Copyright ©D 1985,American Society for Microbiology
A
1.6-Kilobase-Pair
Fragment
in the Genome
of the tsl Mutant of
Moloney Murine Leukemia Virus
TB
That
Is
Associated
with
Temperature
Sensitivity,
Nonprocessing
of Pr80env,
and
Paralytogenesis
P. H. YUEN, D. MALEHORN, C. KNUPP, ANDP. K. Y. WONG*
Department of Microbiology and SchoolofBasicMedical Sciences, University of Illinois, Urbana, Illinois 61801
Received 13 November1984/Accepted 21 January 1985
tslandtS7, twotemperature-sensitive mutants of Moloney murine leukemiavirus strain TBinducehind-limb
paralysis in 100% ofCFW/D mice injected. These two paralytogenic mutants also share a defect in their
inability to process the env precursor protein, Pr8Oe"v, at the restrictive temperature. To identify the
mutation(s) in the genomes of the paralytogenic mutants whichcausetheinabilitytoprocessPr8Oe"v efficiently
and confer the ability to cause hind-limb paralysis instead of lymphoma, we constructed chimeric genomes
between tsl and Moloney murine leukemiavirus or the TB strainofthevirus.Weidentifieda3.9-kilobase-pair
HindIII-PstI sequence from nucleotides 4895 through 8264 and 1through567oftsl, comprising the3' end of
thepol and all of the env genes, the long terminal repeat, and the 5' noncoding sequence, as being responsible
for the temperature sensitivity, the inefficiency in processing Pr8Oe"v, and the induction of paralysis. We
extendedthese findings by demonstrating that the 1.6-kilobase-pairpol-gp7O HindIII-BamHI DNAsequence
fromnucleotides4895 through 6537 oftsl within the3.9-kilobase-pairHindIII-PstI fragment is necessary for tsl toinduceparalysis.Inaddition,we showed thatthis1.6-kilobase-pair fragment also controls theprocessing
of Pr8Oe"v and thetemperaturesensitivity oftsl.
Murineleukemia viruses(MuLVs)are acomplex groupof viruses with the ability to induce a variety of diseases in
mice. For example, Moloney MuLV (MoMuLV) induces T-cell lymphomas (9), and members of the Friend virus
complex induce erythroleukemias. On three separate
occa-siuns we have isolated from MoMuLV-TB, a strain of
MoMuLV, a group of temperature-sensitive mutants, tsl,
ts7, and tsll,
which
induce hind-limb paralysis in 100% ofCFW/D mice injected with them. This is in contrast to the
induction of lymphoma byMoMuLV-TBin the same mouse
strain. Inductionof paralysis has also been reported for the
Cas-Br-E strain of MuLV, whichwasinitially isolated from
the brain of a paralyzed wild mouse (for a review, see
reference 3), and for four clones of MuLV isolated from a
paralyzed Fischerrat whichhad beeninfected with
rat-pas-saged Friend leukemia virus (5). The mechanism by which
theseneurotropic viruses induce paralysis is still unknown.
The tsl group ofmutant viruses is especially suited for
investigations intothe molecularbasis of this
retrovirus-in-duced neurological disease because (i) these paralytogenic
mutants were derived from a nonparalytogenic parent, so
that thechange (ormutation) in the gene responsible for the
induction ofparalysis
could
be identified by comparing thegenome ofthe mutants with that ofthe parent, (ii) the tsl
groupofmutants seemtobemorepotentthan theCas-Br-E
strain of MuLV (they cause paralysis in every one of the
mice injected, and their latent period is relatively shorter
than that ofthe Cas-Br-E strain of MuLV); and (iii) this
groupofparalytogenic mutants share a common
character-istic in their inability to process the precursorPr8Oe'7'(20). If
the accumulation ofPr80e'l' polypeptides can be correlated
with theinductionof paralysis in vivo, it will provide a basis
*Correspondingauthor.
for further investigation into the molecular mechanism of
this retrovirus-induced paralysis.
Toidentifythe mutation(s)inthe genomesof the
paralyto-genic mutants which confer temperature sensitivity,
ineffi-cientprocessing of
Pr8Oehz',
and theabilityto causehind-limbparalysis, we constructed chimeric genomes between tsl
and MoMuLV or MoMuLV-TB. Using this approach, we
have found that replacing the pol-gp7O 1.6-kilobase-pair (kbp) HindIII-BamHI DNA sequence from nucleotide (nt) 4895 to nt 6537 oftsl with thehomologous sequence from
either MoMuLV or MoMuLV-TB was sufficient to correct
temperature sensitivity and inefficiency in the intracellular
processing ofPr8Oe'7' in tsl-infected cells. In addition, the
constructsalsofailedtoinducehind-limbparalysis.
Further-more, hybrid viruseswhose genomescontained
the
1.6-kbpHindIII-BamHI tsl sequence from nt 4895 to nt 6537
re-mained temperature sensitive and inefficient in processing
Pr8Oe"'. This 1.6-kbp tsl subgenomic fragment is also
nec-essaryfor the induction ofhind-limbparalysis. The
follow-ingis areport onthese studies.
MATERIALSAND METHODS
Cells. Viruses werepropagated in mouseTB
cells,
athy-mus-bone marrow cell line derived from CFW/D mice (1),
andassayedon 15Fcells,amurinesarcoma-positive,
leuke-mia-negative cell line (23). NIH 3T3 cells were used for
transfection experiments. All cell lines were maintained in
Dulbecco modified Eagle medium supplemented with 8%
fetal calfserum.
Viruses. The strain of MoMuLV used in this study, MoMuLV-TB,wasisolatedasdescribedby Wongetal.
(19)
from the tissue extract of a sarcoma produced in an
MoMuLV-infected BALB/c mouse provided by Dr.
