JOURNAL OFVIROLOGY, Apr.1993, p.2336-2343 0022-538X/93/042336-08$02.00/0
Copyright
X) 1993, AmericanSociety
for MicrobiologyA
Cellular Cofactor
Facilitates Efficient 3CD
Cleavage
of the Poliovirus
P1 Precursor
WADE S.BLAIR, XIAOYULI, ANDBERT L. SEMLER* Departmentof Microbiology and Molecular Genetics, College of Medicine,
University
of California,
Irvine,
California
92717-4025Received6 November1992/Accepted 11January 1993
The production of poliovirus capsid proteins from a capsid protein precursor (P1) is mediated by virus-encoded proteinase 3CD and involves a complicated set ofproteinase-substrate interactions. In addition to
substrate and enzymatic determinants required for this interaction, we describe a cellular cofactor, which facilitates 3CDrecognitionof thePlprecursor.Cellular cofactoractivityis 3CDdependentand saltdependent. Ouranalysisshows thatproteolytic cleavageof the P1precursorattheVPO/VP3cleavagesite exhibitsagreater
dependencyonthecellular cofactor thancleavageattheVP3/VP1site. Suchagreaterdependencyoncellular cofactoractivity canbe relieved (in part) bythe substitution ofanAla residue for the Pro residue at the -4 positionof theVPO/VP3 cleavagesite.However,mutantvirusescontaining Pro-to-Alasubstitutions at the -4 position oftheVPO/VP3 site exhibit defects in viralgrowth.
Poliovirus, like all members of thefamily Picornaviridae, depends entirelyonprotein processingforthe productionof
mature gene products. In the infected cell, the poliovirus
genomic RNA (7.5 kb in length) is translated into a single
polyprotein (240kDa insize)whichis cleaved into functional viral polypeptides atY. G and Q. G amino acid pairs by virus-encoded proteinases 2A and 3C, respectively (8, 10, 18, 25, 26). Viral proteinase 3C exhibits highly specific activitywhichrecognizes few cellularproteins (15, 27) and cleaves the viral polyprotein onlyat a subset of the Q G aminoacidpairstoyieldauthentic viralgeneproducts.Such 3Cproteolytic fidelity requiresacomplexsetofinteractions, whichdependon (i) recognition ofQ. Gcleavage sites, (ii)
otherprimaryamino acid sequence determinants surround-ing Q. G sites,and(iii)structural determinants surrounding Q. Gcleavagesites.
Thecomplexityofpoliovirus proteinase-substrate interac-tions ismostapparentforcleavageofthe P1(capsid protein)
precursor (Fig. 1), which requires polymerase sequences
(3D) in addition to 3C proteinase sequences in the form of proteinase 3CD (13, 29). 3CD recognition of P1 dependson a complete and intact substrate. Carboxy-terminal trunca-tion (31, 32), incomplete processing ofP2sequences from the P1 carboxy terminus (22), or disruption of P1 ,B-sheet
secondary structures by amino acid insertion mutagenesis (30) results in the disruption of P1 processing in vitro. Interestingly, disruption of ,3-sheet secondary structuresin
any onecapsid protein presentin the P1 precursordisrupts
cleavage of the entire P1 molecule. These observations suggest that P1 tertiary structure may be important for
correctpresentation of the Q. G cleavage sitestoproteinase 3CD or, alternatively, that P1 tertiary structure may be
specifically recognized by proteinase 3CD. In addition to
structural determinants, primary sequence determinants
other than the Q. G cleavage sites have been defined for proteinase 3CD recognition of the P1 precursor. Efficient
cleavage of the VP3/VP1 cleavage site depends on the
presence of an Ala residue in the -4 position (4 residues
proximal to thecleaved Q - G scissilebond) (3). The latter
*Correspondingauthor.
primarysequencedeterminant alsoseemstoberequiredfor efficient cleavage of P2 and P3 precursor polypeptides. Synthetic peptides containing authentic poliovirus (24) or
rhinovirus (6) P2 or P3 cleavage sites are cleaved more
efficiently in the presence of homologous purified 3C
pro-teinases whenanAlaresidue is presentatthe -4positionof theQ Gcleavagesites.
Despite a preliminary understanding of the nature of 3CD-P1 interactions, fundamental questions, such as the role ofpolymerasesequences(3D)in3CDrecognitionofP1 and the composition of the 3CD-P1 proteolytic complex, remainlargelyunanswered. Inan attemptto further under-stand 3CD-P1 recognition,weexaminedanadditional
deter-minant required for 3CD-mediated P1 processing. In this study,wedemonstrate that efficient 3CD-mediatedcleavage of the P1 precursor requires a cellular cofactor. We also showthatproteolytic cleavageattheVP0/VP3 cleavagesite exhibits a greater dependencyon cellular cofactor activity
than cleavage at the VP3/VP1 site. Such an increased
dependencyoncellular cofactoractivitymayresult from the lack of an Ala residue at the -4 position of the VPO/VP3
cleavage site. We also demonstrate that the single amino acid substitution ofanAla residue for theProresidueatthe -4 position of the VPO/VP3 cleavage site results in an
increasedefficiencyofproteolytic cleavageatthat site under conditions limiting for the cellularcofactor in vitro.
How-ever, such a single amino acid substitution haspleiotropic
effects. We showthatmutantvirus, which contains a
Pro-to-Ala substitution at the -4 position of the VP0/VP3 cleavage site, exhibitsdefects in virus growth.
MATERIALS ANDMETHODS
Construction ofthesingleaminoacid substitutionmutation pT7P1-1B(P4A). Synthetic oligonucleotides 20 residues in lengthwereused inthe heteroduplex methodof site-directed
mutagenesis describedby Inouye and Inouye (12) to intro-duce a single amino acid substitution in poliovirus
subge-nomic cDNA pT7-P1 (30) at poliovirus nucleotide 1754 (C-to-Gtransversionmutation), resultinginplasmid pT7P1-1B(P4A). Plasmid pT7P1-lB(P4A) was digested with
endo-nucleasePflMI,anda3,129-bp fragment containing
poliovi-2336
Vol.67, No. 4
on November 9, 2019 by guest
http://jvi.asm.org/
POLIOVIRUS P1 CLEAVAGE COFACTOR 2337
P1 I P2
I
VPO I VP3I
VPI IN@ -,*
97
kD-63 kD
-60 kD
-37 kD
-26 kD-34kD
-I1CD
|1
VPO
I
I
IEvVP3_]
lJFIG. 1. Diagram of potential cleavage products resulting from 3CD-mediated cleavage of P1 precursors in vitro. P1 precursors
synthesized in vitro from subgenomic poliovirus cDNApT7-P1and incubated in thepresenceof proteinase 3CDarecleaved into capsid
protein VPO, VP3, and VP1 orpolypeptide intermediate 1ABC (a
VPO-VP3 fusion) or 1CD (a VP3-VP1 fusion). Molecular sizes of
capsid proteins and polypeptide precursors areshownonthe left.
