0022-538X/90/073509-07$02.00/0
CopyrightC) 1990, American Society for Microbiology
NOTES
Expression of Collagenlike
Sequences by
a
Tumor
Virus,
Herpesvirus Saimiri
PETERGECK,lt* SCOTTA. WHITAKER,1t MARIAM. MEDVECZKY,'t ANDPETER G. MEDVECZKY" 2t
Departments of
Pharmacology'
and Molecular Genetics and Microbiology,2 University ofMassachusetts Medical School,Worcester, Massachusetts 01655
Received 20 December 1989/Accepted2April 1990Sequencing demonstrates
that the oncogenic regions ofa groupA strain anda groupC strain of herpesvirussaimiri are nonhomologous. A bicistronic viral mRNA from this region is transcribed in tumor cells
transformed
byahighly oncogenicgroupC virus. The firstopenreading frame ishomologoustocollagen; nosuch
sequences werefound
ingroupAorB strains. This is thefirstreportthatavirus encodes forsequencessimilar
tothose of
aconnective tissue protein.Unlike
alphaherpesvirus (e.g., herpes simplex virus)
andbetaherpesvirus (e.g., cytomegaloviruses),
members of thegammaherpesvirus subfamily (Epstein-Barr virus,
herpesvi-rus
saimiri, herpesvirus ateles, and herpesvirus
sylvilagus) areable
toinduce lymphoproliferative diseases
(39, 40).Herpesvirus saimiri is used
as amodel for studying
humanlymphoproliferative malignancies, since its genetic
structureand molecular biology
arerelated
tothose of
the humanEpstein-Barr virus (14). Herpesvirus saimiri shows high
oncogenicity
inNew World
monkeys (13) and New Zealand
Whiterabbits
(9, 30), The transformed cells of the
acutelymphomas and leukemias induced by the virus
areCD8-positive
Tlymphocytes
with NKactivity (21, 24) and
carry acircularform
of the originally linear virion DNA with 110
kilobase
pairs (kbp) of unique
sequences(L-DNA, 36%
G+C) flanked
by
tandemrepeats(H-DNA,
71% G+C)(3).
The
rightmost L-DNA region (polarity analogous
tothat of
Epstein-Barr
virusDNA)
isthought
tocontain
theonco-gene(s). This hypothesis is based
onfour lines of evidence:
(i)
theregion
isalways
retained in immortalized cell lines(23,
44),
(ii)deletions in thisregion
result in loss ofoncogenicity
without
anyeffect
on virusreplication (11, 25), (iii)
variabil-ity
of thissequenceforms threehomology
groups(A, B,
andC)
with differentoncogenic potencies (30, 32),
and(iv)
inintergroup recombinants
thecharacteristiconcogenic
trait isconveyed by
thisregion (30).
In strain 11 ofgroupA, unique
features established sofar in thisregion
involve five genesfor
U-type
small RNAs(27, 51),
adihydrofolate
reductasegene
(50),
and anopenreading
frame with nohomology
to known sequences but with apresumed
function in immor-talization(35),
as shownby
deletionmapping. Only
theU-type
small RNAs aretranscribed,
and nocorresponding
mRNAs have been detected in strain 11-transformed cells. Efforts in this
laboratory
were focused on therightmost
L-DNAregion
of group C strain484-77,
which ishighly
oncogenic
inrabbits. Severallymphoblastoid
tumor-derived cell lines have been establishedfromthymuses
andspleens
*Correspondingauthor.
tPresent address: Department of Medical Microbiology and
Immunology, CollegeofMedicine, UniversityofSouthFlorida,Box
10, Tampa,FL33612-4799.
of
tumor-bearing animals (30) and cultured in RPMI medium
supplemented
with20% fetal calf
serumand 10 ,ug of
gentamycin sulfate
perml.
The cell line used in these
experiments (484-77T) had been continuously cultured for
approximately
1yearprior
toRNA isolation and
wasshown
tocarrynolinear viralgenomes
but
approximately 50 copies
of circular viralgenomes per
cell. No virus
production
wasdetected by cocultivation
onowl monkey kidney cells,
solytic-cycle
genesareprobably
notexpressed.
Cloning
the6.4-kbp
484-77right-hand
terminal L-DNA.The
6.4-kbp clone (containing 6.2 kbp of L-DNA)
wasgenerated by
PstIdigestion
of a cloneconsisting
of therightmost terminal 14-kbp L-DNA in
a21-kbp insert of
484-77cloned into thevector
pHyg,
which confers resistance tohygromycin
B(30, 47).
Theoriginal 21-kbp 484-77 clone
was
generated by Sau3AI partial digestion of the full-length
484-77 viralgenomefollowed
by
sucrosegradient
size frac-tionation andcloning
into thepHyg
vector(14).
The6.4-kbp
fragment
was subcloned into the PstI site of theBluescript
M13-SK vector(Stratagene).
A setof small subclones wasthen created
by taking advantage
of themapped
restriction endonucleasesites,
and alibrary
of clones with "nesteddeletions"wasalso
generated by
standardprotocols (18, 19)
(Fig. 1).
A
virus-specific
mRNA transcribed in tumor cell lines ismapped
inaregion
of the genome involved inoncogenicity.
Cytoplasmic
RNA was isolatedby
Nonidet P-40lysis,
asdescribed
previously (12),
from the cell line(484T)
derived from strain 484-77-induced tumor, fromthe strain 11-trans-formed cell line(1670),
and from uninfectedlymphocytes.
