0022-538X/85/020634-11$02.00/0
Copyright X) 1985, American Society for Microbiology
Localization of Epitopes of Herpes Simplex Virus Type
1Glycoprotein
D
ROSELYN J.
EISENBERG,',2*
DEBORAH LONG,2'3 MANUEL PONCE DE LEON,2'3 JAMES T. MATTHEWS,1'2'3t PATRICIA G. SPEAR,4 MARYLOU G. GIBSON,4 LAURENCE A. LASKY,5 PHILLIP BERMAN,5 ELLISGOLUB,2'6
ANDGARYH.COHEN2'3
Department of Pathobiology, School of Veterinary Medicine,1 and Department of Microbiology,3 Department of
Biochemistry,6 andCenterfor Oral Health Research,2 School ofDentalMedicine, University of Pennsylvania,
Philadelphia, Pennsylvania 19104; Department of Microbiology, The University of Chicago, Chicago, Illinois606374; and
Departmentof Vaccine Development, Genentech, Inc., South SanFrancisco, California940805
Received 20August 1984/Accepted 23 October1984
Wepreviously defined eight groupsof monoclonalantibodies which reactwithdistinctepitopes of herpes
simplex virusglycoprotein D (gD). One of these,groupVIIantibody,wasshownto reactwithatype-common
continuous epitope within residues 11 to 19 of the mature glycoprotein (residues 36to 44of the predicted
sequenceofgD). In thecurrentinvestigation,wehavelocalized the sitesofbinding oftwoadditional antibody groups which recognize continuous epitopes of gD. The use of truncated forms of gD aswell ascomputer
predictions of secondarystructureandhydrophilicitywereinstrumental in locatingtheseepitopes andchoosing
syntheticpeptidestomimictheirreactivity. Group II antibodies, whicharetypecommon,reactwithanepitope
within residues 268to287of thematureglycoprotein (residues 293to312 of the predicted sequence). Group
Vantibodies, whicharegD-l specific,reactwithanepitope within residues 340to356 ofthematureprotein
(residues365to381 of the predicted sequence). Fouradditional groupsofmonoclonalantibodiesappear to
reactwithdiscontinuousepitopes of gD-1, since the reactivity of these antibodieswaslost when theglycoprotein
wasdenatured byreduction and alkylation. Truncatedforms of gDwereusedtolocalize these four epitopesto
the first 260 amino acids of the mature protein. Competition experiments were used toassess the relative
positions of binding of various pairs of monoclonal antibodies.Inseveralcases,whenoneantibodywasbound,
therewas nointerference with the binding ofanantibody from anothergroup,indicating that the epitopeswere
distinct. However, in other cases, there was competition, indicating that these epitopes might share some
commonamino acids.
Glycoprotein D (gD) of herpes simplex virus (HSV) is a
structural component ofthe virion envelope which
stimu-lates production of high titers of virus-neutralizing activity (7, 9, 11, 15-17, 34) and islikelytoplayanimportant rolein
the initialstagesof viralinfection. It wasrecently shown that
anti-gD antibodiescanblock the fusion ofinfected cells(39). Inaddition, mice immunized with gDare protectedfroma
lethalHSV challenge(4, 30, 34, 40).Tryptic peptide analysis
(2, 16) and amino-terminal sequencing(14) showed that gD
of HSVtype 2(HSV-2) (gD-2) is structurally similar though
notidenticaltogDof HSV-1(gD-1). Recently,the genesfor gD-1 and gD-2werelocalized and sequenced (29, 32, 50, 51). Although the deduced amino acid sequences for the two
glycoproteins were shown to be 85% homologous, little is
knownaboutthesecondaryortertiarystructureof thesetwo
proteins. Using monoclonal antibodies (MCAb), we previ-ously defined eight epitopes within gD (15), some of which
aretype common and others of which are typespecific. Our
goalis to relate the structureoftheproteinto itsbiological
functions. The high degree of amino acid sequence
homol-ogy between gD-1 and gD-2 probably accounts for the
immunologicalcross-reactivity of polyclonal antibodiesand
MCAb directed againstgD (15, 41). On the otherhand, the
type specificity of other MCAb is undoubtedly related to
differences in the structures of gD-1 and gD-2 and,
conse-quently, inamino acid sequence.
*Correspondingauthor.
tPresentaddress: DepartmentofPathology, HarvardUniversity School ofMedicine, Boston, MA02115.
Recently (7, 12),using synthetic peptides, the type
com-mongroup VIIepitopewaslocalizedtoresidues 11to19of
the mature form of gD (residues 36 to 44 ofthe predicted
sequence [29, 50, 51]). Polyclonal sera to certain of the
synthetic peptidesin the region ofthefirst23amino acids of gD-1 and gD-2alsoexhibited type common virus-neutraliz-ing activity (7), and immunized micewere
protected
from alethalviruschallenge(R. J. Eisenberg, G.H. Cohen, andB.
Dietzschold, unpublished data). During these
studies,
welocalized twoadditional epitopes within the first 23 amino acids of gD-2(7, 12). Thetype 2specificity dependedon two
amino acid differences between gD-1 and gD-2 (29, 50, 51).
The purpose ofthecurrent
investigation
was tocontinue
todelineatethelocationandcharacteristics of theantigenic
epitopes
of gD. Threedifferentsetsoftermshave beenusedto
distinguish epitopes
which are lost underdenaturing
conditions, such as reduction and alkylation, from thosewhich are retained:(i) conformationalversussequential (47);
(ii) discontinuousversuscontinuous (1); and
(iii)
assembledtopographic versus segmental (3). We have chosen to use
discontinuous and continuous as operational
definitions,
without any implication that this terminology is preferredoverany other. We havelocalizedthesite ofbindingoftwo
additional MCAb groups. As with group VII, these also recognize continuous epitopes. Groups II and V (15) bind
specifically to residues 268 to 287 ofgD-1 and gD-2 and
residues 340 to 356 of gD-1. In addition, we carried out
competition studies to map the relative positions offour
discontinuousepitopes, correspondingtoMCAb ingroups
I,
III, IV,and VI (15).
634
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
Cells,
virus,
and radioactivelabeling procedures.
Condi-tionsfor the
growth
and maintenance of BHK and KB cellsandforthe
propagation
of virushavebeen described(8, 11).
For
infection,
aninput
multiplicity
of20 PFUofHSV-1(HF)
and 10 PFU of HSV-2
(Savage)
per cell was used.Proce-duresfor
labeling
ofHSV-infected cells with[35S]methionine
(specific
activity,
600Ci/mmol) (Amersham
Corp.)
and[2,3-3H]arginine (specific
activity,
15Ci/mmol)
(Amersham)
havebeen described
previously
(9, 13, 15, 16).
Preparation
ofpolyclonal antibody
and MCAb togD.
Anti-gD-1
andanti-gD-2
serawereprepared
in rabbitsagainst
immunosorbant-purified
preparations
ofgD-1
andgD-2 (17).
MCAb HD-1
(group
I)
and MCAb 170(group VII)
weresupplied
by
L. Pereira(15, 41).
MCAb55S
and 57S(group
V), 11S
(group
III),
41S(group IV),
and45S(group VI)
weresupplied
by
M.Zweig
(48).
MCAb DL6(group II)
wasprepared
from mice immunizedintraperitoneally
(34)
with 6 ,ugofimmunosorbant-purified
gD-1
(17).
Threedays
afteranintravenous boost of1 jig, the
spleen
was removed and thecellswerefusedto
SP2/0
cellsby
theprocedure
ofMcKearn(28).
Hybridomas
were cloned in soft agarose(25),
andascites fluids were
prepared
fromPristane-primed
miceimmunized
intraperitoneally
with cloned cells.To prepare
immunosorbants,
immunoglobulin
G(IgG)
waspurified
from ascites fluids of MCAb fromeach group(15)
andlinkedtocyanogenbromide-activatedSepharose
4B(Pharmacia
FineChemicals, Inc.).
The amountscoupled
ranged
from 5to 12mg ofIgG
per g ofSepharose.
Purifiedimmunoglobulins
(50
Rxg)
were iodinated with 1251(Amer-sham)
by
eitherthe chloramine-T(19)
orthelactoperoxidase
(36)
method. For certainMCAb,
thechloramine-T methodinactivated the
binding
activity
oftheantibody.
Group
IV MCAb were inactivatedby
bothprocedures.
Synthetic peptides.
Synthetic peptides
toresidues 1to23ofgD-1
(1-23[1])
andgD-2
(1-23[2])
wereprepared
asdescribedpreviously
(7).
Thepeptides
representing
residues340to356of
gD-1
(340-356[1])
and 268to287ofgD-1 (268-287[1])
wereprepared
by
PeninsulaLaboratories,
Inc.Cysteine
wasadded to the amino terminus of
340-356[1]
and to thecarboxy
terminus of268-287[1].