(+), 3CD-mediatedcleavageevents. NCR, noncodingregion.
rus sequences 496 to 3625 was introduced into a T7
transcription vector containing a full-length poliovirus
cDNA(pT7-PV1) (9), resulting in plasmid pT71-1B(P4A). In vitro transcription of subgenomic and full-length polio-virus cDNAs.SubgenomiccDNAswerelinearized by
diges-tionwithHindIII, while full-length cDNAs were linearized
by digestion with EcoRI at sites located 3' of poliovirus
sequences. In vitro transcriptions of subgenomic and full-lengthcDNAswerecarriedoutwithbacteriophage T7 RNA polymerase asdescribed previously (19, 30).
In vitro translation of RNAs derived from subgenomic cDNAs and cleavage of in vitro-synthesized P1 precursors.
Translation of RNAs derived from subgenomic cDNAs pT7-P1 and pT7P1-1B(P4A) was performed as described
previously (3, 30). Either 1 or 2.5
RI
of the resulting P1 invitro translation reaction mixture was incubated in the presence of either 1
p,l
of PV1-infected HeLa cell extractdiluted 10-fold in HD buffer (20 mM N-2-hydroxyethylpi-perazine-N'-2-ethanesulfonic acid [pH7.4]-1 mM dithiothre-itol) or 1.5,ul ofpoliovirustype 1 (PV1)-infected HeLacell
extract, respectively.The above cleavage reactionmixtures
weredilutedtoafinalreactionvolumeof5,10, 25,50,or100
,ulin thepresenceof HDbufferandincubatedfor 3 hat30°C, orwhen indicated, cleavage reaction mixtureswere diluted
100-fold inHDbuffercontaining 150 mMpotassium acetate
(KOAc), HeLaS10cellextract(totalprotein, 400,ug), HeLa S200 cell extract (total protein, 400
p,g),
or fractionatedHeLa S200 cell extracts (total protein, 50
p,g)
prior toincubation. Cleavagereactionswereterminated by addition
of equal volumesof 2x Laemmlisamplebuffer, andcleavage
productswere analyzed by sodium dodecyl sulfate
(SDS)-polyacrylamide gelelectrophoresis (17). Autoradiogramsof SDS-polyacrylamide gels were analyzed with an LKB-2
Ultrascanlaserdensitometer.
Fractionation ofHeLa S200 cell extracts. HeLa S200 cell
extracts were prepared bycentrifugation of HeLaS10 cell
extracts, preparedasdescribedpreviously(4, 7), in a
Beck-man 50 Ti rotor at 200,000 x g for 1.5 h. HeLa S200 cell
extracts were dialyzed in buffer C (Tris-hydrochloride
[pH
7.9]-1 mMdithiothreitol) and loadedon aphosphocellulose
column.Afterelutionwith bufferC, cellular cofactor
activity
was detected by using the above-described cleavage assay with theflowthroughfraction, whichwassubsequently sub-jected toDEAE-cellulose(DE52)chromatography. Cellular cofactor activity was then eluted from the DEAE-cellulose columnwith 0.4 MNaCl in buffer C. All fractionated HeLa S200extracts weredialyzed inHDbufferpriortoanalysis.Transfection of RNAs derived from full-length poliovirus
cDNA pT71-1B(P4A) and mutant virus stock preparation.
RNAs derived from pT71-1B(P4A) were used to transfect subconfluent HeLa cellmonolayersasdescribed
previously
(3, 28). Transfected HeLa cell monolayers were overlaidwitheitherDulbeccomodified Eagle mediumcontaining 10%
fetal calf serum or a semisolid medium consisting of
Dul-becco modified Eagle medium, 6% fetal calf serum, and 0.45% agarose. The transfectedmonolayerswereincubated at 33°C until cytopathic effects or plaques were visible. Mutant virus stocks were prepared by either picking well-isolated plaques 3days aftertransfectionor
harvesting
liquid
overlays 2days after transfection. Liquidoverlay
harvests were usedtoinfectfresh HeLa cellmonolayers,whichwere thenoverlaidwith semisolid medium.Well-isolatedplaques
were then picked as described above and wereclonally
purified by asecond round ofinfection andplaque isolation.The plaque-purified stocks were expanded
by
two serial passagesthrough HeLa cell monolayers at33°C. The titers ofthestocks were determinedon60-mmplatesof HeLa cell monolayers under semisolid mediumat33°C.Sequencing of mutant viral RNA. Viral RNA was har-vested by Nonidet P-40 lysis as described by Campos and Villarreal (5). RNA was sequenced by extension of a 20-nucleotideprimercorrespondingtonucleotides1810
through
1830ofpoliovirus RNA by using[a-32P]dATP,
dideoxynu-cleotides, and reverse transcriptase.[35SJmethionine
pulse-labeling of infected HeLa cells.[35S]methionine pulse-labeling of HeLa cells infected with either PV1 or Sel-lB-06 wascarriedout as described
previ-ously(1). Briefly, HeLacellswere infectedat amultiplicity
ofinfection of 30 and incubatedat33°Cfor 6 h. After the 6-h incubation, infected cells were labeled with 60pCi
of[35S]methionine,
incubated for 1 h at either 33 or39°C,
and then harvested. Pulse-labeled viral proteinswere diluted in Laemmli sample buffer and analyzed on SDS-containing12.5% polyacrylamide gels.
One-stepgrowth curve
analysis.
HeLa cellmonolayers
on60-mmplateswereinfected witheither PV1 orSel-lB-06 at a multiplicity ofinfection of 30. After a 30-min
adsorption
period, infected monolayers were rinsed with phosphate-buffered saline and overlaid withliquid
medium. The in-fectedmonolayerswere incubated at33or39°C.
After2,
3,
4, 5, and 6 hat39°Cor2, 4,6, 8,
and 10 h at33°C,
cells and supematants were harvested. Plaque assays were used to determine PFU percellat eachtimepoint.
