Vol. 66, No. 8 JOURNALOFVIROLOGY,Aug. 1992, p. 4679-4685
0022-538X/92/084679-07$02.00/0
Copyright©1992, AmericanSociety forMicrobiology
Mechanism of
Attenuation of
a
Chimeric Influenza
A/B Transfectant Virus
GUANGXIANGLUO, MICHAEL BERGMANN, ADOLFO GARCIA-SASTRE,t ANDPETERPALESE*
Departmentof Microbiology, MountSinaiSchoolof Medicine,
1 Gustave L. LevyPlace, New
York,
NewYork10029-6574Received 27 March 1992/Accepted 19April 1992
The ribonucleoprotein transfection system for influenza virus allowedus to construct an influenza A virus
containingachimeric neuraminidase (NA)genein which thenoncodingsequenceis derived from theNS gene of influenza B virus(T. Muster, E. K.Subbarao,M. Enami, B. P. Murphy, and P. Palese, Proc. Natl. Acad. Sci. USA88:5177-5181, 1991). This transfectant virus is attenuated in mice and growstolower titers in tissue culture thanwild-type virus.Since such a virus has characteristics desirable foraliveattenuated vaccinestrain, attemptsweremadetocharacterize this virus at the molecularlevel. Our analysis suggests that the attenuation of the virus is duetochanges in the cis signal sequences, which resulted in areduction oftranscription and replication of the chimeric NA gene. The majorfindingconcerns a sixfoldreductioninNA-specific viral RNA
inthe virion,causing a reduction in the ratio of infectious particles to physical particles compared with the ratio inwild-type virus. Although the NA-specific mRNA level is also reduced in transfectantvirus-infected cells, it doesnotappear tocontribute to theattenuationcharacteristics of thevirus. The levels of the other RNAs and theirexpression appeartobeunchanged for the transfectant virus. Itis suggested thatdownregulation of the synthesis of one viral RNA segment leadstothegeneration ofdefective viruses during each replication cycle. We believethatthis represents a general principle for attenuation which may be appliedtoother segmented virusescontaining either single-stranded or double-stranded RNA.
Influenza, anacuteinfectious respiratory disease, remains
a constant worldwide threat to human health and, in the absence ofeffective antiviral agents, can best be prevented
or limited byvaccination (1, 13). The influenza virus vac-cines currently in use in the United States are derived from inactivated influenza virus, and they provide only limited protection in vaccinees (2, 13). An alternative
ap-proachwould involve the development of safe live attenu-atedvaccines; these vaccines should be longer-lasting and
afford greater protection (1). An ideal way to generate suchanattenuatedvaccinewould beto usegenetic engineer-ing techniques. However, until recently, influenza
virus-likeall othernegative-strand RNAviruses-wasnot
amena-ble to site-specific genetic manipulation. The situation has
changed for influenzavirus,becausesyntheticRNAs
recon-stituted in vitro withpurifiedviralpolymerase complexcan now be rescued intoinfectious influenza virus particles (5,
10, 12).
Applying thisnovel reverse genetic approach, we gener-ated a transfectant influenza virus which possesses a chi-meric neuraminidase (NA) gene containing the coding
se-quenceof theNAgeneofinfluenzaA/WSN/33virus andthe
noncoding sequence from the NS segment of influenza B/Leevirus. The viruswas showntobeattenuated inmice
and to induce a protective immune response against
chal-lenge virus. These features suggested that the virus has many properties desirable for an attenuated live influenza virus vaccine (14). In order to understand the molecular characteristics responsible for the attenuation of this
trans-fectant virus, we carried out a series of experiments to
*Correspondingauthor.
tOnleaveofabsencefromtheDepartmentofBiochemistryand MolecularBiology, FacultyofBiology, University ofSalamanca, Salamanca, Spain.
analyze the virusatthe molecular level. The data presented inthisreport suggestthatattenuation results from the effect of the altered cis elementsonthereplication of the chimeric NA gene.Alow level ofthe NA gene in the virus preparation results in a higher proportion of defective particles than is foundin awild-typeviruspreparation. Inaddition, the data support a random mechanism for the packaging ofvirion RNAs(vRNAs)into influenza virus particles.
MATERIALSANDMETHODS
Viruses and cells. Thestocks ofinfluenzaA/WSN/33virus and the NA/B-NS transfectant virus were prepared from
purified plaques by growing them in Madin-Darby bovine
kidney (MDBK) cells in reinforced minimal essential
me-diumcontaining 2 ,ug oftrypsin per ml. MDBKcells were used for the plaquing of viruses and the study of
virus-specific RNAsynthesis.