Theprocedures
forcoupling
ofpeptides
tokeyhole
limpet
hemocyanin
(KLH)
weredescribed
previously
(7,
33).
Briefly,
themaleimidegroupofthe
peptide
wasincorporated
into KLH withM-malimido-benzol-N-hydroxysuccinimide
esterandtheM-malimidobe-nzol-N-hydroxysuccinimide
ester-modifiedproteins
wereal-lowed to reactwith a 20 Mexcess ofthe
peptide
(22).
Thecoupling
ratio ofpeptide/carrier
waspreviously
determined to be 8(7).
Allpeptides
were dissolved in 0.1 M Tris(pH
7.8)-0.15 M NaClforassay
by
theimmunoblotmethod(see
below).
Preparation
of antisera tothesynthetic peptides.
AfemaleNew Zealand White rabbit was immunized with
peptide
340-356[1]
coupled
to KLHatthreeweekly
intervals withatotalof2.4mgof
coupled peptide.
The animalwasgiven
anintravenous boosterdose of120
p.g
ofthecoupled peptide
3 to4days
before eachbleeding.
Atotal of five bleeds wereobtained. Two female New Zealand White rabbits were
immunized with
peptide
268-287[1]
coupled
toKLHatthreeweekly
intervals withatotalof1.4 mgofcoupled peptide
perrabbit.Theanimalswereboosted
intravenously
oncewith 70 ,ugofcoupled peptide
and then three times with 500 ,ug of freepeptide.
Preparation
of native and denaturedgD.
gD-1
andgD-2
were eachpurified
fromcytoplasmic
extracts of infectedcells by affinity chromatography, using a previously
de-scribed procedure (17). For our purposes, the
proteins
eluted from the immunosorbant column with KSCN and
dialyzed against 0.01 M Tris (pH 7.5)-0.15 M NaCI-0.1%
Nonidet P-40 (TSN buffer) are designatedas "native." For
denaturation, purified gD-1 orgD-2 was suspended in
dis-rupting bufferto yield a final concentration of 3% sodium
dodecyl sulfate (SDS)-100 mM Tris (pH 7.0)-10%
2-mercaptoethanol-0.5% glycerol. The sample was boiled for
5 min. lodoacetamide (0.1 M in 0.1 M Tris, pH 8.0) was
added to give a final concentration of 33 mMiodoacetamide,
and the mixture was incubated for 1 hat roomtemperature.
The samples were dialyzed extensively against TSN buffer.
Preparationof truncated forms of gD. (i) The 38Kfragment
ofgD-1. Preparation of the 38K fragment wasby a
modifi-cation of a previously described procedure (15). Briefly, a
cytoplasmic extract(100 ,ul) of HSV-1-infected cells labeled
with [3H]arginine was added to 50
RI
ofHD-1-IgG-Se-pharose (125 ,ug of IgG). The immunosorbant was washed
extensively with 0.01 M Tris (pH 7.5)-0.1% Nonidet P-40-0.5
M NaCl-0.1 mM phenylmethylsulfonyl fluoride (washing
buffer)and then with V8 enzyme buffer (50 mM Tris,pH8.0)
and incubatedwith 50 p.g ofStaphylococcusaureusprotease
V8 in enzymebufferat37°C for 2 h. The immunosorbantwas
washedextensively withwashing buffer. To test the
prepa-ration for the presence of uncleaved gD, a portion of the
immunosorbant was suspended in SDS-disrupting buffer,
boiled for 3 min, and analyzed by SDS-polyacrylamide
gel
electrophoresis (PAGE). V8 proteolysis resulted in a 38K
fragment (15) and no full-length gD. The 38K fragment
linked to HD-1-IgG-Sepharose was then used to test the
bindingof other MCAb asdescribed in Results.
(ii) TruncatedgD-1, residues 1 to 275.The gene forgD-1
wascloned intoapBR322-simianvirus40 shuttle vector and
included a DNA fragment from a HindlIl site upstream of
thegD gene to a Hinfl site atresidue 300 of the predicted
gD-1 sequence (residue 275 of the mature protein). When
this plasmid is grown in Chinese hamster ovary cells, the
glycoproteinis secreted(30). The glycoproteinwaspurified by affinity chromatography, usingagD-specificMCAb(17).
(iii)
Truncated gD-1, residues 1 to 287. Truncated gD,residues 1 to 287, was producedand secreted by the HSV
insertion mutant designated 111 (M. G. Gibson and P. G.
Spear, J. Cell Biochem. Suppl. 8B, in press; Gibson and
Spear, 13th Ann. UCLA Symp. 1984, abstr. no. 1337, p.
191).
The virus was constructed by methods previouslydescribed(18)exceptthatatruncated form of thegD-1 gene
(extendingfrom theSaclsite upstream of thegD gene to the
NarI siteatresidue 312of thepredictedsequenceorresidue
287 of thematureprotein)wasinserted into theBglIIsite of
the
thymidine
kinase gene. The expressed proteincontains48 amino acids at its carboxy terminus that are translated
from the noncodingstrand of HSV-1 DNA atthe 5' end of
thethymidine kinase gene. Theproteinwasaffinity purified
from the medium ofHEp-2cellsinfected with 111 virusbya
previously
described method (17), using a gD-1-specific MCAbdesignated
11-436-1 (39).Immunoprecipitation
andSDS-PAGE. gDwasimmunopre-cipitated
from HSV-1-orHSV-2-infected cellextracts(cellsinfected for 6 h) prepared as previouslydescribed (26, 42),
using
antiseraorMCAb and S. aureusproteinA(IgG Sorb; NewEngland Enzyme
Center). SDS-PAGEwascarriedoutin slabs of 10%
acrylamide
cross-linked with 0.4%N,N'-diallyltartardiamide
(13, 49).Forautoradiography, gelsweredried on filter paper and
placed
in contact with KodakXAR-5 film. For
fluorography,
the gels were treated withon November 10, 2019 by guest
http://jvi.asm.org/
Amplify (Amersham), dried on filterpaper, andexposed to
Kodak XAR-5 film at -70°C.
Immunoblot and neutralization assays. The immunoblot
assay was done as previously described (7, 20), using
antisera or MCAb and iodinated protein A (Amersham).
Virus neutralization assays (50%plaque reduction method),
using HSV-1 (HF) or HSV-2 (Savage), were carried out as
previously described (9, 11).
Competition assays. Twocompetition assays were used.
(i)Immunosorbants. Atotal of 100to 200,ul(representing
150 to 600 ,ug of IgG) of MCAb linked to Sepharose was
incubated for 2 h at37°C with 100,ulofacytoplasmic extract
of unlabeled HSV-1-infected cells (9, 13, 15, 16). The
im-munosorbant was washed 10 times with washing buffer, and
the iodinated second antibody (at least 250,000 cpm) was
added. Thecomplex was washed exhaustively andcounted
in a gammacounter.
(ii)Nitrocellulose. PurifiedgD-1(0.45 to 15 ng) was spotted
onto nitrocellulose strips; the strips were washed as
previ-ously described (7) and incubated with unlabeled first
anti-body and then with iodinated second antibody (ca. 250,000
cpm). The spots were located by autoradiography, cut out,
and countedin a gammacounter. The counts were subjected
to linearregressionanalysis, and the slope of the line (counts
per minute versus concentration of gD) was calculated.
RESULTS
Comparison of the predicted secondary structures of gD-1
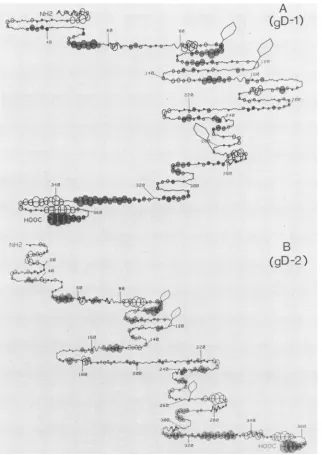
and gD-2. Figure 1 shows a computer representation (7) of
thesecondary structure ofgD-1(Fig.1A)and gD-2(Fig.1B),
derivedfromthepredictedamino acidsequences (29,50, 51)
and rules established by Chouand Fasman (5, 6). It should
be noted that these predictive "rules" of secondary
struc-ture result inan accuracyof predictionin a three-state model
(helix-sheet-turn)ofapproximately50versus33%forchance
(24). However, in the absence of any additional structural
information, we have found that these predictions have
heuristicvalue in that they focus attention on certain regions
of the glycoprotein. Furthermore, we have also analyzed
both glycoproteins with a second empirical analysis (Fig. 1)
which assumes that hydrophilic regions of protein structure
have a greater immunological potential (22). For these
calculations, the first 25 amino acids of the predicted
se-quence were excluded from consideration, since direct
N-terminal sequence analysis showed that, for both gD-1 and
gD-2, lysine residue 26 of the deduced sequence was the
aminoterminus of the mature protein (14). In ournumbering
system, this lysineisresidue 1. The criteria used for
predict-ing the probability of
a-turns
(7) were modified to increasethe likelihood of locating possible epitopes. The
modified-turn criteria predict four additional
n-turns
in gD-1 atresi-dues 200, 225, 255, and 298. None of these additional turns
involves highly hydrophilic regions of the protein.
