Copyright © 1988,American SocietyforMicrobiology
A
Deletion Mutant of L-A Double-Stranded
RNA
Replicates
Like M1
Double-Stranded RNA
ROSA ESTEBAN* ANDREED B. WICKNER
Section onGeneticsof SimpleEukaryotes, Laboratory ofBiochemicalPharmacology, National InstituteofDiabetesand
Digestive andKidney Diseases, Building 8, Room209,Bethesda, Maryland 20892 Received 16October1987/Accepted29 December1987
Xdouble-strandedRNA(dsRNA)isa0.52-kilobasedsRNA moleculethatarosespontaneouslyinanonkiller strainofSaccharomyces cerevisiae originally containingL-Aand L-BC dsRNAs(L-BCisthesamesizeasL-A butsharesnohomologywithit). X hybridizedwithL-A,and direct RNAsequencingofX showed that thefirst 5' 25basepairs(of theXpositive strand)andatleast thelast 110 basepairsof the 3' endwereidentical tothe ends of L-A dsRNA. X showed cytoplasmic inheritance and, like M1, was dependent on L-A for its maintenance. Xwasencapsidatedinviruslikeparticleswhosemajorcoatprotein wasprovided byL-A(asis trueforM1),and Xwasfound in viruslike particleswithonetoeightXmolecules perparticle. Thisfinding confirmsour"head-fullreplication"modeloriginally proposedfor
Ml
andM2.LikeM1orM2, X lowersthecopynumber ofL-A, especiallyinaski host.Surprisingly,Xrequiresmanychromosomal MAKgenesthatare necessaryforM1 butnotforL-A.
Uptofive different double-stranded RNA(dsRNA) fami-lies can be found in strains of the yeast Saccharomyces cerevisiae,namely,L-A, L-BC, M, T, andWdsRNAs,three ofwhich (L-A, L-BC, and M) areencapsidated in
intracel-lular viruslike particles (VLPs) about 39 nm in diameter.
Genes that code for killer toxin production and resistance (immunity) reside on M dsRNA, the size of which varies around 1.5(M2)or1.8 (M1)kilobases(kb), dependingonthe killer determinant. M maintenance or replication requires
the presence of L-A dsRNA (4.5 kb), which encodes the majorcoat protein in which both M and L-A dsRNAs are
separately encapsidated. WhereasMalso needs theproducts ofatleast32 chromosomalgenes(calledMAK[maintenance ofkiller] genes), L-Areplication requires onlythreeof these
genes, namely, MAK3, MAKIO, and PET18. Other genes
(called SKIgenes) negatively control dsRNA replication in such a way that ski mutants show a superkiller phenotype
withanincreased Mcopynumber (reviewedinreferences 21
and22).
Deletion mutantsof M1 dsRNA lacking the toxin immu-nityprecursorcoding region have been described (5, 6, 10, 12). These mutants (called S dsRNAs) interfere with (sup-press) the replication of the wild-type M1genome (9, 10). S
dsRNAs, therefore, are analogous to defective interfering virus particles, the deletion mutants ofanimal viruses that limitreplication of the standard virus during coinfection (for
a review, see reference 19). Unlike defective interfering
particles, however, S dsRNAs donotdependontheparent
M dsRNA for their replication; rather, both M and S are
dependenton L-Aformaintenance orreplication, probably
because both are encapsidated in VLPs whose majorcoat
protein is L-Aencoded.
All S dsRNAs characterized so far are derived from M1
dsRNA. Partial sequence analysis (12) and heteroduplex
analysis (5) ofone ofthem, S3, shows that 229 base pairs (bp)arefromthe 5' end of theM1positivestrand and therest
(about 550 bp) are from the 3' end of the positive strand.
