Copyright© 1992, American Society for Microbiology
Gag Proteins of the Highly Replicative
MN
Strain of
Human
Immunodeficiency Virus Type 1: Posttranslational Modifications,
Proteolytic Processings, and Complete Amino Acid Sequences
LOUIS E.HENDERSON,1* MICHELLE A. BOWERS,' RAYMOND C. SOWDER II,' STEFAN A.SERABYN,1
DONALD G. JOHNSON,' JULIAN W. BESS, JR.,' LARRY0. ARTHUR,' DUNCAN K. BRYANT,2
ANDCATHERINE FENSELAU2
AIDS Vaccine Program, Program Resources, IncorporatedlDynCorp, National CancerInstitute-Frederick Cancer Research and Development Center,
Frederick,
Maryland21702-1201,1 andDepartmentof Chemistryand Biochemistry, University of Maryland, Baltimore County, Baltimore, Maryland 212282 Received 3 September 1991/Accepted 17December 1991
The MN strain of human immunodeficiency virus type 1 was grown in H9 cells, concentrated by
centrifugation, and disrupted, and proteinswerepurified byreversed-phase high-pressure liquid
chromatog-raphy. Complete amino acid sequences were determined for the mature Gag proteins, showing natural proteolytic cleavage sitesand the orderof proteins (pl7-p24-p2-p7-pl-p6)in the Gagprecursors.At leasttwo
sequencevariants ofp24 and eightsequencevariants ofp17weredetected. Thetwo mostabundant variantsof
p24 andp17 representedatleast 50%Yo 5% and20% ± 5%oftheirtotals, respectively. These datasuggest
heterogeneityin thevirus population,with 50% of the total viruscontainingthemostabundantforms of p17
and p24 and 20% of the virus containing the second most abundant forms. The Gag precursors ofthese suggested viruses differ from each otherby only3amino acid residuesbut differ from theprecursorspredicted
by the published MN proviral DNA sequence by 10 residues. Electrospray ionization mass spectrometry
analysisof the purified p24forms showed that the measured molecularweightof theproteinwas200 ± 50
atomic mass units greater than the calculated molecularweight. The source ofadditional massfor the p24 forms was not determined, but the observation is consistent with previous suggestions that the protein is phosphorylated. Greater than 98% of the total recovered p17 was myristylated at the N-terminal glycine
residue,and the measured molecularweights (as determined byelectrosprayionizationmass spectrometry)of themostabundant formswerewithin 3atomic mass units of the calculated molecularweights (15,266). Human immunodeficiency virus type 1 (HIV-1) is the
etiologic agent ofAIDS (1, 25, 29), and the MN strain of HIV-1 (HIV-lMN) (13) has been shown to be a prevalent
prototypeinthe U.S.population,bothbyserologicalstudies (7)andbyvirusisolation andsequencingofthe V3regionof theenv gene(23). Because of itsprevalence, HIV-lMNhas beensuggestedastheprototypevirus for thedevelopmentof vaccines and other antiviral strategies. Standardized stocks of this strain will be neededaschallengereagentsfortesting the effectiveness of various developed procedures; it is highly desirable that these stocks be well characterized. Challenge stocks ofHIV-lMNgrown in H9 cells have been prepared andarebeingevaluatedat thebiologicallevel. At themolecularlevel,atleastonecompleteinfectiousprovirus
has been isolated from cells infected withHIV-1MN, and its nucleotide sequence has been determined (28). However, infected cells generally contain multiple sites of proviral integration, and it is difficult to determine which of the integrated proviruses produces the mostabundant virus in tissue cultures. One method for characterizing the most abundant virus istoisolate viralproteinsand determine their amino acidsequences.
The HIV-1viralgenomecontains threelargeopenreading
frames, designatedgag,pol,andenv,that code forproteins
ultimatelydestined forincorporation into thematurevirus. Gagand Polproducts togetherwith RNAmakeup the viral core, and Envproducts arepart of the lipid envelope. Pol
*Corresponding author.
proteins are enzymes necessary for viral replication. Gag
proteins are often referred to as structural proteins and
representamajor portionof thecorestructure.Theproteins analyzedhere areproductsofthegag genepurified directly fromHIV-lMNgrownin tissue cultures. Theprimary trans-lationalproductof thegag gene isapolyproteinprecursor,
designated Pr55Fag, that is ultimately cleaved by the viral protease tothematureGag proteinsfound in the virus. Here weshow thatPr559agofHIV-1MNis cleavedtosixproducts, including the matrix antigen (MA), core antigen (CA),
nu-cleocapsid (NC) protein,and threepeptides, designatedpl, p2,andp6,and the order oftheproductsin theprecursoris MA-CA-p2-NC-pl-p6, aspreviously suggested (15, 18).We reporton the purification andcomplete chemical structural analysisof the sixmatureGag proteins purifieddirectlyfrom HIV-1MNviralparticles.
MATERIALS AND METHODS
Viruspreparation.H9cellschronicallyinfected with
HIV-1MNwere
provided by
R. C. Gallo(Laboratory
of TumorCellBiology, Division ofCancerEtiology,NationalCancer Institute, Bethesda, Md.) and were grown in roller bottles
underbiosafetylevel 3laboratoryconditionsatthe National Cancer Institute-Frederick Cancer Research and
Develop-ment Center, Frederick, Md. Virus production was in-creasedapproximately50-foldby supplementinginfectedH9 cells withanequalnumber of uninfectedH9cellsafter each harvest (2). Virus was harvested from the culture medium
every 3 to 4 days, isolated by continuous-flow sucrose
1856
on November 9, 2019 by guest
http://jvi.asm.org/
(saturated at room temperature), the pH was adjusted to 8.5 by the addition of concentrated Tris hydrochloride buffer,
and 2-mercaptoethanolwas added to a final concentration of
2% (vol/vol). The resulting clear solution was transferred to a clean sterile tube and removed from the biosafety level 3 environment.
Protein and peptide purification. All samples for reversed-phasehigh-pressure liquid chromatography (rp-HPLC) were dissolved in saturated guanidine hydrochloride containing 2%2-mercaptoethanol at pH 8.5 and sonicated at
50°C
for 15 min before injection into the chromatographic column. Sep-arations were performed on,uBondapak C18 (Waters Asso-ciates) rp-HPLC supports by use of a liquid chromatograph (Pharmacia LKB) equipped with a rapid spectral detector (model 2140). Elutions were accomplished with 0.05% (vol/vol) trifluoroacetic acid at pH 2 and with a gradient of increasing acetonitrile concentrations. Eluted proteins and peptides were detected by UV absorption at 206, 280, and 294 nm and collected, and the solvents were removed by lyophilization.