Interest-ingly, avery hydrophilic region, residues 77 to 95, is not in
a predicted
a-turn.
When the criteria were relaxed evenfurther, this stretch ofamino acids was still not predicted to
be in a
p-turn.
The working hypothesis is that epitopes arelikely to be located in regions where highly hydrophilic
residues are present in ,-turns (45). If, in addition, the
epitope is continuous, synthetic peptides could be used to
mimic the reactivity of the epitope. The program has also
beenexpanded to indicate the positions of predicted
N-aspar-agine-linkedcarbohydrates (shown as balloons) based on the
sequence Asn-X-Thr or Asn-X-Ser (23). For gD-1 and gD-2,
all three positions are glycosylated (10). Predictions for
hydrophilicity use the same criteria as before (7, 22). The
homologyin aminoacid sequence is reflected in similarities
in both secondary structure andregions ofhydrophilicity in
the two proteins. In at least one case, however,two
differ-ences in amino acid sequence in region 1 to 23 have been
correlated with bothchanges in predicted secondary
struc-ture andantigenicity (7, 12, 29, 50, 51). For bothgD-1 and
gD-2, there are two regions in which ,-turns intersect a
highly hydrophilic region, i.e., residues 11 to 19and 265 to
282. A third region in gD-1, residues 340 to 356, is
hydro-philic and contains a predicted ,-turn overlapping the
hy-drophilic region.Thesea-turnsarepresentevenwhenmore
stringent criteria for
predicting
turns areapplied
(7).IngD-2,
however, there is no 3-turn in this region, even with themodified criteria.
Reaction of MCAb in groups I to VII against native and
denaturedgD. To test the structural
predictions,
weused theimmunoblot assay(7, 20) todetermine which of thegroups
of MCAb reacted with discontinuous or continuous
epi-topes. Previous studies indicated that
only
certain MCAbgroups reactedwith denatured gD (32, 37;J. T.
Matthews,
G. H.Cohen,and R. J. Eisenberg, unpublished
data).
NativegD-1(Fig.2)reacted withpolyclonal
anti-gD
serumand withMCAbin groups I to VII(rows 1 to8, lanea). Native
gD-2
(lane b) reacted withpolyclonal anti-gD (row 1,lane
b)
andalsoreactedwithMCAbin groupsI,II, III,
V,
and VII(rows
2, 3, 4, 6, and8, lane b). The reaction of nativegD-2
withgroup V wasunexpected, since group V failedto immuno-precipitategD-2from infectedcell extracts(15,48).
Further-more, denatured gD-2 reacted either
weakly
or not at allagainstgroup VMCAb, whereas native anddenatured
gD-1
reactedequallywellagainstthe sameantibodies. Inaddition
to group V, MCAb in groups II and VII
recognized
thedenaturedformofgD-1andgD-2(lanescand
d).
Thus,
threegroups of MCAb, II, V, and VII,
appeared
torecognize
continuous
epitopes,
andfourgroupsofMCAb,
I,III, IV,
andVI,apparentlyreactedwith discontinuous
epitopes
thatrequirethenative conformation of
gD-i
(34).It should be noted that under the
denaturing
conditions used considerablesecondary
andtertiary
structuremight
remain ingD. Therefore, thattheprotein
retainedantigenic
activity for antibodies ingroupsII, V,
and VII is notproof
per se that these
epitopes
are continuous. In the case ofgroup VII, the proof was
provided by
thereactivity
ofasynthetic
peptide
mimicking
residues8to23 ofgD-1
against
group VII MCAb 170 (7). One ofthe
goals
ofthe presentstudy was toobtain similar
proof
fortheepitopes
specified
by MCAb ingroups II and V.Prediction ofthe location of the groupV epitope. Several
linesof evidence enabled ustolocalizethegroup V
epitope.
Previously,
using
V8proteolysis,
we found that group VMCAb reacted with a 15K
fragment
ofgD-1
(15).
Tryptic
peptide
analysis
showed that thisfragment
represented
thecarboxyterminus of the
protein
(15;D.Long,
G. H.Cohen,
and R. J. Eisenberg,
unpublished
data).
Further evidenceindicated that theepitopewaslocateddownstream fromthe
membrane-anchoring
region
(i.e.,
presumably
after residue 339[51]). First, group V MCAb reacted withfixed,
but not withunfixed, HSV-1-infected cells (37). Thissuggested
that theepitope isnotexposedonthe externalface of theplasma
membraneof infected cells. Second,when
gD-1
wassynthe-sized and processed in an in vitro system, the
processed
protein was partially protected fromproteolysis by
trypsin
(37). Approximately 3,000 daltons of theprotein
wasre-moved bythis treatment, and the
trypsin-resistant
fragment
could not be
immunoprecipitated by
group V MCAb.Fur-thermore, when truncatedformsofthe
gD
gene,lacking
theon November 10, 2019 by guest
http://jvi.asm.org/
NH2 A
2
gnx
~~~(gD-1)
,w-,q-8-ia v
HOOC~~ ~ ~ ~ ~ ~ ~ H
(gD-2)
*-but4(1
Wit,e=t4;$r~ ,^JA5~ . 4*ea
FIG. 1. Predicted secondary structure and hydrophilicity maps ofgD-1 (A) and gD-2 (B). Secondary structures were predicted by a
computerprogram,usingthe rules of Chou and Fasman fordetermining Pt, Pa,and Pb(5,6). Probabilities for theoccurrenceofa-turnswere evaluatedby usingmodified conditions: Pt>7.5 X 10-'orPt>5 x 1O-5 pt>Paand Pt> Pb.Shaded circles indicatehydrophobic regions;
opencircles indicatehydrophilicareas. Theradiusofacircleoveraresidue isproportionaltothe meanhydrophilicityascalculatedfor that residueplusthenextfive residuesaccordingtothemethod ofHoppand Woods(22). Thevalue is therefore distortedatthe C-terminal end.
Thehexagonalballoons indicatepredictedsites (Asn-X-ThrorSer)of
N-asparagine-Iinked
glycosylation (23).information for the transmembrane-anchoring region plus thecarboxyterminus, were cloned into Escherichiacoli, the
expressed gD-like protein was not recognized by 57S
anti-body (R. J. Watson, J. H. Weis, J. H. Salstrom, and L. W.
Enquist,J. Invest. Dermatol., in press). These results, taken together, suggested that the group V epitope was located
between residues 340 and 369 of gD-1.
Tolocalize theepitope further, we relied on the computer
predictions (Fig. 1and 3)and differences in the sequences of gD-1 and gD-2 at the carboxy terminus (Fig. 3) (29, 50, 51).
Weargued that the epitope is largely type 1 specific, based
on theimmunoprecipitation data (15), but that certain
simi-larities in sequence might account for the reactivity of gD-2
againstgroupVMCAbin the immunoblot. Region 340 to 356
of gD-1 ishighly hydrophilicandcontainsapredicted j-turn
atresidues 346to349(Fig. 1and 3). The homologous region
of gD-2 is also hydrophilic, but does not contain this
predictedturn. In addition, the sequences of gD-1 and gD-2
in the region 340 to 356 (Fig. 3) show similarities (e.g.,
residues 346to349 areAla-Pro-Lys-Arg and residues 351 to
356 are Arg-Leu-Pro-His-Ileu-Arg in both proteins) and
differences (e.g., residues 343 to 345 are Thr-Arg-Lys in
gD-1 butareAla-Gln-Met ingD-2). These differences appear
tohaveaprofound effectonthe predictedsecondary
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.148.470.74.529.2]N
ct
J
Xa
0
-0
3
I
I
10
.X.. .v.'V.
0
-7
340-356[1] contains the group V epitope; (ii) this
epitope
appears to be immunogenic in the native protein since
several polyclonal seraprepared against gD, including one
against gD-2, reacted with the peptide; and (iii) there is
limitedantigeniccross-reactivity withananalogousregionof
gD-2.
Anti-340-356[1] serum reacted in the immunoblot assay
(Fig. 5A) against nativegD-1 (lane a), gD-2(laneb), andthe
syntheticpeptide 340-356[1](lane h). The specificity ofthis
serumfor thecarboxyterminus of gD isdemonstrated by the
lack ofreactivity against truncated gD-1, residues 1 to 275
(lane e), and the synthetic peptides 1-23[1] and 268-287[1]
(lanes f and g, respectively). The reactions against native gD-1 and gD-2 were stronger than those
against
thedena-A 3338 340
N H
2
M.H.,9.^
".R.