Plasmids. In order to generate riboprobes, several
plas-midswereconstructed. pSP64-NS DNA contains the entire NS segment ofA/WSN/33virusinserted into the SalI site of the pSP64vector(Promega, Madison, Wis.) in the orienta-tionbywhichpositive-senseNS RNAcanbe madebyusing
SP6 RNApolymerase. To create plasmid pIBI30-NS, the entire NS DNAfragment derived from pSP64-NS was in-serted between theHindIII and EcoRI sites of the pIBI30 vector(IBI,NewHaven,Conn.)in the orientationbywhich
negative-senseNSRNA canbegenerated byusingT7RNA polymerase. In order to obtain plasmid AT3NAv, which produces the negative-sense NA-specific RNA probe, the
DNAin thepT3NAvplasmid(5)was shortenedby deleting
nucleotides 1 to 1096 of the NA vRNA. In
addition,
wecreated a plasmid, designated IBI30-NA, by
inserting
the entireNAfragmentderived frompT3NAv(5)
into theXbaI 4679on November 9, 2019 by guest
http://jvi.asm.org/
4680 LUO ET AL.
and EcoRI sites of the IBI30 vector. This plasmid can produce positive-sense NA-specificRNAbyT7 RNA poly-merasetranscription.
Virus purification and RNA extraction. Influenza A/WSN/33 virus and NA/B-NS transfectant virus were growninMDBKcells and thenpurified by30 to60%sucrose gradient centrifugation as described previously (5). Virus
purifiedfrom four175-cm2flasks of MDBKcellswas
resus-pended in0.3 mlof TMK buffer (10mM Tris [pH 7.5], 1.5 mMMgCl2, 10 mMKCl) anddisrupted byincubationwith 9 pul of 10% sodium dodecyl sulfate (SDS) and 7.5 pul of
proteinase K (10 mg/ml) at 56°C for 10 min, followed by
addition of 35 ,ulof SLN buffer(5% SDS, 1.4 M LiCl, 100 mMsodiumacetate [pH 7.0]). Virion RNAswereextracted with phenol-chloroform and collectedbyethanol precipita-tion. For isolating viral RNAs from infected cells, MDBK cellswere infected with either influenzaA/WSN/33virusor NA/B-NStransfectant virusatamultiplicityofinfection of 1 and harvested at the indicated time points. Cells were
washed twice with ice-cold phosphate-buffered saline and
lysed with 4 M guanidinium isothiocyanate (Sigma). Total RNAwas then purified by equilibrium centrifugationin 5.7 Mcesium chloride (Fisher) (9).
Determination ofthe ratio of infectious andphysical parti-cles. In order to determine the total number of physical particles ofinfluenza A/WSN/33virus andNA/B-NS trans-fectant virus, the virus preparations were mixed with an equal volume of a suspension of carboxylate polystyrene beads (0.1 pum in diameter) at aconcentration of4.5 x 109 particles per ml (Polyscience, Inc., Warrington, Pa.) and then stained with phosphotungstic acid. Theratio of virus andpolystyrenebeadswasdeterminedby countingthe two differentparticles under the electronmicroscope. For mea-suringthenumber of infectiousparticlesin thepreparations, the virusstockswereseriallydiluted andplaquedinMDBK cells.
RNA electrophoresis. RNAs extracted from influenza A/WSN/33virus andNA/B-NStransfectantviruswere
elec-trophoresed on a 3% polyacrylamide gel containing 7.7 M ureaat150 V for 2 h.The RNAsegmentswerevisualizedby silverstaining as described previously (5).
RNaseprotection assay. The RNaseprotection assaywas used forquantitationof virion RNA in viralparticlesaswell as for the measurement of viral mRNAs and cRNAs in infected MDBK cells. The NS segmentwas chosen as an internal control. FormeasuringvirionRNAs, positive-sense NS- andNA-specificRNAprobesweregenerated byrun-off
transcription with phage SP6 or T7 RNA polymerase and plasmid pSP64-NSDNA linearizedbyNcoI(nucleotide [nt] 265) or plasmid IBI30-NA digested with FokI (nt 240), respectively. The NS-specific RNAprobe spans theregion between nt 1 and265 of the NScRNA,and theNA-specific RNAprobe covers nt 1 to 240 ofthe NAcRNA.
Todetermine the amounts of mRNAsandcRNAs, nega-tive-sense NS- and NA-specific RNA probes were tran-scribed from DdeI (nt 465)-digested IBI30-NS DNA and PvuII (nt 306 in pUC19)-cut AT3NA DNA, respectively, with phage T7 or T3 RNA polymerase, respectively. The negative-sense NS-specific RNAprobe is470nt long, con-tainingnt465to890 of theNSvRNAandavectorsequence
of45 nt. The negative-sense NA-specificRNAprobeis403 nt long, coveringtheregionbetweennt1096 and1409 of the NAvRNAand containing a vectorsequence of 90nt.