RESULTS
Proteolytic processing ofin vitro-synthesized P1 precursor polypeptides under diluted cleavage reaction conditions. To examine efficiencies of
cleavage
attheVP0/VP3
siterelative totheVP3/VP1 site, P1 cleavage assayswerecarriedout in vitro under various dilution conditions. The rationale for these experiments was that under dilutecleavage
reaction conditions, the kinetics ofvirus-specific
cleavage
of P1(a
trans-cleavageevent)would be
effectively
reduced,
allowing
a careful analysis of relative
cleavage
efficiencies at theVOL. 67,1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.59.296.74.244.2]2338 BLAIR ET AL.
M 1 1//1/5V10 125 /50 1/100
1CD-
vPo-
VPl-
VP3P-1 2 3 4 6 7
FIG. 2. 3CDcleavageof in vitro-translated P1precursorsunder
diluted reaction conditions. As describedinMaterialsandMethods, RNAs transcribed from awild-type poliovirus
subgenomic
cDNA (pT7-P1)weretranslated in vitro in thepresenceof[ 5S]methionine. In vitro P1 translation reaction mixtureswerediluted5-, 10-, 25-, 50-,or100-fold(asindicatedatthetop)in HD buffer and incubated in the presence of PV1-infected cell extract for 3 h at 30°C. Following incubation, cleavagereaction mixtureswerediluted in 2xLaemmlisample buffer,and theresulting productswereanalyzed by
SDS-polyacrylamide gel electrophoresis. M,marker lane ofextracts
from[35S]methionine-labeled, PV1-infected cells.
VPO/VP3 andVP3/VP1sites. As describedin Materials and Methods,P1precursorsweresynthesizedinvitrofrom RNA transcriptsderived from awild-type poliovirus subgenomic
cDNA (pT7-P1) in the presence of [35S]methionine and incubated in thepresenceof PV1-infected HeLa cellextract
(a source ofproteinases 3C and 3CD) at various cleavage
reaction mixture dilutions in HD buffer. As shown inFig. 2, virus-specific cleavageatthe VPO/VP3cleavagesiteappears tooccurlessefficientlyrelativetocleavageat theVP3/VP1 site under dilutecleavagereaction conditions. With increas-ing cleavagereaction mixture dilutions (5-, 10-,and25-fold) (Fig. 2,lanes 3to5),acorrespondingdecrease inproteolytic
cleavage efficiency at the VPO/VP3 site relative to the VP3/VP1sitewasobserved. This is shownbyan accumula-tionofintermediate 1ABC(aVPO-VP3 fusionproduct
[Fig.
1]) and a decreasedproductionofcapsid proteinsVPO and
VP3relativetoVP1. Atcleavagereactionmixture dilutions of 10- or 25-fold virus-specific cleavage occurred almost
exclusively at the VP3/VP1 cleavage site, and at cleavage reaction mixture dilutions of 50- or 100-foldvery little P1
cleavagewasobserved(Fig. 2). The decreased efficiency of cleavage at the VPO/VP3 site relative to the efficiency of cleavage at the VP3/VP1 site is more pronounced than
appears in Fig. 2because ofunequal capsid protein methi-onine content. The methionine contents ofcapsid proteins VPOandVP3are1.8- and2.3-foldgreater,respectively, than that ofcapsid proteinVP1.Therefore, capsid proteins VPO
and VP3 appear to be present in greater molar quantities
thanVP1 when labeled with [35S]methionineand analyzed
on
SDS-polyacrylamide gels.
Laser densitometer analysisand correction forcapsid proteinmethioninecontentshowed thatcleavageofP1is three-tofivefoldmoreefficientatthe
VP3/VP1site thanat the
VPO/VP3
site in cleavagereaction mixturesdiluted 10-fold(datanot shown).These datadem-onstrated that
virus-specific proteinase
interaction is lessefficientat the
VPO/VP3
site thanattheVP3/VP1site underdilute
cleavage
reaction conditions. Such differential cleav-age efficiencies may result from an increasedstability
ofvirus-specific
proteinase
interaction attheVP3/VP1
cleav-age site relative to theVPO/VP3 site, possibly
due to the presenceofanAlaresidueatthe -4position
of theVP3/VP1
site(see below).
Identification ofacellularcofactor
required
for 3CD-medi-atedcleavage ofP1. The datapresented
above are reminis-centofaloss in3CD-specific activitywithincreasing cleav-age reaction mixture dilution.3CD-specific activity
can bedefinedbyefficient
cleavage
at theVPO/VP3
andVP3/VP1 cleavage sites asopposed
to3C-specific activity,
whichmediates inefficient
cleavage
at theVP3/VP1
site(21).
Todetermine whether such a loss in
3CD-specific activity
resulted from thedilution ofsomefactorendogenous
tothe cleavagereaction other than enzymeorsubstrate, cleavage
reactionmixtureswerediluted100-fold andincubated under various conditions. At 100-fold
cleavage
reaction dilutionconditions,
thebackground
levels of 3Cactivity (detectable
with lower dilutions ofcleavage
reactionmixtures)
are reducedsufficiently
such thatonly
3CD-specific
activity
(relatively
efficientcleavage
at both theVPO/VP3
and theVP3/VP1
sites)
is detected underoptimal
conditions. Inaddition, in P1 cleavagereaction mixtures diluted
100-fold,
the contributionsofputative
cofactors present inthesources of enzyme andsubstrate areminimized(thus allowing
fora more sensitivecleavageassay).
As showninFig.
2(lane 7)
andFig.
3A(lane 4),
invitro-synthesized
P1 precursors diluted100-foldin HD bufferorHDbuffercontaining
KOAc(at
afinal concentration of 150mM) (Fig. 3A,
lane6)
and incubated inthepresenceofPV1-infectedcellextract werecleaved very
inefficiently
at either theVPO/VP3
or theVP3/VP1
siteby proteinase
3CD.However,
whencleavage
reaction mixtureswere diluted 100-fold in HD buffercon-taining
150 mM KOAc and HeLaS10extract(400
,ugof total protein per 100 ,ul of reactionmixture)
P1 precursorswere cleavedefficiently
atboth theVPO/VP3
and theVP3/VP1
cleavagesites(Fig.
3A,lane9).
These data demonstratethat acellular cofactor presentin HeLaS10cytoplasmic
extractsfacilitates efficient
3CD-specific activity
onP1 precursors. Apreliminary
characterization of the cellular cofactor revealed thatitsactivityisdependentonthe presence of salt(150
mMKOAc)
and the presence ofproteinase 3CD. As showninFig.
