Analysis of the Termini of the DNA of Bovine
Parvovirus:
Demonstration of Sequence Inversion
at
the Left Terminus and
Its
Implication for the Replication Model
KATHERINE C. CHEN,* BRUCE C. SHULL, MURIEL LEDERMAN, ERNEST R. STOUT,
AND ROBERT C. BATES
Department of Biology, VirginiaPolytechnicInstituteandState University, Blacksburg, Virginia24061 Received29October1987/Accepted6July 1988
The distribution of terminal-sequence orientations in the viral DNA of bovine parvovirus (BPV), an autonomousparvovirus,wasstudiedby endlabeling and restrictionenzymedigestion and also by cloning. The
left (3') end of theminus strandofBPVwasfound intwoalternativesequenceorientations(designatedasflip
andflop, whichare reversecomplementsof each other), witha10-foldexcessof flip. This is incontrasttothe autonomous rodent parvoviruses whichencapsidate'minus-strand DNAwithonly the flip orientation atthis
end. Theright(5') end of the minus strand ofBPVcontainedbothsequenceorientationswith equalfrequencies, asin the rodent parvoviruses. Sequenceinversionswerealsodetected'atboth endsof the plus strand,'which
makesupabout10% ofthe encapsidated BPV DNA. Each terminus of BPV DNA hadacharacteristic ratioof
flip toflop forms, and thisratiowasrestored in the progeny DNA resulting fromtransfection withgenomic
clones of different defined terminal conformations. Replicative-form DNA showed the same distribution of
terminal-sequence orientationsasthe reannealedplus andminusvirion DNAs,suggestingthatthedistribution offlip and flop forms observedinvirionDNA isnotduetoselectiveencapsidation, but rathertothespecific distribution ofreplicative forms. Thecurrentreplicationmodelforautonomousparvoviruses,whichwasbased onthe available datafor the rodent parvoviruses,cannotaccountfortheobserveddistribution ofBPVDNA.
Analternativemodelis suggested.
Parvoviruses are small icosahedral viruses with
single-stranded DNA genomes containing terminal palindromic sequences.Theseterminal regions have cis signals important in various steps of replication (6, 8, 9, 14). A number of related models have been proposed for the replication of parvoviruses (1, 3, 7, 8, 14, 16, 21, 30, 31). An essential feature included in all these models is a hairpin transfer
mechanism originally proposed by Cavalier-Smith (11) for thereplication of linear molecules containing terminal palin-dromes. According to this mechanism, ifa terminal
palin-dromeisimperfect,twoterminal sequenceswillresult from thereplicationprocessand each will be thereverse comple-ment of theother, arbitrarily assigned asfliporflop (3, 25).
Ifarestrictionenzymerecognitionsequencelies withinsuch anunpaired region in thesingle-stranded DNA, that
restric-tion enzymewill bediagnostic for the flip and flop
orienta-tions, since itwill cutdouble-stranded DNAattwo alterna-tivesitesdepending onthe orientation (21).
The dependent parvovirus, adeno-associated virus (AAV), encapsidates equal amounts of plus- and minus-strandDNA. The left andright terminiareidentical, and the two alternative sequence orientations are found withequal
frequenciesatboth endsof theplus- and minus-strand DNA, consistent with thehairpin transfer mechanism (8, 17). The autonomous parvoviruses encapsidate various amounts of
minus-strand DNA, from 99% for the rodent parvoviruses, which include minute virus of mice (MVM), H-1, and rat virus,to90% for bovine parvovirus (BPV),to50% forLuIlI (14, 28). The left and right termini of the autonomous
parvovirusesarenotidentical. Theterminal-sequence
orien-tations have sofar beenstudied indetailonlyfor therodent
parvoviruses (2-4, 24). The right (5') termini ofthe minus
*Correspondingauthor.
strands of these viralgenomes have been shownto exist in either orientation with equal frequencies, but the left (3') termini areunique. Amodified rolling hairpin model (1, 14)
was developed to account for the absence of sequence
inversion at the left end. However, the generality of this model for the otherautonomousparvoviruses remains tobe determined.
In thisstudy, the distribution of terminal-sequence orien-tations in the viral DNA population of BPV, a nonrodent parvovirus, wasinvestigated by end labeling and restriction enzyme analyses and also by cloning. The results
demon-strated the presence of both flip and flop conformations at
the left end of the minus strand ofBPV, incontrast to the
unique conformationatthis endfor therodentparvoviruses.
The relation of this finding to the models of parvovirus replication is discussed.
MATERIALS AND METHODS
Cloneconstruction. The construction and characterization
of BPV infectious genomic clpnes pVT501, pVT502, and
pGCSma20 have been described previQusly (27). Left and
right terminal fragments ofBPVwerecloned intopUC8,and
terminal SmaI fragments were cloned'into M13mpl8 by
standard procedures (18).