The RNA probes were labeled with [a-32P]UTP (800 Ci/mmol; DuPont, NEN, Boston, Mass.). Ingeneral,50ng
of virion RNA extracted from purified virus was
hybrid-ized to 5 x 104 cpm each of the positive-sense NS- and NA-specific probes, or 5 p.g of total RNA isolated from virus-infected cells was hybridized to 5 x 104 cpm each of the negative-sense NS- and NA-specific probes. After 12 h ofincubation at 45°C, the hybridization mixture was digested with RNases A and
Tl,
following the manufactur-er'sinstruction (Ambion Inc., Austin, Tex.), and the result-ing productswereanalyzed on a 6% acrylamide denaturing gel (9).Primer extension. The genomic RNAs (vRNAs) of the NA and NS segments of influenza A/WSN/33 virus and of
NA/B-NS transfectant virus were quantitated by primer extension (11). The primers for detecting NS- and NA-specific vRNA are 21 nt long and are complementary to
negative-sense vRNA. The NS primer, 5'-GGGAACAATT
AGGTCAGAAGT-3', spansthe region between nt 695 and 714 of the NS cRNA. The NAprimer, 5'-GTGGCAATAAC TAATCGGTCA-3',coversnt1151to1171 of the NA cRNA. Both the NS and NA primers were 5'-end labeled by incubation with[_y32P]ATP (3,000 Ci/mmol; Du Pont, NEN) and T4 DNA kinase(Biolabs, Beverly, Mass.). Then, 100 ng of RNA extracted from virus or 5 ,ug of total RNA isolated from infected MDBK cells was reverse transcribed by RNase H-minus murineleukemia virusreversetranscriptase
(BRL, Gaithersburg, Md.) in the presence of 3 x 105 cpm each of the5'-end-labeled NSand NA primers. After incu-bationat37°Cfor2h,thereactionwas stopped by addition of EDTAto 10 mM, and the reaction mixwassubjected to
phenol-chloroformextraction and alkalitreatment(11). The
productswereanalyzedon a6%polyacrylamide gel
contain-ing 7 M urea. The amount of product was measured by
directly counting the radioactivity of the gel piece corre-spondingtoeach band onthe film.
Neuraminidase assay and Western immunoblot analysis. For the assay of neuraminidase activity of the influenza
A/WSN/33
andNA/B-NStransfectantviruses,2'-(4-methyl-umbelliferyl)-at-D-N-acetylneuraminicacid(Sigma)wasused
asthe substrate. Thereaction mixture consisted of 25 ,ul of 2 mMsubstrate,25 ,ulofvirus,and50 ,u of 0.2 Mphosphate
buffer(pH 6.0) containing2 mM CaCl2.Afterincubation at
37°C
for 10min,thereactionwasstopped byaddition of 2 ml of 0.5 M glycine-NaOH buffer (pH 10.6), and then the neuraminidase activity was determined by measuring thefluorescencewithexcitation at365 nm and emission at450 nm, with methylumbelliferone as a standard. The protein concentration of the viruseswasmeasured with the Bio-Rad
proteinassay kit. For the Western analysisofthe NA and HA proteins of the WSN/33 and NA/B-NS viruses, viral
proteins were electrophoresed on a 10% polyacrylamide
Laemmligel (7) andsubsequentlytransferredto a nitrocel-lulose membrane (Schleicher & Schuell, Keene, N.H.). A monoclonal antibody directed against carbohydrates was
usedtodetect theNAandHAglycoproteinsof the viruses. TheWesternblotwasdevelopedwitha ratantibody against mouse kappa chains, which was labeled with 125I and was
generously provided byThomas Moran.
Analysis ofprotein synthesisin infectedcells. MDBKcells
(35-mm dish) were infected with either WSN/33 virus or NA/B-NStransfectant virus at amultiplicity of infection of
approximately 3. This multiplicity was used because the
NA/B-NStransfectantvirus didnotgrowtohigher titer. At indicated times, the proteins were labeled in cysteine-free medium with
[35S]cysteine
(1,027 Ci/mmol;DuPont, NEN) at 100 ,uCi/ml of medium for 30 min. The cellswere then washed twice with ice-cold phosphate-buffered saline and lysed in 150pul
oflysisbuffer containing 1% Nonidet P-40,J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
ATTENUATION OF CHIMERIC INFLUENZA A/B VIRUS 4681 NA of Influenza A WSN/33 Virus
5AGUAGAAACAAG AQUUUGAACAAACUA
3- UCGCUUU C GUCCUCAAAUJIAC
NA of NA/B-NSTransfectant Virus
5AGUAGUAACAAGAGGAuuuuuAUUUUACAUUCA
3UUCCUUCGUCUCCUA
AAUAAAUCAGUGACCGUUUGCCUUUCUAC
FIG. 1. Noncodingsequencesof the NAsegmentsofinfluenza
AIWSN/33virus and the NA/B-NStransfectant virus. The 5'- and 3-terminal sequences are drawn in a panhandle structure, which consists of twobase-pairedstemsand onemismatchedinternalloop in themiddle. The noncoding nucleotides of thechimericNA gene of the NA/B-NS virus arederived from the NS gene ofinfluenza B/Lee virus. The large letters indicate nucleotidesin the 5'- and
3'-terminal regionswhicharedifferentfor thetwo NA genes. The opentrianglemarks the altered Kozaksequencein theNA geneof theNA/B-NS virus.