Endoproteinase digestion. Proteins to be digested were dissolved in 50 mM NH4HCO3 at pH 8 to a final concentra-tion of 1 mg/ml. When a protein was initially insoluble, the pH was raised to 11 (concentrated NaOH) to solubilize the protein and thenadjusted to 8 to 9 (concentratedHCl) before addition of the enzyme and buffer. Digestions were initiated by adding theendoproteinase to a final concentration of 0.02 mg/ml and were continued for 18 h at room temperature. When a precipitate developed during the course of the digestion, the process was repeated for a second 18-h period. Digestions were stopped by adding concentrated hydrochlo-ric acid to a final pH of 2, and the peptides were separated by rp-HPLC on ,uBondapak C18 columns (3.9 by 300 mm). The endoproteinases used were endoproteinase Glu-C, endopro-teinase Lys-C, and trypsin (Boehringer, Mannheim, Germa-ny).
Amino acidanalysis. Approximately 1 ,ug of purified pro-tein or peptide washydrolyzed in 200,ulof 6 M hydrochloric acidcontaining 0.1% phenol in vacuo at 110°Cfor 24 h. The samples were dried in vacuo and analyzed on a Beckman model 6300 automated amino acid analyzer in accordance with themanufacturer's recommendations.
Gel electrophoresis. Sodium dodecyl sulfate-polyacryla-mide gel electrophoresis (SDS-PAGE) was performed as
previously described (22).
Edmandegradation. Automatic N-terminal Edman degra-dationswereperformed onapproximately 1-nmol samples of purified protein or peptide by use of a pulsed, liquid-phase protein sequencer (model 477A; Applied Biosystems Inc.) equipped with an on-line phenylthiohydantoin analyzer (model 120A) inaccordance with themanufacturer's recom-mendations.
FAB mass
spectrometry.
Fast atom bombardment (FAB) massspectrometry of Gag proteins was performed by use of the firsttwosectors of anHX110/HX110massspectrometer (JEOL, Tokyo, Japan) at an accelerating voltage of 10 kV,tween the two mass spectrometers (4), with helium used as the collision gas at a pressure sufficient to attenuate the precursor ion beam by80%. The collision cell was floated at 4kV. The collision-induced dissociation spectra were re-corded at a resolution of 1:1,000 and with 100-Hz filtering. For both scanning modes, 50 to 100 pmol of peptide in 0.1% trifluoroacetic acid (1
p,l)
and 3-nitrobenzyl alcohol (1p,l)
were used for the analysis.The computer program RESIDUES, written by David Heller (Middle Atlantic Mass Spectrometry Center, Johns Hopkins University, Baltimore, Md.), was used to assist with the interpretation of tandem mass spectrometry data, while the computer program MacProMass (written by Terry D. Lee and Sunil Vemuri, Beckman Research Institute, City of Hope, Duarte, Calif.) was used to identifyp17 proteolysis peptides in the conventional magnetic scans.
ESI-MS.Electrospray ionization mass spectrometry (ESI-MS) was performed with a Vestec electrospray source fitted to a Hewlett-Packard 5988A quadrupole mass spectrometer. Sample p17 (from pool A) was dissolved in an aqueous solution containing 1.5% acetic acid and 50% methanol. This solution was introduced into the source with a Harvard 22
syringe infusion pump.
RESULTS
Isolation of HIV.lMN proteins. As part of an ongoing program to characterize HIV-lMN and to prepare challenge stocks for future studies, the virus was produced in the AIDS Vaccine Program virus production facility at the National Cancer Institute-Frederick Cancer Research and Development Center by cocultivating infected and unin-fected H9 cells. Compared with tie amount of virus pro-duced from H9 cells chronically infected with
HIV-1MN,
this cocultivation method results in approximately a 50-fold increase in the amount of virus 7ecovered and has been adopted as a standard production method (2). Concentrated purified virus was disrupted and solubilized in saturated guanidine hydrochloride (pH 8.5) containing 2-mercaptoeth-anol, and the resulting clear solution was injected into a high-pressure liquid chromatograph for separation and puri-fication of the viral proteins. Figure 1 shows a typical chromatogram for the rp-HPLC separation of the proteinsassoci.ted
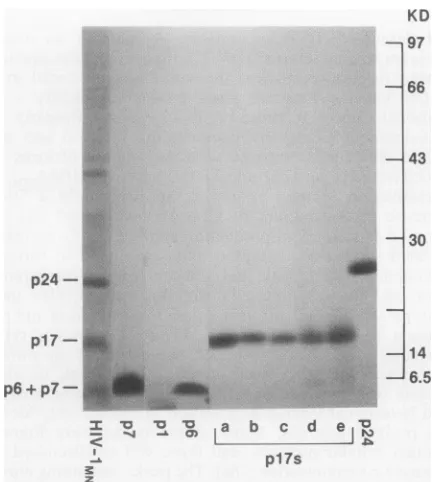
with the purified virus. Each peak of eluted protein was analyzed by SDS-PAGE, amino acid analysis, and N-terminal Edman degradation to qualitatively identify the purified proteins. Many of the peaks were found to contain cellular proteins, and these will be discussed in a separate communication (18a). The peaks containing purified HIV-lMN Gag proteins p17, p24, p2, p7, pl, and p6 are indicated inFi,,.
1. Protein in each of the labeled peaks (Fig. 1) was analyzed by SDS-PAGE, and the results for peaks p7, pl, p6,pl7athrough pl7e, and p24 are shown in Fig. 2. Peak pl7f was not included in this gel but yielded results indistin-guishable from those yielded by pl7e (data not shown); peak p2 yielded no detectable bands in the SDS-PAGE analysison November 9, 2019 by guest
http://jvi.asm.org/
E
C w
(0
0
0
400 800 1200 1600
Minutes (5.0 mllmin)
1923
FIG. 1. Protein separation by rp-HPLC. HIV-1MN virions were disrupted, and the viral proteins and peptides were separated by preparative rp-HPLC.Approximately175 mg ofpurifiedHIV-1MN wasdissolvedand reduced in 40 ml of saturated guanidine hydrochloride containing2% (vol/vol) 2-mercaptoethanolandinjectedintoa,uBondapak C18 column (19 by150mm). Proteinsandpeptideswereeluted with agradientofincreasingacetonitrile concentrations atpH 2.0(0.05% trifluoroaceticacid)and detected by UV absorption at 206 nm. The UV peaksassociated withpurifiedviralproteinspl7a throughp17f,p24, p2, p7, pl, and p6 are indicated. The pool of p24 taken for further analysis is indicatedby a bracket. OD, optical density.
p24- p17-p6+p7
-14 6.5
a b c d e
pl 7s
z
FIG. 2. Coomassie blue-stained gel of purified proteins.
HIV-1MNproteins purified as shown in Fig. 1 were analyzed by
SDS-PAGE on6 to 18% polyacrylamide gradient gelsand detected by staining with Coomassie brilliant blue R-250(18). Lane HIV-1MN
contained wholedisrupted virus,and lanesp7,pl, p6, p24,andpl7a through pl7econtainedapproximately3to5,ugofprotein purified
inthecorresponding peaklabeled inFig. 1.