Thepreparations
werepoly(A)
selected andprobed
onNorthern
(RNA)
blotsby using
standardprotocols (31, 38,
49)
with the6.4-kbp
sequencecontaining
the6.2-kbp
right-most L-DNAregion.
Theprobe
did nothybridize
to strain11-specific
oruninfectedcytoplasmic RNAs,
butin thecells derived from 484-77-inducedtumor,a1.2-kilobase(kb)
RNA withpoly(A)
tailwasdetected(Fig. 2) (four
smallRNAs intotal unselectedRNA
preparations
werealsodetected;
datanot
shown).
Mapping
andanalysis
of the mRNA was carried outby
RNaseprotection
assays andprimer
extension.Nearly
50overlapping
subclones were used forstrand-specific
RNA3509
on November 10, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 10, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 10, 2019 by guest
http://jvi.asm.org/
3510 NOTES
L-DNA (110 kb)
EcoRV XbaI PvuII BamHI Scal
I I I 1-
1-H - DNA
I111111111T
^viral
£LLLLLLLUgenome
SmaI PstI
I
Subclones Mayposition
p484PRO.8 (
p484RX0 .4 p484XBO. 5 p484BPsO. 7 p484ScSmO.4
p484PP1.3
p484RPsl.5 ( pHBND1.9 pHBND1.8 pHBNDl.6
pHBNDI.5
pHBND1.3 pND1.0 pNDO.9 pNDO.8
(2081 to 1269) (1269 to 889) (889 to 429) (429 to -254) (365 to -60) (2081 to 751)
(1269 to -254) (1929 to 429) (1785 to 429) (1589 to 429) (1552 to 429) (1262 to 429) (999 to -254) (910 to -254) (805, to -254)
pNDOU.5 )522 to -254)
FIG. 1. Map of subclones ofthe rightmost 2-kbp L-DNA region. Exact positions (Map position) of the subclones and restriction
endonuclease sites used forsubcloningareindicated. Twosetsof clonescreatedbynested deletionsareshown. One set(pND1.0topNDO.5)
wasobtained by unidirectional deletions only, andthe other(pHBND1.9topHBND1.3)hadtheBamHI-PstIfragmentdeletedalso.
probe synthesis by
establishedprotocols (33).
Thehighly
labeled, single-stranded
RNAprobes
werehybridized
in solution to the RNApreparations
from 484T cells. RNase treatmentby
the method of Zinnetal.(55)
reduced the sizes of theprobes,
andonly
thosefragments
of theprobes
whichbecame
protected by
the mRNA remained intact. The totalregion of protection finally
identified themRNA-coding
M
11
484
Uninftected
7.5-
4.4-
2.4-
1.4-
0.3-FIG. 2. Northern blot hybridization of poly(A)+ RNAs from uninfected and tumor-derived cell lines.RNA (4,ug)wasapplied in
eachlane forformaldehyde-1% agarosegel electrophoresis. Num-bers at left indicate sizes in kilobase pairs. Lane M, RNA size marker (six [32P]pCp-labeled synthetic poly(A)-tailed RNAs
[Be-thesdaResearch Laboratories, Gaithersburg,Md.]); lane 11,
cyto-plasmic poly(A)-selected RNA of a group A strain (strain
11)-transformedmonkey cell line (1670); lane 484, cytoplasmic, poly(A)-selected RNA of a group C strain (strain 484-77)-transformed, thymus-derived, rabbit tumor cell line (484T); lane Uninfected, cytoplasmic, poly(A)-selected RNA ofan uninfected human
lym-phoblastoid cell line (Jurkat). The nick-translatedDNAprobe
rep-resented clonedrightmost L-DNAsequencesbetween nucleotides1 and6200, withaportion of theadjoining H-DNA (-1to -254).
DNA sequences. The results of such RNase
protection
assayswithsome of the subclonesare shownin
Fig.
3.Thedata show that the transcribed
region
is an intronlessse-quencebetween nucleotide
positions
1336 and 140(position
+1 is the first nucleotide leftward from the last intact
repetitive
unit of theright-hand H-DNA;
seeFig. 5).
Right-ward
transcription
was indicatedby
the strandedness ofprotection
assays.The exact 5' end was determined
by primer
extensionanalysis by
standardprotocols (22, 29).
Anoligonucleotide
primer complementary
to asequenceclose tothe 5' end of themRNA(as
estimatedby preliminary data)
betweenmappositions
1248 and 1268(5'
CCTGTTTCTTCAGTTGGATAT3')
wassynthesized,
labeled with[y-32P]ATP by
T4 polynu-cleotidekinase, hybridized
at22°C
to 4,ug
of oligo(dT)-selectedRNA,
and extendedby
reversetranscriptase
toward the
5' end of the mRNA to serve astemplate.
Electrophoresis
of the extendedprimer
with asequencing
marker identified the number ofincorporated
nucleotides(Fig. 4).
The data demonstrate that theprimer
wasextendedwith
68 nucleotides from the
3'-terminalnucleotide of
theprimer
atmapposition
1268. In additiontothemajor
band at 68nucleotides,
minor bandsat67 and 66nucleotides referto theirregular
5' end of the mRNA(5'
capstructure),
wherethe reverse
transcriptase occasionally
preterminates.
Nucleotidesequenceand
predicted
aminoacid
sequenceof
ORF1
of the 1.2-kb mRNA arehomologous
tocollagen.