Mo-loney. Since itsisolation, it has beenpropagatedin TB cells. MoMuLV-TB has been single-virus, single-cell cloned on
severaloccasions. CloneLV30, usedin the present
studies,
364
on November 10, 2019 by guest
http://jvi.asm.org/
A R U5 R US U3 III P15
p121|-p30--.plOk-
pOl*-i--
9p70-4.Pl5E
-1IU3
lUlf
0 1 2 3 4 5 6 7 8 Kb
P Sm K PSA Bg SXHPSM Kp S
p
1) ( (\ P Xh o A SC K SP H x 9BC >X
Sc Sc
BSc A
PXh A Sc K \ SP
1 I1
B0 BgaKSm Sm Bg K
Xh
BScA
PXh AB Sc K \\SC SP II 1111
11S
p Sge eSmx Sm
II
I
H LTR H DNA
k
.H
pKC7
II
PS
HLTR
+ HindIII
PstI
PS
Ps
pUC9
+ HindIII
H
J
Pstl
H Ps
I _ I HI PS
+ ligase
H-C +HindIII
La S;m
n
liga
H
Sm H
+ HindlII
ft
Pst
IH H PS H PS
A4
1,Is
FIG. 1. (A)EndonucleaserestrictionmapsofMoMuLV(I),MoMuLV-TB(II),and tsl(III). (B)Schematicpresentationofthe molecular
cloningof thetsl, MoMuLV,andMoMuLV-TBSmaI-HindIl sequencefromnt31to nt4894 intopKC7 (I) and the HindIII-PstIsequence
fromnt4895to nt8264 and nt1to nt567intopUC9 (II). Detailsaredescribedinthetext. (C) Constructionofchimericgenomeswith the
HindIII-PstIsequenceof tsltotheHindIII-SmaI sequencesofMoMuLV-TB and MoMuLVand-viceversa.Detailsaredescribed in thetext.
Abbreviations: A, Aval; B, BamHI; Bg, BgII; C,ClaI; H, HindlIl; Hp, HpaI; K, KpnI; P, Pi'uII; S,Sall; Sc,Sacl; Sm, SmaI; X, XbaI; Xh,XhoI.
p Sm PS
P Sm PS
~\) (K \
Sc
B
K p Sm
a () Hp C(tX K
K p Sm
el
>pHr
C (')I
gSc
I i t
rl==--
1---- I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.84.508.88.654.2]was one of the isolates obtained in the most recent clonal
isolation. It hasbeen shown by Shields and co-workers (14) and confirmed by us (unpublished data) that MoMuLV-TB
can be distinguished from standard MoMuLV by the
elec-trophoretic mobilityof the p30protein.Inthis report the two
strains ofMoMuLV will be referred to as MoMuLV and MoMuLV-TB.
tsl is a spontaneous temperature-sensitive mutant of
MoMuLV-TB isolated as described by Wong and
co-workers (19). The tsl strain used in the present studies was also recently purified by cloning TB cells infected with tsl at amultiplicity of infection of 0.01.
Virus assay. The 15F virus assay has been described
previously (18, 21).
Viral DNAs. The permuted genomesof tsl and
MoMuLV-TB were molecularly cloned into Charon 21A as described
by Yuen et al. (22). The infectious recombinant viral DNAs
tsl-19, tsl-20, and wt-25, each with one copy of the long
terminal repeat (LTR), were used in the studies reported here. Plasmid p8.2, an infectious permuted MoMuLV
ge-nome with one copy of the LTR cloned into pBR322 at the
HindIll site (12), was a gift from D. Baltimore. The viral
DNA ofp8.2is referredto as wt-8.2.
Isolation,purification, and analysis of viral DNAs.
Restric-tion fragmentswere separated byagarosegel electrophore-sis. The desired fragments were electroeluted into dialysis bags. The DNA was filtered through glass fiber to remove
agarose debris and then concentrated by ethanol
precipita-tion. The DNA pellet was suspended in TE buffer(10 mM
Tris,1 mM EDTA,pH 8.0) and then extracted with
phenol-chloroform, phenol-chloroform, and ether.
Restriction enzymes were purchased from Bethesda
Re-search Laboratories, Gaithersburg,Md., and the conditions
forendonuclease digestionwerethose recommended bythe
supplier. Gel transfer and filter hybridization were carried
outbythe method of Southern (16).
DNA transfection. The calcium-phosphate precipitation
method of
Graham
and van der Eb (4) was used. Details oftheprocedures were as described in Yuen et al. (22).
Recombinant plasmid and chimeric genome construction.
Restriction fragments were cloned into pUC9 (17), pKC7
(10), or pBR322. Transformations into Escherichia coli
HB101 werecarriedoutby standardprocedures. Construc-tion of recombinant plasmids and chimeric genomes is describedbelow.
Metabolic labeling, immunoprecipitation, and SDS-PAGE.
Immunoprecipitation of intracellular virus-specific proteins
and sodium dodecyl sulfate-polyacrylamide gel electropho-resis (SDS-PAGE)were performed asdescribed previously (20,21). Gelswerefluorographedand exposed to X-ray film at -70°C.
Mouse strain andinoculationprocedure. Theinbred CFW/D
mice and the inoculation procedureused in this studywere
described in Yuen et al. (22).
RESULTS
Construction of chimeric genomes between molecularly
cloned tsland MoMuLV or MoMuLV-TB DNAs. The
restric-tion maps of themolecularly cloned tsl, MoMuLV-TB, and
MoMuLV,
designated
tsl-20, wt-25, and wt-8.2,respec-tively, are shown in Fig. 1A. These molecularly cloned
genomes,
together with tsl-19, a tsl genome obtained in aseparate cloning experiment (22), were used in the studies
reported here.
As a first step in identifying the mutation(s) in the tsl
genome responsible forthe temperature-sensitive function,
1 2 3 4 5 6 7 8 9 111213141516.1718 Kb
23-4.3
2.3_- 2.0- 1.3-
0.6-FIG. 2. UV fluorescence photograph of the ethidium bromide-stained gel of the endonuclease digests of wt-25, p14, p12, and p18. The DNAs(restriction endonuclease) used in each lanewere: 1and 11,wild-type A(Hindlll)and4X174 (HaeIII);2, wt-25, a recombi-nantCharon21Awith thefull-sizeMoMuLV-TB genome(HindIII and PstI);3, 3.9- and4.1-kbp fragments of wt-25 (PstI);4, pUC9
(HindIII); 5, p14, a recombinant pUC9 plasmid with the 3.9-kbp
HindIII-PstIsequenceofwt-25(HindIII and PstI); 6,p14 (HindlIl); 7, p14 (HindlII and XhoI); 8 and 9, p14and pUC9, respectively
(HindIII andBamHI);12, pKC7(HindIll); 13,p12,apKC7
recom-binant plasmid with the full-size wt-25 genome (HindlIl); 14, p12 (SmaI); 15, p18, a pKC7 recombinant plasmid with the 4.9-kbp SmaI-HindIIIwt-25 DNAsequencefromnt31 to nt 4894(HindlIl);
16,p18 (SmaI);17,p18 (HindlIl andSmaI);18and,p18 and pKC7,
respectively (KpnI andHindIll).