3A, invitro-synthesized
P1 precursorsdiluted100-fold in HD buffer containing HeLa S10 extract (total protein,400
,ug)
andincubated in the presence ofanextract ofPV1-infected cellswerecleaved veryinefficiently by 3CDinthe absence of salt
(150
mMKOAc) (lane 7).Similarly,
P1precursorsdiluted 100-fold in HD buffercontaining150 mM KOAc and HeLa S10 cell extract and incubated in the absence of PV1-infected cellextract (thesourceof
protein-ase
3CD)
remaineduncleaved(Fig. 3A,lane8). Thecellular cofactor, therefore, does not appear to exhibit proteolyticactivity.
Thecellular cofactor is also heat labile andphenol soluble. HeLa S10 cellextract that was incubated at 600C(Fig.
3A,lane10)
orphenol-chloroformextracted(Fig. 3A,lane
11) prior
to analysis exhibited no cofactor activity. Cellularcofactoractivity
was retained in the S200 fractionfollowing centrifugation
of a HeLa cell cytosolic extract. J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.112.254.78.314.2]POLIOVIRUS Pl CLEAVAGE COFACTOR 2339
A
zC:
0 + M z
Kl IN
U o o
+ - +
0 Co
CO
C
_ o
co 0 Co) co
P1-
1ABC-
VPO-VP1-- _
V P3- &
B XL DE52
M K CiW U- U- o o o
P1=5
1ABC- di
1CD-X v
vPO-VP1-.
VP3- _
1 2 3 4 5 6 7 8 9
UV~~~..
1 2 34568~~~~:7
9 10 11
Fractionation of HeLaS200extractsbyionexchange chro-matography showed that the P1 cleavage cofactor activity
waseluted in theflowthrough fraction ofaphosphocellulose
column (Fig. 3B, lane 4) and was eluted with 0.4 M NaCl
from a DEAE-cellulose column (Fig. 3B, lane 7). Such a
preliminary characterizationsuggeststhatthe cellular cofac-torisaspecific proteinorprotein complex ofayetunknown molecularweight.
Fractionationof the HeLa S200 cellextract also revealed cellular proteases with enriched specific activities, which cleaveP1toyield nonauthentic viral polypeptides of
appar-entmolecularweightssimilartothoseofviral proteinsVPO and VP3 (Fig. 3B, lanes 8 and 9). The activities of such cellularproteases are distinct from the 3CD-dependent P1 cleavage cofactor (discussed above) and seem to be 3CD independent (data notshown). Cellular protease activity is notdetectedwith HeLaS10orS200 cellextractpresumably becauseofverylowspecificactivities in theseextractsprior to fractionation. It is unlikely thatthese cellular proteases haveroles in P1processing.
Proteolyticprocessingof P1precursorscontaininga
Pro-to-Ala substitution atthe -4 position of the VPO/VP3 cleavage site. Todetermine whetherdifferential 3C-or3CD-mediated
cleavage efficiencies at the VPO/VP3 and VP3/VP1 sites under conditions limiting for the above-described cellular cofactorcanbe attributedtothepresenceorabsence ofa-4
Alaresidue,aPro-to-Ala singleamino acidsubstitutionwas
introduced atthe -4position oftheVPO/VP3cleavage site [pT71-1B(P4A)] (Fig. 4).P1precursorsderived from pT7P1-1B(P4A) or pT7-Pl (wild type) were synthesized in the
presenceof[ 5S]methionine andincubated in thepresenceof
FIG. 3. In vitro 3CD-Pl cleavage reaction mixtures incubated undervarious dilutionconditions. Pl precursors were synthesized in vitroin the presenceof[35S]methioninefrom wild-type poliovirus in vitro-synthesized mRNAs. (A) In vitro Pl translation reaction mixtureswerediluted100-fold inHDbuffer(lanes 4 and 5) ordiluted 100-foldin HD buffercontaining150mMKOAc(lane 6), HeLaS10 extract(totalprotein,400pLg)(lane 7),150mM KOAcand HeLaS10 (total protein, 400 p.g) (lanes 8 and 9), 150 mM KOAc and heat-treated(60°C)HeLaS10extract(total protein, 400ptg)(lane10),or 150mMKOAc andphenol-chloroform-extracted HeLaS10extract (lane 11). Diluted Pl precursors were incubated either in the presence(lanes4, 6, 7,and 9 to11)orin the absence ofPV1-infected (INF) cellextract(lanes5 and 8). Lane 3, in vitro-synthesized Pl precursors incubated in the presence of PV1-infected HeLa cell
extractunder undiluted(U) cleavage reaction conditions.No RNA, in vitro translation reaction mixtures incubated in the absence of RNA.(B)Invitro-translated Pl precursors were diluted100-foldin HDbuffer containing150mMKOAc(lane 2), HD buffer containing 150 mM KOAc and HeLa S200 cell extract (total protein, 400 ,ug) (lane3), orHD buffercontaining150mM KOAc and fractionated HeLaS200 cellextracts(total protein,50,ug) (lanes4 to9). Lane 4, flowthrough (FT)fraction afterphosphocellulose chromatography; lane5, theFTfraction after DEAE-cellulose chromatography;lanes 6to 9,fractions eluted with 0.2, 0.4, 0.6, and1 MNaCl, respec-tively,fromaDEAE-cellulosecolumn. DilutedPl precursors were incubated in thepresenceofanextractfromPVl-infected cells for 3 hat30°C. M,marker lane ofan extractof[35S]methionine-labeled, PV1-infectedcells.
PV1-infected cell extractundervarious dilution conditions. As shown in
Fig.
5, proteolytic cleavage of P1 precursors derived frompT7P1-lB(P4A) atthe VPO/VP3 cleavage site was more efficient relative tocleavageat theVP3/VP1 site (lanes 9 to 12) under conditions limiting for the cellular cofactor thancleavageattheVPO/VP3 site of P1 precursors derived frompT7-P1 (lanes4 to7).This ismostapparentforcleavage reaction mixtures diluted 10- or 25-fold. Laser densitometer analysis showed that P1 precursors derived from pT7P1-lB(P4A) are cleaved three- to fourfold more
efficiently at the
VPO/VP3
site than P1 precursors derived from the wild type in cleavage reaction mixtures diluted VOL. 67, 1993on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.349.529.76.330.2]2340 BLAIR ET AL.
P1
| VPO | VP3 | VP
WT:
< I.L
z Z
0 0
M z z
P1-ij[-.
1ABC-
ICD-* t
. *. Leu Pro Arg Leu Gln Gly LeuPro*.* 268269 270 271 272 1 2 3
1 1/5 1/10 1/25
LL
z
0
z
1B( P4A)
1 1/5 910 /25
..
..
,1g....
.';-'i'
ts
.. _ :
st
_ iilii Xaxief iw.> A,,
-, 7';'.
.j4,,,,..Qts:.