150 mM NaCl, 50 mM Tris-HCl (pH 8.0), and 1 mM phenylmethylsulfonyl fluoride. About 1/20 of the sample was loadedonto a 10% polyacrylamide Laemmli gel (7).
RESULTS
Ratio of infectious to physical particles.The genome of the transfectant NA/B-NS virus differs from that ofwild-type influenza A/WSN/33 virus onlyin the noncoding region of the NA gene (Fig. 1). It is thus likely that the altered
biological properties of the transfectant virusarethe result of alteredcis signals located in thenoncoding regionof the chimeric NA gene (9, 15, 18). Specifically, it was noted earlier that the transfectant virus grewto lower titers than wild-type virus in MDBK cells, MDCK cells, and mice. In addition, the low-multiplicity growth curves in tissue
cul-ture weresignificantly delayed relativetothoseofwild-type
virus (14). We then proceeded to examine whether the
transfectant virus had a temperature-sensitive phenotype,
which could explain the altered growth characteristics.
However, the pattern of the growth curve at 30, 33, 37,
and 38°C for NA/B-NS virus was not different from that ofwild-type virus at the corresponding temperatures. We
next asked whether defective particles were present in theNA/B-NSvirus
preparation.
Theviruswascharacterized by counting the physical particles under the electronmicroscope and comparing this number with the PFU of the preparation. Interestingly, the NA/B-NS transfectant virus showed a similar number of physical particles as
the wild-type virus but consistently lower PFU titers (14) (Table 1). Thus, theNA/B-NS virus grown in MDBK cells shows at least a 5- to 10-fold-lower infectious
particle-to-physical particle ratio than is seen with the WSN/33 wild-typevirus.
[image:3.612.60.300.682.727.2]Characterization of theRNAof the NA/B-NS virus. Since
TABLE 1. Ratioof infectiousparticlestophysical particlesof influenzaA/WSN/33virus andNA/B-NStransfectant virus Virus Physical(pp) (no./ml)particles Infectious particles Ratio, pp/ip
(ip) (no./ml)
WSN/33 1.8 x 109 1.2x 108 15
NA/B-NS 1.2 x 109 1.0 x 107 120
the NA/B-NSvirus contains many defective particles, the viral RNA was examined for the presence of defective RNAs. Following extraction from purified virus, the ge-nomic RNA was separated on a 3% polyacrylamide gel
containing 7.7 M urea. As shown in Fig. 2A, the chimeric NA RNA of the NA/B-NS transfectant virus is almost invisibleon the gel, whereas theother sevensegments are
present in approximately equimolar concentrations. When theamountof RNAonthe gelwasincreased,the chimeric
NARNAwasshown tomigrateatthesamepositionasthe control RNAin lane3(datanot shown). However,
electro-phoresis andsilverstainingdidnotpermitthequantitationof
thechimericNA RNApackagedinvirions.For this purpose, the RNase protection assay and primer extension experi-ments wereperformed.
Positive-sense NS-specific and NA-specific probes were hybridizedinthesamereactiontopurifiedvirion RNA from
eitherWSN/33 virus(Fig. 2B,lane3)orNA/B-NS transfec-tantvirus(Fig. 2B, lane4) and then digestedwith RNases A and T1. The resulting products were analyzed on a 6% polyacrylamide gel
containing
7 Murea(9),
andtheamountsof vRNAwerecalculatedby countingtheradioactivityof the
gel slices
corresponding
tothe bandson the film. As shownin Fig. 2B, the probe protected bythe chimericNA RNA
migratesfaster than thatof thewild-typegenebecausethe 5
noncoding sequences are different in the two NA genes.
Compared with the NA segment of WSN/33
virus,
theamount of chimeric NA RNA in the transfectant virus is about sixtimeslower, with the NSgene usedasaninternal
control.The
primer
extensionexperiment,
asshown inFig.
2C, showsasimilarreductionin theamountofchimericNA RNApackagedinvirions relativetotheNS gene,suggestinga
specific
lowerrepresentation
of the chimericNA gene inthetransfectant virus preparation.
Characterization of theproteinof theNA/B-NSvirus. The
nextquestionwaswhether theNA/B-NStransfectantvirus also contained less neuraminidase
protein.
We carried out neuraminidase assays and a Westernanalysis
of the NAprotein in virus particles. For the enzymatic assayof the
neuraminidase, the totalconcentration of viral
proteins
wasfirst determined by the
protein
assay, and then the same amount ofpurified
viruswas used. TheNA/B-NS
transfec-tantvirus showedonlyatwofold-lower NA
activity
thanthe WSN/33 wild-typevirus. A similarfinding
was obtainedbyWestern analysis, which showed 1.9-fold less NA
protein
but the same amount of HA
protein
in virions of theNA/B-NS transfectant virus
compared
with those ofWSN/33virus(see Fig.
5A).