(datanotshown).These results(Fig. 1 and 2) show that pl,
p2, p6, and p7 are eluted as highly purified proteins in well-separated and nearly symmetrical peaks. p24 is also elutedas ahighly purified protein,butthepeakis asymmet-rical. The trailing edge of the p24 peak contained
contami-nating proteins (visualized by SDS-PAGE analysisof
indi-vidual fractions) and was omitted from the pool of highly
purified p24 (bracketmarked Pool inFig. 1)taken for further
analysis. On the basis of UVabsorption,we estimated that
thepool containedatleast70% of the total p24 eluted in the
asymmetrical peak. In contrast, p17 was eluted from the
column as a group of partly separated peaks labeled a
through f, as shown in Fig. 1. The protein in each peak
migratedas asinglebandatabout 17 kDa in theSDS-PAGE
analysis(Fig. 2)andreacted withapolyclonalrabbit
antise-rumprepared against theprotein purifiedinpeakaand also
with a mouse monoclonal antibody to HIV-1 p17 (data not
shown). Of the total p17 eluted from the column, we
esti-mated that 66%was eluted inpeak pl7a, 5%waseluted in
peak pl7b, 9% was eluted in pl7c, 9%was eluted in peak
pl7d, 8%waseluted inpeakpl7e, and 3%waseluted inpeak
pl7f. Tobetter understand the natureof the apparent chro-matographic heterogeneityof the p17forms and to charac-terize purified pl, p2, p6, p7, and p24, we subjected the proteins to complete amino acid sequence analysis and examined them for posttranslational modification by mass
spectraland chemical methods.
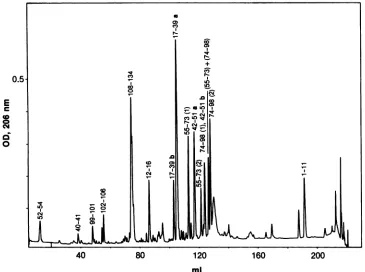
The protein in peak pl7a (Fig. 1) was digested with
endoproteinase Glu-C, and the peptides wereseparated by
rp-HPLC (Fig. 3). Each purified peptide was analyzed for amino acidcomposition andamino acid sequenceby N-ter-minal Edman degradation and mass spectrometry. The re-sults are summarized in Fig. 4, in which the determined
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.149.486.73.321.2] [image:3.612.82.298.419.660.2]E(
is~~~~~~~~~~~~~~~~~~~~~ICbJ
40 80I1201
o 0~~~~~~~~~~~~~~~c
I-co~~~~~~~~t
ejmIlLO 0
40 80 120 160 200
[image:4.612.123.488.74.346.2]ml
FIG. 3. Separationofpeptidesresultingfromendoproteinase Glu-Cdigestionofpl7a.Theproteinpurifiedinpeakpl7a(Fig.1 and2)was
digestedwithendoproteinaseGlu-C,andthepeptideswereseparated by rp-HPLC.Elutedpeptidesweredetectedby UV absorptionat206 nm.Thepeptidematerialin each UVpeakwasidentifiedbyaminoacidsequencing (see the text),andpeptidesarenumberedaccordingto thepositionsoftheirfirstand last residues in the amino acid sequencealignment showninFig.4.Peptidesrepresenting identical regions of p17(Fig. 4) butdifferingby amino acid substitutions aredistinguished by the lettersa and b (seethe text). Peptides containing cysteine residues(positions56 and86; Fig. 4)wereisolated intheir monomeric form,indicatedby(1),and intheirdisulfide form, indicated by (2). The peptide designated (55-73)+(74-98) is a heterologous disulfide dimer. When two peptides coeluted in the samepeak, both peptides are
indicated andseparated byacomma.OD, opticaldensity.
amino acid sequences for thepeptides arecomparedwitha
predicted amino acid sequence based on the nucleotide
sequence of codons 2through 135 of theHIV-1MN proviral
gag gene(28).Thealignmentshows that the determined and
predicted amino acid sequencesarein agreementfor 130 of
134 compared positions and prove that the protein in peak
pl7a(Fig. 1)is derived from the N-terminal end of the Gag
precursor. These data also show that the protein in peak
pi7a
is amixture ofat least twoclosely related p17forms that differ from each other by substitutions of valine forisoleucine at positions34 and 46.
The two forms ofp17 present in peak pi7a (Fig. 1) are revealed by the data in Fig. 3, which shows sets of two
peptides that differ from each otherbya single amino acid
residue. Onesetiscomposedofpeptides17-39a and 17-39b
(Fig. 3).Thesepeptideshave identical amino acid sequences
corresponding to positions 17
through
39 in Fig. 4, exceptthatpeptide 17-39a has isoleucine atposition34 andpeptide
17-39bhasvalineatposition34.Anothersetiscomposed of
peptide 42-Sla, with a valine at position 46, and peptide
42-51b, with an isoleucine at position 46. The peak
desig-nated 74-98(1),42-51b in Fig. 3is amixture containingboth
peptide 74-98 and peptide 42-51b. Edman degradation
re-vealedtwo residues at each step, confirming thesequences of the twopeptidesandindicatingthatpeptide74-98 repre-sented greater than50% of the totalpeptide in themixture.
In addition, mass spectral analysis of peak 74-98(1),42-51b
(Fig. 3) confirmed the sequence of peptide 42-51b. The
determined mass spectra of peptide 42-51b and peptide
42-Sia are shown in Fig. 5. The two spectra are identical,
exceptthat themassesofionsd4to
dg
and a4toag
in 42-Sia(Fig. Sa) are14 massunits greaterthan those of the
homol-ogousions in42-Sib(Fig. Sb),confirmingthesubstitution of valine for isoleucine. The amount ofpeptide 17-39a recov-ered is about four times the amount of peptide 17-39b
recovered(seepeakheightsinFig. 3). Similarly,theamount
ofpeptide42-51a recovered is about four times the amount
ofpeptide 42-Sib recovered(Fig. 3). On thebasis of these
recoveries,weconclude that about 70to80%of theprotein
in pl7a (Fig. 1) has isoleucine at position34 and valine at
position 46 and that 20 to 30% of the protein hasvaline at
position34andisoleucineatposition46; together, thesetwo
formsmake up atleast66%of the totalp17eluted from the column. Inaddition,the determinedamino acid sequencesof
thesetwoforms differ from thepredictedsequencesatthree
otherpositions, atwhich theproteins havelysine in placeof asparagine(position 17) (peptides 17-39a and17-39b),
argin-ine in place of leucine (position 74) [peptides 74-98(1) and
74-98(2)], and lysine in place ofglutamic acid (position 92)
(peptide 74-98). Since both the 17-39a and the 17-39b
pep-tides havelysineatposition17 andnootherforms ofpeptide
74-98 were detected, we conclude that both forms of p17
present in thepl7a peak (Fig. 1)havelysineatpositions 17 and 92 and arginineatposition74.