*'':: ;'JF_*
SO;"
pT7P1-1B(P4A,
. . . LeuAla Arg Leu Gln GlyLeu Pro * *
[image:5.612.63.300.73.230.2]288269270 271 272 1 2 3
FIG. 4. Diagram of theP1 precursorillustratingthesingleamino acid substitution mutationpT7-lB(P4A).Aminoacid residues span-ning theVPO/VP3cleavage site forwild-type (WT) P1 precursors andP1precursorsderived frompT7Pl-lB(P4A)aredepicted.*, -4 residue;V,cleaved
Q0
G amino acidpair.10-fold (data not shown). Additionally, incubation of P1
precursors derived from pT7P1-1B(P4A) diluted 25-fold in
HDbuffer in the presenceofanextractfrom PV1-infected cellsresulted inproteolytic cleavageattheVP0/VP3site and theproduction ofcapsid proteinsVPOandVP3(Fig. 5,lane 12), while very little cleavage at the VPO/VP3 site was
observed when P1 precursors derived from the wild type
were incubated under the same conditions (Fig. 5, lane7). These data demonstrate thatanAla residue introducedatthe -4 position of the VPO/VP3 site increases 3C or 3CD cleavage efficiency atthat siterelative totheVP3/VP1 site under conditionslimitingfor the cellular cofactor.Theyalso suggestthat the cellular cofactorserves acompensatory role tofacilitate efficient 3CDcleavageatthewild-type VPO/VP3 cleavage site in the absence ofa -4Alaresidue.
[35Sjmethionine pulse-labelingofcells infected withmutant virus Sel-lB-06 or PV1. The Pro-to-Ala single amino acid
substitution mutation at the -4 position of the VP0/VP3 cleavage site was introduced into a full-length poliovirus
cDNAto construct plasmid pT71-B(P4A). After transfec-tion ofRNAs derived frompT71-1B(P4A)into cell
monolay-ers,mutantvirusSel-lB-06, which contained the Pro-to-Ala amino acidsubstitution, was recovered. Mutant virus
Sel-1B-06 displayed a small plaque phenotype at both 33 and 39°C. [35S]methionine pulse-labeling analysis was used to examinetheproduction ofvirus-specific proteins in
Sel-1B-06-infected cells at 33 and 39°C. Cells were infected with either Sel-lB-06 or PV1 for 6 h at 33°C, labeled with [35S]methionine, and incubated ateither33or39°C for 1 h.
After the 1-h incubation, cells were harvested, and
virus-specific polypeptideswereanalyzedonSDS-polyacrylamide
gels. [3S]methionine pulse-labeling analysis showed that protein processing in cells infected with mutant virus Sel-1B-06at33°Corin cells infected with Sel-lB-06at33°C with
ashiftto39°Cimmediately following the pulse with
[35Slme-thioninewasphenotypically similartothat for the wildtype
(Fig. 6). However, a slightly decreased accumulation or
production of 1ABC (a VPO/VP3 fusion product) in cells infectedwithSel-lB-06 compared with that forthewildtype
was observed. Such a slightly decreased accumulation of 1ABCwasconfirmed byimmunoprecipitationanalysis using
antiserum directed against capsid protein VP3 (data not
shown) and presumably results from anincreased cleavage
efficiency attheVPO/VP3 site. These data, therefore,
dem-1 2 3 4 5 6 7 8 9 10 11 12
FIG. 5. 3CDcleavageofPlprecursorsderived frompT7-Plor
pl7Pl-1B(P4A). Pl precursors were synthesized in vitro in the
presenceof[35S]methioninefrom mRNAs derived fromsubgenomic pT7-Pl (wildtype[W+])orpT7Pl-lB(P4A)cDNAasdescribedin
Materials and Methods. PlprecursorsderivedfrompT7-Pl (lanes4
to7)orpT7Pl-lB(P4A) (lanes9to12)werediluted5-,10-,or25-fold (asindicatedatthetop)in HDbuffer and incubated in thepresence of PV1-infected cellextract.Lanes 3 and8, Plprecursorsderived
frompT7-PlorpT7P1-1B(P4A)wereincubated in the absence(no
INF) of an extract from PV1-infected cells. No RNA, in vitro
translation reaction mixtures incubatedinthe absence of RNA.M, marker lane of an extract from [35S]methionine-labeled,
PV1-in-fected cells.
onstratethataPro-to-Alaaminoacid substitutionatthe -4 position of the VP0/VP3 site results ina slightly increased
cleavage efficiency atthat site andareconsistent withwhat
wasobserved invitro(Fig. 5).
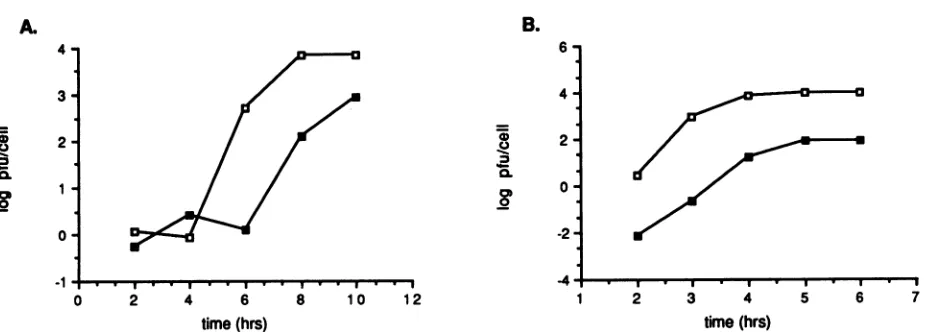
One-step growth analysis of mutant virus Sel-1B-06. To determinethe effects of the-4Pro-to-Alasubstitutionatthe VPO/VP3 siteonviralgrowth,one-stepgrowthanalysiswas
carried out at 33 and 39°C as described in Materials and
Methods. The results of suchananalysis showed thatmutant virus Sel-lB-06 exhibited delayed kinetics of virus produc-tioncomparedwith thatof thewild typeat33°C (Fig. 7A).In addition, 10-fold and 100-fold reductions in maximum viral yield from cells infected withmutantvirus Sel-lB-06
com-pared with the yield for the wildtypewereobservedat33°C (Fig. 7A) and 39°C (Fig. 7B), respectively. These data, therefore, demonstrate thata Pro-to-Ala single amino acid substitutionatthe -4positionof theVPO/VP3 cleavagesite results in a temperature-sensitive defect in viral growth,
mostlikelyatsomestepof the viral replication cycle other thanproteolytic processing.It ispossiblethat suchanamino
acid substitution interferes with capsid protein function at thelevel ofprotomer-protomer interactions.
vP0-~
tw
VP - 1 _
.~
VP3-_NOW
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.320.556.75.380.2]POLIOVIRUS P1 CLEAVAGE COFACTOR 2341
(0.