,F.
3e0 K
[iV' 355 350
365 ,~
[image:5.612.73.287.72.472.2]~~~~
~
0 0
H
FIG. 2.Immunoblot analysis of monoclonal antibodies directedat
HSVgD. The antibodies used (groupedasdescribed in reference15)
were asfollows: row1,anti-gD-1 (rabbit 1); row2,groupI, HD-1; row3,groupII, DL6;row4,groupIII, 11S;row5,groupIV,41S; row6,groupV,57S;row7,groupVI,45S;row8,groupVII,170. Antigens: lanea,immunosorbant-purified (17) (native) gD-1 (15ng); laneb, native gD-2 (15 ng); lane c,denatured gD-1 (15ng);laned, denaturedgD-2 (15 ng); lanee,truncated gD-1, residues 1to275 (30) (60 ng).
tureof thedownstream residues of thetwoproteins. Based
onthisinformation,residues340to356of gD-1 appeared the
mostlikelyto contain thegroup V epitope.
Recognition of the synthetic peptide 340-356[1] bygroup V
MCAb andby polyclonal anti-gDsera.GroupVMCAb (Fig.
4, row 7) reacted with purified native gD-1 (lane a), gD-2
(lane b),and thesynthetic peptide correspondingtoresidues
340to356 ofgD-1 (lane e).This antibodydidnot reactwith
synthetic peptides corresponding to other portions of gD-1
(lanes c andd). Two polyclonal seraprepared against gD-1
(rows 1and2) reactedwiththis peptide. Inone case(row 1)
thereaction was strong, and in the other case the reaction
was weak(row 2). In addition, the peptide reacted weakly
with one polyclonal serum prepared against purified gD-2
(row 3), but failed to react with a second anti-gD-2 serum
(row 4). These results show that (i) the synthetic peptide
B 340
NH2
V350
355 360
i: 'i | b R
l...
R
.. .345 P
365
HOOC
.Y~' L"
FIG. 3. Comparison of amino acid sequence and predicted
secondary structures ofcarboxy-terminalsequences of gD-1
(resi-dues 338to369)andgD-2(residues 338to368). Probabilities for the
occurrenceofP-turnswereevaluatedasinthelegendtoFig. 1. The single-lettercode designations are asfollows: A, alanine; R,
argi-nine; D, aspartic; N,asparagine; C, cysteine; E, glutamic acid; Q, glutamine; G, glycine; H, histidine; I, isoleucine; L, leucine; K,
lysine; M, methionine; F, phenylalanine; P, proline; S, serine; T, threonine; W, tryptophan; Y, tyrosine; V, valine.
I
2
Adik
qw
A
*
4
4
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.318.557.240.648.2]a
cde
A
abcdef gh
* *
*
S
B
S
a
b c d e f
5.5,i
6
ee-7
S
-,Y ._ ..__
FIG.4. Immunoblot analysis ofsynthetic peptides which mimic portions of gD-1, using polyclonal antibodies and MCAb. The antibodies usedwere asfollows: row1,anti-gD-1 (rabbit 1);row2, anti-gD-1 (rabbit 2); row3, anti-gD-2 (rabbit 3); row4, anti-gD-2 (rabbit 4);row5, group VII,170;row6, groupII,DL6;row7, group V, 57S.Antigens: lane a, native gD-1 (15 ng); lane b, native gD-2 (15 ng); lane c, 8-23[1] (500 ng); lane d, 268-287[1] (1 p.g); lane e,
340-356[1] (100ng).
tured forms oftheseproteins (lanescandd).Anti-340-356[1]
serumimmunoprecipitated both gD-1 (Fig. SB, lane c) and
gD-2 (lane d) from infected cellextracts. Lanesaand b(Fig.
SB) represent negative controls in which the HSV-1 and
HSV-2 extracts were tested with serumobtained from the
animal before immunization with
340-356[1].
As positive controls, the same extracts were immunoprecipitated withanti-gD-1serum(laneseandf).Itis clear thatanti-340-356[1]
wasmorereactiveagainst
gD-1
than gD-2. Aswithgroup VMCAb,theanti-340-356[1]serumfailedtoshow any
neutral-izing activity against HSV-1orHSV-2 (data notshown).
Localization of the groupIIepitope. Group II,represented
byDL6MCAb, reacted withthe native anddenaturedforms
of both gD-1 and gD-2 (Fig. 2, row 3), exhibited
type-commonmembrane immunofluorescence (datanotshown), and neutralized HSV-1 at 1:50 dilution and HSV-2at 1:20,
usinga50% endpoint(9, 11). The continuous epitope
recog-nizedbyDL6MCAbwasdistinct fromthoserecognized by
either group VIIorV, sinceDL6didnot reactwith either the
8-23[1]
or the 340-356[1] synthetic peptide (Fig. 4, row 6,lanes c and e). Preliminary localization of the group II
epitope was accomplished by testing the reactivity oftwo
[image:6.612.64.298.74.406.2]truncatedforms ofgD-1, representingresidues 1 to275and
FIG. 5. Analysis of anti-340-356[1] serum by immunoblot (A) and SDS-PAGE (B). For immunoblot analysis, the antigens were: a, native gD-1; b, native gD-2; c, denatured gD-1; d, denaturedgD-2; e, truncated gD-1, residues 1 to 275; f, 8-23[1]; g, 268-287[1]; h,
340-356[1].Theconcentrations of theseantigenswerethesameasin
Fig. 2. For SDS-PAGE analysis (B), extracts from HSV-1-infected cells are inlanes a, c, and e and extracts fromHSV-2-infectedcells are in lanes b, d, and f. The sera used were: lanes a and b, preimmunization bleed from rabbit immunized with 340-356[1]; lanes c andd,anti-340-356[1];lanes e and f, anti-gD-1 (17).
1 to 287, against MCAb (Fig. 6). Both forms reacted with
group VII antibody (row 1, lanes c to e), indicating the
presence of residues 8-23 in the truncated proteins. As
expected, neitherform reactedwithgroup V antibody(row
3, lanesc to e). The truncated form, 1-275[1], also failedto
a
bcd e
I
2
*
*S
.0
FIG. 6. Immunoblot analysis of truncated forms of gD-1, using MCAb.Theantibodies usedwereasfollows: row1, group VII,170; row2, groupII,DL6;row3, groupV,57S.Antigens: lane a, native gD-1 (15 ng); lane b, native gD-2 (15 ng); lane c, truncated gD-1, residues1 to275(30),60ng;laned,truncatedgD-1,residues 1 to 287(GibsonandSpear, in press), 100ngofprotein;lanee, residues 1 to287,200ngofprotein.
I
2
3.*
4.-m60K
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.315.559.76.312.2] [image:6.612.319.559.502.664.2]react with group II MCAb (row 2, lane c). However, this
antibody didreactwith 1-287[1] (row 2, lanes d and e). These
results suggest that thegroup IIepitope is probably located
between residues 275 and 287, although it could include
several residuesupstream. Fromcomputerpredictions (Fig.
1),theregion from residue 266toapproximately 282
repre-sents a potential epitope in both gD-1 and gD-2 since this
region is highlyhydrophilic and contains a,B-turn (45).This
combination of predictions and data suggest that a peptide
consisting of residues 268to287 wouldencompassthegroup
IIepitope.
Localization of the group II epitope by using synthetic
peptides. Toconfirm thelocation of thegroupIIepitope,we
tested the reactivity of a peptide representing residues
268-287[1]. This peptide reacted withgroupIIMCAb (Fig. 4,
row6,lane d) butnotwithantibodiesingroupV(row7,lane
d) or VII (row 5, lane d). Thus, the group II epitope is
located between residues 268 and 287 of gD-1. In addition,
thepeptide reacted against three of the fourpolyclonalsera
prepared against gD (Fig. 4, rows 1 to3, lane d).
Peptide 268-287[1] was coupled to KLH and used to
immunize tworabbits. After the initial immunization
proto-col andoneintravenous booster dose, serum samples were
assayedfor anti-gD and anti-peptide antibodies by the
im-munoblotassay. Neitherserumreacted with nativeor
dena-tured gD (data not shown). The animals were then given
three intravenous booster doses of the free peptide and
successive serumsampleswere assayed again. Again, none
of theserareacted with native ordenatured gD. One serum
sample reacted with the synthetic peptide 268-287[1] (data
not shown). None of the sera exhibited any neutralizing
activity (data not shown).