the inefficiency in processing Pr80en,', and the ability to
induce paralysis in mice, chimeric genomes were
con-structed by exchangingthe entire env gene between tsl and
the wild-type virus genomes. Since the permuted tsl,
MoMuLV-TB, and MoMuLV proviruses were cloned into
Charon 21A or pBR322 at the HindIll site, we decided to
fragment each genome into halves: (i) a HindIII-PstI
frag-mentfromnt4895 to nt 8264andnt 1 to nt567,consistingof
the3' end of thepol gene, the entire env gene, the LTR, and
the amino-terminal portion of the gag gene (5' noncoding
sequence), and(ii)aSmaI-HindIllfragmentfrom nt 31 to nt
4894, consistingof part of the Rregionof theLTR, U5,the
gaggene, and the major portion of thepolgene.
Molecular cloning ofthe HindIII-PstI sequence from nt
4895 to nt 8264 and nt 1 to nt 567 oftsl-19, tsl-20, wt-25,and
wt-8.2isshown inFig. 1BII.Thevarioussteps in the actual
cloning of p14, pUC9 recombinant plasmid with the
HindIII-PstI sequence ofwt-25,arepresented inFig.2.The
permuted genome ofwt-25 was restricted from its Charon
21A vector atthe HindlIl site and further cleaved with PstI
(lane 2). The purified 3.9-kbp fragment from nt 4895 to nt
8264andnt1 tont567 and the4.1-kbpfragmentfromnt744
to nt 4894of the PstI-restricted wt-25 genomeare shown in
lane 3. These DNA sequences were not separated before
ligation with PstI-HindIII-restricted and dephosphorylated pUC9 DNA (lane 4). The ligated DNAs were transformed
intoE. coli HB101. Recombinants with the HindIII-PstI 3'
halfof the viral genome could bedistinguished bythesize of
the viral insert, which was about 3.9 kbp (lane 5), the
absence of an XhoI site(lanes6and7), and thepresence of
asingleBamHI site. Due to the presenceofaBamHI site in the polylinkerofpUC9 10 ntfrom theuniquePstI site, the
viral HindIII-PstI fragment was cleaved into two
subfrag-ments of about 1.6 and 2.3 kbp after BamHI restriction
(lanes 8 and 9). Recombinant plasmids pl, p5, and p15,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.323.562.70.215.2]+ ligase
J./
H X B LTRPs
L _ ~~p15
x x
p14
Sm
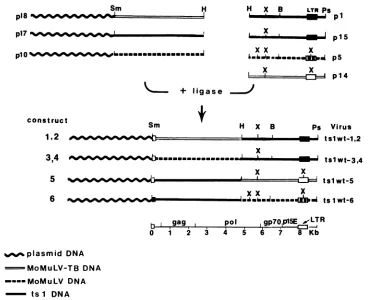
construct
1,2
3,4
5
6
gag
0 1 2
H X B
x
'WI _V ...I . . .o. m
ub
mmm mommmmmm---x x
__s- --_ _ _1 _ _ WE, Xx x
Ps Virus
IJ tslwt-1,2
tSlwt-3,4
tslwt-5
ts1wt-6
pol
gp7OI1L5E
wLTR3 4 S 6 7 8 Kb
plasmid DNA -MoMoNuLV-TB DNA
----MoMuLV DNA
ts1 DNA
FIG. 3. Chimeric viral genomes constructed from the 3' pol-env-LTR and the 5' U5-gag-pol sequences of tsl, MoMuLV-TB, and MoMuLV. Infectious viruses tslwt 1 and tslwt 2 were recovered from NIH 3T3 cellstransfected with constructs 1 and 2, which were
constructed by ligating the SmaI-HindIll sequence ofp18 (MoMuLV-TB) to the HindIII-PstI sequence ofpl (tsl-19) or p15 (tsl-20), respectively. Infectiousvirusestslwt 3 andtslwt 4wereproducedwhenconstructs3 and4,obtainedbyligatingthe SmaI-HindIIIsequence ofplO (MoMuLV) with theHindIII-Pstlsequenceofpl(tsl-19)orp15 (tsl-20),respectively,weretransfected into NIH 3T3 cells.Infectious
viruses tslwt5and tslwt 6wereobtainedby transfecting the reciprocalconstructs5and6, generated by ligatingtheSmaI-HindIIIsequence ofp17 (tsl-20) and the HindIII-PstI sequenceofp14(MoMuLV-TB)orp5(MoMuLV), respectively. Abbreviations:B,BamHI; H, HindIll; Ps,PstI; Sm, SmaI; X, XbaI.
carrying the HindIII-PstI sequence oftsl-19, wt-8.2, and
tsl-20, respectively, wereobtained similarly.
Molecular cloning of the SmaI-HindIIl nt sequence of
tsl-20, wt-25, and wt-8.2 is shown inFig. 1BI.The
proxim-ity of the two PstI sites at the 5' end of the gag coding sequence necessitated adifferent strategy for clonally
puri-fyingthegag-polhalves of the genome sothat the full-size
genome could be restored when the cloned 5' halves were
ligated tothe HindIII-PstI 3' halvespreviously cloned(Fig.
1C). The permuted genomes of tsl-20, wt-25, and wt-8.2
wereisolatedandpurified from theirrespectivevectorsand
cloned into pKC7 at the HindIII site. The recombinant
plasmids obtained were restricted with SmaI to determine
the orientation of the viral insert. Recombinant plasmids
with the viral insert in the desired orientation (Fig. 1B la)
werecharacterized bythepresenceofa9.5-kbp SmaI-SmaI
fragment, which consisted of the viable portion of pKC7,
including the ampicillin resistance gene, and the 4.9-kbp
SmaI-HindIII 5' half of the viral genome. Therefore, to isolate the 4.9-kbp SmaI-HindIII viral sequence,
recombi-nantplasmidswiththecomplete viral genomein the correct
orientation were restricted with SmaI, and the 9.5-kbp
fragment was isolated, purified, religated at the SmaI site,
and transformed into E. coli HB101. Ampicillin-resistant colonies were screened for the presence of the 9.5-kbp
plasmids. Recombinant plasmids containing the
SmaI-HindIII viral sequence were then purified for chimeric
genome construction.