0
9
(0
0
m
P1 - _ _ g*
P3---3CD
1ABC
-3D- _...
VPO-VP2
-VP3-
_ _
2A-- _
[image:6.612.127.226.77.435.2]1 2 3 4
FIG. 6. [35S]methionine pulse-labeling analysisof cells infected
with PV1 or Sel-1B-06. As described in Materials and Methods,
cellswere infected with either PV1 (lanes 1 and 3) or Sel-1B-06
(lanes 2and 4), incubatedat33°C for6h, pulsedwith
[35S]methio-nine, andincubatedateither33°C (lanes 1 and 2)or39°C(lanes 3 and
4). Cellswereharvested andresuspended in Laemmli sample buffer,
and viralpolypeptides were analyzed by SDS-polyacrylamide gel
electrophoresis.
It is unlikely that the observed temperature-sensitive defectinSel-1B-06 virus growth results from lesions in the virusgenomethatmayhave beenintroduced during cDNA
manipulations or virus stock preparation other than the
primary mutation. The Pro-to-Ala mutation was
recon-structed in different full-length poliovirus plasmid back-grounds, and a mutant virus which exhibited a growth
phenotype similar to that of Sel-1B-06 was generated. In
addition, multiple plaqueswere isolated during theoriginal
construction ofmutantvirusSel-1B-06, and all virus stocks derived from such plaque isolates exhibited similar growth phenotypes.
DISCUSSION
Proteinase 3CDrecognition and cleavage of the P1
precur-sor is a complex reaction, involving proteinase-substrate
interactions at multiple levels. Structural determinants as
well as definedprimary amino acid sequence determinants
arerequired forthisinteraction. Inthis article, the
require-ment for a cellular cofactor, an additional determinant for efficient 3CD cleavage ofP1, was described. Such a cofactor appears to be a specific protein or protein complex which was eluted in theflowthroughof aphosphocellulose column and was eluted with 0.4 M NaCl from a DEAE-cellulose column. Cellular cofactor activity is 3CD dependent and dependent on salt (150 mM KOAc). Proteolytic cleavage of the P1 precursor at the
VPO/VP3
site exhibited a greater dependency on cellular cofactor activity than cleavage at theVP3/VP1 cleavage site. We believe that such a greater dependency results from the absence of a -4 Ala residue (which is present at the VP3/VP1 site) at the
VPO/VP3
site. Consistent with this conclusion, the substitution of an Ala for the Pro residue at the -4 position of theVPO/VP3
site resulted in an increased cleavage efficiency at that site under conditions limiting for the cellular cofactor. However, mu-tant virusSel-1B-06
containing such a Pro-to-Ala substitu-tion at the -4position of theVP0/VP3 site exhibited defects inviral growth, resulting presumably from defects in viral capsid proteinfunction rather than defects inP1 processing. The cellular cofactor described in this study seems to increase3CD-specific activity. 3CD-specific activity is func-tionally differentiated from 3C activity on the basis of the efficiency ofP1cleavage. Proteinase 3CD activity mediates efficient cleavage at both the VPO/VP3 and the VP3/VP1cleavage sites, while proteinase 3C mediates inefficient cleavage only at the VP3/VP1 cleavage site (16, 21). It is probable that in P1cleavage reaction mixtures diluted less than 25-fold (Fig. 2) a sufficient amount of 3C-specific activity is present to mediate inefficient but detectable amounts of VP3/VP1 cleavage. The observed preferential proteolytic cleavage at the VP3/VP1 site relative to cleavage at theVPO/VP3siteunder suchcleavage reaction conditions mayresult from areduction of 3CD-specific cleavage activ-ity to levels near or below background levels of 3C activactiv-ity. However, inP1cleavage reaction mixtures diluted 100-fold (a more sensitive assay system)inefficient 3C-specific cleav-age activity on P1 precursors is diluted to nondetectable levels and the assay detects exclusively 3CD-specific P1 cleavage activity. Under such assay conditions, which allow thedistinction of 3C- versus 3CD-specific activity, very little 3CD-mediated P1 cleavage was detected in the absence of cellular cofactor activity (Fig. 3). Our data, therefore, dem-onstrate that 3CD-specific activity is greatly facilitated by the presence of cellular cofactor activity. Therequirement of acellular cofactor for efficient 3CD-mediated cleavage ofP1 may explain the results of a recent study describing the purification of bacterially expressed 3CD proteinase. Pro-teinase 3CD, partially purified from genetically engineered
Escherichia
coli, exhibited lower than expected levels of 3CD- versus 3C-specific cleavage activity on P1 substrates (11), possibly resulting from the absence of3CD-P1cleavage cofactor activity in bacterial extracts (3a).As a function of facilitating 3CD-specific activity, the cellular cofactor appears to mediate recognition of a nonideal(VPO/VP3)cleavage site. Asdiscussed above, a -4 Alaresidue is present at most of the cleaved
Q.
Gpairs on thepoliovirus polyprotein (23) and seems to be animportant substrate determinant required for 3C- or 3CD-mediatedsubstraterecognition (3, 24). On the basis of these criteria, theVP0/VP3cleavage site, whichcontains a -4 Pro residue, might bedefined as a less than ideal(nonideal) cleavage site. Despite the absence of a -4 Ala residue, 3CD-mediated
cleavageof theVP0/VP3 siteappearstobeefficient in vivo (2) and in vitro under nondiluted cleavage reaction
condi-tions (Fig. 2). However, under dilute cleavage reaction
VOL. 67,1993
on November 9, 2019 by guest
http://jvi.asm.org/
2342 BLAIR ET AL.
B.
6-
4-CD
2-0.
0--2
I . . I . . I . . I . I '
.--2 4 6 8 10 12
time(hrs)
I , . I , . .I I
1 2 3 4 5 6 7
time(hrs)
FIG. 7. One-stepgrowth analysis of cells infected with either PV1 orSel-1B-06. Cellswereinfected with either PV1 (E) ormutant
Sel-1B-06(U)at33'C(A)or39'C (B), andcells and supernatantswereharvestedatspecifiedtimes after infection.PFUpercellateach time pointwere determinedbyusing plaque assays.
conditions (10- or 25-fold) proteolytic cleavage at theVPO/
VP3 site appears to be less efficient than cleavage at the VP3/VP1 cleavage site, which contains a -4 Ala residue.