End-labelanalysis.Procedures forisolatingDNAs (single-and double-stranded virion DNAs, as well as
replicative-form[RF] DNA)weredescribedpreviously (12). DNA,1 to
2 pugper50-,ulreaction, was'labeledatthe3'-hydroxylends
with 50 ,uCi of [35S]ddATP (Dupont, NEN Research
Prod-ucts,Boston, Mass.)and 20U'ofterminal
deoxynucleotidyl-transferase(Dupont)inas-olutioncontaining100 mM potas-sium cacodylate, 25 mM Tris hydrochloride, 0.2 mM dithiothreitol, and 1 mM CoCl2(pH-7.6) (23). After incuba-tionat37°C for2h,DNAwasprecipitatedandwashedthree 3807
0022-538X/88/103807-07$02.00/0
Copyright© 1988, American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
3808 CHEN ET AL.
times
with
70% ethanol.The labeled DNAwasdigested
withthe restriction enzymes
diagnostic
for theflip
andflop
orientationsasindicated in Results.Thedigestion
products
were
analyzed
on asequencing gel (10) by using M13mp18
sequencing
reactions as size markers. End-labeled restric-tionfragments
would be one baselonger
than the sizespredicted
from the DNA sequence as a result ofthe extrabase
(ddA)
added
by
theterminal transferase.RESULTS
BPV
encapsidates approximately
90% minus-strandDNAand 10%
plus-strand
DNA,
and virion DNAs ofopposite
polarity readily
reanneal under the conditions used forisolation
(12).
VirionDNA was recoveredintwofractions:double-stranded
DNA,
consisting
of reannealedplus
and
minus
strands,
andsingle-stranded
DNA of minuspolarity
only.
Weassumethat theminusstrands in reannealedvirionDNAare a
representative sample
of the total minus-strandpopulation. By convention, parvoviral
genomes are pre-sented withthecoding
sequence(plus strand) reading
fromleftto
right; thus,
the 5' endoftheplus
strandorthe 3' end of the minus strand is referred to as the left end. Thisconvention will be followed heretodiscuss the distribution
(the
proportion
of various components in apopulation)
ofterminal-sequence
orientations ofBPV DNA.Sequence
heterogenity
oftheleftendofBPVdemonstratedby
cloning
of terminalfragments.
Toobtain apopulation
of cloned terminalfragments,
reannealed double-strandedvi-rion DNA of BPV was used to generate clones of the left-terminal
(map
units 0through 17)
andright-terminal
(map
units92through
100)
EcoRIfragments
ofBPV.For theright
end,
clones with eitherflip
orflop
orientations werereadily
obtained. For the leftend,
most of the clonesobtained were in the
flip
conformation.Surprisingly,
clonep3'V8
containing
a34-base-pair (bp)
deletionwas found tobe in the
flop
conformationatthis terminus. This conforma-tion hadnot been observed from otherautonomous parvo-virus genomes.Sinceasequenceinversion withintheleft terminus ofBPV could occur
during
cloning
as anartifact(27),
itwasneces-sarytoconfirm thepresence ofthe
flop
conformation in the native DNAby
other means. Theunique
SmaIrecognition
sequence
(CCC/GGG)
ofBPVlies withinanunpaired region
of the left
hairpin (Fig.
1A). Hence,
the enzyme will cutdouble-stranded
DNA at either 44 or 106 nucleotides(nt),
depending
on whetherthe DNA is in theflip
orflop
form. Theability
to clone a left terminus of 106 nt from thedouble-stranded virion DNA after
digestion
with SmaIwould
provide
direct evidence for the existence of theflop
conformationatthe left endofboth strands of DNA. Whenaseriesofterminal
SinaI
clonesweresequenced,
onewitha96-bp
insert was found. Thisclone,
M13Smal99,
had theexpected
sequence for theflop
conformation but with a10-bp
deletion at the terminus.Thisfinding
provided
directproof,
by cloning,
of the existence of theflop
conformationattheleft endofboththe
plus
andminusstrands. Sequence inversion duringcloning
could notgive
rise to a 96-bpfragment
froma44-bp fragment.
Demonstration ofboth
flip
andflop
conformationsatthe left(3')
endof
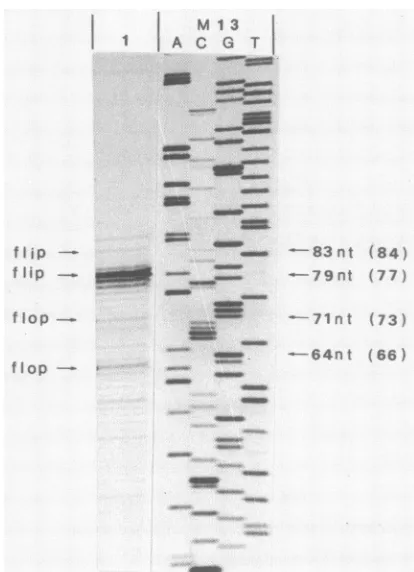
the minus-strandDNAby end-labelanalysis. The occurrenceoftheflip
orflop
formscanalsobevisualizedandthe relative amounts ofeach can be assessed by end-label
analysis
asdescribed inMaterials
and Methods. On thebasisof the sequence data
(12),
the left end of the minusstrandofBPVDNAcanexist inamoststable
(maximum
basepairing)Y-shaped configuration (Fig.