Another S dsRNA, S14, has been cloned and completely sequenced(6). The break points in S3and S14areonly 21 bp
*Correspondingauthor.
apartin the 5' end of theM1 sequenceand10 bpapartin the 3' end. All the S dsRNAspreserve aportion of the 5' end of the M1 positive strand (a minimum of 44 bp) and a much larger portion (up to 550 bp) of the 3' end of the positive strand. The fact that S dsRNAs retain both ends of the original molecule implies that these ends probably play an
essential role in the processes known to be necessary for these moleculestobemaintained: transcription, replication, andencapsidation.
Since L-A produces the capsid protein essential for its
own encapsidation, one might expect to find L-A deletion mutants equivalentto the S mutants, but that would notbe completely suppressive,sincetheparentalmoleculeremains essential. Such L-A deletions have not, until now, been
observed.
We report hereX,the first deletionmutantof L-AdsRNA. We show thatX is similarto S mutantsin that itoriginated
as aresultofalargeinternaldeletionof theparentmolecule. Whereas X is derived from L-A, its replication and its relationtohostgenesresemble those of Mmorethenthose of L-A.
MATERIALS AND METHODS
Strains and media. Strains of S. cerevisiae used in this study are listed in Table 1. Media were as previously
described(20).
Genetic methods. Standard methods of genetic analysis
wereused (7).
Cytoduction. A cytoplasmic genome can be transferred fromonehaploidstrain toanother withoutdiploidizationor
otherchangeofnucleargenotypeby transient heterokaryon formation (cytoplasmic mixing, cytoduction) by using the karl-imutant,defective in nuclear fusion(2). Theprocedure
wasthesame asthatdescribed by WesolowskiandWickner
(18).
Assay ofkilling activity. Killing activity was assayed as
previously described (4).
VLPpreparationand RNApolymerase activityassay.VLP
purification was as previously described (4), and the RNA
polymerase activity associated with VLPs was assayed by
themethod of Welshetal.(17), with slightmodifications(4). 1278
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Strains ofS. cerevisiaeused
Designation Genotype Sourceof reference
a his4 karl-I L-A-HNBa L-BCM1
ahis4 karl-i L-A-HNB L-BCM-o
aargi ski2-2 L-A-oL-BC M-o
aski2-2L-A-HNB L-BC M-o X
aleul karl-iL-A-orhoo a his4karl-i L-A-HNL-BC
Ml
aargi ski2-2 L-A-o L-BC M-o
RE411 aleulL-A-HNB L-BC X
RE455 aargi karl-iski2-2
L-A-HNB L-BC X 2285 akarl-i leulclo- L-o M-o
RE327 ahis4 leulkarl-i
clo-L-A-HNB L-BC-o M1 RE464 aargi L-A-HNL-BCX RE516 aargi ski2-2 clo-L-o
RE558 ahis4 leul karl-i
clo-L-A-HNB L-BC-o X 3950-3B aade2leu2 mak2l-l
2597 aura3trpl adel makl6-1 2593 aleu2ura3makl8-1
2611 alysi mak27-1
3958-3B aura3mak26-1
1868 aleu2 cdci6 makll-l
TF78-4A aural leu2cani maklOts
2321 aaro7mak3-1 leul his3
ura3 2444 aura3mak6-1
2540 atyri makl-2
2362 alys2 adel mktl mak4-1
1747 aarg9maklO-I L-o
2186 amkti lysi L-A-o L-B
1686 aargi thrl L-A-oL-C
YHU-1652 aura3 hisS L-A-HN M-o
P24-28C--1020; 16 1368
AK-Meiotic segregant
fromcross2404
x 2507
A364A-*1020 Meioticsegregant
fromcross 1368
x 2507AK-
H-Segregant from
cross64-4B x
1089 RE411--RE57 This work
This work
This work RE464-*RE327
16
sis in5% acrylamide gels aspreviously described (14), with slightmodifications. RNA was denatured at 90°C for 1minin the presence of 7 M ureainstead of30% dimethylsulfoxide.
The samples were quicklychilled on ice and applied to the gel. Positive and negative strands were separated at 8 V/cm for 6 to 8 h.
32P-labeled positive-strand X ssRNA obtained in an in vitrotranscription reaction performed by purified X VLPs was used to determine which band was the positive and which was the negative strand. The faster-moving band was thepositive strand.