Cleavage efficiencyatthe nonidealVPO/VP3 cleavagesite is increasedindiluted cleavage reaction mixtures by either the additionofcellular cofactoractivity (Fig. 3)orthe substitu-tion ofanAla residueforthe Pro residue atthe -4position of the VPO/VP3 cleavagesite(Fig. 5). Sucha-4Pro-to-Ala
substitution also results ina slightly increasedefficiency of
cleavageatthe VPO/VP3 site in vivo(Fig. 6). Wepropose,
therefore, that in addition toincreasingoverall 3CD
recog-nition ofP1,the cellularcofactor alsoservestocompensate for the lack ofa -4Alaresidueby facilitatingefficient 3CD interactionspecificallyat theVPO/VP3 site.
Although thepresence ofa -4 Pro residuemay
compro-mise cleavage efficiency at the VPO/VP3 site under condi-tions limiting for cellular cofactor activity, the -4 Pro residue appearstobeimportant forcapsid protein function. Protein processing of virus-specific polypeptides in cells infected with mutant virus Se1-1B-06, which contains a
Pro-to-Alaamino acid substitution atthe -4position of the VPO/VP3 site, is similartothat for the wildtype. However, mutant virus Se1-1B-06 exhibited a temperature-sensitive
defect inviral growthat39°C. It is likely, therefore, that the -4Pro residue(located atthe carboxy terminus of VP2) is structurally important for viral capsid protein function. Previous studies havereported defects in viralcapsid
assem-bly (1) and viral RNAencapsidation and release (14) result-ing from alterations at the carboxy terminus of capsid proteinVP3 andattheamino terminus of VP1,respectively. These observationssuggestafunctional requirementfor the
Pro residueatthe -4position of the VPO0VP3 cleavagesite (conserved in all three serotypes of poliovirus) despite a
resulting dependency on cellular cofactor activity for
effi-cient3CD-mediated cleavage atthat site.
Asaresult ofongoing effortstopurifythe cellular cofactor to homogeneity, preliminary data show that the cellular cofactoractivity is 3CDdependent and, therefore, doesnot
appeartobeaproteaseitself. Cellular cofactoractivity also
has a saltrequirement (100to 150 mMKOAc) for activity. Although the cellular cofactor requires salt for activity and
appears stable at high salt concentrations, salt
concentra-tions above 250 mM partially inhibit its activity (data not
shown). The molecular weight of the cellular cofactor is
presently unknown, butthe cofactor isexpectedtobelarger than 12 kDa in size. Extracts from HeLa S10 (data not shown)and HeLaS200andfractionatedHeLaS200extracts dialyzed in HD buffer priorto analysis (Fig. 3B) by using dialysis tubingwitha12-to14-kDa-molecular-size exclusion retained cofactor activity. The latter observations exclude thepossibilitythat cofactoractivityresults from the addition of small molecules such as divalent cations, nucleotides, NAD+, or other small coenzymes to the diluted cleavage reaction mixtures described in this study. Thepresence of cofactor activityin HeLa S200 extracts (devoidof cellular membranes) and the observed inefficient cleavage of P1
precursors incubated in diluted cleavage reaction mixtures
containing 150 mM KOAc and canine microsomal
mem-branes(atfinal concentrations of 0.2eq/,ul)demonstrate that cofactor activity is not exclusively attributable to cellular membranes(datanotshown). Additionally,the detectionof cofactor activity in specific fractions after HeLa S200
ex-tractsweresubjectedto ionexchange chromatography sug-geststhatthe cellular cofactor isaspecific proteinorprotein
complex. A recent study reported the association of heat shockprotein70 withpoliovirusP1 ininfected cells(20),but preliminary evidence suggests that heat shock protein 70
maynotexhibit cofactor activity(2a). Mechanistically, the cellular cofactormayact tostabilize 3CD-P1 interactionsby facilitating the formation of a cofactor-3CD-P1 proteolytic complex. Alternatively, the cellular cofactormay act inde-pendentlyonP1orproteinase3CDtoinducearecognizable
substrate conformation orto confer P1 cleavage activityto 3CD.
ACKNOWLEDGMENTS
We are grateful to Holger Roehl for critical comments on the
manuscript andtoHung Nguyen forexperttechnical assistance. This workwassupported by Public Health ServicegrantAI22693
from theNational Institutes of Health. W.S.B. wasa predoctoral
trainee of the Public Health Service (grant A107319). X.L. was
supported bytheChineseVisiting Scholars Program of the Irvine HealthFoundation.
REFERENCES
1. Blair,W.S., S.-S. Hwang, M. F. Ypma-Wong, and B. L. Semler. 1990.Amutantpoliovirus containinganovelproteolytic cleav-agesitein VP3 is altered in viralmaturation. J. Virol. 64:1784-1793.
4.
3-0.
-1
J.VIROL.
a
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.76.543.75.241.2]POLIOVIRUS P1 CLEAVAGE COFACTOR 2343 2. Blair, W. S., X. Li, and B. L. Semler. 3CD cleavage of the
poliovirus P1 precursor: amodel forcomplex proteinase/sub-strate interactions. In L. Carrasco, N. Sonenberg, and E. Wimmer(ed.), Regulation of gene expression in animal viruses, in press. Plenum Press, London.
2a.Blair, W. S., D. G. Macejak, P. Sarnow, and B. L. Semler. Unpublishedobservations.
3. Blair, W. S., and B. L. Semler. 1991. Role for the P4 amino acid residue in substrate utilization by the poliovirus 3CD protein-ase.J. Virol.65:6111-6123.
3a.Blair,W.S., and B. L. Semler. Unpublished obsevations. 4. Brown, B. A., and E.Ehrenfeld. 1979.Translationofpoliovirus
RNA in vitro:changes incleavage pattern andinitiationsitesby ribosomal saltwash.Virology97:396-405.
5. Campos, R., and L. P. Villarreal. 1982. An SV40 deletion mutant accumulates late transcripts in a paranuclear extract. Virology 119:1-11.
6. Cordingley, M. G., P. L. Callahan, V. V. Sardana, V. M. Garsky, and R. J. Colonno. 1990. Substrate requirements of human rhinovirus 3Cprotease for peptidecleavage in vitro. J. Biol. Chem.265:9062-9065.
7. Dorner, A. J., B. L. Semler, R. J. Jackson, R. Hanecak, E. Duprey, and E. Wimmer.1984.In vitrotranslationofpoliovirus RNA:utilization ofinternalinitiation sitesinreticulocyte lysate. J.Virol. 50:507-514.