1A).
There is aunique
HhaI (GCG/C) site inoneof itsarms. Endlabeling(whichaddedaddA basetothe3'-hydroxylend and made the
fragment
onebaselonger)of the minusstrand,followedby digestionwith
HhaI, shouldyieldbands of 77 and 66 nt,correspotidingto
theflipandflop conformations, respectively.
Whenthe single-stranded BPV DNA(minuspolarity
only)
was analyzed in this way, bands at 79 and 64 nt, which corresponded well with the expected lengths, were ob-served, along with a number ofsecondary bands (Fig.
2).
The band corresponding to theflip conformation at the 3' terminus was about 10-fold more intense than that
corre-sponding to theflop conformationatthe 3' terminus. There-fore, roughly90% of the minus strands had the flip orienta-tion, and only 10% had the flop orientation at the left end. HhaI tends to make single-strand cuts within the
duplex
region containing the recognition site (3). Weaker bands in the end-label analysis (Fig. 2), at nt 71 and 83, agreedwell withthe expected values of 73 and 84predictedfor
cleavage
atthe distalsides of theHhaI sites(Fig. 1A).Bandsat77,
78,
and80nt(Fig. 2)werethe result of variations in the
position
of the terminal nucleotide at the left end, as reported for other parvoviruses (3, 15). The discrepancies between the expected terminal-fragment lengths and the observed ones
aredue, at least in part, to the effect ofsecondarystructure
on themigration of thesefragmentsinasequencing
gel.
Equalfrequencies of flip and flop forms at the right(5')end ofthe minus strand.The conformation of theright (5')end of the minusstrand can be demonstratedby end-labelanalysis
of invitro-replicated minus strand. In vitro replication would yield a double-stranded DNA whose conformation at the right end is defined by the conformation of the minus strand. NdeI(CA/TATG) digestion would cut the invitro-replicated
BPVDNA at nt 5437 inthe flip orientation and at nt 5422 in the flop orientation (12). Replicated BPV virion DNAwas
thus analyzed, and two bands of 55 and 73 nt, with approx-imately equal intensities, were observed (12).
Theright terminus of BPV was previously assignedto nt
5491 on the basis of the data mentioned above and the sequenceanalysis of cloned Si nuclease-resistantfragments
ofsingle-stranded virion DNA(12). However, in vitro rep-lication of palindromic DNA by Escherichia coli DNA
polymerase I Klenow fragment is often incomplete and arrests at specific sites that are capable of forming hairpin
structures,aspreviously reported by Cotmoreand Tattersall
(13)and asdescribedbelow. Duringcloning both of theright
terminus of BPV and of infectious genomic clones from
double-stranded virion DNA (27), several of the clones obtained had therightterminus 26bp beyond thepreviously assignedterminus ofthe genome. Also, one clone of theSi
nuclease-resistant right terminus containing the same 26 additionalbp wasfound(K. C. Chen, unpublishedresults). In
addition,
the fragment lengths of end-labeled,NdeI-digesteddouble-stranded virion DNA were found to be 20 to 22bp longerthan thoseobtainedfrom the in vitro-replicated DNA(seebelow). On thebasisof this evidence, the revised terminus of BPV isplacedatnt 5517 (Fig. 1B).
Even thoughthe terminal fragments of NdeI-digested in
vitro-replicated minus-strandDNA were truncated because of
incomplete
replication,thebandintensitiesindicatedthat both theflipandflopforms occur with equal frequencies at theright
end ofthe minus-strand virion DNA, as demon-stratedfor otherparvoviruses (1, 17, 21, 24). This result isexpectedifthehairpintransfer mechanism were involved in the synthesis of minus strand with the plus strand as the
templateand its right-endpalindrome as the self-primer.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
(A) Left (3') terminus (1) Flip conformation
A 80-A G
G C,_ C G HhaI
G C 100 120 140 160
C G
IIII
G CGCGTAACA'CCGGATTAGCCCGTCGGTTATACCACCTACGTCCTTTACTCATTATTTTTATATCTCGGACAC---5'
C
A TCGCATTATT-GGCC----CGGGCAGCCAATATGGTGGATGCAGGAAATGAGTAATAAAAATAT 3'
G C\
C G 60 SmaI 40 20 1
G C G C C G G A C
(2) Flop conformation G
C T G C C G C G
80-G C 100 SmaI 120 140 160
C G T
T AGCGTAATA-ACCGG----GCCCGTCGGTTATACCACCTACGTCCTTTACTCATTATTTTTATATCTCGGACAC- ' G
C GCGCATTGTAAGGCCTAATCGGGCAGCCAATATGGTGGATGCAGGAAATGAGTAATAAAAATAT 3'
C G 60 40 20 1
G C HhaI C G T C T
(B) Right (5') terminus
(1) Flip conformation
5341 5360 5380 5400 5420
3'--GAAATATAAAAATATGTGACTAAGTGAGAAAGGTGGTAGAAAATCAACATAAGAATCAATAGTTATTTCCGCGCTTCGCGGTATTTTTTA A 5'
fTATTTTTATACACTGATTCACTCTTTCCACCATCTTTTAGTTGTATTCTTAGTTATfAATAAAGGCGCGAAGCGCCIATATAT
5517 5500 5480 5460 5440
NdeI
(2) Flop conformation
5341 5360 5380 5400 5420 NdeI
II
I
I
I
A T T3'--GAAATATAAAAATATGTGACTAAGTGAGAAAGGTGGTAGAAAATCAACATAAGAATCAATAGTTATTTCCGCGCTTCGCGGTATACTCTA T
5'
ATATTTTTATACACGGATTCACTCTTTCCACCATCTTTTAGTTGTATTCTTAGTTATCAATAAAGGCGCGAAGCGCCATAAAAAAT
5517 5500 5480 5460 5440
FIG. 1. Nucleotidesequencesof theflipandflopconformationsatthe left(3')terminus(A)andtheright (5')terminus(B)oftheminus strandof BPV. RestrictionenzymerecognitionsequencesforHhaI,NdeI,andSmaIareindicated. Thepositionof the
previously
assignedright (5') terminus (*) is shown.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.90.534.82.655.2]3810 CHEN ET AL.