RNA sequencing. The 3'-end-labeled positive- and nega-tive-strand X ssRNAs separated in a 5% polyacrylamide gel as described above were cut and eluted as follows. After
completion of electrophoresis, thegelwasimmersed inTBE buffer (90 mM Tris borate [pH 8.0] with 4 mM EDTA) containing 0.5 ,g of ethidium bromide per ml for 15 min. ssRNAbands were visualized and photographed under UV light(approximately 350 nm). The gel then was subjected to
autoradiography for1min at room temperature, and labeled bands werelocalized. They were cut out from the gel with an
RNase-freerazorblade, crushed on a sterile glass plate, and extracted overnight with 0.5 ml of 0.5 M ammonium acetate-1 mM EDTA at37°C.Extracted RNAs were cleaned
fromsmallpieces ofcontaminatingpolyacrylamide by
filtra-tionthroughglass wool, andthen2.5 volumes of ethanol was
addedtoprecipitatetheRNAs. Afterbeingwashed with 70%
ethanol,RNAs were dissolved in 10 to 20
RI
ofdiethylpyro-carbonate-treated H20 and used to determine the RNA sequence.
The enzymes used to partially digest the RNA and their basespecificitieswere asfollows: RNaseT1 (G), RNase U2 (A), RNasePhyM(A +U), RNaseBacilluscereus(C +U), and RNase CL3 (C). They were purchased in an RNA
sequencing kitfrom BethesdaResearch Laboratories. Con-ditions used for partial digestions and separation of the products by acrylamide gels were as recommended by the manufacturer.
Transfection withpurifiedVLPs.Transfection experiments
with purified X VLPs were carried out by the spheroplast
procedure of El-Sherbeini and Bostian (3).
aNatural variants of L-A dsRNA carrying various combinations of the
[HOK](orH), [NEX](orN),[EXL](orE), and[B]cytoplasmicgeneshave
been identified(reviewedinreference21;Uemura andWickner,inpress). dsRNA purification. dsRNAs were purified by cellulose
chromatography as previously described (15). To analyze
dsRNA directly in cells, the rapid method for extraction describedby Fried and Fink (5)wasused, exceptthat LiCI
precipitationwasomitted;extracted dsRNAswereanalyzed on1to1.5%agarosegels. When thepresence orabsence of certain dsRNAs could not be unequivocally established because ofthelarge amount of rRNA and tRNApresent in the samples, samples were digested with pancreatic RNase A(10 ,ug/ml) for1 hat37°Cinhigh salt (0.5 MNaCI)before
agarosegel electrophoresis. Under theseconditions, single-stranded RNA (ssRNA)moleculesarecompletely digested,
whereas dsRNAs areresistant tothe treatment.
Labelingofthe3' ends of RNA. The 3' termini of dsRNA
werelabeled withcytidine[3',5'-32P]diphosphate by usingT4 RNA ligase under conditions suggested by the supplier (Bethesda Research Laboratories, Inc.).
The strands of X dsRNA bearing 3'-terminal cyti-dine[3',5'-32P]diphosphate were separated by
electrophore-RESULTS
X is a deletion mutant of L-A. In the course of another study, total nucleic acids fromtwotetradsofthecross2404
x 2507 (ahis4 karl L-A-HNB L-BC x aargl ski2-2 L-BC) were extracted and analyzedon a1.5%agarosegel (Fig. 1).
We noted aband (called X)migratingat0.52 kb(relativeto lambdaHindIlI fragments) only in the case ofspore clone 64-4B(Fig. 1,lane6).XwassensitivetoRNase A in low salt butnotin high salt (datanotshown), indicating that it isa
dsRNAmolecule.
The amount ofL dsRNA in strain 64-4B was very low comparedwith that in the othersporeclones from thesame
tetrad, even lower than in the SKI wild-type segregants.