Localization of discontinuous epitopes of gD-1.Previously,
we developed a procedure (15) to partially fragment gD,
using S. aureus protease V8, an enzyme which cleaves
specifically at glutamic acid residues. In that procedure,
metabolically labeled gD was immunoprecipitated with a
MCAb plus S. aureus-bearing protein A and the complex
wastreated with the protease (15). Theantibody protected
thatportion of gDtowhich itwasbound, and the bound and
unbound fragments were characterized by SDS-PAGE and
tryptic peptide analysis. We found thatgroupI, IV, and VI
MCAb remained bound to a 38K fragment which, on the
basis ofN-terminal amino acid sequencing (14; D. Long,
G. H. Cohen, R. Hogue-Angeletti, and R. J. Eisenberg,
unpublisheddata), wasfound tocontaintheamino terminus
of pgD (the precursor form ofgD). Analysis of the tryptic
glycopeptides (10) of the 38K fragment showed that
glyco-peptides 1 and 2 were present but that glycopeptide 3 (at
position 262)was missing (data not
$hown).
Thus the 38Kfragment appearstobe located betweenresidues 1 and 262.
A possible V8 cleavage site is located at glutamic acid
residue 260. The V8 experiments implied that the epitopes
reacting withgroup I, IV, VI, and VII MCAbwerelocated
within this portion of gD-1. Antibodies in groups II and III
couldnotbelocalized by this technique, since nofragments
remained associated with them after V8 proteolysis.
How-ever, we nowknow thatgroup II islocated downstream of
the38K fragment. Thus, it wasalsopossible that thegroup
IIIepitopewaslocated downstream of thecarboxy terminus
of the38K.
Figure 2(lane e) shows thatgroupI, III,IV, VI, and VII
MCAb boundto anothertruncated form of gD-1 consisting
ofresidues 1to275(30). These resultsagreewiththoseofV8
proteolysis (15) with the exception ofgroup III(11S). One
[image:7.612.316.556.89.148.2]possibility was that thegroup III epitope included residues
TABLE 1. Localization of the 11S epitope on the 38K fragment 1251 bound(cpm)b
Sample" HD-1_251
iiS_125i
57S_125I
Control 9,459 10,205 57,433
gD-1-HD-1 (no V8) 7,868 690,120 350,784 gD-1-HD-1 (+ V8) 10,898 589,136 54,826 a The controlsampleconsistedofHD-1-Sepharose with no gD-1 added.
Foreach assay, 100p.1of acytoplasmic extract ofHSV-1-infected cells was added to 50 ,u1 of HD-1-IgG-Sepharose. The immunosorbant was washed with washing buffer and incubated with 50 ,ug of S. aureus V8 protease in 50 mM Tris (pH 8.0) for 2 h at 37°C. The complex was washed with washing buffer andthenincubated with iodinated MCAb. This complex was washed andcounted in a gamma counter.
between 260 and 275. Anotherpossibility is thatbinding of
group III occurred within the 38K fragment, but that this
binding didnotprotect thefragment
frorn
furtherproteolysis
(15). Todetermine directly whether group III MCAb could
bind to the 38K fragment, we carried out the following
experiment. A group I MCAb (HD-1) immunosorbant was
used to bind purified gD-1. This complex was washed
extensively, and a portion was treated with S. aureus
protease V8 and washed again. Then various iodinated
MCAb were added, and the complex was washed
exten-sively and then counted inagamma counter. Table1 shows that, in the HD-1 control
(HD-1
as immunosorbant andiodinated probe), no significant counts bound above
back-groundineithertheV8-treatedor theuntreated sample. We also probed the V8-treated and untreated samples with
group Vantibody(57S)andfound that theuntreatedsample
containedasignificantnumberofcountsandtheV8-treated sample
contained
no counts above background. Thisindi-catedthat the
proteolysis
wascomplete andthat thegroup Vepitopewasnotpresent afterV8 treatment. Withiodinated
11S(groupIIIMCAb)as theprobe, approximately thesame
number ofcounts bound to the V8-treated and untreated
gD-1-HD-1 complexes. These results show that the group IIIepitope ispresent onthe38Kfragmentand,furthermore,
that thisepitope isdistinctfromthe group I epitope.
Topographical relationship
ofepitopes
located in residues 1to 287 of gD-1. Previous
experiments
indicated that sixepitopes ofgD-1 werelocated withinthefirst 287residuesof
theprotein,twoofwhich werecontinuousand fourofwhich werediscontinuous. Twodifferent experimentswerecarried
out tobegintodefinetherelativepositions oftheseepitopes.
These will be referred to as competition experiments,
al-though we recognize that the term does not accurately describethetypeofanalysis being
performed.
First, representative MCAb from the six groups were
covalently bound to Sepharose. A preliminary
experiment
was carried out todetermine the amount of each
immuno-sorbantrequired to bind agiven amountof gD-1 present in
infected cell extracts. The appropriate amount of each
immunosorbant was used tobind similar amountsof
unlab-eled gD-1. After this, a different and iodinated second
antibodywasadded,and thecomplexeswerewashed
exten-sively and counted in a gamma counter. The underlined
values in Table 2show that each antibody groupcompeted
against itself, sinceonly background levels of counts bound
when the same antibody was used as immunosorbant and
iodinatedprobe.Threetypesofresultswereobtained: (i)no
competition, in which a significant number ofcounts were
bound, e.g., usinggroup III as immunosorbant and group I asprobe, orvice versa; (ii) completecompetition, in which
only a background number of counts were bound, e.g.,
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 2. Competition analysis usingMCAblinkedtoSepharose 4B'
lodinatedantibodies (cpm of1251bound)
Immuno-sorbant Group I Group II GroupIII Group VI Group VII (HD-1) (DL6) (llS) (45S) (170)
Control 3,166 2,227 710 7,800 1,066
I(HD-1) 2,914 62,135 34,158 15,806 37,909
II(DL6) NDb 3,175 ND ND ND
III(11S) 30,619 80,231 700 2,958 10,972
IV(41S) ND ND 578 32,265 ND
VI(45S) 17,558 85,565 3,304 9,819 30,581 VII(170) 27,618 80,048 30,581 25,435 1,027
aAntibodiesareincluded in groups accordingto originaldefinitions (15). Foreachassay,100 to 200p.lof MCAb linkedtoSepharosewasincubatedfor 2h with100p.lofacytoplasmicextractofHSV-1-infected cells (9,13,15,16).
The immunosorbant was washed and the iodinated second antibody (ca. 250,000 cpm) was added. Thewashed complexeswerecountedin a gamma counter. The underlined values show that eachantibody group competed against itself.
bND, Not done.
comparinggroup IV against group III; (iii) partial competi-tion, in which some counts were bound, e.g., comparing
group VI and groupI. Inthisassay, thereisnoway toknow
what the maximal level of binding should be.
As asecond approach (Table 3), different concentrations of purified gD-1 were spotted onto nitrocellulose strips
which were then incubated with an excess of unlabeled
antibody. The strips were washed and incubated with
iodi-nated second antibody. The maximal level ofbinding (no
competitionor0%in Table 3) wasdetermined fromacontrol inwhichthestrip was incubatedonly with labeledantibody.
The values underlined in Table 3 represent the percent
competition which occurredwhen the same antibody, both
unlabeled and labeled, was used to compete against itself.
Theoretically,these valueshouldapproach 100%. Forgroup
II, this value was 93%; however, forgroups I and III, the
value was
70%.
Thereasonfor this isnot understood,since,in each case, the firstantibody was presumably present in
excess. Itmay be a problem of antibody affinity or
presen-tation ofthe antigen on nitrocellulose. Nevertheless, when
heterologous antibody groups were compared, the results
agreedwith the resultsinTable 2. There was nocompetition
between groups I and III or I and VII. Furthermore, there
was partial competitionbetween groups I and IV and I and
VI. Group III showed partial competition with groups IV
and VI. Group II MCAb showed slight
competition
withgroup VI and possibly group I. Thus, the two kinds of
experiments leadtothesameconclusions and form thebasis
fortopographical positioning of these epitopesingD-1 (Fig.
7).
DISCUSSION
Inprevious studies, wedefinedeight antigenic epitopes of
gD, based on an analysis with a panel of MCAb (15). We
attempted toassociate the binding ofparticular MCAb with
differentfragments of the protein and found that several of
themwere ina38Kfragment whichencompassesthe amino
terminus. We further defined the position ofonecontinuous
epitope of gD, amino acid residues 11 to 19, which reacts
with group VII MCAb (7, 12). Localization of that epitope
was based on the V8 proteolysis studies (15) as wellas the
use of computer predictions of secondary structure and
hydrophilicity in choosing an appropriate synthetic peptide
to test the predictions. Here, our goal was to localize the
precise locations oftwo other continuous epitopes,
recog-nizedby group II and V MCAb, andto begin to define the
location of discontinuous epitopes of gD-1. Computer
pre-dictions were instrumental in helping to choose synthetic
peptidestodemonstrate thelocation ofcontinuousepitopes.