Peptide 1-11 (Fig. 3) had an amino acid composition consistent with thepredictedsequence (based onthe DNA
on November 9, 2019 by guest
http://jvi.asm.org/
10 20 30 40 50 60
[image:5.612.160.472.68.336.2]---l---KR---I---I"I--V---
|l---KV----'''!'--I
K---l---.I---N---I---I----
--!
!---' ---v----'
----N
.---I.---.
---IK---1
pool
] A B
I C
I--- RK---I----I I--V--- i_ --- D
K .
i
~~~~-K---I---',--I---
11---70 80 90 100 110 120 130
SIQTGSEELKSLYNTVATLYCVHQKIEITKEALEKIEEEQNKSKKKAQQAAADTGNRGNSSQVSQNY
K.---I'""
I1 ]I
-K---KI-
'-'__----' '-1--'
---K.---R
K----
. -R-R---E D
l---K---Hl---l
---K----FIG. 4. Amino acidsequenceofp17.The amino acid sequences of thechromatographic forms ofp17purifiedasshown inFig. 1(forms pl7atopl7d)weredetermined byanalysis ofpeptidesasdescribed in the legendtoFig. 3. The deduced sequence based on the proviral DNA sequenceis shown in boldface type inthetopline. The determined aminoacidsequencesof the peptides are shown in pool A for pl7a, pool Bfor pl7b,pool C forpl7c,andpoolD forpl7d.Dashes indicate positionsatwhichthe deduced and determined sequences agree.
sequence) butwas "blocked" to N-terminal Edman
degra-dation. The peptide sequence was determined by mass
spectralanalysis.All sequence ions derived from the amino
terminus of thepeptide were 211massunits greater than the
sum of their amino acid residues. In addition, the spectra
showedamyristyl-Gly
[CH3(CH2)12CONHCH2CO']
ion at 268mass units and amyristyl[CH3(CH2)12CO']
ion at 211 mass units, as previously observed for other myristylatedpeptides (16, 19). The N-terminal peptide eluted late from
the column, consistent with the covalent attachment of a fattyacid,andwasimmediately preceded byanotherpeptide
(unlabeled in Fig. 3). Analysis of this preceding peptide
revealed that itwas apartial cleavage productconsisting of residues 1 to 16 and was also myristylated. No other
peptidesrecovered from thedigestcontained the N-terminal
residue. The molaryields of the N-terminal peptides (1-11
plus 1-16) were equal to the molar yields of the other
peptides
recovered inFig. 3. Therefore, weconcluded thatat least 95% of the protein in peak pl7a(Fig. 1), including
both forms ofp17, contained the N-terminal myristyl moi-ety. Except for disulfide-bonded peptides (Fig. 3)that may haveformed during the digestion, noother posttranslation-allymodified forms of theproteinweredetected.
Thetechniques (endopeptidase digestion, peptide
separa-tion andanalysis byEdmandegradation, andmassspectral
analysis)usedabove for theanalysisofproteininpeakpl7a
wereused for theanalysisofprotein purifiedinpeaks
pi7b,
pl7c, andpl7d (Fig. 1),and theresultsof theseanalyses are
alsosummarized inFig. 4.These results confirmedthatthe proteins purified inthepeakswereviralp17but alsoshowed that eachpeakcontainedamixture ofmultiple forms of the
protein. Peakpl7bcontained the forms ofp17 identified in
peak
pl7a
plus a form of the protein with asparagine atposition17and isoleucineatposition34. Inaddition, peptide
93-98was identified inpeakpl7b, suggestingthat aportion
of the proteincontainedglutamic acid at position92.
Anal-ysis of peak pl7c revealed additional forms with leucine,
serine, orasparagine at position74. The leucine at position
74 is in agreement with the nucleotide sequence, but the
peptidealso containedlysineatposition92, indisagreement
with the nucleotide sequence. The recoveries ofpeptides
from the analysis of peak pl7c suggested that this peak
contained a form of the protein with valine residues at
positions 34 and 46, in agreement with the nucleotide
se-quence. Peakpl7calsocontained forms of theproteinwith
glycine at position 89 and glutamine at position 104. Peak
pl7dcontained additional forms of theproteinwithlysineat
position 108and possibly glutamicacid atpositions 74, 84,
and 92(also indicated inpl7b).
Thetotalnumber of amino acid sequencevariantsof
p17
present in the virus population cannot be estimated from these results, because some variants may be present in amountsbelowourlimits of detection. However, the
analy-sis of the variouschromatographic forms of
p17
revealedat least eight different amino acid sequence variants of the protein. In addition, the proteins in peaks pl7e andpl7f
werenotanalyzed butprobablycontained additional variant forms. Nevertheless, at least one-half of the total
p17
is contributed by the major form in peak pl7a, and about one-fifth of thetotalp17
iscontributedby theminor formin pl7a. None of the forms identified were found to be in complete agreement with theproteinsequencededuced fromtheproviralDNAsequence ofHIV-1MN.Atleast98% of the
protein analyzed contained the N-terminal myristyl
post-translationalmodification,butpeptidesthatlacked the
mod--
---_I
ZV-
---
on November 9, 2019 by guest
http://jvi.asm.org/
* 40
u
a3
A.1-d5
L F
1.
b,
a3
as
11
.1
I.
I
.
a7a8 dg
a9
a
1.11
200 400 6Be BOB 10B0
m/z
[M+H]H
Arg
PhelAla
Val Asn Pro Gly Leu Leu Glu JR
II,
as
F
b,
a8 d9 ag
-1
.LI adUim6IWIA&h i a,I~Ii& I1IAL,r II.L IL
a ..
288 408
. I . I
689
a7
m/z
FIG. 5. Tandemmassspectraofpeptides 42-51a and 42-Sib(Fig. 3). Both peptidesrepresentpositions42to51 inFig. 4; residue 45 is isoleucinein42-51a(spectrum a) and valine in42-Sib(spectrum b). The fragment ions that contain residue 45are14massunits heavier in
spectrumathan in spectrum bbecauseofthepresenceofthe isoleucineinplace ofavaline.
ified N-terminalglycineresiduewerenoted in theanalysisof
pl7bandpl7c.