Strain 64-4B is a ski2-2 segregant, and the amount of L dsRNA(mainly L-A)in suchstrainsis about fivefoldhigher than in wild-type strains (1) (compare lanes 1, 3, 5, and 7 with lanes2, 4,and8, Fig. 1).Thecopynumber ofXdsRNA (0.52 kb) in strain 64-4B wasevengreaterthan thatofL-A (4.5 kb) in otherski2-2 segregants.
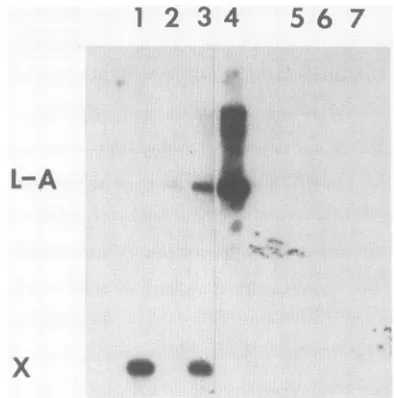
We showedbyNorthern(RNA)blothybridizationthatX dsRNA hadhomology with L-A. L-A VLPs wereprepared
from a clo L-A L-BC-oM1 strain (RE327)
(clo
strainslose L-BC), andthe VLPtranscriptasewasusedtomakeanL-A 13682404 2507 64-4B
1089 1101 RE57
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.62.298.92.502.2]positive ssRNAprobe. Strong hybridization with X dsRNA
(lanes1and3,Fig.2) and L-Afromacontrol strain(lane4) was observed, but no hybridization was seen with several
L-BC preparations (lanes 5, 6, and 7, Fig. 2). Thus, X is a deletion mutantofL-A. Further proof ofthisconclusion is presented below. A small amount of full-length L-A was detected in the strain carryingX (lane 3). Its copy number was only about 1/20 thatofX in the same strain. This and otherdata (see below) showthat Xlowers the copy number
ofL-A. Strain 64-4B (carrying X and L-A) was subcloned
three times in succession, and nucleic acids from six
sub-clones wereanalyzed byNorthern blothybridization asfor
Fig. 2. All clones gave the same resultas in lane 3, Fig. 2
(data not shown). Thus, X and L-A are
stably
maintained together.Xdependson L-Aforits maintenanceand isindependent of L-BC. The dependence ofX on L-A for its maintenance, replication,orbothwastestedbytwomethods.(i)Usingthe
mak3 and maklOfs mutant strains 2321 and TF78-4A(Table 1), which are unable to maintain L-A dsRNA, we showed that neither L-AnorXwasfound after introductionof both
molecules by cytoduction into such strains and that by meiotic crosses the segregation pattern obtained was 2L-A
X:2L-A-o X-o. When X was introduced into the MAK3 or MAKJO wild-type strains, all cytoductants acquired X, and
in meioticcrosses, alltetradswere4X:OX-o.Thisalsomeans that X, like L-A, shows nonchromosomal inheritance. (ii)
Growth of strain RE455(aargi ski2-2 L-A L-BCX)at39°C
1
2
34
5
6
7
8
DNA
rRNA
rRNA
x
ABC
DA
BC
D
ski
+ - + -+
- +FIG. 1. Initialdetection of X dsRNA. Total nucleic acids were
extracted from clones
~of
two tetrads dissected from the diploidstrain 2507
x
2404
(a
argi
ski2-2 L-BC
x
othis4-15
karl-i L-A
L-BC)andanalyzedon a1.5% agarosegel.Lanes 1to4areclones fromtetrad1,andlanes5to8areclones fromtetrad 4.A, B,C,and Dindicatethe fourspore clones of eachtetrad. Thesymbols -and+ represent ski2-2and wild-typeSKI+ meioticsegregants, respec-tively. Lmeansmainly L-AdsRNA;onlyaminoramountofL-BC ispresentinthesepreparationsandisindistinguishablefromL-Aby size on thistype ofgel. X is thespontaneously dele'ted L dsRNA moleculepresentonlyin spore clone64-4B (lane 6).