GTM1 3
|A C G T|
_~_
f jp -_
f
Ilip_--flop
flop
-83nt (84)
--79nt (77)
71n t
-64nt
(73 )
(66 )
FIG. 2. End-label analysis of the left terminus of the minus
strand of BPV. Single-stranded virion DNA was end labeled, digested with HhaI, and analyzed as described in Materials and
Methods. Lane 1 contains wild-type virion DNA. Bands corre-spondingtotheffipandfloporientations areindicatedontheleft.
The observed and the expected (shown in parentheses) sizes are indicatedontheright.
Demonstration ofsequence inversions at both ends of the
plus strand by end-label analysis of double-stranded virion
DNA. The distribution ofterminal-sequence orientations in
theplus-strand virion DNAcanbededuced from the analy-sis of double-stranded (reannealed plus and minus strands) virionDNA, provided thatthedistributionwithin the minus strandis known, asillustrated below.
Reannealing ofBPV virion DNAplus and minus strands
would result indouble-stranded molecules whichare either
fullybasepaired (in the fliporflop conformations)orpartly
unpaired in terminal regions because of various
terminal-sequence orientations. The former would be cut by the diagnostic restriction enzyme and produce fragments char-acteristic of the sequence orientation, but the latter would
not. Forarestrictionenzymetogenerate afragment witha
given conformation, either flip (i) or flop(o), both plus (c)
andminus (v) strands in the double-stranded (d) DNA have
to be inthesame orientation. Theprobability (P) of cutting
atsuchaconformation is the product of theprobabilities that eachstrand is inthatconformation,asgiven byPd(i)=
PC(i)
x P,(i) andPd/o) = Pc(o) x Pv(o), wherePd(i) denotesthe
probability of double-stranded DNA being in the flip orien-tation. The ratio of flip to flop observed in the double-stranded DNA is then the product of the ratios for the individual strands,
Pd(i)/Pd(o) = [P,(i)/P,(o)] X
[PA(i)IPA(o)]
(1)Endlabeling and SmaI digestion ofdouble-stranded,
rean-nealed virion DNAfrom wild-type virus gaveamajor band
at 42 ntand a minor band at 103 nt(Fig. 3A, lane 1), which
corresponded well with the expected 45 and 107 nt for the flip and flop conformations at the left end, respectively. Bands at 40, 41, and 43 nt were the result of terminal
variations of the leftend,as observedwith HhaI digestion. Theexistence ofabandcorrespondingtotheflop
conforma-tion ofthe left end from double-stranded DNA provided
additional evidence for its presence in the minus strand. The relative intensities indicated that the flip conformation was predominant at the left end. The flip-to-flop ratio in double-stranded, reannealed virion DNA was about the same as that
observedin theminus strand after digestion with HhaI(Fig. 2). Sincethe two ratios were about thesame, this suggested, from equation 1, that theratio for the plus strand mustbe closeto 1.Thatis, for the virion DNA thatwasencapsidated
as plusstrand, half of itwasin theflip orientationatits left
end, and the other half was in the flop orientation. This
would be the result expected if plus strands were synthe-sized bythehairpin transfer mechanism with minus strandas thetemplate and its 3'hairpin asthe self-primer.
Similaranalysis oftherightendofdouble-stranded virion
DNAwith NdeIasthediagnosticenzymeshowedtwobands of77 and 93 nt (Fig. 3B, lane 1). With the reassigned right end of BPV, theexpected sizes of theflip and
flop
fragmentswould be 81 and 96 nt(Fig. 1B),whichwere in reasonable
agreement with the observed values. The flop band was
about twice as intense as the flip band. Since the minus strand had equal proportions of flip toflop atthis end, the ratio observed in the double-strandedDNA mustresult from
the flip-to-flop ratio in the plus strands (see equation 1).
