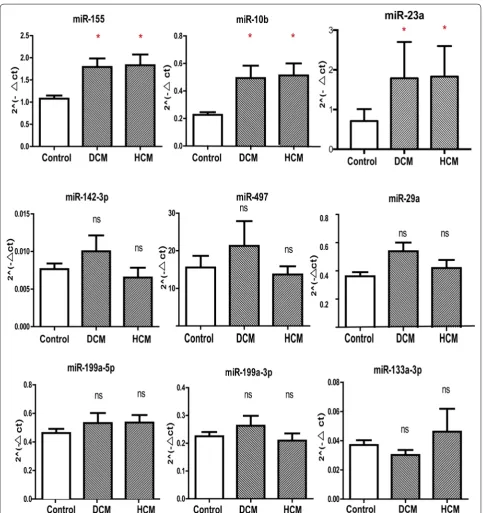
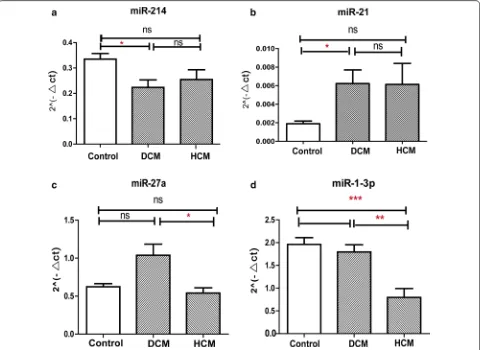
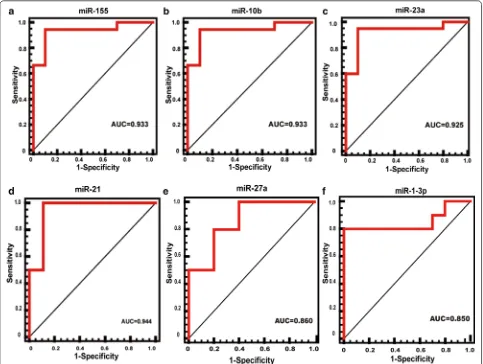
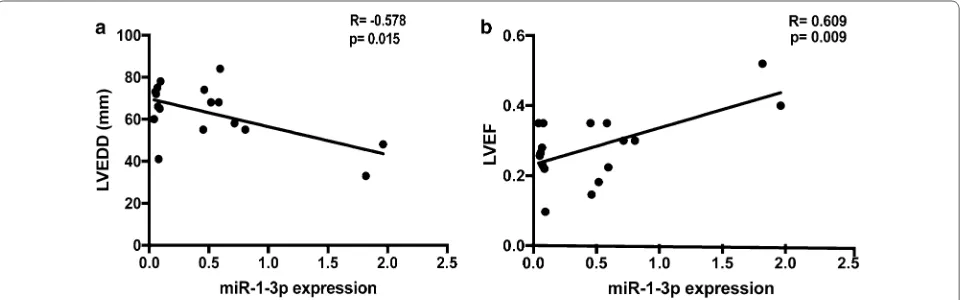
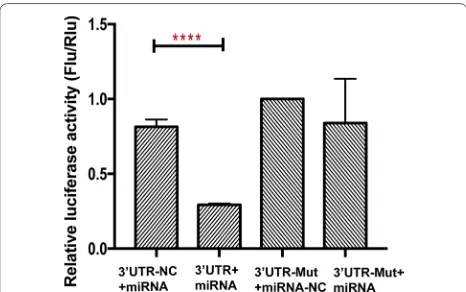
MiR 1 3p that correlates with left ventricular function of HCM can serve as a potential target and differentiate HCM from DCM
Full text
Figure
Related documents
Without describing this new form at length, it is here that I wish to comment on three statements in Rabkin's paper: "Member States submit to rules and regulations drawn
The curriculum- integration project was premised ‘on the conviction that there is presently limited critical awareness of HIV and AIDS as well as human rights-related matters
Survival analysis according to the treatment for the recurrent disease showed that local therapy (repeat hepatectomy or ablation) for intrahepatic recurrence improved OS compared
• What impact does invasive home mechanical ventila- tion have on the quality of life of caregivers/parents of the affected patients.. • Are there differences in the quality of life
– Emerging channels like online chat, text messaging, and smartphone apps now used by nearly 1/3 of all policy holders. I l il i d
When blastn-searched against the nt database in GenBank, many sequences in Table 1 and other taxonomically uncertain OTUs from this study returned
The lender will ask the judge for a Possession Order - the right to take your property, evict you and anyone living there, and to sell the property to recover the mortgage and