Thegene and
flanking regions
of the1.2-kbp mRNA
werese-quenced by
thedideoxynucleotide-chain
termination method
by
standardprotocols (43, 48)
andanalyzed with the
DNASTARcomputerprogram(DNASTAR, Inc., Madison,
Wis.) (Fig. 5).
The establishedmappositions of the mRNA
wereconfirmed
by the
consensus sequencesfor
transcrip-tion
regulation.
Atypical
TATAbox
wasfound 28
nucleo-tides upstream of the first
nucleotide of
the mRNA,
atposition 1335;
aprimary-transcript
terminalcleavage signal
I-DNA
111111111I
PvuI I
2.0 1.5 1.0 0.5 0 kb
J. VIROL.
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.76.559.57.311.2] [image:2.612.114.245.452.594.2]A
1540-
1380-
1050-
835-
500-M
1
2
3
4
5
B
*NM,p ~
I.
M
1
2
501-
am
404-
_-
331-242-
_
190-C
L-DNA (110 kb)
I
iiProt
l
H-DNA
mm
mII
1 viral.LL.L..LLLLJgenome
ected
0
_- l Probes
800 to -254
1269 to -254
2081 to 751
- 2414 to 429
5099 to -254
522 to -254
365 to -254
Protected
800-140
1269-140
1336-751
1336-429
1336-140
522-140
365-140
Lanes
An A2 A3 A4 A5
B1
B2
FIG. 3. Mapping of mRNA by RNase protectionassay. (A)Autoradiogram from a denaturing 3% polyacrylamide gel electrophoresis
displaying the larger protected fragments. Numbersatleft indicate lengths innucleotides. LaneM,RNAsize marker synthesized by T7 RNA polymeraseontemplates of determined size; lanes 1 through 5,protected RNA fragments (map positions of probes and protected fragments are depicted in panel C). (B) Autoradiogram from adenaturing 6% polyacrylamide gel electrophoresis displaying the smaller protected
fragments. Lane M, DNA size marker(pUC19 HpaII digestion, labeled with [32P]dCTP by using the Klenow fragment of Escherichia coli DNApolymerase); lanes 1 and 2, protected RNA fragments (map positions of probes and protected fragmentsaredepicted in panel C). (C)
Map of probes and protected fragments. Numbersonthemapindicate kilobase pairs. Theexactmappositions of probes (-) andprotected
fragments (-)arealsoshown, with referencestothecorresponding lanes in panels A and B.
TGTGTGTG
sequence waslocalized
atthe 3'
end of the
mRNA;
and apoly(A) addition signal AATAAA
sequence wasidentified 40 nucleotides
upstream. Twomajor
openreading
frames (ORFi and ORF2)
wererevealed. ORFi
codes for99 amino acids between
nucleotides
1290 and994
(Fig. 2).
Thecoding
capacity of ORF2
is 214 amino acids from nucleotides 812 to 171. The latter is not discussedfurther,
since nosignificant homology
was found in the GenBank data.Inthe sequence
of
ORF1, however,
a remarkablestruc-ture has
been
revealed.
Ofthe total
297nucleotides,
162 code forarepetitive
sequence withanelement of 18 nucle-otidesrepeated
intandem ninetimes,
withminor irregular-ities inthe first and last units(Fig. 5A). High similarity
wasfound between ORFi and different forms of
collagen
of diverseorigin (not shown).
The greatestsimilarity
to ORFiwas
displayed by
acollagen
sequenceof
anematode,
Cae-norhabditiselegans (26),
forwhich 116 of162nucleotidesin therepetitive region
were identical(71.6%);
theflanking
nonrepetitive
sequencesofORFi also showedsimilarity
tocollagen (Fig. SB). High homology
wasdemonstratedwhen amino acid sequences of various knowncollagens
and theputative herpesvirus
saimiriprotein
sequence werecom-pared (Fig. 6).
Collagenlike
sequences arepresent
in group C strains ofherpesvirus
saimiri and absentingroupA andB strains. To obtain virion DNA for Southernhybridization,
OMK cells.0
Sequencing
_
markers
LZ
G A
T C
68N-54N1-li
FIG. 4. Primerextension analysis of the 5' end of the mRNA.
Autoradiogram from a denaturing 6% polyacrylamide gel
electro-phoresis showingtheextendedprimer togetherwithasize marker
sequencingladder. Only aportionof the sequencing gel isshown.
The sequence itself is unrelated and is used merelyas a marker.
LaneExtension, Extendedprimer;lanes G, A, T,andC, partofa
size markersequencing ladder,shownonlyfrom nucleotide(N)54.
G, A, T,and C label the individualchain termination reactions.