Restriction analysiswasperformedonp12, arecombinant
plasmid with the permuted wt-25 genome in the desired
orientation, andp18,adeletion mutantofp12withonlythe
SmaI-HindIll 5' half of the wt-25 genome (Fig. 2, lanes 12
through 19). The full-size wt-25 genome restricted from its
pKC7vector attheHindIIIsites isshown in lanes 12 and 13.
OnSmaIdigestion, p12wasfragmentedintoalarge 9.5-kbp
andtwosmallerfragmentsofabout 2kbpeach(lane 14).The presenceofunique HindIII (land 15)andSmaI(lane 16)sites in deletion mutant p18 caused it to be cleaved into two
fragmentsof about 4.8 kbpeach afterdoubledigestion with
SmaI and HindlIl (lane 17). AfterHindIII and KpnI
diges-tion, p18wascleavedinto threefragments, consistingof the
deleted pKC7 vector with a few nt from the virus genome andtwo viral sequencesof about2.0 and 2.9kbp (lane 18).
pKC7 did not have a KpnI site (lane 19). Recombinant
plasmids plO and p17, with the SmaI-HindIII sequence of
wt-8.2 and tsl-20, respectively, were similarlyisolated. The chimeric genomes between tsl and MoMuLV or MoMuLV-TB were constructed asshown in Fig. 1C. Each of these constructs (Fig. 3) had an overlapping sequence from theSmaI siteat nt30tothe PstI siteat nt567,derived from both tsl and MoMuLV or MoMuLV-TB DNA. The
ligated DNAs were transfected into NIH 3T3 cells without
pl8
p17
Sm H
_ -- ___0-_ - I_l
40--i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.124.492.69.369.2]further treatment. Infectious viruses obtained from each
construct werecharacterized. The mechanism for generating
the infectious viruses after transfection of these hybrid
DNAs into NIH cells is not known. Although a complete
LTRsequence is at the 3' end, only part of R and the entire
US region are present at the 5' end of the genome. A
recombination of theoverlapping region (nt 30 through 567),
which is duplicated in the constructs to generate the
pro-moter atthe 5' endofthe genome, may be necessary. If this
is the case, the parental origin of the overlapping region in the resulting viruses may vary from virus tovirus.
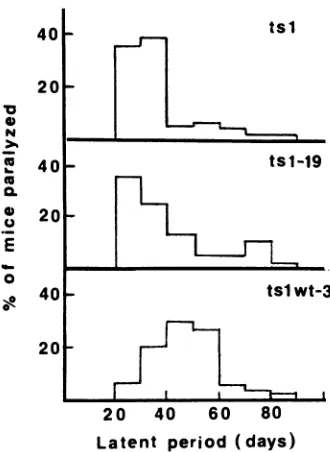
Characterization of the infectious reconstructed viruses.
The reconstructed viruses tslwt 1, 2, 3, 4, 5, and 6 were
testedfortemperature sensitivity, abilitytoprocess Pr80en'
at the restrictive temperature, and ability to induce
hind-limb paralysis in mice (Table 1). tslwt 1, 2, 3, and 4,
producedby the constructscomposed ofthe 3' halfofthe tsl
genome and the 5' half ofthe MoMuLV or MoMuLV-TB
genome, remained temperature sensitive. NIH 3T3 cells
transfected with any of these four viruses accumulated
Pr80en", with protein profiles indistinguishable from that of tsl-infectedcells (see Fig. 8). In addition, 100%of the mice
injected with tslwt 1, 2, 3, or 4became paralyzed. The latent
period oftslwt 3 was also examined because ofthe large numberofmiceinjected with this virus. More than80% of
themice becameparalyzed by60dayspostinjection (Fig. 4), suggestingthat tslwt3 may have alonger latentperiodthan
eitherthe cloned or noncloned tsl. The significance ofthe
apparentlyextended latentperiodshownbytslwt3 needs to
be investigated further.
Incontrast, tslwt 5 and 6, produced byconstructs
com-posed ofthe 5' halfofthe tslgenome and the 3' half ofthe
MoMuLV-TB orMoMuLV genome, respectively, were not
temperature sensitive and processed Pr80en' like wild-type
virus-infected cells (see Fig. 8). In addition, tslwt 5 and 6
failed to inducehind-limb paralysis in mice. Of the CFW/D mice infected with tslwt 6, 100% died oflymphoma 3 to 5
monthspostinjection. Noneof the 10CFW/Dmice infected
withtslwt 5 showed any sign of disease atthistime (97 days
postinjection). Therelativelyshort latentperiodrequired by
tslwt 6 to induce lymphoma is in agreement with that
reported for the standard MoMuLV (9) but in contrast to
[image:5.612.356.521.71.297.2]that reported forMoMuLV-TB (7).
TABLE 1. Characterization of virusesproduced by chimeric genomes intransfectedNIH-3T3cellsa
Virus
(34aC/39aC)y
Titer ratio processingbPr80e"' No. of mice %of mice injected paralyzed'wt-25d 1.4 P 20 0
tsll9d 3.0 x 103 NP 28 100
tslwt 1 1.5 x 103 NP 16 100
tslwt2 2.5 x 103 NP 14 100
tslwt 3 1.2 x 102 NP 52 100
tslwt4 1.4 x 102 NP 22 100
tslwt5 4.3 P 10 0
tslwt 6 1.2 P 22 0
aAppropriatelydiluted virus was inoculated into two setsofplatesseeded with 15F cells and allowed to adsorb for 45 min at34°C.One setofplateswas
incubatedat 34°C and the other at 39°C. The assay was read 4 to 5days postinfection.
bInfectedcells grown at39°Cwerepulsedfor 15 min with[35S]methionine
andchased for 3 h. Cell extracts wereimmunoprecipitatedwithantiserumto MoMuLV gp7O, analyzed by SDS-PAGE, fluorographed, and exposed to Kodak X-ray film. P, Pr80M processed togp7Oandp1SE;NP, Pr80M""not
processed.
cCFW/Dmice within 48 h of birth were inoculated intraperitoneallywith
105to106 infectiousvirus in 0.1 ml ofgrowthmedium. d Parentalvirus.
40
20
~0
N
(U
E
-0
0
40
20
40-tsl
ts1-19
tsl wt-3
201
20 40 60 80
Latent period
(days)
FIG. 4. Histogram ofthe percentage of mice paralyzed by the
non-molecularly cloned tsl, the molecularly cloned tsl-19, and hybridvirustslwt3, with the indicated latentperiods.