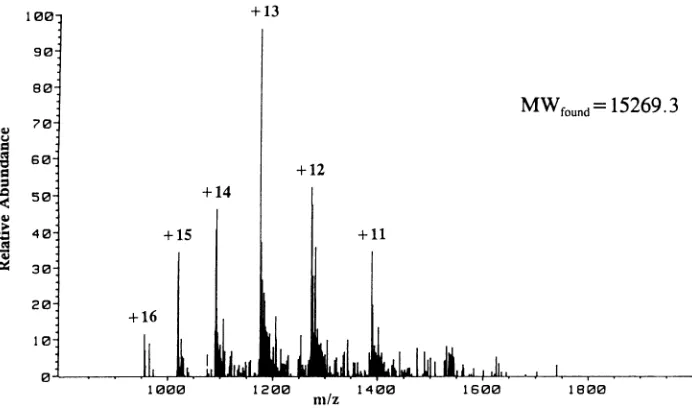
ESI-MS ofpl7.Theprotein purified inpeak pl7a (Fig. 1)
was analyzed byESI-MS. This isa highlyaccuratemethod for determining the molecular weight of a protein and is useful forverifyingamino acidcompositionsofproteinsand fordetectingthepresenceofposttranslationalmodifications presentbeforeproteolysis (26). Figure6 showstheESI-MS spectrum obtainedfrom the analysisofpl7a (Fig. 1). Anal-ysisof the iongroupswith differentcharged statesrevealed a molecularweight of 15,269.3 mass units for the protein. This value is in near-perfect agreement with the molecular weightof15,266.3massunits calculated from thedetermined
structuresof bothforms ofp17 (Fig. 4)present inpeak p17a (Fig. 1). Close agreement between the determined and calculated molecularweights strongly suggests that thep17 analyzed here (pl7a) is posttranslationally modified bythe addition of an N-terminal myristyl group but contains no other modifications. However, this conclusion cannot be extendedtothe otherchromatographicforms ofp17 (pl7bto
pl7f; Fig. 1), since they were not analyzed by ESI-MS. Phosphorylated forms of p17 have been detected by 32P
labeling of
HIV-lvAv
viral proteins (27), and it is quite possible that one or more of the minor chromatographic forms ofp17 contain phosphorylatedresidues. The finding that thepredominantforms ofp17arenotphosphorylatedis consistent with an earlier analysis of p17 purified fromHIV-1IIIB in which we did not detect the presence of phosphorylatedamino acids(15). Our previous analysiswas done with nonradioactive methods and would not have detected phosphoproteins present in <20% of the total protein analyzed.
Apoolofhighly purified p24 (markedinFig. 1) containing
at least 70% of the total p24 eluted from the columnwas
digested with endoproteinase Glu-C or trypsin. For each
digest, the resulting peptides were separated by rp-HPLC
(datanotshown)and analyzedasdescribed for thevarious p17 forms. The determined amino acid sequences were
aligned with the amino acid sequence predicted by the proviral DNA sequence (Fig. 7). Only the relevant and
(b)
8_
c
"a 68
.0
> 41
Cu L1
R
I I 11 -1 L AlI i..ai
I
.NM
AA Ak1
11
sZSi
1-I
rs
+
1-a6
I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.119.494.76.470.2]+13
+14
+15
+16
11.
1.|11A
MWfound=15269.3
+12
I.
+11
-J IIa* JiL,....LhMa.LELI.I.A,&ll"Milp*.l NM 1000 1200 1400
nmlz 1600 1800
FIG. 6. ESI mass spectrum of pl7a (Fig. 1). Each peak in the envelope is a molecular ion with the indicated charge state. A mass
spectrometermeasuresmass/charge; hence,themolecularweight ofalarge analyte, suchasp17,canbedetermined if the analyte hasahigh
chargestate(26). These datayieldadeterminedmolecularweight of 15,269.3 for pl7a (Fig. 1).
nonredundant peptides are shown. These analyses revealed
twoamino acid sequencevariants ofp24. Thepredominant form represented about 70% of the total protein analyzed and contained alanine at position 86; theother form repre-sented about30%of the totalproteinanalyzedand contained valineatposition86. Both forms differed from thepredicted amino acid sequenceat threeotherpositions: position 7, at which theprotein containedglutamine in place ofglutamic acid; position 184,atwhichtheproteincontainedtryptophan in place ofarginine; andposition 185, atwhich the protein containedmethionine inplaceof threonine.
Protein in the p24 pool (Fig. 1) was also analyzed by
ESI-MS, andamolecularweight of25,752 + 50was deter-mined (datanotshown). Thecalculatedweightsfor the two
forms ofp24 (Fig. 7)were25,551 (alanineatposition 86)and 25,579 (valineatposition 86). Thus,themeasured molecular weight is about 200 greater than the calculated molecular weight. Previous reportshave shown that HIV-1 p24 (from strain IIIB or BRU) contains phosphoserine and
phospho-threonine (15, 27) and exhibits multiple isoelectric forms containing phosphate (24). The ESI-MSdatadonotdirectly determine phosphates; however, the results of the analysis
are consistent with the previous proposals that the p24
protein isphosphorylated. The observedvariation in molec-ularweight (±50)isgreaterthantheprecisionof the ESI-MS method(26)and is inpartduetothesequenceheterogeneity in thesamplebut also couldsuggestthat thesamplecontains a mixture of diphosporylated and triphosphorylated
pro-teins. Unfortunately, mass spectral analysis of the proteo-lytic peptidesderived fromp24 (asinFig. 7)failedtoreveal any evidence of posttranslational modifications, including phosphorylations. Itseemspossiblethatputative phosphate
groups were lost through hydrolysis during the enzymatic
digestionofthe protein orseparationof the peptides. The otherGag proteins purifiedinFig. 1 (pl, p2, p6, and p7) were also analyzed by Edman degradation of purified
peptides and mass spectral analysis of peptides (data not
shown). The results showed that the amino acid sequences
1 10 20 30 40 50 60 70
PIVQNIEGQMVHQAISPRTLNAWVKVVEEKAFSPEVIPKFSALSEGATPQDLNTIIMTVGGHQAAMQNLKETINEEA
j Q---I- I---
---_____Q---
l-0-80 90 100 110 120 130 140 150
AEWDRLHPVHAGPITPGQNREPRGSDIAGTTSTLQEQIGWNTNNPPIPVGEIYKRWIILGINICVRNYSPSSILDIR
- --|j--A--
--V---A----I---
---A----1 ---1 ---H-H---'
---A----A-160 170 180 190 200 210 220 230
QGPKEPFRDYVDRFYKTLRAEQASQEVINRTTETLLVQNANPDCKTILKI.IQPAATLEEMKTACQGVGGPGHKARVL
---I II II--- WM---H
---FIG. 7. Amino acidsequenceofp24. Peptidesderived fromtrypsinandendoproteinaseGlu-Cproteolytic digestionsofp24 (Fig. 1)were
isolatedby rp-HPLCandsequenced byEdmandegradationandmassspectral analysis.The determined amino acidsequencesarealignedwith
the sequencededuced from the proviral DNA sequence shown in boldface type in the top line. Dashesindicate locations at which the
determinedsequences agreewith the deducedsequence.Two variantsofpeptide83-96werefound(aminoacidsequencevariations atposition 86). Themostabundant form ofpeptide83-96(approximately70% of thetotal) contained alanine atposition86.