1
2
34
567
L-A
."W [image:3.612.336.533.77.276.2]x
FIG. 2. Northernblothybridizationof X anddifferentL dsRNA preparationswitha32P-labeledL-ApositivessRNAprobeobtained
as an in vitro transcript of purified L-A VLP-associated RNA polymerase from strain RE327 (a his4 leul karl-i clo L-A M1 L-BC-o).Lanes: 1,XdsRNApurifiedfrom clone64-4B (Fig.1,lane 6)by electroelutionfromanagarosegel;2,LdsRNA from thesame
clone, alsoelectroeluted from the gel; 3, total nucleic acids from clone64-4B(thesamesampleasinlane 6 inFig. 1);4,crude extract of clone64-4D(lane8 inFig. 1)thatdoes not contain XdsRNA;5, 6,and7,total nucleic acids extracted from strains 2186(L-A-oL-B), 1686(L-A-oL-C), and 2507 (L-A-oL-BC),respectively. Thesewere included toconfirm that the L-Aprobewasnotcontaminated with L-BC.
for 3 days resulted in loss of both X and L-A from most clones. Whereas loss of X without loss of L-A was often
observed,therewas nocaseinwhich XdsRNAwasfound in theabsence ofL-A, indicating that X depends on L-A(data not shown). As we show below, X is encapsidated in the L-A-encodedmajorcoatprotein, and this is presumablywhy
X requires L-A. By Northern blot hybridization, wefound that when X was curedby growth at high temperature, the copy number of L-A increased to the level of the parental
X-o strain. Thisagain shows that X lowers the copy number ofL-A dsRNA.
In thecase ofL-BC, the same type of experiments were carried out
by
cytoducingX into strain 2285 (a karl-i leul clo-L-o). Xwasmaintained in all cytoductants eventhoughL-BC wasabsent. That L-BCwas absent in those cytoduc-tants was proved by cytoducing their cytoplasm again into themakJO-I L-o strain 1747. When analyzed on agarosegels,
those cytoductants were shown to be L-o, and since the maklO mutation only affects L-A maintenance, not L-BC,
thedonors must havelacked L-BC.
X isencapsidated in L-A-encoded VLPs. Stationary-phase
cells of a 4-day culture were used to prepare VLPs from strain RE455. This strain contains L-A, minor amounts of L-BC, and X dsRNAs (Table 1). The last step in the purification is acesium chloride densitygradient
equilibra-tion sedimentation (4), andparticle bands are located in the gradient according to their density, which depends on the RNA/protein ratio of the particles.
Figure 3shows an ethidiumbromide-stained agarose gel of the dsRNAs of the VLPs present in thefractions collected fromacesiumchloride gradient (A) and anautoradiogram of theproducts of the RNApolymeraseactivity associated with them(B).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.82.275.373.630.2]A
Bottom
L-A
dsRNA
X
dsRNA-X
ssRNAO
B
Top
(W
L-A
ssRNA
X
dsRNA%,
X
ssRNA'---5 7 9 11 13 1ssRNA'---5 17 19 21 23
6 8
10 12 14 16 18 20 22
FRACTION
NO
FIG. 3. Ethidium bromide-stainedagarosegel(A) and autoradio-gram (B) of the dsRNAs contained in purified VLPs from strain RE455 by cesium chloride density gradient centrifugationand the products of theRNApolymerase activity associated withthe VLPs. The bottom shows the denser fractions, and the top shows the lighterones. A sample of eachfractionwas incubatedin an RNA polymerase reaction mixture that contained[32P]UTP,andthe other three ribonucleotides for 2 h at 300C. After completion of the reaction, RNAs were phenol extracted and separated on a 1.5% agarosegel, and the gelwassubjectedtoautoradiography. Inpanel A, aband of dsRNAis seen,peakingin fraction 14, whose molecular size is 2.3 kb, consistent with that of W dsRNA (18), which is presentin this strain. Stains lackingL-Aand L-BC do not show a W-size band atthis place in the gradient even though the strain carriesWdsRNA.Wedonothave direct evidencethat thisband is WdsRNA.