The above results indicated that the 3' half ofthe tsl
genome contained the sequence(s)
responsible
fortheinef-ficiency
inprocessing
Pr8Oenl' atthe restrictivetemperature,the induction of paralysis, and the
temperature-sensitive
function.Theobservationthat tslwt1, 2, 3, and 4,
although
possessing
theU5and the 5'portion
ofthe gag geneofboth tslandwild-typegenomes,remainedphenotypically
like tslsuggeststhat thisoverlapping
portion
ofthe constructfromthe SmaI site at nt30 to the PstI site at nt 567 may not be
involved in producing the
phenotypic
differences observedbetween tsl and wild-type virus.
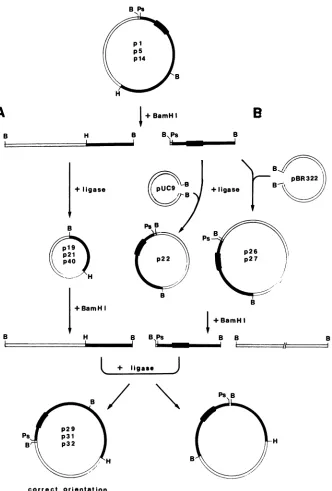
Constructionof chimericHindIII-PstI sequences of tsl and
MoMuLV-TB or MoMuLV. To further delimit the DNA
sequence of the tsl genome which confers temperature
sensitivity,
inefficiency inprocessing Pr80ens,
andability
toinduce paralysis, chimeric genomes between tsl and
MoMuLVorMoMuLV-TB (see
Fig.
7) were constructed.The isolation and purification of the HindIII-BamnHI and
BamnHI-PstI sequences of the tsl, MoMuLV-TB, and
MoMuLV genomesareshowninFig.5and 6. Todistinguish
constructs p29 and p32, which consist of a
wild-type
HindIII-BamHI sequence and a tsl BamHI-PstI sequence,
from constructp31, which consists ofatsl HindIII-BamHI
sequenceandawild-typeBamHI-PstI sequence
(Fig.
7), thechimeric HindIII-PstI
fragments
wererestricted with XbaI.We have previously shown (22) that the HindIII-PstI
se-quencesoftsl,MoMuLV-TB,and MoMuLV may be
distin-guishedbyrestriction with XbaI. tsl has lost theXbaI siteat
nt8113 in the U3regionof MoMuLV-TB and MoMuLV.In
addition, the XbaI site at nt 5325 in the
carboxy-terminal
portionof thepolcoding
sequenceof the MoMuLV genomeis absent in both thetsl and MoMuLV-TB genomes.
pUC9
had noXbaI restriction site(Fig.
6, lanes 8 and9).
Whereas
p31
(lanes 10and 11)wascleavedby
XbaI intotwofragmentsof about 2.3 and4.4
kbp,
XbaI restrictedp29 (lane
12) into two fragments of about 6.3 and 0.4kbp
andp32
(lane 13) only once.To restore the complete genome, the HindIII-PstI viral
sequences isolated from p29,
p30,
andp32
wereligated
toI I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.62.301.531.630.2]B Ps
p1
p5
p14
B
A +BamHI
B H B B\Ps B
B
pBR322
+ligase pU9 +ligase
'B
B Ps B
p~~ ~ ~ ~ ~~~B p26
p4 p22 p27
B
B
I+BamH I
+ ligase )
Ps B
B
p29 Ps p31
B p32
~~~~B
[image:6.612.138.470.81.572.2]correct orientation
FIG. 5. Schematicpresentationof themolecularcloning of the HindlIl (H)-BamHI (B)(panel A)and BamHI-PstI(Ps) (panel B)sequences
oftsl,MoMuLV-TB, and MoMuLV, and the construction ofachimericHindIII-PstI viralsequence.Recombinantplasmidspl,p5,andp14, containingthe HindIII-PstIviralsequencesoftsl, MoMuLV, and MoMuLV-TB, respectively,wererestrictedwithBamHI, generatingtwo
fragments of about 2.2 and 4.1 kbp (see Fig. 6, lane 2). The 2.2-kbpfragment consists of the BamHI-PstI viralsequenceand 10ntfrom the polylinker of pUC9. The4.1-kbp fragment consists of the viral HindIII-BamHIsequenceandpUC9. The4.1-kbp fragmentwasreligatedat
the BamHI site, transformed into strain HB101 cells, and generated recombinant plasmid p19 from pS, p21 frompl, and p40 from p14. The 2.2-kbp fragment derived from pl wascloned into pUC9at the BamHI site, and the 2.2-kbp fragment from p5 and p14 was clonedinto
pBR322, alsoatthe BamHI site. The recombinantplasmid with the BamHI-PstIsequencefromtslwasdesignated p22, and the recombinant
plasmids with the MoMuLV and MoMuLV-TB BamHI-PstI sequences weredesignated p26 and p27, respectively. Recombinant plasmids p22, p26,andp27wererestricted with BamHI.TheBamHI-BamHI fragment containing the BamHI-PstI viralsequence wasseparated by agarose gel electrophoresis and isolated by electroelution. Recombinant plasmids p21, p19, and p40, carrying the HindIII-BamHI viral sequence of tsl, MoMuLV, and MoMuLV-TB, respectively, were restricted with BamHI and dephosphorylated with calf intestinal phosphatase.TheBamHI-BamHI tslsequencefromp22wasligatedtop19orp40, and after transformation into strain HB101 cells generated the6.7-kbp recombinants p29 and p32. Similarly, recombinant plasmid p31wasconstructedby ligating p21tothe BamHI-BamHI fragment
of MoMuLV.
on November 10, 2019 by guest
http://jvi.asm.org/
1 2 3 4 5 6 7 8 910111213
94 -6.6 -4.3
1.3
-0.6
-FIG. 6. Endonuclease restriction analysis of cloned viral se-quences. Lanes: 1, HindIll digest of wild-typeXandHaeIII digest of 4X174DNAs; BamHIrestriction ofpl(lane 2),p21 (lane3), p22 (lane4), pUC9(lane 5), p27 (lane 6), and pBR322 (lane 7); pUC9 restricted with HindIll (lane 8) and doubledigested with Hindlll and XbaI (lane 9); XbaI restriction of p31(lanes 10 and 11), p29 (lane 12),and p32 (lane 13). Sizes are shown in kilobases.