In each case, the epitope was found to be located within
stretches ofhighlyhydrophilic amino acid residues making
uppredicted ,-turns (45).
The groupVepitope was localized to residues 340to356
ofgD-1 which isdownstreamfromthemembrane-anchoring
region (50, 51). A synthetic peptide consisting of this
se-quence was found to bind specifically to group V MCAb.
The location of this epitope is thus on the portion of gD-1
which faces the inside of the virion or infected cell and
confirms predictions of its location based on other studies
(15, 37; Watsonetal., J. Invest. Dermatol., inpress). Thus,
the failure ofgroupVMCAbtoneutralizevirusortobindto
thesurface of HSV-1-infected cells is due tothe
inaccessi-bility of thisepitope when gDisassociatedwithmembrane.
Recently, Rector et al. (44) used a group V antibody (55S)
(15, 48) to examine whether non-neutralizing antibodies
could be protective in passive immunization studies. Their
data showed that 55S was not protective. Since group V
MCAb wouldnot have reacted with intact virions orintact
infected cells, their result is not surprising.
The results of the presentstudy indicate that thegroupV
epitope is in itself immunogenic in purified gD, since the
synthetic peptide reacted with several polyclonal sera
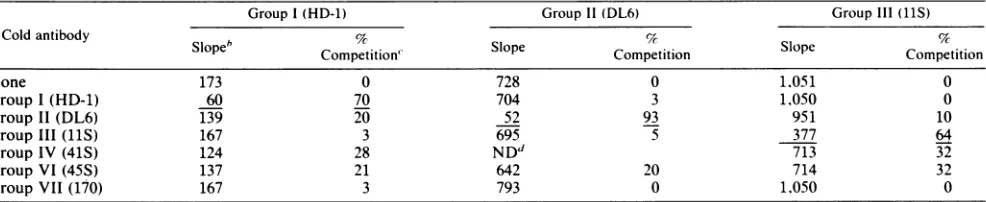
pre-TABLE 3. Competition analysis usingthe immunoblot assay
Group I (HD-1) Group11(DL6) GroupIII(11S)
Cold antibody Slopeb Competition" Competition Competition
None 173 0 728 0 1,051 0
GroupI(HD-1) 60 70 704 3 1,050 0
Group II(DL6) 139 20 52 93 951 10
Group III (11S) 167 3 695 5 377 64
Group IV(41S) 124 28 ND" 713 32
GroupVI(45S) 137 21 642 20 714 32
GroupVII (170) 167 3 793 0 1,050 0
"Ineach assay, various concentrations of gD-1, ranging from 0.45 to 15 ng, were spotted onto nitrocellulose, incubated with cold antibody for 2h,and then re-actedwith iodinated antibodies for1 h.The spots were cutfrom the nitrocellulose and counted in a gamma counter. The results were plotted as counts bound ver-susconcentration ofgD.
bSlopeisgivenin counts per minute per nanogramofgD.
cPercentcompetitionwasderived by the following equation: 100-(counts per minute pernanogram of gD bound with unlabeledantibody present/counts per minute per nanogram of gD bound with no unlabeled antibody). Values underlined represent the percent competition which occurred when the same antibody,
both,unlabeled andlabeled,was used to compete,against itself.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.66.559.570.671.2]NH2' Vil
11-19
. V
V
3340-356
A
OEl~
COOH [image:9.612.60.303.70.195.2]268-287
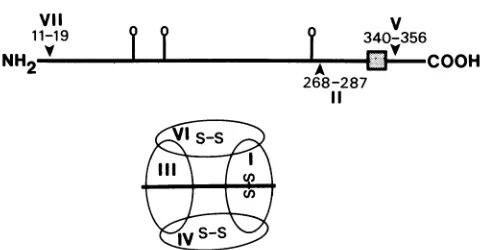
FIG. 7.Topographicmapof HSV-1 gD. Thepositionsofepitopes
bindingtoMCAbingroupsVII, I1,andVareshown. Alsoindicated
(as balloons)are the threeN-asparagine-linked glycosylationsites.
The transmembrane region is depicted as abox. The positions of discontinuous epitopes (ellipses at bottom)were derived fromthe competition experiments. Three of these epitopesappeartoinvolve S-S bonds.
pared against gD, including an antiserum prepared against
gD-2. More recently, we have tested additional polyclonal
sera prepared in rabbits against gD-1 and gD-2 and have
found that six ofeight sera, including two prepared against
gD-2, reacted with this peptide. In addition, antiserum
prepared to the peptide reacted with gD-1 and gD-2 in
immunoblot and immunoprecipitation assays, although the reaction was much stronger against gD-1. A somewhat
puzzling observation was that this antiserum was more
reactive againstthe native than the denaturedforms ofgD-1
and gD-2 (Fig. 5A, lanes a to d). It is possible that the
conformations ofpeptide 340-356[1] on KLH might well be nearertothe conformation of thesamesegmentinthenative
protein thantothe different conformationalensemble of the
denatured protein.
Anotherpuzzling observation was the reactivityofgroup
V MCAb against gD-2detectedbyimmunoblot. Previously,
these antibodies were considered type 1 specific based on
immunoprecipitation and immunofluorescence assays.
Al-though most of the amino acids in the epitope specified by
group V antibodies may be unique to gD-1, it is clear from examination of the sequence in this region that several
amino acids in the epitope must be common to gD-1 and
gD-2. The more sensitive immunoblot assay might better
detect thispartial overlap. Alternatively, gD-2 mightassume
different conformations under the conditions used in dif-ferentassays(27, 38). Thefewtypecommonresidues in this sequence maybearrangedclosetogetherinthe nativegD's, sothataweak typecommonreactivitycanbeseen,although the sequencesotherwise differ substantially. Ifso, it would
explain why group V MCAb appeared to be more reactive
against native than against denatured gD-2 (Fig. 2, row 6,
lanes b and d).Finemappingofthe group Vepitope should take intoaccount the differences in sequence betweengD-1
andgD-2in thisregion.
Localization of the group II epitope was accomplished
first by analyzingthe reactivity oftruncated forms ofgD-1
against DL6 MCAb. The antibody failed to react with a
truncated form ofgD ending at residue 275 of the mature
protein (30)but didreactwithaform ofgD endingatresidue
287(M.G. Gibson and P. G. Spear,inpress). These results
suggested that the epitope was between residues 275 and
287. Thecomputerpredictions (Fig. 1)showedthatgD-1and gD-2 each contained a region with hydrophilic residues within a P-turn. Interestingly, this region is rich in acidic
residues
(both
aspartic
andglutamic
acids)
andproline,
butlacks the basic amino
acids,
such aslysine,
that are oftenassociated with
epitopes (31).
Inchoosing
asynthetic
pep-tide for confirmation ofthe
location,
we assumed that theepitope
could include some residues upstreamof 275. Pep-tide268-287[1]
reacted with DL6 MCAb and with threepolyclonal
seraprepared against gD.
Thispeptide
alsoreacted with several other
polyclonal anti-gD
rabbitsera, atotal of four to
gD-1
and two togD-2.
However,
whenrabbitswereimmunized withthis
peptide coupled
toKLH,
there was no
antibody
response. After severalinjections
with the free
peptide,
one ofthe seraexhibited
reactivity
against
thepeptide
but notagainst gD.
In this case, it ispossible
that none of the conformations of thepeptide
coupled
to KLHcorresponded
to the structure of thepeptide
as it is foundinnative ordenaturedgD.
The
synthetic
peptide approach
wetook inthesestudies isnot yet
likely
to be fruitful inlocalizing
discontinuousepitopes
(1,45).
This is because theseepitopes depend
onacertain
tertiary
structure ofgD,
which in part involvesdisulfide bonds. The
position
of these bonds is not yetknown,
but such information should aid in localization.However,
we do know that fourdiscontinuousepitopes
ofgD-1
are located within the first 260 amino acids of theprotein
(Fig. 7),
since antibodiesingroupsI,
III,
IV,
and VIreacted with truncated
gD-1 (1
to275)
as well as with the38K
fragment
(presumably
residues 1 to260)
generated
by
V8proteolysis.
Six ofthe sevencysteine
residues ofgD-1
arelocated within residues66to202.Itis
quite
possible
that disulfide bonds formedby
these sixcysteines play
arole inthe structure of discontinuous
epitopes.
Cysteine
atresidue333ofthemature
protein
is withinthetransmembraneregion
of
gD-1
and is notinvolved in formation of theseepitopes.
Thus,
our datapredict
that thiscysteine
isprobably
notinvolved in intramolecular disulfide bonds in
gD-1.