There was only one type of L-A-containing VLP; it banded around fraction6, anditsdensitywas, aspreviously reported, 1.4076(4). However, Xwasfoundthroughoutthe
cesium chloride density gradient. That Xwas encapsidated inside VLPs and wasnot naked dsRNAwas proved bythe RNA polymerase activity present in each fraction that
synthesizedXdsRNA inadditiontoXpositivessRNA(Fig. 3B). Furthermore, the density of naked dsRNA is higher
than thatoftheparticles,andin this typeof cesiumchloride
gradientitwouldpelletatthe bottom ofthe tube. By sodium dodecyl sulfate-polyacrylamide gel electrophoresis, we foundthat XVLPshad thesameprotein compositionasL-A VLPs, withamajor coatproteinsubunit of about 81 kilodal-tons.
"Head-full replication" ofXdsRNA.M1dsRNA,which is less than half the sizeof L-A,is found in VLPs with either one or twoM1 dsRNA molecules per
particle.
These arise because M1 positive ssRNAtranscripts
made in VLPscarrying only one M1 dsRNA molecule are often retained within the VLP, where they are then copied to make a second M1 dsRNA molecule. VLPs with two M1 dsRNA
molecules per particle are full, however, and all newpositive strands are extruded from the particles (4). This head-full replication mechanism predicts that any small genomelike X dsRNAreplicating in these VLPs with an L-A coat should generate multiple VLP species, in this case, the largest integer less than (molecular weight of L-A dsRNA)/(mo-lecularweight of X dsRNA). The results of Fig. 3 and 4 show that this prediction was fulfilled. The banding of X VLPs throughout the gradient was due to the presence ofmultiple discretespecies with different densities. Rebanding of three differentfractions from the initial cesium chloride gradient of Fig. 3 in threenewcesium chloride gradients (Fig. 4) showed that they banded true and all three had RNA polymerase activity. Equal aliquots of gradient fractions were used for measurementof dsRNA (Fig. 4, left), RNA polymerase (Fig. 4,right),andVLPproteins (Fig. 4, right, insets). These data show that thedifferences in the banding densities of different VLPfractions are due to differences in the ratio of RNA to protein in the VLPs. Six peaks of X positive
ssRNA-synthesizing activitywereobserved in the gradient shown in
Fig. 3(five peakscanbeseeninthefigure, and a sixth peak was present in fraction 26 [not shown]). The density of particles peaking in fraction 9 corresponds to seven X dsRNA molecules per particle. A small amount of
X-con-taining particleswaspresent atthe same density as the L-A VLPs, although it did not form a discrete peak.
Theratio of RNA polymerase activity producing X ssRNA and dsRNA(Fig. 3B)to XdsRNA (Fig. 3A) was highest in the upper part of thegradient and decreased with increasing
density. Thisis the expected result of particles with several XdsRNAtemplate molecules per particle are limited in the RNA synthesized by having only one RNA polymerase molecule.
Also aspredicted by the head-full replication mechanism, XdsRNA synthesis occurred in particles ofmany distinct
densities in the cesium chloride gradient of Fig. 3. Thus,
40 20
0
U
40 Z
I--20 M
e4 40C.)