the HindIll-restricted and dephosphorylated plasmid plO,
p17, or p18(recombinant plasmidswith theSmaI-HindIII
5'-U5-gag-pol sequence of MoMuLV, tsl, and MoMuLV-TB,
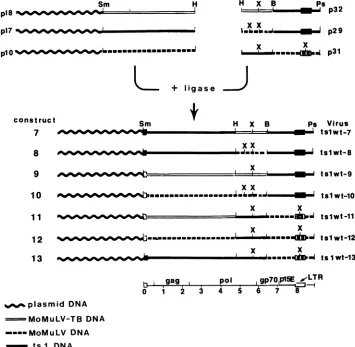
respectively) to generate the constructs shown in Fig. 7.
pl8
p17
p10
Sm H
- -Jwwww^""
L
TABLE 2. Characterization of viruses produced by chimeric genomesintransfectedNIH 3T3cellsa
VTiterratio Pr80en%,
No.
of % ofVirus
(34TC/390C)
processing mice miceinjected paralyzed
tslwt 7 2.1 P 12 0
tslwt 8 2.5 P 62 0
tslwt 9 1.8 P 27 0
tslwt10 1.6 P 24 0
tslwt 11 100-1,000 NP 20 NAb
tslwt 12 100-1,000 NP 10 NA
tslwt13 100-1,000 NP 22 PAC
a See Table 1, footnotes a and b. Data for the parental viruses were the
same asin Table1.
bNA, Notavailable. Mice 40 to 120 days postinjection with tslwt 11 or 12 haveasyet shownnosignofparalysis.
C PA,Partially available. Of 22 mice injected with tslwt 13, 4 have begun to showsignsofparalysis.
Infectious viruses
produced
aftertransfection oftherecon-structedviralgenomes were
designated
tslwt 7through
13.Characterization ofinfectious viruses tslwt7
through
13. The infectious viruses tslwt 7 through 13 were tested fortemperature
sensitivity, efficiency
inprocessing
Pr80env atthe
nonpermissive
temperature, andability
to induce hind-limb paralysis (Table2).
tslwt7,
8, 9, and 10 were not temperature sensitive and processed Pr80eMvintracellularly
H X B
.-x .-x
= p32
. p29
x-ouD-OJ.X p31
+
ligase
4f
H X B_
_.
_X X
._ -1
xx
-- -0%0 I aumummmmmun~mu
,1 _
J--!
tslwt-8_-' tslwt-9
ml tslwt-10
x x
- - 04-04-04 IL I. .IdmmmuuuJ
x X
'W_N- _- - -
--L---tslwt-11
tslwt-12
....
X.&1
tsIwt-13b gag pol gp70p15E, LTR
0 1 2 3 45 7 8
las0plasmid
DNA-MoMuLV-TB DNA
----MoMuLV DNA
ts1 DNA
FIG. 7. Schematic presentationofconstructs tslwt 7through 13. B,BamHI; H, HindIll; Sm, SmaI; X, XbaI; Ps, PstI.
construct
7
Sm s Virus
tslwt-7 8
9 10
1 1 12
13
Pe
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.113.259.73.211.2] [image:7.612.322.564.95.194.2] [image:7.612.129.484.360.707.2]1 2 3 4
[image:8.612.116.239.75.247.2]Pr80O W
5VT
_gp7O
FIG. 8. Metabolic labeling and immunoprecipitation with anti-gp7O ofintracellularviralproteins of cellsinfected withtsl(lane 1),
tslwt 8fromtwo separatetransfectionexperiments (lanes 2 and3),
and MoMuLV-TB (lane4).
at the nonpermissive temperature. The intracellular viral
proteins ofcells infected with tslwt 8, a representative of
these constructs, were immunoprecipitated with anti-gp7O
(Fig. 8). At thetimeofwriting,miceinjected withtslwt7, 8, 9,or10were130to150dayspostinjection,andnoneof them
had become paralyzed. In contrast, tslwt 11, 12, and 13
remainedtemperature sensitive andfailedtoprocessPr80en'
intracellularly atthenonpermissive temperature.Theresults
ofimmunoprecipitation were similar tothose shown in Fig.
8 andthereforeare notshown. Atthetime of writing, 4 of 22
CFW/D mice 39to95days postinjectionwith tslwt 13, have
beguntoshowearlysignsof paralysis. However, mice 40to
120 days postinjection with tslwt 11 and 12 have as yet
shown no sign of paralysis. It is possible that the latent
periodinthese hybrids isprolonged. Thedifferencebetween thegenomesoftslwt13andtslwt11or12isthat thegag-pol
region oftslwt 13 is derived from tsl, whereas the same
genomic segment of tslwt 11 and 12 is derived from
wild-type virus. Whetherthissegmentof thegenome playsarole
inmodulating theonsetofparalysis in tslwt 11 and 12 needs
to be investigated further.
DISCUSSION
Toidentify the mutation(s) in thegenome of the
paralyto-genic mutant tsl which confertemperature sensitivity,
inef-ficiency in processing Pr80enl', and the ability to cause
hind-limb paralysis, we constructed an assortment of
chi-meric viral DNAgenomes between the molecularly cloned
genome of tsl (22) and those of its parental wild-type MoMuLV-TB or MoMuLV. Viruses obtained from these
chimeric genomes were tested for their temperature
sensi-tivity, ability to process Pr80en', and ability to induce
hind-limb paralysis in mice. We showed that the
HindIll-PstI sequence from nt 4895 to nt 8264 and nt 1 to nt 567, which comprises the 3' end of the pol gene, all ofthe env
gene,andthe LTR, containsthe sequence(s)responsible for
the temperature-sensitivefunction, the inefficiency in
proc-essing Pr80env at the nonpermissive temperature, and the
induction of paralysis in mice. These results are in
agree-ment with the observations ofDesGroseillers and
co-work-ers(2) thatthe 3.9-kbpSaIl-ClaI fragment comprising the 3'
endofthepoland all ofthe env sequenceofthe neurotropic
Cas-Br-E MuLVwassufficienttoconferthe
paralysis-induc-ing
potentialto the virus.We have extended the above studies and showed that
constructs with the 1.6-kbp HindIII-BamHI sequencefrom
nt 4895 tont 6537 of either MoMuLV orMoMuLV-TB are
nottemperaturesensitive, process Pr80e"vininfectedcells at the restrictive temperature, and do not cause hind-limb paralysis. This HindIII-BamHI sequenceconsists of the 880
base pairs (bp) of the carboxyl terminal ofthe pol coding sequence and 660 bp ofthe amino-terminal portion ofthe
gp70 coding sequence. Conversely, hybrids with the
HindIII-BamHI sequence derived fromtslremained
temper-aturesensitiveandinefficient in theintracellular processing
of Pr80eMv at the restrictive temperature. At the time of
writing, 4 of 22CFW/D mice 60 dayspostinjection withone
of these hybrids, tslwt 13, have begunto showearlysigns of paralysis.