Virus-specificRNAsynthesisininfectedcells.Basedonthe vRNAanalysisof theNA/B-NStransfectant
virus,
there is little doubt that lesschimeric NA RNA is contained in the viralparticles. Thelower level of the chimeric NARNAin the virions could be caused eitherby
achange
in thepackaging signal, leading
to less efficientpackaging
of theNARNA, orby a changein the
synthesis
ofgenomic
NA RNA,whichcouldbe the result ofinefficientrecognition
of the influenza B virus-specificpromoter by the influenza Aviral polymerase. To
pinpoint
the exactmechanism,
weexamined the vRNA
synthesis
of the chimeric NA gene in infected MDBK cells.Considering
that vRNA ismainly
synthesized
in the latephase
of infection(19),
weusedRNAextracted from cellsat7,
8.5,
and 10 hpostinfection.
Total RNA(5,ug)
extracted fromvirus-infected cellswasused for theprimerextensionanalysis.
The data inFig.
3show that vRNAsynthesis ofthe chimeric NAgenewasremarkably
decreased relative tothat of control virus.Compared
withVOL.66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
4682 LUO ET AL.
A.
WSN/33 NA/B-NS
eI
ControlF....
i-NA/B-NS C.
310 281 27 1 234 194
B.
(0c Un o c. Z .0a) *
*_
Z_n
ML-0
a. O ulz
NS
NA d Z NS
0
-NA+4NAIB-NS
2 3 4
X n
s z)
)-.
4-NA
[image:4.612.115.480.78.303.2].--NS
FIG. 2. Characterization of theRNA oftheNA/B-NS virus. (A)RNAelectrophoresis. The RNAs extracted from purifiedviruseswere analyzedon a3%polyacrylamidegelcontaining7.7 M urea andvisualized by silver staining. Lane 1, RNA of influenza A WSN/33 virus; lane 2, RNAofNA/B-NStransfectantvirus; lane 3, RNAobtained by run-off transcription fromplasmidpT7NA/B-NS, which produces the chimericNARNA.(B) Analysis ofNARNA invirions by the RNase protection assay. RNA (50ng)extractedfrompurified virus was used in thehybridization reactionwith positive-sense NS-andNA-specific riboprobes asdescribed in Materials and Methods. The protected probeswereelectrophoresed on a6% acrylamide gel containing 7 M urea. Lane 1, riboprobes without RNase A orT1digestion; lane 2, riboprobes following RNase A andT1digestion; lane 3, riboprobes protected by the RNA of influenza A/WSN/33 virus; lane 4, riboprobes protected by the RNA ofNA/B-NS transfectantvirus. (C) Quantitation of NA-specific RNA in virus by primer extension. The vRNA extracted from either WSN/33 virus (lane 2) or NA/B-NS transfectant virus (lane 3) was reverse transcribed by RNase H-minus reverse transcriptasewithNS andNAsegment-specificprimersasdescribed in Materials and Methods. The products for the NS RNAs are 195 nt long,andthose forthe NA RNAs areapproximately 260ntlong.The products wereanalyzedona 6%polyacrylamide gel containing 7 M urea. Size markers are shown onthe left(in nucleotides).
the synthesis of the NA RNA of WSN/33 virus, synthesis from thechimericNA geneis reduced by a factor of 9, 8, and 8at7,8.5, and 10 hpostinfection,respectively. However, no
reduction of vRNAsynthesis was observed with respect to
w)
WSN/33 NA/B-NS 1 7 8.510 1 7 8.510
Vicnr
3 Z
Virion p.i.(hr)
310
281 4-NA
271
234
23
4
194w O*, .*
~ ~ ~ 4
FIG. 3. Analysis of NA-specific vRNA synthesis in infectedcells by primerextension. RNA isolated from infected cellswasreverse
transcribed with reverse transcriptase and NA and NS vRNA-specific primers, asdescribed in Materials and Methods. vRNAs extracted from purified viruswere used as a control (right). The
resulting products were displayed on a6% acrylamide denaturing gel. Thereversetranscripts of the NS RNAsare 195 ntlong, and those of theNA-specificsegmentsareapproximately260ntlong,as
indicatedbyarrows atthe right. The numbersonthetopindicate times postinfection. Virion represents the RNA obtained from purified virus. Sizemarkersareshown in nucleotides.
the NS segment of the NA/B-NS transfectant virus. This result suggeststhat the reduction of thechimeric NA RNA in transfectant virus is similar to the reduction in
synthe-sis observed in infected cells. Thus, the lower
representa-tion of the chimeric NA segment in virus particles is most
likelynotdueto adefect in the packaging of the chimeric NA RNA.
In ordertodetermine the level of mRNAsynthesis of the chimeric NAgene, MDBK cellswere infected with either WSN/33 virus or NA/B-NS transfectant virus, and total RNAwasthen isolated from virus-infected cellsatdifferent timespostinfection. Subsequently, the level ofvirus-specific
mRNAs was quantitated by the RNase protection assay.