H-DNA
16~~~~~~~~~~~~~~~~~~~~~~~~~~
6 5 4 3 2
I I- .- I _._ I
C.-i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.83.528.76.360.2] [image:3.612.377.502.467.651.2]3512 NOTES
A
gagtttccaaaatgtagtaagctaacaTATAAATatcagc agaaaatagagtaacAcaagaaacagctaacaagagcaacacaaacacgc ttgaaagaaaATGGCAAGCGAGCCAAACCTAAGATATCCAACTGAAGAAA CAGGAGATCGAGGACCTCAAGGACCTCCAGGACCTCCAGGACCCCAAGGA CCTCCAGGACCCCAAGGACCTCCAGGACCCCAAGGACCTCCAGGACCCCA AGGACCTCCAGGACCCCAAGGACCTCCAGGACCCCAAGGACCTCCAGGAC CCCAAGGACCTCCAGGACCTTCAGACTCCAGGATTGTTTGTAACAAACTT ATTGCTTGGAATCATAATTTTACTCTTATTAATTATAGTTGCGATCTTAC TGGTGTCTAAttagtagtaaactaagaacagagcaagcagtatagaagag agtaattatgctagcttaatttcatctagtagtcagttgtgtagaacttt ttattagtaaatctagaagtgttaagaaaaaggtagagaaagtagtgacc tttattagctgcatcttcatatctctaaattagtaagtATGGAAAACCAG AGAGAAGAAATAGAACTAACAGAAATTCCAGAGACTGAGAAAAAAAGAAC AGCTGAAGAAAAATTATTATCATGCTCTGCTGAAACTGCTGAAGAAAAGG
TATCATTATGCTCTGAAGAAACTACAGACACGTCAAGCAGTTCTAGCAGT GAGCAAACACCTGCTCCTATAGAAGTAAACGTTAACATTCAAACGAGCAC ATACTTGCCTCAGAACGCGGCAACAAACTTGAACAGTTTGTACACAAGTT TTGAAGACGCAAGGGCACAAGGAAAAGGACTGGTGCGACACAATTCGGAC GATTTAAAAAGTTTTTTAGAAAAATACCCACCAGATTATAGAAAACCTAA ACGGGACTTGTCAGAATCATGGGATCCAGGTATGCCGAAACCAACTTTGC CTCCTAGGCCTGCGAACCTAGGTGCAAGCCAGGCAAGTACTGTAAGACGG CATGTAAGAGAACAAAATTTTAAACAGCTTAGAGAACGTAAAGCTAATGA AGGGAAAATAGTAAAAGATTTGAAAAGACTTGAGTACAAAGTTAATATAA TTCTTTGTTTAGTAGTTGTTATATTAGCTATAATATTATTACTAACTGGG
CTAAGTATACTGTTCATAAGAATAAAAAGTTAAtttaacaaagaatttaa caaataTGTGTGTGcatggtcattaaaagaagcgggccccaagcagtggg cgctgggcagctggaaaaatgtcctgggcgtggctagagcgagctcgggg ggcaaagcgagcagcgagcggctgcggagggagggggaagggattgagcT gccccgcccctctggggtgagattccagagcccctgttgtgtccccaggc caccgcaggcccggggctgtgcccctgtgcttggagccagtccccagagc cctggctaggccgaggcgctgctagagtaagggtcccacaactgttcagg cccccccagggggagagcccagcgcagcgctctgccgggtaggggggggg tagctggctccccccccacctctgcgtgaagcagagtgcatgcagcccct
1351 1301 1251
1201 1151 1101
1051 1001
951
901
851
801
751
701
651 601 551 501 451 401 351 301 251 201 151
101
51 1
-50
-100 -150 -200 -250
TATA-box
Start of mRNA
Initiator codon
ORF 1
Terminator codon Initiator codon
ORF 2
Terminator codon Cleavage signal
of mRNA
"H" sequences
B
ORF 1 ATGGCAAGAAA ATAGGAG CG
collagen 1109 GCTATTT
GCAGGAG
CCCCCGA
TCORF 1 GGACCTCCCGC
collagen 1159
cCCAGGACdA1CCAGG
CGACGAORF 1 AGGAC G CAGGAC G
collagen 1209 AGGAC G CAGGAC GAC C
ORF 1 GA T c GGAC CCAGGA
collagen 1259 GA
GAC
GGACdCAG
ORF 1 CG CTTC CT ATT CAAACTT A
collagen
1309 C5AGGAG cc T GG TORF 1 AT T GCG ATC C
collagen 1359 TATCAGGCTCTCG G C
FIG. 5. Nucleotide sequence of the mRNA coding region. (A) Nucleotide sequence between nucleotides -250 and 1390. Transcription consensus sequences and open reading frames are shown in capital letters. A (at nucleotide 1335),Start point of transcription; T (at nucleotide 1), first nucleotide of L-DNA. The initiation and termination codons are printed in boldface letters. In ORF1, each repetitive unit is underlined. Negative numbers refer to a portion of the first intact H unit. (B) Comparison of
ORFl
and a collagen sequence (C. elegans; GenBank identifier, CELCOL1G). In the collagen sequence, an insert of AGTGAGT atnucleotide 1335 is omitted. Boxes represent full homology.were
infected with the following strains representing the
three
subgroups: strains 484-77, 487-77, and 505-77 (all group
C);
strain 11 (group A); and strain S295C (group B). DNA
wasisolated from the virions as described previously (30).
Restriction endonuclease digestions, electrophoresis,
vac-uumblotting, and hybridization with nick-translated DNA
probe
were
performed by using standard protocols (31, 38,
49).
The
ORFl
sequence was present in the three group C
strains
tested, with no variability in length of the
ORFl-containing
fragments (Fig. 7).
As
expected, no hybridization was detected in a
repre-sentative
from group A or B. No homology was found
between
ORFi
or
ORF2 nucleic acid or predicted protein
sequence
and the
published
right-end sequence of strain 11
(group A) (35)
with
AALIGN or ALIGN programs
(DNASTAR,
Inc.)
orFASTA programs ALIGN,
RDf2,
or
PLFASTA (37)
(data not shown).