Our observations clearly indicate that the 1.6-kbp
se-quence not only controls the processing of Pr80env and
encodes the temperature-sensitive function, but is also
nec-essary for the induction of paralysis.
Restriction mapping of tsl, ts7, and the wild-type
MoMuLV-TB genomes with 11 endonucleases failed to
show any difference in the restrictionpattern of the 1.6-kbp
HindIII-BamHI sequence. It is entirely possible that the
phenotypic differences may have resulted from mutations
which were not detected at this levelofrestriction analysis.
Alterations in a single base which lead to a change in
phenotype and lossofinfectivity have beenreported(8). To
determine whether one or more mutations haveoccurred in
the tsl genome, acomparison of the nt sequences between the 1.6-kbp HindIII-BamHI sequences of tsl and those of MoMuLV-TB is now inprogress. Ifmorethanone mutation is found, chimeric genomescould beconstructed by exchang-ing subfragments of theHindIII-BamHI sequence.
In vivo studies (submitted forpublication) oftsl-injected CFW/D mice showedthatthe titer ofinfectious virus
recov-ered from the plasma and spleen was maximal by 10 days
postinjection. In contrast, the infectious virus titer in both
the spinal cord and brain cells gradually increased and
reached a level which exceeded the maximal concentration
ofvirus foundintheplasma at 25 to 30dayspostinjection.At
this time, paralysis became evident in tsl-infected mice, whereas the wild-type virus-infected mice appeared to be
normal; the virus titer was about 2 orders of magnitude greater in tsl-infected spinalcord samplesand about 1 order of magnitude greaterintsl-infectedbrainsamplesthaninthe same tissues infected with the wild-type. The infectivity detected in the tsl-infected spinal cord and braincells could
not be due to the presence of endogenous viruses, since
these infectious viruses were found to possess the
pheno-type of the tsl virus, i.e., they were temperature sensitive andimmunoprecipitation ofthe intracellular protein ofthese
cells with anti-gp7O showed that Pr80env had accumulated.
These findings clearly indicate that tsl virus can infect and
replicate muchmore efficientlythanwild-typeMoMuLV-TB
in the central nervous system of infected mice. This
en-hanced neurotropism of tsl virions was further indicated
when it was found that tsl grew to high titers in primary
cultures of neurons from CFW/D mice, in contrast to the
barely detectable infectivityfound in similarprimaryneuron
cultures infected with wild-type virus. It may be speculated
that alterationsin theconformationofgp70in tslmay enable
tsl virions to gain entry into neurons. Alternatively, the
failure to process PM80env efficiently, which results in
re-duced amounts ofgp7O and piSE on the virion envelope of
on November 10, 2019 by guest
http://jvi.asm.org/
tsl (20), may allow tsl virions to bypass the blockage at the
cell surface and gain entrance to the neurons. Once tsl is
able to enter the neuron and establish replication, the
accumulation ofPr80env in the infected neuron may disrupt
its normal function, resulting in paralysis. In addition, tsl and wild-type virus may differ in their ability to integrate in the nerve cells, or a change(s) in the tsl genome may render
its replication in nerve cells much moreefficient than that of
wild-type virus. Experiments to resolve these possibilities are under way.
As yet no mammalian retrovirus protein has been shown
to possess the protease activity for processingPr80en'.Thus,
it has been assumed that Pr80env processing is carried out by
a cellular enzyme. The present studies have shown that a
mutation(s) in the nt sequence of tsl, extending from the
HindIll site (nt 4894) at the 3' end ofpol to the BamHI site
(nt 6537) of gp7O, retards the processing of Pr8Oen" at the
nonpermissive temperature. These findings suggest that the
HindIII-BamHI sequence of tsl may encode a heat-labile
env protein cleavage enzyme which functions inefficiently at the nonpermissive temperature or, alternatively, that the
sequence encoding gp7Ois altered so that the precursor has
an altered conformation which cannot be cleaved efficiently at the nonpermissive temperature.
As pointed out by Shinnick and co-workers (15), thepol
gene has a coding capacity much larger than necessary to
encode the 80,000-dalton reverse transcriptase molecule.
Recent studies bySchwartzberg and co-workers (13) showed
that mutants carrying deletions at theSacl(nt2558) andStuI
(nt 2927) sites released virus with no detectable reverse transcriptase activity, in contrast to mutants with deletions
at the HindlIl site (nt 4894), which produced virions with
normal reverse transcriptase and RNase H activities. These
findings demonstrated that the reverse transcriptase
mole-cule is encoded at the 5' end of the pol gene. Thus, the
sequence extending from somewhere between the StuI and
HindIIIsites to the beginning of the env sequence encodes a different function.
It has also been demonstrated by Kopchick and
co-work-ers (6) that Rauscher MuLV virions contain endo-DNase
activity in a 40,000-dalton protein, p40. Kopchick and
co-workers suggested thatp40 may be involved in integration of
the viral genome by making staggered cuts in the host and
viralDNAs. Recently, Donehower and Varmus (Proc. Natl.
Acad. Sci. U.S.A., in press) reported that the integration efficiency of the proviruses of two MoMuLV mutants, one with a base substitution at nt 4950 and the other with deletion mutations at nt 4950 and nt 4951, was reduced
10-fold. Thus, it appears that the 3' end of the pol gene
encodes the 40,000-dalton endonuclease. However, as
pointed out by Kopchick et al. (6), the reverse transcriptase
andendonuclease molecules together still do not accountfor
all the coding potential of thepol gene. The demonstration in
the present study that replacing the HindIII-BamHI
se-quence of tsl with the homologous wild-type sequence
corrected the defect in the processing ofPr80etn suggests the
possibility that the 3' end of the pol gene may in addition encode a protease responsible for cleaving the envelope precursor protein. The alternate explanation for our
obser-vationsis a conformational change in Pr80en' resulting from
a mutation(s) in the 5' end of the gp7O coding sequence
which renders it unable to be processed to gp7O and
p1SE.
