Table 1, column 5).
L
. DISCUSSIONAutoradiographic analyses
ofelectrophoreti-cally
separated
polypeptides
inpolyacrylamide
gels
revealed that HEp-2 cells infected with HSV-1(F) makeat least49polypeptides,
num-bered in order of decreasing size from 1 to49.
Some evidence forvirus
specificity
isavailable for47ofthesepolypeptides
and consists ofoneor more of the following: (i)
synthesis
of thepolypeptide
isstimulated
after infection,(ii)
the amount and in some instances the
electro-ffi phoreticmobilityare
determined
by thevirusin40-43 that these parameters vary
depending
on thevirus type and intratype variant infecting the
vals computed from data shown in Fig. 9 by the
lli
procedure
described in the text. The relative molar il amountsof polypeptides (RM, see text for methodof 1.racomputation) classified
asS,
U, and NS arerepre-36
D sented by "solid"lines, "dotted" lines, and "dashed" lines,respectively.Thepolypeptidesareidentified byfected cell ICP number (above bars) and themolecular weight
?ling inter- estimate
(abscissa).
VOL. 12, 1973
1361
-W 4D "d
4A
4D
z
21-%A
4D
7m Z. i
-LI',
10
.a..
a
0-
1--4D
E
6..cli
-2
40
E
IU
Z
4D 09
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:15.497.71.231.54.640.2]0 2 4 6 8 10 0 2 4 6 8 10 0 2 4 6 8 10 Time (hrs)
FIG. 11. Patternsof synthesis of ICPinHSV-1(F)-infected cells. The ICPweregrouped bythepattern of synthesis designatedAtoF. SincetheRMfor the ICPcomprisingeachgroupare notthesame, theplotswere
normalized. However, themaximum RMobservedduringthereproductive cycle is shownunder column Min the boxedlegendnext to thenumber andsymbolicrepresentation of the ICP.
cell, and (iii) the polypeptides are precipitable by anti-HSV-1 antisera of known specificity.
The remaining two polypeptides (ICP 34 and
35) are probably host polypeptides, a
con-clusion reinforcedbytheobservation that these polypeptides comigrate with major proteins of
the uninfected cell.
These 47 virus-specific polypeptides havean
aggregate molecular weight corresponding to
that of about 41,000 amino acids. Assuming
that the distribution of "sense" information in
the HSV-1 DNA is asymmetricassuggestedby
analyses of transcription (6), HSV DNA can
specify the sequence ofa maximum of 55,000
amino acids. On the basisofthesecalculations
the47polypeptidesaccountforabout75% ofthe
maximum information contentofHSV-1 DNA.
Of the 27 known virion polypeptides, 24
"ana-logues," i.e., polypeptides with identical
elec-trophoretic mobilities, were provisionally
iden-tified among the47 ICP. Of the remainder, 15
did not correspond to any known virion poly-peptide and 9 remained unclassified, although
it islikely that most are nonstructural.
Polypep-tides specified by the remaining 25% of HSV
genetic information areeithermadeinamounts
too small to be detected or are obscured by
major polypeptides. Although the resolution
andidentification ofpolypeptides is incomplete
and their classification is provisional pending
furtherstudies, severalaspectsof the synthesis
of ICP are apparent from results of this paper
and meritdiscussion.
Posttranslational modification of viral polypeptides. At least three kinds of
posttrans-lational modifications may alter the mobility
ofviruspolypeptides, i.e., (i) rapid
posttransla-tional cleavages such as those seen in
picor-navirus-infectedcells (9, 25, 26), (ii) slow
post-translational cleavages determined by intracel-lular translocation or occurring during assembly
ofthevirion, and (iii) conjugation or addition of
prosthetic groups, e.g., glycosylation,
phospho-rylation, amidation, acetylation, methylation, etc. Studies presented in this paper have
ex-cluded rapid posttranslational cleavages of the
type seen in picornavirus-infected cells.
How-ever, a prolonged chase revealed modifications
inthemobilityofsomepolypeptides, notablyof
ICP 17and of a component ofICP 23 (Fig. 9).
The magnitude ofthese
changes
suggest thatICP 17 is cleaved to asyet unknown products.
Thenatureofthe conversion ofacomponent of
ICP23 (to ICP24) is uncertain. Ingeneral, the
operation of slow modificationsrequiresfurther
study in view of the considerable number of HSV polypeptides which are substrates for
glycosylation (18,22) and
phosphorylation,
andparticularlysince wehavenotidentifiedprotein
22A ofB-capsids,theputative precursor toVP
on November 10, 2019 by guest
http://jvi.asm.org/
[image:16.497.112.394.61.300.2]PROTEINS SPECIFIEDBYHERPES SIMPLEXVIRUS
22 (8; W. Gibson and B.
Roizman,
J.Virol.,
submitted
forpublication)
inourpresent analy-sis.Kinetics of synthesis of structural and nonstructural polypeptides.