Hence, for the plus strand, roughly one-third of itwas
flip
andtwo-thirds of it was flopatthe right end.
Terminal structures of BPV RF DNA. In vivo RF DNA (mostly of monomer length) (22) was isolated from BPV-infected cells 20 to 24 hpostinfection byamodification of the Hirt procedure (29). This DNA was subjected to end-label
analysis by usingSmaI for the left end and NdeIfortheright end. The restriction fragments from these reactions were
comparedtothoseobtainedbydigesting
reannealed,
double-stranded virion DNA with the same enzymes
(Fig.
3A andB).The ratio offliptoflopateach terminuswasthesamefor
both RF and double-stranded virion DNA. Selective
encap-sidation could not be responsible for the observed ratios of terminal-sequence orientations in virion DNA, because the same ratios of flip to flop forms were observed in the RF
population. Furthermore, the sizes of terminal NdeI
frag-ments obtained from the two DNAs were the same. There
was noextensionof the right endofBPV RF DNAcompared withthe encapsidated virion DNA (1), incontrast to MVM. Fate offlip and flop forms of transfecting genomic clones.
Full-length genomic clones pVT501 (flop at left end) and
pVT502(flip atleft end) have the same right end (in the flip form) and differ only in the orientation at the left end (27). When the left ends ofthe progeny DNA of amplified
trans-fections initiated with either pVT501 or pVT502 were ana-lyzed byend labeling and SmaIdigestion, bothflip andflop conformations wereobserved and theratio ofthe twoforms
wasidentical to that found in a normal infection (Fig. 4). This suggested that, at the left end, one conformation could generate the otherand that the flip form always accumulated preferentially in the viralDNA, regardless of the conforma-tion present in the clone used for transfection. Similar
analyses were performed on a different infectious genomic
clone, pGCSma2O (flipatboth ends [27]), which is deletedby
3 bases at the left end and 35 bases at the right end. Both ends wererepaired towild-type lengthand againtheratio of
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.73.280.71.357.2]TERMINAL-SEQUENCE ORIENTATIONS OF BPV DNA
A LEFT END
B
RIGHT END__ _
_R
.v_
_5
_ _
__
_ _
_!! __
_
_ _
[image:5.612.75.552.76.406.2]
-..S.
...
-_ _
-_,.,>..
= _
4 _
_w _
_ _
----103 nt ( 107)
flip
ml S1
1 A C G T
am
--fo
cow -..V_
M .M
,. _
000
-93 nt (96)
77nt (81)
- 42 nt (45)
f p
FIG. 3. End-label analyses of double-strandedvirion DNA and in vivoRFDNA.Reannealedplus- and minus-strandvirion DNA (lane 1)
and RFDNA(lane2)wereendlabeledanddigested either with SmaI for analysis ofsequenceinversionsatthe leftend (A)orwithNdeI for
analysisofsequenceinversionsatthe right end (B). The positions and observed and expected (shown inparentheses)sizes ofthe flip andflop
conformationsareindicated.
fliptoflopateach terminus wasindistinguishable from that for wild-type virion DNA (data not shown). The same
phenomenon has been reported forAAV (26) andthe right
terminus ofMVM (cited in reference 14).
DISCUSSION
Thedistribution ofterminal-sequence orientations of BPV virion DNA was analyzed by end labeling and restriction enzyme digestion. The results demonstrated specific ratios offlipandflop conformationsatboth endsfor DNAofeither
plusorminuspolarity.Incontrast,sequenceinversionatthe
left end of the minus strand was absent in all the rodent
autonomousparvoviruses studiedto date (14).
Thepresenceofapproximately equalamountsofflipand
flopforms attherightendsof the minus strands, aswellas at the left ends of the plus strands, suggests that hairpin transferprocesses occurforsynthesisof BPV virion DNA of eitherpolarity.Ineithercase,thecomplementtotheplusor minus strand would serve as the template and self-primer from its 3'-terminalpalindrome.
Transfection with genomic clones of BPV containing
definedconformations atthe endsresulted inprogenyvirion
DNAs with distributions identicaltothatfound inanormal infection. The same ratio ofplus tominus strands (27), as well as the same ratios offlip to flopforms atthe ends of
reannealed virionDNAs (Fig. 4),were observed. This indi-cated that there must be inherent constraints on DNA
replication fortheestablishment of such aspecific distribu-tion. Evidence exists thatbothcellular andviral factors are
involved intheseprocesses. Both H-1 and LuIll can infect NBEcells;however, H-1 encapsidates predominantly minus strands, whereas LuIlI encapsidates equalamountsof plus
and minus strands (5, 19, 20). The cellular influence is
indicatedbythe observation that H-1 encapsidates various
amounts ofplus strand when propagated in different cell
types (S. Rhode, personal communication).