8. Emini, E. A., M. Elzinga, and E. Wimmer. 1982. Carboxy-terminalanalysis of poliovirus proteins: terminationof
poliovi-rusRNAtranslationandlocation ofuniquepoliovirus polypro-tein cleavagesites. J. Virol.42:194-199.
9. Haller, A. A., and B. L. Semler. 1992. Linker scanning muta-genesisof the internalribosomeentrysite ofpoliovirusRNA. J. Virol.66:5075-5086.
10. Hanecak, R., B. L.Semler, C. W. Anderson, and E.Wimmer. 1982. Proteolytic processing of poliovirus polypeptides: anti-bodies to polypeptide P3-7c inhibit cleavage at glutamine-glycine pairs.Proc.Natl. Acad. Sci. USA 79:3973-3977. 11. Harris, K. S., S. R.Reddigari,M.J. H.Nicklin, T.Hammerle,
and E. Wimmer. 1992. Purification and characterization of poliovirus polypeptide 3CD, aproteinase and a precursor for RNApolymerase.J. Virol. 66:7481-7489.
12. Inouye, S., and M. Inouye. 1987.Oligonucleotide-directed site-specific mutagenesis using double stranded plasmid DNA, p. 181-206.In S. A. Narang (ed.), Synthesis and applications of DNA and RNA. AcademicPress, Inc., New York.
13. Jore, J., B. DeGeus,R.J.Jackson, P. H. Pouwels,and B. E. Enger-Valk. 1988. Poliovirus protein 3CD is theactive protease forprocessing of theprecursorprotein P1invitro. J.Gen. Virol. 69:1627-1636.
14. Kirkegaard, K. 1990.Mutations inVP1ofpoliovirusspecifically affect both encapsidation and release of viral RNA. J. Virol. 64:195-206.
15. Korant, B.D., J. Langer,andJ.Powers.1980. Proteinsynthesis and cleavage in picornavirus-infected cells, p.257-288. In G. Koch and D. Richter (ed.), Biosynthesis, modification and processing of cellular and viralpolyproteins. Academic Press, Inc.,New York.
16. Kriiusslich, H. G., C. Holscher, Q. Reuer, J. Harber, and E. Wimmer.1990. Myristoylation of thepoliovirus polyproteinis requiredforproteolytic processing of thecapsid and for viral
infectivity. J. Virol. 64:2433-2436.
17. Laemmli, U. K.1970.Cleavageofstructural proteinsduring the assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
18. Larsen, G. R., C. W. Anderson, A. J. Dorner, B. L. Semler, and E.Wimmer. 1982. Cleavage siteswithin the poliovirus capsid protein precursors. J. Virol. 41:340-344.
19. Lawson, M. A., B. Dasmahapatra, and B. L. Semler. 1990. Species-specific substrate interaction of picornavirus 3C pro-teinase suballelicexchange mutants. J. Biol. Chem. 265:15920-15931.
20. Macejak, D. G., and P. Sarnow. 1992. Association of heat shock protein 70 with enterovirus capsid precursor P1 in infected humancells.J. Virol.66:1520-1527.
21. Nicklin, M. J. H., K. S. Harris, P. V. Pallai, and E. Wimmer. 1988.Poliovirusproteinase 3C: large-scale expression, purifica-tion, and specific cleavage activity on natural and synthetic substratesin vitro. J.Virol. 62:4586-4593.
22. Nicklin, M. J. H., H. G. Krausslich, H. Toyoda, J. J. Dunn, and E.Wimmer. 1987. Poliovirus polypeptide precursors: expres-sion in vitro and processingby exogenous 3C and 2A protein-ases. Proc. Natl.Acad. Sci. USA 84:4002-4006.
23. Nicklin, M. J. H., H. Toyoda, M. G. Murray, and E. Wimmer. 1986. Proteolytic processing in the replication of polio and related viruses. Bio/Technology 4:36-42.
24. Pallai, P. V., F. Burkhardt, M. Skoog, K. Schreiner, P. Baxt, K. A.Cohen, G.Hansen, D. E. H.Palladino,K. S.Harris,M.J. Nicklin, and E. Wimmer. 1989. Cleavageofsyntheticpeptides by purifiedpoliovirus3Cproteinase. J. Biol. Chem. 264:9738-9741.
25. Semler,B.L.,C. W. Anderson, N. Kitamura, P. G. Rothberg, W. L. Wishart, and E. Wimmer. 1981. Poliovirus replication proteins: RNA sequence encodingP3-lb and the sites of pro-teolyticprocessing. Proc.Natl. Acad.Sci. USA78:3464-3468. 26. Toyoda, H., M. J. H.Nicklin, M. G. Murray, C. W. Anderson, J. J. Dunn, F. W. Studier, and E. Wimmer. 1986. Asecond virus-encodedproteinase involvedinproteolyticprocessingof poliovirus polyprotein. Cell 45:761-770.
27. Urzainqui, A., and L. Carrasco. 1989. Degradation ofcellular proteinsduring poliovirus infection: studiesbytwo-dimensional gelelectrophoresis.J. Virol.63:4729-4735.
28. Vaheri, A., and J. S. Pagano. 1965.Infectious poliovirusRNA:
asensitive method ofassay.Virology27:435-436.
29. Ypma-Wong, M. F., P. G. Dewalt, V. H. Johnson, J.G.Lamb, and B. L. Semler. 1988. Protein 3CD is themajor poliovirus proteinaseresponsibleforcleavage of the P1 precursor. Virol-ogy166:265-270.
30. Ypma-Wong, M. F., D. J. Filman, J. M. Hogle, and B. L. Semler. 1988. Structural domainsofthepoliovirus polyprotein
are major determinants for proteolytic cleavage at Gln-Gly pairs.J.Biol. Chem.63:17846-17856.
31. Ypma-Wong,M.F.,and B. L.Semler. 1987. In vitromolecular genetics as a tool for determining the differential cleavage specificities ofthepoliovirus3Cproteinase. Nucleic Acids Res. 15:2069-2088.
32. Ypma-Wong, M. F., and B. L. Semler. 1987. Processing deter-minants required for in vitro cleavage of the poliovirus P1 precursortocapsid proteins.J. Virol.61:3181-3189.
VOL. 67,1993




![FIG. 6.withcells4).(lanesnine,and [35S]methionine pulse-labeling analysis of cells infected PV1 or Sel-1B-06](https://thumb-us.123doks.com/thumbv2/123dok_us/1302386.83382/6.612.127.226.77.435/withcells-lanesnine-methionine-pulse-labeling-analysis-cells-infected.webp)