Analysis
of the kinetics ofsynthesis ofICP revealed six basic patternsdesignatedAthroughF.Of these six, Fisprobably artifactual, aconsequence of multi-ple comigrating polypeptides synthesized
ac-cordingtopattern E and
B,
C orD. Pattern E could describe equally well host proteins with progressivelydecliningratesofsynthesis
aswellas virus proteins synthesized very
early
ininfection. Most of virus-specific polypeptides follow pattems A
through
D,
but not in arandom fashion. Thus, structural proteins and theirlikelyprecursorsalmost all followpatterns A and C, whereas NS polypeptides
clearly
identified by two or more criteria as virus-specific follow patterns B, D, and E. Several
commentsshould be made
regarding
patternsofsynthesis ofthese
polypeptides
and theirfunc-tion. Briefly, (i) it is convenient to regard S
polypeptides comprisinggroupC and the virus-specific NS polypeptides comprising groups E
as early.
Conversely,
the Spolypeptides
com-prising group A arelate, whereas the NS poly-peptides ofgroups Band D
might
bedesignated
stable and
unstable, reflecting
their rates of synthesis late in infection. (ii)Comparison
ofthe rates of synthesis of the S polypeptides at
varioustimeintervalswiththeamountsofthese
polypeptides
in the virion show thatthey
are notsynthesizedinproportiontotheir incorpora-tion intovirions. As seen fromdatapresentedinFig. 9, the probability of incorporation is not
uniform and appears to be much
higher
formembersofgroup A
(late)
polypeptides
thanforgroup C (early)
polypeptides.
Theinterpreta-tion of this observation is
presently
uncertain;wemayspeculate that
early S polypeptides
mayhave functions unrelated to their role in the
virion in addition to
having
aggregation rateconstants
markedly
different from those ofgroup Apolypeptides.
Temporal and abundance control of viral protein synthesis. In parallel with studies on
the control oftranscription (6, 7, 21), we are
concerned with two aspects of viral protein synthesis, i.e., temporal control ofprotein
syn-thesis and control of viral protein abundance.
Theexistence ofatemporalcontrol ofprotein
synthesis is apparent from analyses of the
kinetics ofsynthesis of viral proteins
summa-rized in Fig. 11. As discussedearlierinthetext,
the distribution of S, NS, and U
polypeptides
among the different kinetic classes is
nonran-dom, with NSproteinsfollowingpatternsB, D,
and
E,
and Sproteins largely following patternsA and C. Because S polypeptides are
synthe-sized both early (pattern C) and late (pattern
A), there is no clear temporal differentiation
between S and NS proteins.The incorporation
of amino acids into S polypeptides early is therefore appreciable; S polypeptides incorpo-rated 53, 67, and 72% of total amino acids
during the labeling intervals beginning at 2.5,
3.75, and 11.0 h postinfection,respectively. We
conclude from the foregoing analyses that (i)
temporal controls ofHSV-1 gene expression do
exist, (ii) these controls definethesynthesis of atleast fivegroupsofproteins, and (iii)thereis no simple differentiation of proteins intoearly nonstructural and late structural. Structural
proteins are members of both early and late
groups and form a substantial fraction of total
proteins madethroughout infection.
The search for abundance control ofprotein
synthesis arises fromtheobservation that viral
transcripts form two classes differing in their accumulated mean molar abundance. The
abundant class is transcribed from 14 to 16%
and 19 to 22% of virus DNA at 2 and 8 h
postinfection, respectively, and is from 140- to atleast 15-foldmore
abundant
than the scarce transcripts which arise from about 30% of theDNA bothearly (2 h) and late (8 h) in infection. Abundant andscarcetranscripts arisingfroma
total of 43% of virus DNA were found on
polyribosomes at 8 h postinfection, and it was
found that abundant transcripts were adenyl-ated whereas scarce transcripts were not (21).
These observations raise two questions: (i) is
therea control ofprotein abundanceand (ii) is
protein abundance determined solely by the
transcript concentration or by theoperation of
translational controls.
It seems clear that a protein abundance
control does exist since proteinsare not synthe-sized in
equimolar
amounts. This is evident from acursory examinationofFig. 10and from the fact that the molar ratio of protein 31 toproteins 1 to 2 is 130 at 11 hpostinfection, even
though both proteins follow the same general
patternofsynthesis.
The answer to the second question is at
pres-entuncertainfor two reasons.First,it isdifficult
to comparetheresults of analysesof
transcrip-tional and translatranscrip-tional programs. The first
type ofanalysisisfundamentallymoresensitive
inthat thehybridizationofscarcetranscriptsis
notobscuredbythat ofabundant transcriptsin
the way that trace amounts of oneprotein may
beobscured byproteins in the same size range
present in much largeramounts. However, the
abundant class of transcript has not been
VOL.12,1973 1363