Again, the NSsegment wasusedasacontrol.From the time course, it isapparent thatthe level ofchimeric NA mRNA is markedly reduced relativetothat of thewild-type virus and that it only increases slightly with time (Fig. 4A). At 5 h postinfection,thelevel ofchimericNAmRNA wasfivefold
less than that of WSN/33 virus (Fig. 4B), whereas the NS-specific mRNA synthesis of the NA/B-NS transfectant
virus was similar to that ofWSN/33 virusat the indicated times (Fig. 4C). It should be noted that the NS-specific mRNA synthesis appears earlierand is more efficient than NA-specificmRNAsynthesisinbothWSN/33andNA/B-NS
transfectant virus-infected cells (probes ofsimilar activity
were used in the experiment). This finding is in good
agreement with previous reports (4) and suggests that the mRNA synthesis of differentinfluenzavirusRNA segments
isdifferentially regulated. The conditions of theassaydidnot J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.77.277.500.624.2]ATFIENUATION OF CHIMERIC INFLUENZA A/B VIRUS 4683
A
NS Probe - 470
(-)Sense Sense
N.S...-RNA.... NS mRNA
5
'-Product
AAA(A)n 3'
+ 409
nt*
NA Probe
t=03
4 nt(-)Sense 3 '
NA mRNA 5
'-Product
IAAA(A)n 3'
14-298
nt-W'a) 0
QD
-o0
-CL C)
NSo:
NA-ov
.Vl
WSN/33
Virus
1 3 4 5 6 7
3
lo45
eOQe
NA/B-NS
Virus1 3 4 5 6 7
Hours(p.i.)
-NS
4-NA
B 4000 --- WSN/33
* NA/B-NS
C
a
._
0
0
0
EL
cJ
2000
1000
4 6
Hours (p.i.)
-0--O WSN/33
-a- NA/B-NS
FIG. 4. Timecourseof mRNAsynthesisin MDBKcells.(A) Quantitation ofNA-andNS-specificmRNAsatdifferent timespostinfection (p.i.) bytheRNaseprotectionassay.Thediagramonthetopschematically illustrates the procedure.Both theNSand NAprobesarenegative senseand containsequencescorrespondingtothe 3'-terminalside of thecRNA, flankedbyvectorsequencesatthe 3'terminus,asindicated by theopenrectangles.The sizes oftheprobesareshownonthetop, and the sizes ofthe resulting productsareindicatedatthe bottom of thediagram.The timepointsareindicatedonthe top. Thepositionsofprobes and productsonthegelareindicatedbyarrowsatthe left and right, respectively. (B)TimecourseofNA-specificmRNAsynthesis.Theamountof the NA mRNAwasmeasuredby directly countingthe radioactivity (countsperminute)in thecorrespondingband excised from thegelshown inpanelA. (C) ComparisonofNS-specificmRNA synthesisoftransfectant virus andA/WSN/33virus in infected cells. Theamountof mRNA foreach timepointwasdeterminedasdescribed forpanelB.
permitustoquantitatethe level of cRNA of the chimeric NA
genebecausethe cRNAsynthesiswasfoundtobeonly3to 5% of that of mRNAsynthesis (lOa).
Reduction of NAprotein synthesisinvivo. The NAprotein was found to be reduced by a factor of 2 in virions of the NA/B-NS transfectant virus, whereas thesynthesis of chi-meric NA mRNAwasreducedbymuchmore.Thequestion
thenarosewhether the NAproteinencodedbythe chimeric NA mRNA inside cellsparallelstheamountof itsmRNA,or whether thereisaselective incorporation of the
neuramini-dase into the viral envelope. In orderto answer this
ques-tion, we measured the synthesis of NA
protein
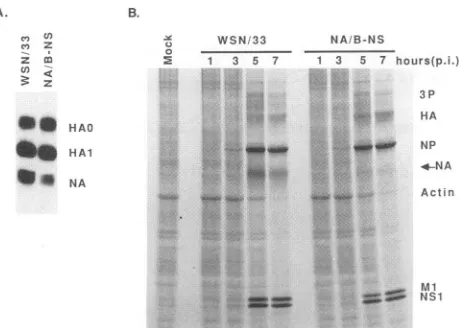
in infected cells. Viral proteinswere labeled with[3Slcysteine
at 1, 3, 5,and 7 hpostinfection (Fig. SB).The celllysateswerethen analyzedon a10%polyacrylamideLaemmligel(7), and theproteinswerequantitatedbyanAMBISradioanalytic imag-ingsystem(11). Synthesisofthe NAproteinof theNA/B-NS
transfectant viruswasmeasured and showntobetwotimes lower than that of wild-type virus at both 5 and 7 h postinfection. From this experiment,we canconclude that
3000
a -E C
a
CD
C M._
0 C)
4 Hours (p.i.)
8
VOL. 66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.69.557.72.511.2]J. VIROL. B.
c., Z z m
en
;z
: z
0 HAO
HAl
fb40
WSNi'33 NA/B-NS
1 3 5 7 1 3 5 7 hours(p.i.)