Sequencing
the
oncogenic DNA region of a highly
onco-genic herpesvirus
saimiri
strain and studying its expression
by transcription analysis
resulted in three major and
poten-tially important
observations. (i) Earlier hybridization data
that the
rightmost
L-DNA sequences are nonhomologous
among
different groups of herpesvirus saimiri have been
confirmed
by sequencing. (ii)
This is the
first
report
in
which
an mRNAfrom the
oncogenic region in transformed cells
has
been detected and characterized. (iii) This is also the first
J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.127.505.85.402.2]HVS TGDRGPQGPPGPPGPQGPPGPQ--
-GPPGPQGPPGPQGPPGPQGPPGPQGPPGPPGPPGPSDSRIVCNKLIAWN
1. P S P AP ND P--- K A D AD GS P A T EKGICPK
2. P EA A FA AD--- Q AK E DT VK DA
3.
S AS A R4. S AS A R
5. S AS A R
SA S KDGLN L I SA A KDGLN L I SA S KDGLD L I
A A
x R RT DA A P
R RT DA V P R RT DA S P
KGP GAAGPPG TG PSGGYDLSF PQPP
PSAGFDFSF PQPP PSGGYDLSFFPEPP
6.
S PA R SA AA KD LNGLP I P R RT EV V P SGFDLDGGGRYY
FIG. 6. Comparison ofamino acid homology in the repetitive region with
alpha(1)
chain sequences of type I collagens from different species. Amino acidsdifferences
areshown relative to the herpesvirus saimiri sequence between amino acid positions 14 and 84. Empty spacesrepresentidenticalamino acids. Lines 1, 2, 3, 4, 5, and 6 are the nematode(CELCOL1G),murine (A23982), bovine(CGBO1S),
human(CGHU1S),
rat (CGRT1S),and avian (CGCH1S) corresponding sequences, respectively (GenBank identifiers are in parentheses). Similar degrees of homology werefound between the saimiri collagenlike sequence and other collagen types (not shown). X, Insert in the murine sequence (IGNVGA). Amino acids 1 to 13 (MASEPNLRYPTEE) and amino acids 85 to 99(HNFTLINYSCDLTGV)
of theherpesvirus saimiri sequence are not depicted.report
in
which a viral mRNA has been shown to have an
open
reading frame
that codes
for a collagenlike sequence.
To
explain the lack of homology between strains 11 and
484-77
in the right-end
1.8-kbp L-DNA region, we favor the
hypothesis
that different cellular sequences have been
ac-quired by these strains.
There is
evidence that herpesvirus
saimiri
readily integrates
cellular sequences into its genome.
The virally
coded
thymidylate synthase (2), dihydrofolate
reductase
(50), and U-type small RNAs (27, 51) are highly
homologous
to
corresponding
human sequences and were
probably
acquired
from the cellular genome. The difference
in
G+C
content suggests
that the
collagenlike
sequence may
also
be of cellular origin (52% G+C,
in contrast to
36%
G+C
in
the
surrounding viral
L-DNA
sequences). However, we
assumethat these cellular sequences then further evolved,
since
no
homology
between cellular and viral sequences can
be detected
by Southern hybridization
(32; P.
Medveczky,
unpublished data).
The
right-end region
of
L-DNA
of
strain
11
(group
A)
has one open
reading
frame with no homology
toORF1
or
ORF2 of
484-77,
asshown,
but
deletion
of the
open
reading frame
results in loss
of
oncogenicity
(35).
These results
suggest
that
the two
viruses
probably
trans-form
T
cells
by different molecular mechanisms.
The viral
message in the tumor-derived
cell line is
1.2 kbA B
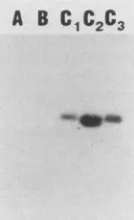
C1C2C3
FIG. 7. Southern hybridizationtodetect
collagenlike
sequences in different strainsrepresenting
thethreehomology
groups. Herpes-virus saimirivirion DNA(about50ng) fromthedifferent
strainswasdigested with PvuII. The nick-translated DNA
probe
represented
clonedsequencesbetweennucleotides 1269and 889. Lane
A,
Strain 11(group A); laneB, strainS295C (groupB);
laneC1,
strain 484-77 (groupC);
laneC2,
strain 487-77(groupC);
laneC3,
strain 505-77 (groupC).long and contains
a3'
poly(A) tail, and our data indicate no
splicing, unless sequences outside the
studied 6.2-kbp region
areinvolved.
ThemRNA
sequence contains two open
reading frames, which is unusual
but not unprecedented
among
eucaryotes.
The
structure
of the
saimiri
collagenlike (SCOL) sequence
is
coherent with basic patterns in
collagen structure (4, 53).
Incollagen,
anine-nucleotide
element codes for Gly-X-Y
amino acidtriplets (X and
Yare
frequently proline and
hydroxyproline, respectively) which are repeated several
hundred
times inindividual
polypeptides of the
triple helical
structure(26, 36; for
areview,
seereference
6). A
charac-teristic element of collagen evolution is
a54-nucleotide unit.
Therelics of
thisevolutionary stage
arestill present in
the exonsof
achicken
collagen,
in
which
mostof the 50
exonscontain
54nucleotides
(6, 36). In
herpesvirus
saimiri,
the162-nucleotide sequence appears
tobe the
result of
exactly
three reiterations of
a54-nucleotide unit. The absence of
splicing
may indicate
a reversetranscription
step
prior
tointegration into the viral genome.