100-
90-
80-
70-
50-402
302
202
101
_I III EL ME1111111-1_MNL&M
1--WN-H
---i
I.M- ti
i.
I-1.11.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.145.491.73.277.2] [image:7.612.156.479.551.677.2]75
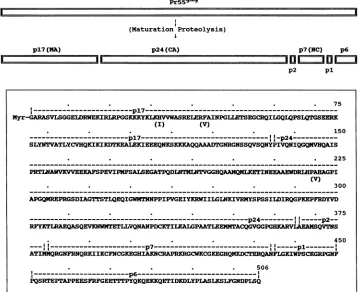
---pl7--- --_---________---________--_--Myr-GARASVLSGGELDRWEKIRLRPGGKKKYKLKHVVWASRELERFAINPGLLETSEGCRQILGQLQPSLQTGSEERK
(I) (V)
150
.- - - - -p17 . --1p24 .
SLYNTVATLYCVHQKICKIKIKEALEXIEEEQNKSKKKAQQAAADTGNRGNSSQVSQNYPIVQNIQGQMVHQAIS
. . . 225
PRIMAWVXVVEERAFSPEVIPNFSALSEGATPQDLNTQMLNGGHQAAMQNLETINEEAAEWDRIHPAHAGPI
(V)
. . . 300
APGQMREPRGSDIAGTTmSTLQEQIGWMTNNPPIPVGEIYKMWIIGINKIVRNYSPSSIDIRQGPKEPFRDYVD
. . . 375
.--- p24---
p2--RFYKTIRAEQASQEVKNWNTETLLVQNANPDCITILKALGPAATLEEMNTACQGVGGPGHKARVIAEAVQVTNS
p7.
. . . 450
-p7 ---
-.pI
----ATI!Q(QRGNFRNQRKIIKCFNCGKEGHIADNIcRAP IKRCNFCGKEGHQDRQ lKIWPSCKGRPGNF
. . . 506
---.p6----_._._._._-_
PQSRTEPTAPPEESFRFGEETTmTPYQKQEKKQETIDKDLYPLASLKSLFGNDPLSQ
FIG. 8. (Top)Schematic diagramshowing the maturationproteolyticprocessingof theHIV-lMNGag precursor to theGag proteins(p17, p24, p2, p7, pl, andp6)found inthe mature virus.(Bottom)Amino acid sequence of a functionalGagprecursor from the most infectious replicative variants of HIV-1MN grown in H9 cells. Amino acid residues in parentheses indicate substitutions that distinguish another competentGagprecursor also present in the infectiousreplicative variants.
ofpurified p2,
p6,
andp7were identical to the amino acidsequences predicted by the proviral DNA sequence, and therewas no evidence of amino acid sequence variants or
posttranslational modifications for these proteins. Purified
pl (Fig. 1) was found to have the amino acid sequence
FLGKIWPSCKGRPGNF, which differs from the predicted sequence at position 13, at which the pl protein contains
proline in place of a deletion, indicated by the DNA
se-quence, andatposition 14,atwhichthe plproteincontains glycine in place of arginine. No posttranslational modifica-tionsorsequencevariants of pl were detected.
In summary, the analysis of the purified Gag proteins shows the complete amino acid sequences and maturation
proteolytic cleavage sites for the Gag precursors that
con-stitute themajorportionof
HIV-lMN
producedbyinfectionandreplicationin H9 cells(Fig.8).The amino acid sequence
of the most abundant form of the Gag precursor
(approxi-mately50% of the total) is shown inFig. 8, and that of the
nextmostabundant form(approximately 20%ofthetotal)is
alsoindicated. The schematicdiagram showing the
matura-tionproteolyticcleavage sites of the Gag precursors (Fig. 8)
indicates the cleavage sites for generating the proteins de-scribed here (p17, p24, p2, p7, pl, and p6). The cleavage sites shown in Fig. 8 are highly efficient sites, since no appreciable partial cleavage products were identified. How-ever,otherpeptidefragments of the Gag proteins indicating less efficientcleavage sites were also noted during the course of this work. These included fragments of p2, pl, and p6, indicating cleavages between residues 369 and 370, 439 and 440, and 490 and 491 inthediagram shown inFig. 8.
DISCUSSION
Products of the retroviral gag gene perform highly com-plex orchestrated tasksduringthe assembly,budding,
mat-uration, and infection stagesof the viral replication cycle.
Duringviralassembly, the proteins form membrane
associ-ations(12,30,32)and self-associations thatultimatelyresult
inbuddingofanimmature virion fromthe infectedcell, and
point mutationsorsmalldeletionsin the gag gene canexert a dominant repressive effect on viral production (35). Gag
precursorsalsofunctionduringviralassembly toselectively
bind andpackage two plus strands of genomic RNA(10, 11).
The assembled immature virion contains approximately
2,000 to3,000 Gag precursors(3, 20) and becomes infectious after activation of theviral protease(21), proteolytic cleav-ageofthe Gag and Gag-Pol precursors, and rearrangement of the products to form the mature virion. In the mature
virion, the Gag proteins associate to form specifically
de-fined structures (9) that are probably necessary for the infection process.
The amino acid sequences ofGag precursors have been deduced from the DNA sequences of proviruses derived from numerousstrains ofHIV-1 (28). Many of these provi-ruses are known tobe infectious; however, it is difficult or impossible to deduce the exact amino acid sequences of highly efficient Gag precursors from the study of proviral DNA sequences. The most direct way todetermine struc-tures of Gag precursors that function efficiently in viral
assemblyistodetermineexactamino acid sequences ofGag
proteins incorporated into highly replicative viruses. H9
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.131.491.79.371.2]cellschronically infected with HIV-lMN contain many inte-grated proviruses (31) but do not shed large amounts of virions into the culture medium (2). However, when chron-ically infected H9 cells are continuously cocultivated with uninfected H9 cells, the amounts of virus shed into the mediumincrease by at least 50-fold. This biological amplifi-cation selects for the most infectious replicative viruses.
Here wehavepurified mature Gag proteins directly from large-scale production lots of HIV-1MN and determined their
completeamino acid sequences andposttranslational
modi-fications, includingthematuration proteolytic cleavagesites
in the precursors. We have usedcomplimentary, but inde-pendent, methods (mass spectrometry and automated Ed-man degradation plus amino acid analysis) (4, 8) to deter-mine the amino acid sequences and posttranslational modifications and have confirmed the results by indepen-dently determining the exact molecular weights of the puri-fied proteins by using ESI-MS. This combined approach creates alarge body of overlapping and redundant data that results inavery high level of confidence in the final deter-mined structures.