3 P
:-;p..^* HA
PA N P .-NA
Actin
NSl
FIG. 5. Analysis of NA protein invirionsand in infected cells. (A) Western analysis of NA protein in virions. As described in Materials and Methods, a monoclonal antibody directed against carbohydratewasusedtoquantitatetheglycoprotein in the viruses. Theproteinsareindicatedby HAO(uncleaved HA),HAl (subunit 1
of HA), and NA at the right. (B) Viral proteins synthesized in infected cells. At different times postinfection (p.i.), the viral proteinswerelabeled with [35S]cysteinefor 30min and analyzedon
a10%acrylamide Laemmli gel. Thedifferent proteinsaremarkedat the right (see Fig. 2 legend). The position of the NA protein is
indicatedbyan arrow. TheamountofNAproteinwasdetermined
by counting the radioactivity in thegel in a radioanalytic AMBIS imagingsystem (11) with the NP and Ml proteinsascontrols.
the assembly of the NA protein into virions parallels its synthesis in infected cells.
DISCUSSION
Inapreviousreport, wedescribedatransfectantinfluenza
Avirus containing achimeric NAgenein which the
noncod-ing sequences are identical to those in the NS gene of
influenza B/Lee virus. This virus has many unique growth
characteristicsin tissue culture, and it is highly attenuatedin mice (14). Since such avirus could be aprototype for live influenzavirus vaccines, attemptsweremade tounderstand the molecular mechanism underlying these changed growth characteristics. Several lines of evidence obtained from theseexperimentssuggestthattheciselements derived from the influenza B/Lee virus gene are responsible for the
dramatic effects on transcription and replication of the
chimeric NAgeneof the NA/B-NS transfectantvirus. Itwas found that the NAgenehadasixfold-lower representationin
the purified viral preparation than did the remaining seven
RNAs. This strikingly lower representation ofone RNAis
compatible with the finding that the NA/B-NS transfectant virus has an approximately 5- to 10-fold-lower infectious particle-to-physical particle ratio than wild-type virus.
It is assumed that an infectious virus would require the
presence of a full complement of all eight influenza virus
RNA segments. Many of the NA/B-NS progeny virus,
however, lackanNAgene, sothatmore defective particles
areformed than is the casein awild-type virus infection. It
isnotclearwhetherthis5-to10-fold reduction in titer is only
areflection of the lower representation of the NA gene or
whether other factors also play a role. For example, some
viruses may contain defective interfering RNAs which would lower the infectivity titer of the preparation.
How-ever, in this context it should be noted that there is no
evidence for the presence ofsignificant quantities of defec-tive interfering RNAs in theNA/B-NSvirus.
The mRNA synthesis of the NA is also considerably
reduced in transfectant virus-infected cells. Surprisingly,
this does not lead to a commensurate reduction in protein synthesis. Both virus-infected cells and purified virus show
only a twofold-lower level of NA protein than wild-type virus-infected cells or purified wild-type virus itself. This higher-than-expected level of NA protein in thetransfectant virusmaybethe result ofagoodKozaksequencepresent in the chimeric NAgene. The chimeric NAgenehasanAin the -3positioninstead of the U found in thewild-typeNARNA (open triangle in Fig. 1). We believe that the twofold reduction in NAactivity doesnot significantlyinfluence the pathogenicity of thevirus, sincewe have constructed
trans-fectant viruses in which the expression of the NA gene is downregulated by a factor of 10 without affecting virus growth in tissue culture (3a). It should be noted that in the transfectant virus, expression of the othersevengenesis not
altered. Specifically, the NA/B-NS virus produces the same level ofHAproteinas found in wild-type virus, and compa-rable amounts of HAare packaged into the envelope ofthe virus. It appears that the attenuation characteristic is best
explained bythe lower synthesis ofNA-specific vRNA and
by the resultinglower representation ofparticles containing afull complement ofthe eight RNAsegments.
How influenza A virus packages its eight RNA genome segments remains an interesting question. In the past, two different mechanisms were proposed for the packaging of
influenza virus RNAs: one suggeststhat theeight RNAsare
selectively packaged, and the othersuggeststhatviral RNAs
arepackaged randomly (3, 8, 20). Evidence isnow accumu-lating to support the random-packaging mechanism. The
idea originated from the fact that influenza viruses have a
lowratio of infectious particlestophysical particles(3, 8). If
one assumes that an average of 11 RNAs are packagedper virion, the expected ratio is compatible with that found in
vivo (6). This modelwas also supported by the findingofa reassortant virus which contained two copies of the same segmentderived from twodifferentviruses(17), andfurther support for this theory came from a recent report which
described aninfluenza A virus which required nineRNAsin
order to be infectious (6). Our data in the present study
concerning the characterization of the NA/B-NS
transfec-tant virus also seem to favor the random-packaging
mecha-nism rather than the selective one. The lower level of
chimeric NA RNA found in virions is consistent with the
reduction of its synthesis in infected cells. In a selective
packaging model, it would be expected that approximately
equimolaramountsofchimeric NA RNA would bepackaged
into virus particles.
In summary, the present data suggest that influenza
vi-ruses for which the synthesis ofavRNA segment is down-regulated produce defective particles during replication.