To test these possibilities, we are currently making
con-structs by replacing the HindIII-XbaI (nt 4895 to nt 5766)
sequence of thepol gene or theXbaI-BamHI (nt 5767 to nt
6537) sequence of gp7O with the homologous sequence of tsl
andvice versa. Inaddition, werecently isolated a
rccombi-nant inCharon21A with adeletion mutationof 50 to 100
bp
at nt 5580. Characterization of these constructs and of the
deletion mutant should be helpful in resolving these
ques-tions.
ACKNOWLEDGMENTS
WewishtothankM. M.Soong forsomeof theproteinanalysis. We also thank the Word Processing Center of the University of Illinois College of Medicine at Urbana-Champaign fortyping this manuscript.
This investigation was supported by Public Health Service
re-search grantCA36293, awardedbytheNational Cancer Institute. LITERATURE CITED
1. Ball, J. K., T. Y. Huh, and J. A. McCarter. 1964. On the statistical distribution ofepidermalpapillomata in mice. Br. J. Cancer 18:120-123.
2. DesGroseillers, L., M.Barrette,and P.Jolicoeur.1984.Physical mappingoftheparalysis-inducing determinantofawildmouse
ecotropicneurotropic retrovirus.J. Virol.52:356-363. 3. Gardner, M. B. 1978.Type-C viruses of wild mice:
characteri-zationand naturalhistory ofamphotropic,ecotropicand
xeno-tropic murine leukemia viruses. Curr. Top. Microbiol. Im-munol.79:215-239.
4. Graham,F.L., and A. J.vander Eb.1973.Anewtechniquefor theassayof human adenovirus-5DNA. Virology52:456-461. 5. Kai, K., and T. Furuta. 1984. Isolation of paralysis-inducing
murine leukemia viruses from Friend viruspassage inrats. J. Virol.50:970-973.
6. Kopchick, J. J., J. Harless, B.S. Geisser, R. Killam, R. R. Hewitt, and R.B. Arlinghaus. 1981. Endodeoxyribonuclease activity associated with Rauscher murine leukemia virus. J. Virol. 37:274-283.
7. McCarter, J. A.,J. K. Ball, andJ.V. Frei. 1977. Lowerlimb paralysis induced in mice by temperature sensitive mutant of Moloney murine leukemia virus. J. Natl. Cancer Inst. 59: 179-183.
8. Miller, A., and I. Verma. 1984. Two base changes restore
infectivity to anoninfectious molecularclone ofMoloney
mu-rine leukemia virus(pMLV-1). J.Virol. 49:214-222.
9. Moloney,J.B. 1960. Biological studieson alymphoid-leukemia virus extracted from sarcoma 371. Origin and introductory investigation.J. Natl. Cancer Inst. 24:933-947.
10. Rao, R. N., andS. G. Rodgers. 1979. Plasmid pKC7: avector
containingtenrestriction endonuclease sites suitable forcloning DNA sequents. Gene7:79-82.
11. Rude, R., G. Gallick, and P. K. Y. Wong. 1980. A rapid screeningtechnique forthe isolation ofpostintegration temper-ature-sensitive mutants ofMoloney murine leukemia virus. J. Gen. Virol. 49:367-374.
12. Schwartzberg, P., J. Colicelli, and S. P. Goff. 1983. Deletion mutantsofMoloney murine leukemia viruswhich lack glycosyl-atedgagproteinarereplicationcompetent. J. Virol.46:538-546. 13. Schwartzberg, P., J. Colicelli, M. Gordon, and S. Goff. 1984. Construction andanalysisofdeletion mutations in thepolgene of Moloney murine leukemia virus: a new viral function re-quired forproductive infection. Cell37:1043-1052.
14. Shields, A., N. Rosenberg, and D. Baltimore. 1979. Virus pro-duction by Abelson murine leukemia virus-transformned
lym-phoidcells. J. Virol. 31:557-567.
15. Shinnick, T. M., R.A.Lerner,andJ. G. Sutcliff. 1981. Nucleo-tide sequence ofMoloneymurineleukemia virus. Nature (Lon-don) 293:543-548.
16. Southern, E. M. 1975. Detection ofspecific sequences among DNAfragments separatedby gel electrophoresis. J. Mol. Biol. 98:503-517.
17. Viera,G.,andJ.Messing. 1982. ThepUCplasmids,an
M13amp7-derived system for insertion mutagenesis and sequencingwith synthetic universal primers. Gene 19:259-268.
18. Wong, P. K. Y., and G. Gallick. 1978. Preliminary
on November 10, 2019 by guest
http://jvi.asm.org/
zation ofa temperature-sensitive mutant of murine leukemia viruswhich produces defectiveparticlesattherestrictive
tem-perature.J. Virol. 25:187-192.
19. Wong, P. K. Y., L. J. Russ, and J. A. McCarter. 1973. Rapid, selective procedure forisolation of spontaneous temperature-sensitive mutants of Moloney leukemia virus. Virology 51:424-431.
20. Wong, P. K. Y., M. M. Soong, R. MacLeod, G. Gallick, and P. H. Yuen. 1983. Agroupof temperature-sensitive mutantsof Moloneyleukemia virus which is defective in cleavage ofenv
precursor polypeptide in infected cells also induces hindlimb paralysisinnewborn CFW/Dmice. Virology 125:513-518.
21. Wong, P. K. Y., M. M. Soong, and P. H. Yuen. 1981. Replica-tion ofmurine leukemiavirus inheterologous cells: interaction between ecotropic and xenotropic viruses. Virology 109: 366-378.
22. Yuen,P. H.,D.Malehorn, C. Nau, M. M. Soong, and P. K. Y. Wong. 1985. Molecular cloning oftwo paralytogenic, tempera-ture-sensitive mutants, tsl and ts7, and theparental wild-type Moloney murine leukemia virus. J. Virol.53:178-185. 23. Yuen, P. H., M. M. Soong, M. S. Kissil, and P. K. Y. Wong.
1984.Restriction of Moloney murine leukemia virusreplication in Moloney murine sarcoma virus-infected cells. Virology
132:377-389.