How-ever, it may be involved in intermolecular
disulfides,
per-haps
information ofthegD-1
dimer(17, 18).
Inthisregard,
it is
interesting
to notethatgD-2,
which lacks thiscysteine,
does not form dimers
(17,
18).
Inpreliminary
experiments,
we have found that the group III
epitope
isdestroyed by
boiling gD-1
in SDS in the absence ofmercaptoethanol,
whereasdestruction ofgroups
I,
IV,
andVIrequired
reduc-tion and
alkylation (M.
Ponce deLeon,
G. H.Cohen,
andR. J.
Eisenberg,
unpublished data).
Competition
experiments
were carried out to determinetherelative orientationofthefourdiscontinuous
epitopes.
Insome cases, the
binding
of oneantibody
togD-1
had noeffect
(no
competition)
on thebinding
of another. Theseresults are further evidence that the MCAb
groupings
arevalid and that there are distinct discontinuous
epitopes
ongD-1.
In other cases, there wascompetition.
This indicatedthat
(i)
someamino acidsin theseepitopes
areshared;
or(ii)
theepitopes
were soclose that therewassteric hindrance in thebinding
of a secondantibody;
or(iii)
binding
of oneantibody
altered the conformation ofgD
so thatbinding
ofthe second
antibody
wasaffected(35).
Previously,
wespec-ulated that
binding
ofgroup III MCAb altered theconfor-mation of
gD-1,
making
the molecule moresusceptible
toprotease V8
cleavage (15).
Thisexplanation
is stillconsistentwith the present results.
On the basis of these
studies,
as well as the studies ofthree continuous
epitopes,
we have constructed a two-parttopographic
map forgD-1
(Fig.
7).
First,
we havedepicted
theprotein essentially
as alinear molecule with thepositions
ofthe three continuous
epitopes
indicated. Thediscontinu-ous
epitopes
have beendepicted
in a separatedrawing
ason November 10, 2019 by guest
http://jvi.asm.org/
ellipses
located downstream fromgroupVII,
each of whichincludesamino acids
prior
toresidue 260. Thediscontinuousepitopes corresponding
to MCAbwhich exhibitedcompeti-tion are shown as
overlapping.
Thus,
group IIIoverlaps
groups IV and VI and group I also
overlaps
IV and VI.Groups
I and III do notoverlap
at all.Preliminary
experi-ments indicate that antibodies in all of the MCAb groups
except
possibly
group IV areabletoimmunoprecipitate
thegD-like
protein produced by tunicamycin-treated,
HSV-1-infected cells
(42;
Matthewsetal.,
unpublished data).
Threeof the
epitopes
aredepicted
asinvolving
disulfidebonds(S-Sin
Fig.
7),
although
wedonotknowhowmanycysteines
aredisulfidebonded in
gD
orhow manyare involved indeter-mining
thestructure ofany oneepitope.
Furtherlocalization of discontinuous
epitopes
willrequire
other
approaches, including
amorecomplete
understanding
of thecontributionofdisulfide bondstothestructureof
gD.
One
possible
approach
will be toanalyze
the amino acidchanges
associated with mutants which exhibit an alteredpattern
ofreactivity
with MCAb. Such mutants wouldinclude those whichare no
longer
neutralizedby antibody,
suchasthe"mar"mutants
(21).
Anotherapproach
wouldbetoexaminethe amino acid
changes
foundin natural isolatesof HSV which exhibit an anomalous
pattern
ofreactivity
with MCAb
(41, 43).
Thisapproach
wasrecently exploited
(43)
toexplain
thereactivity
of an HSV-1 strain with agD-2-specific
MCAbcalled17f3A3(2).
Analysis
ofthe DNAsequence of the
gD
gene ofthe isolate revealed achange
which altered the codon for
asparagine
(residue
72 of the matureprotein)
present
in the prototype HSV-1 strain tohistidine,
normally
present
inthe HSV-2 strain.Grouping
of17,A3
hasnotyet
beenaccomplished.
However,
itmight
bein group VIII
(15).
Ouruncertainty
about thegrouping
of17,A3
illustrates the need for a common classification ofgD-specific
MCAb. Wearenowinthe process ofgrouping
anumberof additional
gD-specific
MCAb fromseverallabo-ratoriesto overcome this
difficulty.
ACKNOWLEDGMENTSThis
investigation
wassupported by
Public Health Service grantsDE-02623 from the National Institute of DentalResearch,AI-18289 from the National Institute of
Allergy
andInfectiousDiseases,
and CA-21776 from the NationalCancer Institute.Aportion
of this workwas
supported
by
agranttoG.H.C. and R.J.E. from the AmericanCyanamid
Co. M.G.G. is a fellow of the Leukemia Society ofAmerica,and J.T.M.was a
predoctoral
traineesupported byPublicHealthServicegrantNS-07180 from the National Institute of
Neu-rological
and Communicative Disorders and Stroke.We thank B.
Hampar,
M.Zweig,
and L. Pereira for monoclonalantibodies,
Wesley
Wilcox forcarefully reading
themanuscript,and Madeline Cohen,Valerie Rinaldt, andMichael Nobelfor excellent technicalassistance.LITERATURE CITED
1. Atassi,M. Z.1978.Precisedetermination oftheentireantigenic
structureof
lysozyme.
Molecular features ofprotein
antigenicstructures and
potential
of 'surface-stimulation'synthesis-a
powerful
newconcept forprotein
binding
sites.Immunochem-istry
15:909-936.2. Balachandran, N., D. Harnish, W. E. Rawls,andS. Bacchetti. 1982.
Glycoproteins
ofherpes simplex
virustype2asdefinedbymonoclonal antibodies. J. Virol.44:344-355.
3.
Benjamin,
D. C., M. A.Berzofsky,
J. East,F. R. N. Gurd, C.Hannum, S.J. Leach,E.
Margoliash,
J. G.Michael, A.Miller, E. M.Prager,
M.Reichlin,E. E.Sercarz,S.J.Smith-Gill,P. E.Todd, and A. C. Wilson. 1984. The antigenic structure of
proteins:
areappraisal.
Annu.Rev. Immunol. 2:67-101.4. Chan,W. 1983. Protective immunization of mice with specific HSV-1glycoproteins. Immunology49:343-352.
5. Chou, P. Y., and G. D. Fasman. 1974. Conformational parame-tersfor amino acids in helical a-sheet and random coilregions. Biochemistry 13:211-222.
6. Chou, P. Y., and G. D. Fasman. 1974. Prediction of protein conformation.Biochemistry 13:222-245.
7. Cohen, G. H., B. Dietzschold, M. Ponce deLeon, D. Long, E. Golub, A. Varrichio, L. Pereira, and R. J. Eisenberg. 1984. Localizationand synthesis ofanantigenic determinant ofherpes simplex virus glycoprotein D that stimulates production of neutralizing antibody. J. Virol. 49:102-108.
8. Cohen, G. H., M. N. Factor, and M. Ponce de Leon. 1974. Inhibition ofherpes simplex virus type 2 replication by thymi-dine.J. Virol. 14:20-25.
9. Cohen,G.H.,M.Katze,C.Hydrean-Stern,and R.J.Eisenberg. 1978. Type-common CP-1 antigen of herpes simplex virus is associated with a59,000-molecular-weight envelope glycopro-tein. J. Virol.47:172-181.
10. Cohen, G. H., D. Long, J. T. Matthews, M. May, and R. Eisenberg. 1983. Glycopeptides of the type-common glycopro-tein gD of herpes simplex virus types 1 and 2. J. Virol. 46:679-689.
11. Cohen,G.H., M. Ponce de Leon, and C. Nichols. 1972. Isolation of a herpes simplex virus-specific antigenic fraction which stimulates the production of neutralizing antibody. J. Virol. 10:1021-1030.
12. Dietzschold, B.,R.J.Eisenberg,M. Ponce deLeon, E. Golub, F. Hudecz, A. Varrichio, and G. H. Cohen. 1984. Fine structure analysis of type-specific and type-common antigenic sites of herpessimplex virus glycoprotein D. J. Virol. 52:431-435. 13. Eisenberg, R. J., C. Hydrean-Stern, and G. H. Cohen. 1979.
Structuralanalysisofprecursor andproductforms of type-com-monenvelope glycoproteinD(CP-1 antigen) of herpessimplex virus. J. Virol. 31:608-620.
14. Eisenberg,R.J.,D.Long, R.Hogue-Angeletti,andG. H.Cohen. 1984. Amino-terminal sequence of glycoprotein D ofherpes simplexvirustypes 1 and 2. J. Virol. 49:265-268.
15. Eisenberg, R.J., D. Long, L. Pereira, B. Hampar, M. Zweig,
and G. H. Cohen. 1982. Effect of monoclonal antibody on limitedproteolysisof nativeglycoproteingD of herpessimplex virustype 1. J. Virol. 41:478-488.