Theresults show that during maturation, the HIV-1 Gag
precursorsare cleavedtoyieldsixmatureGagproducts,as indicated in Fig. 8, and that the order of cleavage products in the precursors is p17-p24-p2-p7-pl-p6. The cleavage sites and number ofproteolytic products are in agreement with previous findings for
HIV-111,,
(15) and are also highlyhomologous to the cleavage sites and number of products previously identified for simianimmunodeficiencyvirus(14)
and related viruses, such as HIV-2 (18). The proteolytic
processing scheme proposed here reveals five cleavage sites
and differs from other proposals indicating fewer cleavage
sites (27, 36). All sixcleavage products wereisolated from
HIV-1MN (Fig. 1) and, on the basis of the amounts of the
proteins, we estimate that approximately equal molar
amountsof eachGagproteinwererecovered fromthevirus. These findings suggestthatcleavage at each site occursby
an efficient process and are in agreement with the equal
molar recoveries ofGagprecursorcleavageproducts
previ-ously reportedfor other retroviruses(14, 17).
Each Gag protein was recovered as a single
chromato-graphic entity,with theexceptionofp17, forwhichatleast
six chromatographically distinct forms were noted (peaks
pl7atop17finFig.1). Analysisofpl7atopl7drevealed that
aminimumofeightdifferent amino acid sequencevariantsof
this protein were present in the total
HIV-lMN population
(Fig. 4). Quantitative recoveries indicated that a single
amino acid sequence variant of
p17 (Fig. 4)
contributed atleast50%to the total
p17
recoveredfrom the virus and that another variant (also identified in Fig. 4) contributed anadditional20to30%tothe total.The remainderof the total
p17
was divided among the other amino acid sequence variants ofp17 (Fig. 4),with no singlevariantrepresentingmore than about 5 to 8% of the total. Analysis of p24
revealed two amino acid sequence variants (Fig. 7). The mostabundant variant contributed at least50% tothetotal p24 recovered from thevirus,and the other variant contrib-utedatleast20% to thetotal (30%of the totalp24 was not
analyzed). No other amino acid sequence variants were
revealedby analysis of the totalprotein recoveredforeach ofthe otherGag proteins(pl,p2, p6, and p7;Fig. 1).Onthe basis ofquantitative recoveries and amino acid sequences, we suggest that approximately 50 to 60% of the total Gag
proteinsrecovered are proteolytic cleavage fragments of a
single
Gagprecursorand thatanadditional 20to30%
of the total Gagproteins
mayoriginate
from anotherGag
precur-sor, as indicated in Fig. 8. The sequence heterogeneity
probably results from the expression of several different
proviruses integratedin the chronicallyinfected cell
popu-lation. The total number ofdifferent virusesexpressedin the mixture cannot be estimated, but the most replicative vi-ruseswillbeexpanded in the cocultivationprocedure. The
data provided here help characterize the more replicative
viruses and show the amino acid sequences ofGag precur-sors that function
efficiently
in the assembly and buddingprocess.
In addition to amino acid sequences, posttranslational
modificationscangreatly influence biological propertiesand mustbe considered when describingthe molecular compo-sitions ofproteins. A
previous quantitative analysis
ofthe IIIB(15) andLAV(27) strains ofHIV-1suggestedthat mostMAproteinsin thematurevirusesweremyristylated. Here
we analyzedfive chromatographicvariants of the MA
pro-tein produced by the MN strain and showed that each
variantwasmyristylated.Together, thesevariantsrepresent
thevastmajorityof the total MAproteinpresent inthe bulk
virus; therefore, these data suggestthat themost prevalent
forms ofthe viruses
produced
from the MN strain utilizemyristylated Gag precursors
during
viralassembly.
Theimportance of the myristylation modification (30, 33) is
underscored by the recent finding that analogs of myristic acid caninhibit virus
production (5, 6).
Asearch for otherpossible posttranslational
modifications was conductedby
comparing experimentally determined molecular weights
(mass
spectrometry)
and calculated molecularweights
(se-quencing)of the
purified
Gagproteins.
The observedmolec-ular
weight
of theprotein
in thepool
ofp24
analyzed
was200 ± 50weight
unitshigher
than the calculated molecularweight, indicatingthe presence ofposttranslational
modifi-cations. The nature of the modifications was not
directly
determined; however,
these data are consistent with theprevious
observations that p24 containsphosphoserine
andphosphothreonine
(15, 27).
The results
presented
here were obtained from asingle
production lot of HIV-1MN and clearly show amino acid
sequence
heterogeneity
in the isolated virus. The methodsused here can be
applied
to otherproduction
lots tohelp
determine sequence variations in the virus that may arise
throughcontinued
culturing.
Other lots have beenanalyzed
by rp-HPLC andshow
chromatographic
evidence ofheter-ogeneity.
However, a morerefinedchromatographic
proce-dure must be developed to analyze and compare the
se-quence
heterogeneity
in each lot. ACKNOWLEDGMENTSWethank Clara M. Dinterman and Patricia Coulter Grove for editorial and clericalassistance.
This research was sponsored, at least in part, bythe National Cancer Institute under contract N01-CO-74102with Program Re-sources, Incorporated/DynCorp. This workwasalsosupported,in part,bygrantBUS87-14238 from theNationalScience Foundation. Mass spectral measurements were performed at the Structural BiochemistryCenter,anNSF-supportedBiologicalInstrument
Cen-ter attheUniversityofMaryland,BaltimoreCounty. REFERENCES
1. Barre-Sinoussi, F., J.C.Chermann,F. Rey,M.T. Nugeyre,S. Chamaret, J. Gruest, C. Dauguet, C. Axier-Blin, F. Vezinet-Brun,C.Rouzioux,W.Rozenbaum, andL.Montagnier. 1983. Isolation ofaT-lymphotropic retrovirusfromapatientat risk for acquired immune deficiency syndrome (AIDS). Science
220:868-871.
2. Bess, J. W., Jr. (Program Resources, Incorporated/Dyn Corp,
on November 9, 2019 by guest
http://jvi.asm.org/
Natl. Acad. Sci. USA 86:8655-8659.
6. Bryant, M. L., L. Ratner, R. J. Duronio, N. S. Kishore, B. Devadas, S. P.Adams, and J.I.Gordon. 1991.Incorporationof 12-methoxydodecanoate into the human immunodeficiency vi-rus1gagpolyproteinprecursorinhibits its proteolytic process-ingandvirus production in achronically infected human lym-phoidcell line. Proc. Natl. Acad. Sci. USA88:2055-2059. 7. Devash, Y., T. J. Matthews, J. E.Drummond, K. Javaherian,
D.J.Waters, L.0.Arthur, W. A. Blattner, and J. R.Rusche. 1990. C-terminalfragmentsofgp120 andsyntheticpeptidesfrom fiveHTLV-III strains: prevalenceofantibodies to the HTLV-III-MNisolate in infectedindividuals. AIDS Res. Hum. Retro-viruses6:307-316.