x
u n
120
°5 10
FRACTION NO
FIG. 4. XVLPsof fractions9(panels A), 17(panels B),and 26 (panelsC) fromthecesium chloridegradient shown inFig.3 were
subjected to three independent cesium chloride density gradient equilibrium sedimentations.Panels: left, onesample of eachof the fractions collected wasphenol extracted andseparated on a1.5% agarosegel;right, RNApolymerase activityassociated with the X VLPsasmeasuredby incorporationof[32P]UTPinto acid-insoluble material. The insets show protein content in main peakfractions analyzedbysodiumdodecylsulfate-polyacrylamide gel electropho-resis. The samevolumes insamplesA, B,andCwereused for all measurements(dsRNA, protein, and RNApolymerase activity).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.63.301.77.365.2] [image:4.612.323.565.459.635.2]TABLE 2. Effect of mak mutationsonXmaintenance
X-containing [B] strain that is: X-containing SKI'strain thatis:
mak
SKI+
ski mak [B] [B-0]mutant X No. of No.of mutantX No. of No. of
tetrads Meiotic tetrads Meiotic No.raof Meiotic tras Meiotic
analyzed segregation analyzed segregation analyzed
egation
analyzedsegregation
makl8-la 2 22X:2X-o 2 1 3X:1X-o makl2d 3 3 4X:OX-o 3 14X:OX-0
14X:OX-o 1 3X:1X-O
mak2-Pla 2 2 2X:2X-o 2 2 3X:1X-o 1 2X:2X-o
mak26-Ja 2 22X:2X-o NDb mak4-1 3 3 4X:OX-o 3 1 4X:OX-o
mak27-la 2 2 2X:2X-o ND 2 2X:2X-o
makJ6-Jc 3 32X:2X-o 3 3 3X:1X-o mak6-1 2 14X:OX-o 3 3 2X:2X-o
12X:2X-o
makll-l 2 1 4X:OX-o 3 23X:1X-o
13X:1X-o 1 4X:OX-o
amak mutations belonging to this group arebypassed byaski mutation in thecaseofM1(16) butnotby [B].Themakl8,mak2l,mak26,andmak27 strains
used were strains 2593, 3950-3B, 3958-3B, and 2611,respectively.
bND, Not done.
Cmakl6-1in the case ofMlwasbypassed by neither ski nor [B]. The result obtained withthe
makl6-1/+
ski2-21+Xdiploidstrain meioticsegregationof 3X:1X-o is thusdifferent from that of M1. The makl6 strain2597 wasused.dmakmutations bypassed by either skior[B]. Themakl, mak4,mak6,andmakilstrains usedwerestrains2540, 2362, 2444,and1868,respectively. ratherthanpreformed XdsRNA molecules beingpackaged
at more than one molecule per particle, we believe that a
singleXpositivessRNA ispackaged and that this molecule
replicates until the package is full.
Xand M1 are incompatible. We studied the behavior of
Ml
andXwhenpresenttogether inthe samestrain. Strains1101
(a his4 karl-i L-AL-BCM1) andRE411 (a leul L-AL-BC
K+
K
L-A
x
1 2 3456 78
FIG. 5. X and M1 incompatibility. Total nucleic acids were
extractedfrom K+(samples 1 and 2) andK-(samples3to8)diploid coloniesobtainedby matingtheX-containing strainRE411and the killerstrain1101 andseparatedon a1.5%agarosegel.Xwaspresent
only inthe K- derivatives, and these contained no M1. All K+
colonieswereX-o.
X) were allowed to mate on YPAD completemedium for 1
dayat30°C; diploidsingle colonieswereselectedby growth
onminimalmedium(SD),and their killeractivitywastested. Over90%of thediploidcolonies were K- (nonkillers); only a few showed the normal killer phenotype of the parental strain 1101; someofthem wereweakkillers. After subclon-ing, nonkillers and normal killers were stable, but weak
killerssegregatedmainly nonkillers andafew normal killers. Wepurified total nucleic acids fromanumberofK+ and K- subclones picked randomly from the SD plates and
analyzedthem on1.5%agarosegels (Fig. 5). Samples1 and 2 were K+, and samples 3 to 6 were K-. Asexpected, M1
dsRNAwasonlypresent insamples1and2,butthey didnot contain X dsRNA.X dsRNA waspresentintheK- samples
(samples 3 to 6), and M1 was lost. Thus, X and M1 are
incompatible.
Wefurther studied thisphenomenon ofincompatibility by
crossing
otherX-andM1-containing strains,
someof whichwere
SKI'
and someofwhichwereski-.Althoughwefoundsome differences in the ratio of nonkiller diploids to killer
ones, nostrain that couldstably maintainXandM1together
was found. In all crosses, most diploid cells lost M1 and retained X,withonly aminority losingXand retaining M1. This effectwas evenmoredramaticwhen X wasintroduced by cytoduction into anM1-containing strain. Practically all cytoductants became K- because ofthe acquisition ofX.