16. Eisenberg, R. J., M. Ponce de Leon, and G. H. Cohen. 1980. Comparative structural analysis ofglycoprotein gD ofherpes simplexvirustypes 1 and 2. J. Virol. 35:428-435.
17. Eisenberg, R.J., M. Ponce deLeon, L. Pereira, D.Long, and G. H. Cohen. 1982. Purification ofglycoprotein gD ofherpes
simplexvirustypes 1 and 2by useofmonoclonalantibody.J.
Virol. 41:1099-1104.
18. Gibson, M. G., and P. G. Spear. 1983. Insertion mutants of
herpes simplexvirus haveaduplicationoftheglycoproteinD
geneandexpresstwodifferent forms ofglycoproteinD. J. Virol. 48:396-404.
19. Greenwood,F. C., W. M.Hunter, andJ. S.Glover. 1963. The
preparation of 131I-labeled human growth hormone of high
specific radioactivity. Biochem. J. 89:114-123.
20. Hebrink, P., F. J. van Bussel,and S. 0. Warnaar. 1982. The
antigenspottest(AST):ahighlysensitive assay for the
detec-tion of antibodies. J. Immunol. Methods 48:293-298.
21. Holland,T.C.,S. D.Marlin,M.Levine,andJ.Glorioso. 1983.
Antigenic variants of herpes simplex virus selected with
glycoprotein-specific monoclonal antibodies. J. Virol. 45:672-682.
22. Hopp, T. P., and K. R. Woods. 1981. Prediction of protein
antigenicdeterminants fromamino acid sequences. Proc. Natl.
Acad. Sci. U.S.A. 78:3824-3828.
23. Hubbard,S.D.,and R.J.Ivatt.1981.Synthesisand processing
of asparagine-linked oligosaccharides. Annu. Rev. Biochem.
50:555-583.
24. Kabsch, W.,and C.Sander. 1983. Howgoodarepredictions of
proteinsecondarystructure?FEBSLett. 155:179-182.
25. Kennett,R. H. 1980. Cloningofhybridomas. Cloningin
semi-solid agarose, p. 372-373. In R. H.Kennett,T.J.McKearn,and
on November 10, 2019 by guest
http://jvi.asm.org/
K. Bechtol (ed.), Monoclonal antibodies. Hybridomas: a new dimension in biological analyses. Plenum Press, New York. 26. Kessler, S. W. 1975. Rapid isolation of antigens from cells with
astaphylococcus protein A antibody adsorbent: parameters of theinteraction of antibody-antigen complexes with protein A. J. Immunol. 115:1617-1624.
27. Kuismanen, E., B. Bang, M. Hurme, and R. F. Pettersson. 1984. Uukuniemi virus maturation: immunofluorescence microscopy with monoclonal glycoprotein-specific antibodies. J. Virol. 51: 137- 146.
28. McKearn, T. J. 1980. Fusionof cells inanadherent monolayer, p.368-369.InR.H.Kennett, T. J.McKearn,andK. B.Bechtol (ed.), Monoclonal antibodies. Hybridomas: a newdimension in biological analysis. Plenum Press, New York.
29. Lasky, L. A., and D. Dowbenko. 1984. DNA sequence analysis of the type-common glycoprotein-D genes of herpes simplex virus types 1 and 2. DNA 3:23-29.
30. Lasky, L. A., D. Dowbenko, C. C.Simonsen,and P. W.Berman. 1984. Protection of mice from lethal herpes simplex virus infection by vaccination with a secreted form of cloned glycopro-tein D. Biotechnology 2:527-532.
31. Leach, S. J. 1983. Howantigenic are antigenic peptides? Bio-polymers22:425-440.
32. Lee,G. T.-Y.,M. F.Para, and P.G. Spear. 1982. Location of the structural genesfor glycoproteins gD and for other polypep-tides in the S component of herpes simplex virus type 1 DNA. J. Virol. 43:41-49.
33. Liu, F. T., M. Zinnecker, T. Hamaoka, and D. H. Katz. 1979. Newprocedures for preparation and isolation ofconjugates of proteins and a synthetic copolymer of D-amino acids and immunochemical characterization of suchconjugates. Biochem-istry 18:690-697.
34. Long, D.,T.J. Madara, M. Ponce deLeon,G. H.Cohen,P. C. Montgomery, and R. J. Eisenberg. 1984. Glycoprotein D pro-tects mice against lethal challenge with herpes simplex virus types 1and 2. Infect. Immun. 37:761-764.
35. Lubeck, M.,and W. Gerhard. 1982.Conformationalchangesat topologically distinct antigenic sitesontheinfluenzaA/PR/8/34 virus HA molecule are induced by the binding of monoclonal antibodies. Virology118:1-7.
36. Marchalonis, J. J. 1969. An enzymic method for the trace iodination ofimmunoglobulins and otherproteins. Biochem. J. 113:299-305.
37. Matthews, J. T., G. H. Cohen, and R. J. Eisenberg. 1983. Synthesis and processing ofglycoprotein Dofherpessimplex virus types1 and2inanin vitro system.J. Virol. 48:521-533.
38. Molday, R. S., and D. MacKenzie. 1983. Monoclonalantibodies torhodopsin: characterization, cross-reactivity and application as structuralprobes. Biochemistry22:653-660.
39. Noble, A. G., G. T.-Y. Lee, R. Sprague, M. L. Parish, and P. G. Spear. 1983.Anti-gD monoclonal antibodies inhibit cellfusion induced by herpes simplex virus type 1.Virology129:218-224. 40. Paoletti, E., B. R. Lipinskas, C.Samsonoff, S. Mercer, and D. Panicali. 1984. Construction of live vaccines usinggenetically engineered poxviruses: biological activity of vaccinia virus recombinants expressing the hepatitis virus surface antigenand the herpessimplex virus glycoproteinD.Proc. Natl. Acad. Sci. U.S.A.81:193-197.
41. Pereira, L., D. V. Dondero, D. Gallo, V. Devlin, and J. D. Woodie. 1982. Serologicalanalysis of herpes simplex virus types 1 and2with monoclonalantibodies. Infect. Immun. 35:363-367. 42. Pizer, L. I., G. H. Cohen, and R. J. Eisenberg. 1980. Effect of tunicamyin on herpes simplexvirus glycoproteinsandinfectious virus production. J. Virol. 34:142-153.
43. Rawls, W. E., N. Balachandran, G. Sisson, and R. J. Watson. 1984. Localization ofatype-specific antigenic site on herpes simplex virus type2glycoproteinD. J.Virol. 51:263-265. 44. Rector,J. T., R. N. Lausch, and J. E.Oakes.1984. Identification
ofinfected cell-specific monoclonal antibodies and their rolein hostresistance to ocular herpes simplex virus type1infection. J.Gen. Virol. 65:657-661.
45. Rose,G. D. 1978.Prediction of chain turns inglobular proteins on a hydrophobic basis. Nature (London) 272:586-590. 46. Ruyechan, W. T., L. S. Morse, D. M. Knipe,and B. Roizman.
1979. Moleculargenetics of herpes simplex virus. II. Mapping of themajorviral glycoproteinsandofthegenetic loci specifying thesocial behavior of infected cells. J. Virol. 29:677-697. 47. Sela, M., B. Schechter, I. Schechter, and A. Borek. 1967.
Antibodiestosequential and conformational determinants. Cold Spring Harbor Symp. Quant. Biol.32:537-545.
48. Showalter, S. D., M. Zweig, and B.Hampar. 1981.Monoclonal antibodiestoherpes simplex virus type1proteins, including the immediate-early protein ICP4. Infect. Immun. 34:684-692. 49. Spear, P. G. 1976. Membrane proteins specified by herpes
simplex viruses. I. Identification of fourglycoprotein precursors and theirproducts in type 1-infected cells. J. Virol. 17:991-1008. 50. Watson,R.J.1983. DNAsequenceof theherpes simplexvirus
type 2glycoprotein Dgene.Gene 26:307-312.
51. Watson, R. J., J. H.Weis, J. S. Salstrom,and L. W. Enquist. 1982. Herpes simplex type 1 glycoprotein D gene: nucleotide sequence and expression in Escherichia coli. Science 218: 381-383.

![FIG.7antibodiesanti-gD-1(rabbitportions340-356[1]ng);V, 57S. 4. Immunoblot analysis of synthetic peptides which mimic of gD-1, using polyclonal antibodies and MCAb](https://thumb-us.123doks.com/thumbv2/123dok_us/1412453.94117/6.612.315.559.76.312/antibodiesanti-rabbitportions-immunoblot-analysis-synthetic-peptides-polyclonal-antibodies.webp)