8. Fenselau,C. 1991.Beyondgenesequencing: analysis ofprotein
structure with mass spectrometry. Annu. Rev. Biophys. Biophys. Chem. 20:205-220.
9. Gelderbloom,H. R., M.Ozel, and G.Pauli. 1989. Morphogen-esis andmorphology ofHIV,structure-functionrelations. Arch. Virol. 106:1-13.
10. Gorelick, R., L. Henderson, J. Hanser, and A. Rein. 1988. Point mutantsofMoloneymurine leukemiavirusthatfail topackage viral RNA: evidence forspecific RNArecognition by a "zinc finger-like" protein sequence. Proc. Natl. Acad. Sci. USA 85:8420-8424.
11. Gorelick, R.J.,S.M.Nigida, Jr., J. W. Bess, Jr., L.0. Arthur,
L. E. Henderson, and A. Rein. 1990. Noninfectious human immunodeficiency virus type 1 mutants deficient in genomic RNA.J. Virol. 64:3207-3211.
12. Gottlinger, H. G., J. G. Sodroski, and W. A. Haseltine. 1989. Role of capsid precursor processing and myristoylation in morphogenesis and infectivity of humanimmunodeficiency
vi-rustype 1. Proc. Natl. Acad. Sci. USA86:5781-5785. 13. Gurgo, C.,H.-G.Guo,G.Franchini, A.Aldovini,E.Collalti, K.
Farrell,F.Wong-Staal, R. C. Gallo, and M. S. Reitz, Jr. 1988. Envelope sequences oftwonewUnited States HIV-1 isolates. Virology164:531-536.
14. Henderson, L. E., R. E.Benveniste, R. Sowder, T. D. Copeland, A. M. Schultz, and S. Oroszlan. 1988. Molecular characteriza-tion of gag proteins from simian immunodeficiency virus (SIVMne). J. Virol.62:2587-2595.
15. Henderson,L.E., T. D.Copeland, R. C.Sowder,A. M.Schultz, andS. Oroszlan. 1988.Analysis ofproteins and peptides purified from sucrose gradient banded HTLV-III, p. 135-147. In D. Bolognesi (ed.), Human retroviruses, cancer, and AIDS: ap-proaches toprevention andtherapy. Alan R. Liss, Inc., New York.
16. Henderson, L. E., H. C. Krutzsch, and S. Oroszlan. 1983. Myristyl amino-terminal acylation of murine retrovirus pro-teins: anunusual post-translational proteinmodification. Proc. Natl. Acad.Sci.USA 80:339-343.
17. Henderson,L.E., R. Sowder, T. D. Copeland, G. Smythers, and
S. Oroszlan. 1984. Quantitative separation of murine leukemia virus proteins by reversed-phase high-pressure liquid chroma-tographyreveals newly described gag and env cleavage
prod-ucts.J.Virol. 52:492-500.
18. Henderson, L. E., R. C. Sowder, T. D. Copeland, S. Oroszlan, andR.E.Benveniste.1990.Gagprecursorsof HIVand SIV are cleaved into sixproteins found in the mature virions. J. Med. Primatol. 19:411-419.
viral infectivity. Proc. Natl. Acad. Sci. USA 85:4686-4690. 22. Laemmli, U. K. 1970. Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
23. LaRosa, G. J., J. P. Davide, K. Weinhold, J. A. Waterbury, A. T.Proty,J. A.Lewis,A. J.Langlois, G. R. Dreesman, R. N. Boswell, P. Shadduck, L. H. Holley, M. Karplus, D. P. Bolognesi, T. J.Matthews, E. A. Emini, and S. D. Putney. 1990. Conserved sequence and structural elements in the HIV-1 principal
neu-tralizing determinant. Science249:932-935.
24. Laurent, A. G., B. Krust, M.-A. Rey, L. Montagnier, and A. G. Hovanessian. 1989. Cell surface expression of several speciesof human immunodeficiency virus type 1 major core protein. J. Virol. 63:4074-4078.
25. Levy, J. A., A. D. Hoffman, S. M. Kramer, J. A.Landis,J. M. Shimabukuro, and L. S. Oshiro. 1984. Isolation of lymphocyto-pathic retroviruses from San Francisco patients with AIDS. Science 225:840-842.
26. Loo, J. A., C. G. Edmonds, and R. D. Smith. 1990. Primary sequence information from intact proteins by electrospray ion-ization tandem mass spectrometry. Science 248:201-204. 27. Mervis, R. J., N. Ahmad, E. P. Lillehoj, M. G. Raum, F. H. R.
Salazar, H. W. Chan, and S. Venkatesan. 1988. The gag gene products of human immunodeficiency virus type 1: alignment within the gag open reading frame, identification of posttrans-lational modifications, andevidencefor alternative gag precur-sors. J. Virol. 62:3993-4002.
28. Myers, G., J. A.Berzofsky, B. Korber, R. F. Smith, and G. N. Pavlakis (ed.). 1991. Human retroviruses and AIDS 1991, a compilation and analysis of nucleic acid and amino acid se-quences. Los Alamos National Laboratory, Los Alamos, N.Mex.
29. Popovic, M., M. G. Sarngadharan, E. Read, and R. C. Gallo. 1984. Detection, isolation, and continuous production of cyto-pathic retroviruses (HTLV-III) from patients with AIDS and pre-AIDS. Science 224:497-500.
30. Rein, A., M. R. McClure, N. R. Rice, R. B. Luftig, and A. M. Schultz. 1986. Myristylation site inPr65ragis essential for virus particle formation by Moloney murine leukemia virus. Proc. Natl. Acad. Sci. USA 83:7246-7250.
31. Reitz, M. (National Cancer Institute). 1991. Personal communi-cation.
32. Rhee, S. S., and E. J. Hunter. 1990. Structural role of the matrix protein of type D retroviruses in Gag polyprotein stability and capsid assembly. J. Virol. 64:4383-4389.
33. Schultz, A., and A. Rein. 1989. Unmyristylated Moloney murine leukemia virus Pr65gag is excluded from virus assembly and maturation events. J. Virol. 63:2370-2373.
34. Toplin, I., and P. Sottong. 1972. Large-volume purification of tumor viruses by use of zonal centrifuges. Appl. Microbiol. 23:1010-1014.
35. Trono, D., M. B. Feinberg, and D. Baltimore. 1989. HIV-1 Gag mutants can dominantly interfere with the replication of the wild-type virus. Cell 59:113-120.
36. Veronese, F. D., T. D.Copeland,S. Oroszlan, R. C.Gallo, and M. G. Sarngadharan. 1988. Biochemical and immunological analysis of human immunodeficiency virus gag gene products p17 and p24. J. Virol. 62:795-801.