The
perfect invariable repeats
are oneof the
intriguing
features of the saimiri
collagenlike sequence,
since incolla-gen
the X and Ypositions
arehighly
variable. Thepresence
of the sequence
wasdetected without
fragment
variability
inall
tested group C strains
by
Southern
analysis.
This
dem-onstratesthe conservation of thesequence
within thesub-group,
which may indicate the
biological
significance
of
thesaimiri
collagenlike
sequence.
No
evidence is available
onthe function of the
saimiricollagenlike
sequence,
and theexpression
of thepolypeptide
has yet
tobe
demonstrated.
It has beensuggested
thatcollagen plays
arole
incellular
growth regulation,
usually by
down
regulating
cell division
(7, 28,
46).
On the otherhand,
very
small
(15,000
to17,000
Mj)
collagenlike polypeptides
that have
stimulating
effects
onembryonic
tissues(42)
andbone
marrow stemcell culture
(52)
have been detected.Mitogenic
stimulation and
celltransformation
after afi-bronectin
fragment
withaffinity
tocollagen
is bound(10, 20)
seem to
indicate
acollagenlike
cell surfacereceptor,
whichmay
actin cell
growth
regulation.
Integrin
receptors
oncytotoxic
T
lymphocytes (nonspecific
adhesionreceptors,
e.g.,
LFA-1,
LFA-3,
VLA-1,
andVLA-2) (5, 16, 17, 34,
41)
may
also be involved
inacollagen-related growth regulation.
The
cellular
and retroviral oncogenes
are derivatives ofon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.109.510.78.202.2] [image:5.612.144.211.540.650.2]3514 NOTES
normal
cellular
counterpartswith physiological
functions
(1,
45).
These proto-oncogenes act atdifferent
stagesof
signal
transduction inthe
inter-
and intracellular pathways
of cell
growth regulation (8, 15, 54).
No experimental evidence sofar has
indicated,
however,that
sequencesrelated
tocon-nective tissue
proteins
areinvolved in
the process.This
report
raises thepossibility
of
agrowth
regulatory
function
for a
virally
encodedcollagenlike
polypeptide.
We thankLarry Hardy, Carel Mulder, Rosemary Vassallo, and
Douglas Waud for critical reading of the manuscript. We thank
DanielMullenfortheexcellent photographic work.
This work was supported by Public Health Service grants RO1 CA43264, RO1 CA39653, and Al 21129fromtheNational Institutes
ofHealthandbiomedicalresearchsupportgrantSOTRRO5712from
theNationalInstitutesofHealth totheUniversity of Massachusetts
MedicalSchool.
LITERATURE CITED
1. Adamson, E. D.1987.Oncogenesin development. Development
99:449-471.
2. Bodemer, W., H.H. Niller, H. Nitsche, B. Scholz, and B.
Fleckenstein. 1986. Organization of the thymidylate synthase
geneofherpesvirus saimiri. J. Virol. 60:114-123.
3. Born4amm,G.,H.Delius, B.Fleckenstein,F.-J. Werner,andC.
Mulder. 1976. Structure of Herpesvirus saimiri genomes:
ar-rangement ofheavy andlight sequences in the M-genome. J.
Virol. 19:154-161.
4. Bornstein, P. 1980. Structurally distinct collagen types. Annu.
Rev. Biochem. 49:957-1003.
5. Buck,C.A.,andA. F.Horwitz. 1987. Cell surfacereceptorsfor
extracellular matrix molecules. Annu. Rev. Cell Biol.
3:179-205.
6. Burgeson, R. E. 1988. New collagens, new concepts. Annu.
Rev.Cell Biol.4:551-577.
7. Buttle, B.J., andH. P. Ehrlich. 1983. Comparative studies of collagenlatticecontractionutilizinganormalandatransformed
cellline.J.Cell. Physiol. 116:159-166.
8. Chiarugi, V., M. Ruggiero, and F. Porciatti. 1987. Oncogenes
andtransmembrane cellsignalling. CancerInvest.5:215-229.
9. Daniel, M. D., L. V. Melendez, R. D. Hunt, N. W. King, M.
Anver,C. E.0.Fraser,H. H. Barahona,and R. B.Baggs.1974.
Herpesvirus saimiri. VII. Inductionof malignant lymphomain
NewZealandWhiterabbits.J. Natl. CancerInst.53:1803-1807. 10. DePetro, G.,S. Barlati, T. Vartio, and A. Vaheri. 1981.
Trans-formation enhancing activity of gelatin binding fragments of
fibronectin. Proc. Natl. Acad. Sci. USA78:4965-4969. 11. Desrosiers,R.C., A.Bakker,J. Kamine,L. A. Falk, R. D. Hunt,
and N. W. King. 1985. A region of the Herpesvirus saimiri
genome requiredforoncogenicity. Science 228:4694-4699. 12. Favoloro, J.,R. Treisman,and R. Kamen. 1980. Transcription
maps ofpolyomavirus specific RNA: analysis by
two-dimen-sionalnucleaseS1 gel mapping.MethodsEnzymol.65:718-749.
13. Fleckenstein, B., and C. Mulder. 1980. Molecular aspects of
Herpesvirus saimiri andHerepesvirusateles, p.799-812.InG.
Klein(ed.),Viraloncology. RavenPress,NewYork.
14. Gompels, U. A., M. A. Craxton, and R. W. Honess. 1988.
Conservationofgene organizationin the lymphotropic
herpes-viruses herpesvirus saimiri and Epstein-Barr virus. J. Virol.