Since the proteins of this virus areunalteredcompared with
those ofwild-type virus, attenuation must be the result of
inefficientcis-acting signals. We believethat thisprinciple of
attenuation canbe applied to otherviruses with segmented
genomes. For example, the introduction of modifications
into the noncodingsequences ofrotavirusgenes orofgenes
of other segmented double-stranded RNA viruses (16)
should also allow the pathogenicity of these viruses to be
altered. With the influenza virus system, we now plan to identify the nucleotide sequences in the NA/B-NS virus
which are responsible for the regulation of the synthesis of
vRNA and mRNA. This should help us to precisely
deter-A.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.63.294.74.238.2]ATTENUATION OF CHIMERIC INFLUENZA A/B VIRUS 4685
mine the cis elements required for the replication of
influ-enzaviruses.
ACKNOWLEDGMENTS
This work wassupported byNational Institutes of Health grants AI-18998andAI-11823toP.P., a Max Kade Foundation fellowship toM.B.,and a NATO fellowship to A.G.-S.
REFERENCES
1. Brown, F. 1990.Introduction: modern approaches tovaccines. Semin. Virol. 1:1-2.
2. Centers for Disease Control. 1992.Outbreak of influenzaAina nursing home-NewYork,December1991-January 1992. Mor-bid. Mortal. Weekly Rep.41:129-131.
3. Compans, R. W., N. J. Dimmock, and H. Meier-Ewert. 1970. An electron-microscopic studyof theinfluenza virus-infectedcell, p.87-108. In R. D. BarryandB.W. J.Mahy(ed.), The biology oflarge RNA viruses.AcademicPress, NewYork.
3a.Doymaz,M., P. Palese et al.Unpublisheddata.
4. Enami, M., R. Fukuda, and A. Ishihama. 1985. Transcription and replication of eight RNA segments of influenza virus. Virology142:68-77.
5. Enami, M., W. Luytjes, M. Krystal, and P. Palese. 1990. Introduction ofsite-specific mutations into thegenomeof influ-enzavirus.Proc. Natl.Acad. Sci. USA 87:3802-3805. 6. Enami, M., G. Sharma, C. Benham, and P. Palese. 1991. An
influenza virus containing nine differentRNAsegments. Virol-ogy185:291-298.
7. Laemmli, U. K. 1970. Cleavageof structuralproteinsduringthe assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
8. Lamb, R., and P. W. Choppin. 1983. The gene structure and replicationofinfluenzavirus. Annu. Rev. Biochem. 52:467-506. 9. Luo, G., W. Luytjes, M. Enami, and P. Palese. 1991. The polyadenylation signal ofinfluenza virus RNA involves a stretch
of uridines followed by the RNA duplex of the panhandle structure.J. Virol.65:2861-2867.
10. Luo, G. X., and P. Palese. 1992. Genetic analysis ofinfluenza virus. Curr.Opinion Genet. Dev. 2:77-81.
10a.Luo, G. X., and P. Palese. Unpublished data.
11. Luo, G., and J. Taylor. 1990. Template switching by reverse transcriptaseduring DNA synthesis. J. Virol. 64:4321-4328. 12. Luytjes, W., M. Krystal, M. Enami, J. Parvin, and P. Palese.
1989.Amplification, expression and packagingof a foreign gene by influenza virus. Cell59:1107-1113.
13. Murphy, B. R., and R.G. Webster. 1990.Orthomyxoviruses,p. 1091-1152. In B. N.Fields andD. M.Knipe(ed.),Virology,2nd ed. Raven Press, New York.
14. Muster, T., E. K. Subbarao, M. Enami, B. P. Murphy, and P. Palese.1991. AninfluenzaAvirus containing influenzaBvirus 5' and 3' noncoding regions of the neuraminidase gene is attenuatedin mice. Proc.Natl.Acad. Sci. USA88:5177-5181. 15. Parvin,J. D., P. Palese, A. Honda, A.Ishihama,and M.Krystal. 1989. Promoteranalysisofinfluenzavirus RNApolymerase.J. Virol.63:5142-5152.
16. Roner, M. R., L. A. Sutphin, and W. K. Joklik. 1990. Reovirus RNAisinfectious. Virology 179:845-852.
17. Scholtissek, C., W. Rohde, E. Harms, R. Rott, M.Orlich, and C. B. Boschek. 1978. A possible partial heterozygote of an influenzaAvirus.Virology 89:506-516.
18. Seong, B. L., and G. G. Brownlee. 1992. A new method for reconstitutinginfluenza polymerase andRNA in vitro:astudy of the promoter elements for cRNA andvRNA synthesis in vitro and viral rescue in vivo. Virology186:247-260.
19. Shapiro, G.I.,T.Gurney, Jr., and R. M. Krug. 1987.Influenza virus gene expression: control mechanisms at early and late times of infection and nuclear-cytoplasmictransport of virus-specificRNAs. J. Virol.61:764-773.
20. Smith, G. L., and A.J. Hay. 1982.Replicationof the influenza virus genome.Virology 118:96-108.
VOL. 66,1992