The specificity of the distribution of terminal-sequence
orientations in virion DNA isdeterminedatthe RFstageand
not atthe encapsidation stage(Fig. 3). Thepresenceofthe sameratios offlip to flopforms in RF DNA and in double-stranded virion DNA for BPVarguesagainst encapsidation
of a subpopulation of progeny DNAs. If this were the mechanism for selection, the ratios in reannealed virion DNAwould beexpectedtodiffersignificantlyfrom thosein RFs. The data indicated insteadthat the distribution ofRFs
with different terminal-sequence orientations isresponsible
for the observed distribution ofterminal-sequence orienta-tions in virion DNA. The sameconclusion was reachedfor MVM (1) from the observation that a unique sequence
orientationwasdetected at the left end of both RF andvirion DNA.
1
~~M
13 l. A C _
*Nw n
.~~~~~~-t am_
flop
VOL. 62,1988 3811
on November 10, 2019 by guest
http://jvi.asm.org/
3812 CHEN ET AL.
M tj
j,A-:s<
LLOP-t
-FIG. 4. Terminal-sequence orientations at theleft enyDNAresultingfrom amplified transfections with
pVT502. Double-stranded virion DNA waspurified I
turesinfectedwithprogenyvirions ofcellstransfected (lane 1),pVT502 (lane2),orwild-typeBPV(lane3). 1 endlabeledanddigestedwith SmaI.
However, themechanisms forestablishingspe progenyDNAdistributions currently proposed
and MVMreplication (1, 14)are notadequatetc
datafor BPV. The replication mechanism for) allowshairpin transfer withequal efficiencyatb
would predict equal distribution in both RFs
DNAs.
Themodifiedrollinghairpinmodel (1, 14),po:
general model for autonomous parvovirus repli posedamechanismforselectivegeneration ofce
accountforthe distribution of virion DNA inM model,twoobligatory dimerintermediates and
nicks and ligations generate, in equal propc monomerRFswhoseleftendsareinthesameoi thatof theinput virionDNA. ThetwoRFsinterc
onetotheotherwhilegenerating minus strands displacement synthesis onplus-strandtemplate: theconstraints that restrict these RFsto minus thesis werenotindicated.
We have observed a complex distributior strandedBPVDNA. All eightpossible configura
flopateachendof plus orminus strands) occu amounts.Consequently, all fourmonomerRFs(I either end) must also be present, in unequal,
serve as templates forprogenyDNA synthesis.
butionofRFs mustbeestablishedandmaintain
atetheparticular distribution of virion DNAob
To adapt the modified rolling hairpin model to our
obser-vations with BPV, we might propose that the site-specific
nickase-ligase would act on the progeny minus
strand,
aswell as the parental minus strand, of thedimerintermediate.
In this way, it would be possible to generate all four
monomer RFs in the distribution observed for BPV by
postulating certain affinities of the nickase-ligase for these - 0 3 n sites. Butwhenthesetemplate RFs areused for the
synthe-sis of both plus and minus strands (as required for
BPV),
their characteristic asymmetric distribution will be altered,
because thetemplate RFschange fromoneformto
another,
depending on the type of progeny DNA
synthesized.
For example, progenyplus-strand synthesis wouldcausethe left endof parental RF tochangefromfliptoflop orvice versa.If plus- andminus-strandsyntheses were equally likely, then eventually a uniform distribution ofRFswould be reached.
More likely, plus and minus strands are synthesized with different efficiencies, inwhichcasetheasymmetric
distribu-tion of RFs depends on these efficiencies as well. Thus it
seems that the characteristic distribution ofDNA forms is determined in a complicated fashion by the rates of all the steps in the replication process.
Rather than adapt the modified rolling hairpin model, we have developed a unified kinetic model forparvovirus rep-lication (K. C. Chen, J. J. Tyson, M.Lederman,E. R.Stout,
4 . andR. C.
Bates,
manuscript
inpreparation;
J. J.Tyson
andK. C. Chen,manuscriptin preparation) based on the simpler AAVreplication model (8), in which RF and progeny DNAs are synthesized via hairpin transfer. In our model, we assume that the rates ofsynthesis of these DNAforms may bedifferent dependingonthe flip or flopconformationatthe 3' terminus of the template. Dimer RF molecules were end of prog- included as possible intermediates in the generation of pVT501 and monomer RFs. At some point in the infectious cycle, the from cell cul- various forms of RF and progeny DNAs would reach
steady-with
pVT501
statedistributions,based on their rates of synthesis and ratesrhe
DNAwasof
conversionto otherforms by hairpintransfer. These rateconstants would be characteristic for each parvovirus and
could be modulated bycellular or viral factors or both.