62:757-767.
15. Heldin, C.-H.,and B. Westermark.1984.Growthfactors:
mech-anismofactionand relationtooncogenes.Cell37:9-20.
16. Hemler, M. E., C. Huang, and L. Schwarz. 1987. The VLA
protein family. J. Biol. Chem262:3300-3309.
17. Hemler, M. E., J. G. Jacobson, andJ. L. Strominger. 1985.
Biochemical characterization of VLA-1 and VLA-2. J. Biol.
Chem. 260:15246-15252.
18. Henikoff, S. 1984. Unindirectional digestion with exonuclease III creates targeted breakpoints for DNA sequencing. Gene
28:351-359.
19. Hoheisel, J., and F. M. Pohl. 1986. Simplified preparation of
unidirectionaldeletion clones. NucleicAcidsRes.14:3605.
20. Humphries, M. J., andS. R. Ayad. 1983. Stimulation of DNA
synthesis
by
cathepsin
Ddigests
offibronectin. Nature(Lon-don)305:811-813.
21. Johnson, D.R., and M. Jondal. 1981.
Herpesvirus-transformed
cytotoxic T-cell lines. Nature(London)
291:81-83.22. Jones,K.A.,K. R.Yamamoto,and R.
Tjian.
1985. Twodistinct transcription factors bind to the HSVthymidine
kinasepro-moter in vitro. Cell 42:559-572.
23. Kaschka-Dierich, C., F. J. Werner,
I.
Bauer, and B. Flecken-stein. 1982. Structure ofnonintegrated,
circularHerpesvirus
saimiriandHerpesvirus atelesgenomesintumorcell lines and invitrotransformed cells. J. Virol. 44:295-310.24.
Kiyotaki,
M., R. C.Desrosiers, and N. L.Letvin. 1986. Herpes-virus saimiri strain 11 immortalizes a restricted marmoset T8 lymphocyte subpopulation in vitro. J. Exp. Med. 164:926-931. 25. Koomey,J. M.,C.Mulder,R. L. Burghoff,B.Fleckenstein,andR. C. Desrosiers.
1984.
Deletion ofDNA sequences innonon-cogenic
variant ofHerpesvirus
saimiri. J. Virol.50:662-665.
26. Kramer,J. M., G. N.Cox,
and D. Hirsch. 1982.Comparison
ofthecomplete sequencesoftwo
collagen
genesfrom Caenorhab-ditis elegans. Cell 30:599-606.27. Lee,S.I.,S. C. S.Murthy,J.L.Trimble, R.C.
Desrosiers,
and J. A. Steitz. 1988. Four novel U RNAs are encodedby
aherpesvirus. Cell 54:599-607.
28. Liotta, L.A.,D.Vembu, H. K.Kleinman,
G.
R.Martin, andC. Boone. 1978.Collagen
required
forproliferation
of cultured connective tissuecells, butnot their transformedcounterparts. Nature (London) 272:622-624.29. McKnight, S. L., and R. Kingsbury. 1982.
Transcriptional
control signals of a eukaryotic protein coding gene. Science 217:316-324.30. Medveczky, M. M., E.
Szomolanyi,
R.Hesselton,D.
DeGrand,P. Geck, and P. G. Medveczky. 1989. Herpesvirus saimiri strains from three DNAsubgroups havedifferentoncogenic potentials
in New ZealandWhite rabbits. J. Virol. 63:3601-3611. 31. Medveczky, P., C.-W. Chang, C. Oste, and C. Mulder. 1987.Rapid vacuum driven transfer ofDNA and RNA
from
gels tosolidsupports. Biotechniques 5:242-246.
32. Medveczky, P., E.Szomolanyi, R.C. Desrosiers, and C. Mulder. 1984. Classification of herpesvirus saimiri into three groups based on extreme variation in a DNA region required for
oncogenicity.
J. Virol. 52:938-944.33. Melton, D. E., P. A. Krieg, M. R.
Rebagliati,
T. Maniatis, K. Zinn, and M. R. Green. 1984. Efficient in vitro synthesis of biologically active RNA and RNA hybridization probes from plasmids containing a bacteriophageSP6
promoter. Nucleic Acids Res. 12:7035-7056.34. Mentzer, S. J., S. H. Gromkowsky, A. M. Kremsky, S. J. Burakoff, and E. Martz. 1985. LFA-1 membrane moleculein the regulationof homotypic adhesions of human B lymphocytes. J. Immunol. 135:9-11.
35. Murthy, S. C.
S.,
J. J. Trimble, and R. C. Desrosiers. 1989. Deletionmutantsofherpesvirus saimiri define an openreading frame necessary for transformation. J. Virol. 63:3307-3314. 36. Ohkubo, H., G. Vogeli, M. Mudryj, V. E. Avvedimento, M.Sullivan, I. Pastan, and B. de Crombrugghe. 1980. Isolation and characterization of overlapping genomic clones covering the chicken alpha 2 (type
I)
collagen gene. Proc.Natl.
Acad. Sci. USA 77:7059-7063.37. Pearson, W. R., and D. J. Lipman. 1988. Improved tools for biological sequence comparison. Proc.
Natl.
Acad. Sci. USA 85:2444-2448.38.
Rigby,
P., M. Dieckmann, C. Rhodes, and P. Berg. 1977. Labeling DNA to high specific activity in vitro by nick transla-tion with DNA polymeraseI.