cificRFand For example, suppose that the rates of RF and progeny
forAAV (8) DNA synthesis were the same regardless of the
conforma-)explainthe tion at the ends of thetemplate. Then a uniform distribution kAV, which of RF and virion DNAwould beestablished,asobserved for )othtermini, AAV. To account for BPV distribution, suppose that the and virion rates of synthesis with the flip form at the left terminus were very slow compared with the rates with the flop form at the stulated as a leftterminusor either form atthe rightterminus. Then most ication, pro- ofthe RF would be in theflip form at the left terminus mainly -rtain RFs to because these RFs with the flipconformationat the left end [VM. In that would beused lessfrequentlyastemplates and remain in the
site-specific flip form. Since the left terminus would be used as the primer )rtions, two andtemplateforplus-strand synthesisand the rightterminus rientation as would be used for minus-strand synthesis, mostly minus -onvertfrom strands with the flip form at the left end would be produced byrepetitive from these RFs as a result of the different rates of synthesis s. However, from the ends. This would be the case for BPV, and if the ,-strand syn- rate were even slower for utilization of the flip form of the left terminus, the distribution seen with MVM could be of single- obtained. Withthis kinetic model, the observed virion DNA ttions(flip or and RF DNA distributions of all parvoviruses studied to r inunequal date, including LuIl (N. Diffoot and B. Shull,unpublished
fliporflop at results), can beaccounted for by assumingdifferent sets of amounts, to rate constants for synthesis of RF and progeny DNA with
This distri- differentprimers-templates.
iedtogener- In the rolling hairpin model, dimer intermediates are )served. obligatory and must be processed by asequenceofnicking
J. VIROL.
vmiu
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.63.298.73.393.2]andligationstepsin ordertogeneratetwomonomer RFs(of MVM) withauniqueconformationatthe left end. However, dimerintermediates may not be obligatory for BPV replica-tion. The four monomer RFs in BPV theoretically can be
generatedfrom any given form of input virion DNA, withor
without a dimer intermediate, by the hairpin transfer pro-cesses. In vivo RF isolated from BPV-infected cells was found tobe predominantly ofmonomer length (22), unlike the large proportion of dimers observed for MVM (32). In
ourkinetic model, dimer RFs are possibleintermediatesbut arenotnecessaryin order to generate aspecificdistribution ofRFs and virion DNA forms. Ifdimer intermediates are formed, they can be processed to two monomers by
stag-gerednickings and repair of both strands.
The observation oftwoalternative orientations at theleft terminus of the minus-strand DNA of the autonomous par-vovirus BPV shows greater diversity in the replication strategy of these viruses than had previously been sus-pected. To account for this diversity, we suggest that the relative abundance of the various viral DNA species are determined by the rate constants for synthesis of RF and progeny DNA from various templates. A more detailed description of this kinetic model forparvovirusDNA repli-cationis inpreparation.
ACKNOWLEDGMENTS
We thank J. J. Tyson for helpful discussions and for critical review of themanuscript.
This researchwassupported by American CancerSocietygrant
MV-220.
LITERATURE CITED
1. Astell, C.R., M. B. Chow, and D. C.Ward. 1985. Sequence analysis of the termini of virionandreplicative forms of minute virus of miceDNA suggests amodifiedrollinghairpin modelfor
autonomousparvovirusDNAreplication.J. Virol.54:171-177. 2. Asteli, C. R., E. M. Gardiner, and P. Tattersall. 1986. DNA sequenceof thelymphotropicvariant of minute virus ofmice, MVM(i), and comparison with theDNA sequenceof the fibro-tropicprototypestrain. J.Virol. 57:656-669.
3. Asteli, C.R., M. Smith,M. B. Chow,and D. C. Ward. 1979. Structure of the 3' hairpin termini of four rodent parvovirus
genomes: nucleotide sequence homology at origins of DNA
replication. Cell 17:691-703.
4. Astell, C. R.,M.Thompson,M.Merchlinsky,and D.C. Ward.
1983. ThecompleteDNA sequenceofminute virus ofmice,an
autonomousparvovirus. Nucleic AcidsRes. 11:999-1018. 5. Bates,R.C., C.E.Synder,P. T.Banerjee,andS.Mitra. 1984.
Autonomous parvovirus Lulll encapsidates equalamounts of plus and minus DNA strands.J. Virol. 49:319-324.
6. Berns,K.I., and R. A.Bohenzky.1987. Adeno-associatedvirus:
anupdate. Adv. Vir. Res. 32:243-306.
7. Berms, K. I., and W. W. Hauswirth. 1982. Organization and
replicationofparvovirusDNA,p. 3-35.InA.S. Kaplan (ed.), Organization andreplication of viral DNA. CRC Press, Inc.,
BocaRaton, Fla.
8. Bems, K. I., and W. W. Hauswirth. 1984. Adeno-associated virusDNA structure andreplication, p. 1-31. In K. I. Berns
(ed.), Theparvoviruses. PlenumPublishingCorp.,NewYork.
9. Berms,K.I.,and M. Labow.1987.Parvovirusgeneregulation.J.
Gen. Virol. 68:601-614.
10. Biggin, M. D., T. J. Gibson, and G. F. Hong. 1983. Buffer
gradient gels and 35S label as anaid to rapid DNA sequence
determination. Proc. Natl. Acad. Sci. USA 80:3963-3965.