J. Mol. Biol. 113:237-252. 39. Roizman, B. 1982. The family Herpesviridae: generaldescrip-tion, taxonomy, classificadescrip-tion, p. 1-23. In B. Roizman (ed.), The herpesviruses, vol. 1. Plenum Publishing Corp., New York. 40. Roizman, B., L. E. Carmichael, F. Deinhardt, G. de-The, A. J.
Nahmias, W. Plowright,
F.
Rapp, P. Sheldrick, M. Tadahashi, and K. Wold. 1981. Herpesviridae definition, provisional no-menclature, and taxonomy. Intervirology 10:201-217.41.
Ruoslahti,
E. 1988. Fibronectin and its receptors. Annu. Rev. J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
Biochem. 57:375-413.
42. Sage,H.1985. Lowmolecular weight fibroblastcollagen:
struc-ture, secretion, anddifferential expressionasafunction of fetal
and cellularage. Biochemistry. 24:7430-7440.
43. Sanger, F., S. Nicklen, andA. R.Coulson. 1977. DNA
sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
44. Schirm, S., I. Muller, R. C. Desrosiers, and B. Fleckenstein.
1984. Herpesvirus saimiri DNA in a lymphoid cell line
estab-lished by in vitrotransformation. J. Virol. 49:938-946. 45. Sibbitt, W. L., Jr. 1988. Oncogenes, normal cell growth, and
connective tissue disease.Annu. Rev. Med. 39:123-133. 46. Smith, B. D., A. P. Mahoney, and R. S. Feldman. 1983. Inverse
correlation of collagen productionto anchorage independence and tumorigenicity in W-8 and M cell lines. Cancer Res. 43:4275-4282.
47. Sugden, B., K. Marsh, and J. Yates. 1985. A vector that replicates as a plasmid and can be efficiently selected in
B-lymphoblasts transformed by Epstein-Barr virus. Mol. Cell Biol. 5:410-413.
48 Tabor,S., and C. C. Richardson. 1987. DNAsequenceanalysis
withamodified bacteriophage T7 DNA polymerase. Proc. Natl.
Acad. Sci. USA 84:4767-4771.
49. Thomas, P. S. 1980. Hybridization of denatured RNA and small DNAfragmentstransferredtonitrocellulose. Proc.Natl. Acad. Sci. USA 77:5201-5205.
50. Trimble, J. J., S. C. S. Murthy, A. Bakker, R. Grassmann, and R.C. Desrosiers. 1988. Agenefor dehydrofolate reductase in herpesvirus. Science 239:1145-1147.
51. Wassarman, D. A., S.I.Lee, and J. A.Steitz. 1989.Nucleotide sequence of HSUR 5 RNA from herpesvirus saimiri. Nucleic Acids Res. 17:1258.
52. Waterhouse, E. J., P. J. Quisenberry, and G. Balian. 1986. Collagen synthesis by murine bonemarrowcell culture. J. Cell.
Physiol. 127:397-402.
53. Yamada, Y., V. E. Avvedimento, M. Mudryj, H. Ohkubo, G. Vogeli, M. Irani, I. Pastan, and B. de Crombrugghe. 1980. The
collagengene: evidence for itsevolutionary assembly by ampli-fication ofaDNA segment containing an exonof 54bp. Cell
22:887-892.
54. Yarden, Y. 1988. Growth factor receptor tyrosine kinases.
Annu. Rev. Biochem. 57:443-478.
55. Zinn, K., D.DiMaio, and T. Maniatis. 1983.Identification oftwo
distinctregulatory regions adjacenttothe human betainterferon
gene.Cell 34:865-879.
on November 10, 2019 by guest
http://jvi.asm.org/
ERRATA
Expression
of
Collagenlike Sequences by a Tumor Virus,
Herpesvirus Saimiri
PETERGECK,SCOTT A. WHITAKER, MARIA M. MEDVECZKY, AND PETER G.MEDVECZKY Departmentsof Pharmacologyand Molecular Genetics and Microbiology, University of Massachusetts
Medical
School,
Worcester,
Massachusetts 01655Volume 64, no. 7, p. 3512 and 3513: The nucleotide sequence in Fig.
5A, between nucleotide positions 1101 and 976, and
the
corresponding
amino
acid
sequencein
Fig.
6,
between
amino
acid
positions
64
and
105,
should read
asshown
below:
CCC
CCA
GGA CCT CCA GGA CCT TCA GGACTG
CCA GGA TTG TTT GTA ACA AAC TTAP P G P P G P S G L P G L F V T N L
TTG CTT GGA ATC ATA ATT TTA CTC TTA TTA ATT ATA GTT
GCG
ATC TTA CTG GTGL
L G I I I L L L L I I V A IL
L VTCT AAA TTA GTA GTA AAC TAA
S K L V V N
Induction of
Human
Immunodeficiency
Virus Type 1
Expression
in
Chronically
Infected Cells
Is
Associated
Primarily
with
a
Shift in RNA
Splicing Patterns
NELSONL. MICHAEL, PAULMORROW,JOSEPH MOSCA, MARYANNEVAHEY, DONALDS.BURKE,ANDROBERT R. REDFIELD
Departmentof RetroviralResearch and DivisionofRetrovirology, Walter Reed Army Instituteof Research, 13TaftCourtSuite 200, Rockville, Maryland20850,andSRA Technologies and
Henry M.JacksonFoundation, Rockville, Maryland20850