11. Cavalier-Smith,T.1974. Palindromic basesequencesand
repli-cation ofeucaryotic chromosome ends. Nature (London)240: 467-470.
12. Chen, K. C., B. C. Shull, E. A. Moses, M. Lederman, E. R.
Stout,and R. C. Bates.1986.Completenucleotide sequence and genome organization of bovine parvovirus. J. Virol. 60:1085-1097.
13. Cotmore, S. F.,and P. Tattersall. 1984. Characterization and molecularcloning ofahumanparvovirus genome. Science226: 1161-1165.
14. Cotmore, S. F., and P. Tattersall. 1987. The autonomously
replicating parvoviruses of vertebrates. Adv. Vir. Res.
33:91-174.
15. Fife, K. H., K. I. Berns, and K.Murray. 1977. Structure and nucleotide sequence of the terminal regions of adeno-associated virusDNA.Virology 78:475-487.
16. Hauswirth, W. M. 1984. Autonomous parvovirus DNA
struc-ture and replication, p. 129-152. In K. I. Berns (ed.), The parvoviruses. Plenum Publishing Corp.,NewYork.
17. Lusby, E., K. H. Fife, and K. I. Berns. 1980. Nucleotide sequenceof the inverted terminalrepetition in adeno-associated virus DNA. J. Virol. 34:402-409.
18. Maniatis, T.,E. F.Fritsch, andJ.Sambrook. 1982. Molecular
cloning: alaboratory manual. ColdSpringHarborLaboratory,
ColdSpringHarbor, N.Y.
19. Muller, D.-E., and G. Siegl. 1983. Maturation of parvovirus LuIll in asubcellular system. I.Optimalconditions for in vitro
synthesis and encapsidation of viral DNA. J. Gen. Virol. 64: 1043-1054.
20. Rhode, S. L. 1978. H-1 DNA synthesis, p. 279-296. In D. C. WardandP.Tattersall(ed.),Replicationofmammalian parvo-viruses. Cold SpringHarborLaboratory, Cold SpringHarbor,
N.Y.
21. Rhode, S. L., and B.Klaassen. 1982. DNAsequence of the 5'
terminuscontaining the replication origin of parvovirus replica-tive formDNA. J.Virol. 41:990-999.
22. Robertson, A. T., E. R. Stout, and R. C. Bates. 1984.Aphidicolin inhibition of the production of replicative-form DNA during
bovineparvovirus infection.J. Virol.49:652-657.
23. Roychoudhury, R., and R. Wu. 1980. Terminal transferase-catalyzed addition of nucleotides to the 3' termini of DNA. Methods Enzymol. 65:43-62.
24. Sahli, R., G. K.McMaster, and B. Hirt. 1985. DNAsequence comparison between twotissue-specific variants ofthe autono-mousparvovirus, minutevirusof mice. NucleicAcidsRes. 13:
3617-3633.
25. Samulski, R. J., K. I. Berns, M. Tan, and N. Muzyczka. 1982. Cloningofadeno-associatedvirus intopBR322: rescue of intact virusfrom therecombinant plasmid in human cells. Proc.Natl. Acad.Sci. USA79:2077-2081.
26. Samulski, R. J., A. Srivastava, K.I. Berns, and N. Muzyczka.
1983. Rescueofadeno-associated virus from recombinant plas-mids:gene correction within the terminalrepeatsofAAV.Cell 33:135-143.
27. Shull, B. C., K. C. Chen, M.Lederman,E.R.Stout, and R. C. Bates. 1988. Genomic clones of bovineparvovirus: construction
and effect of deletions and terminal sequence inversions on
infectivity. J. Virol. 62:417-426.
28. Siegl, G., R. C. Bates, K. I. Berns, B. J. Carter, D. C.Kelly, E. Kurstak, and P. Tattersall. 1985. Characteristics and taxonomy
ofparvoviridae. Intervirology23:61-73.
29. Siegl, G., and M. Gautschi. 1978.Purification andproperties of replicative-form andreplicative-intermediateDNAmolecules of
parvovirus LuIII, p. 315-325. In D.C. Ward and P. Tattersall (ed.), Replication of mammalian parvoviruses. Cold Spring HarborLaboratory, Cold SpringHarbor, N.Y.
30. Straus, S. E., E. D.Sebring,andJ.A. Rose. 1976.Concatemers of alternating plus and minus strands are intermediates in adenovirus-associated virus DNA synthesis. Proc. Natl. Acad.
Sci. USA73:742-746.
31. Tattersall, P., and D. C. Ward. 1976. Rolling hairpin model for replication ofparvovirusand linearchromosomal DNA. Nature (London) 263:106-109.
32. Ward, D. C., and D. K.Dadachanji.1978.Replicationofminute virusof mice DNA, p. 297-313. In D.C. Ward andP.Tattersall
(ed.), Replication of mammalian parvoviruses. Cold Spring
HarborLaboratory, Cold Spring Harbor, N.Y.