Xrequiresmany MAK gene products needed byM1but not by L-A. M1 requiresatleast 30chromosomal geneproducts for its maintenance or replication, only 3 of which are needed by L-A (MAK3,
MAKJO,
and PET18). As shownabove, X could not be maintained when introduced into mak3 and maklO mutant strains 2321 and TF78-4A by
cytoduction. The discovery that X also excluded M1 from thecytoplasm ofakillerstrainledus tostudy the role of the other MAKgenes, needed only by M1, on X maintenance.
The MAK genes tested are listed in Table 2. They belong tothree differentcategories,according totheir properties in M1 maintenance. Group 1 includes MAK18, MAK21, MAK26, and MAK27, among others; mutations in such genes show clear2M1:2M-o meiotic segregation when both
parental strains are wild type for SKI genes; they are
bypassed, however, by ski- recessive mutations so that a double mutant(e.g., mak18-1 ski2-2) can thenmaintainM1.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.93.264.402.678.2]Mutations in thisgroup are not bypassed or suppressed by
the cytoplasmic genetic element [B] (16).
maki,
mak4, mak6,makil,
and several other mak mutations belong togroup 2, which is bypassed by either aski- mutation or the extrachromosomal element [B], known to be on L-A (H. Uemuraand R. B. Wickner, Mol. Cell. Biol., in press). Only
the makl6-1 mutation is suppressed by neither ski- muta-tionsnor[B] (group 3).
We crossed strains belonging to these three categories
withappropriate X-containing strains, some of which were
SKI'
andsomeof which wereski-. Aftertetraddissection,total nucleic acids were extracted from spore clones and
analyzed on agarose gels. The results obtained are
summa-rized in Table 2. Briefly, mak mutant group 1 gave the
meiotic segregation 2X:2X-o in all tetrads examined when
both parents were SKI (two or three in each cross); when one of the parental strains was a ski- mutant, the most
frequenttypeoftetradformedwas the tetratype 3X:1X-o,as
expected forsegregation of mutations in two unlinked loci, oneof which bypasses the other. The second groupofmak
mutations was presumably bypassed by the extrachromo-somal element [B], and most of the tetrads showed4X:OX-o segregation. We suspected that all of our X-containing
strains carried [B], because the deletion ofL-A that
pro-duced X occurred in a meiotic segregant of the
[B]-con-taining strain 2404x 2507 (Table 1;Fig. 1). Toconfirm that the result found was due to the [B] present in the
X-containing strain andnot tothefact thatXdidnotrequirethe MAKgene products, wecarried outthe following transfec-tion experimenttointroduceXintoa [B-o] strain.
X VLPs purified from strain RE455 (fraction 26 in the cesium chloride gradient shown in Fig. 3A) were used to
transfect spheroplasts prepared from the L-A-containing, [B-o], X-ostrainYHU1652 (Uemura andWickner, in press) by the method of El-Sherbeini and Bostian (3). Among the URA3+ transformants obtained, 34 were testedfor acquisi-tion ofX VLPs by growth on YPADplates for 2days and analysis of their nucleic acidsonagarosegels.Wefound that
12 (35% ofURA3+ transformants) had acquired X VLPs. We then introduced by cytoduction the X and L-A VLPs
from one X-containing transfectant into standard tester
strains thatwereL-A-oandX-oand used thosecytoductants
tostudy the effect of the mak mutationsbelongingtogroup
2onX maintenance when [B] was notpresent. Theresults obtained withmakl-2, mak4-1, mak6-1, andmakll-l strains
aresummarized in Table 2. When [B]waspresent, mostof the tetrads (8 of10analyzed)were
4X:OX-o,
and when [B]was absent, only a few (3 of 13 analyzed) gave the same
segregation
pattern.
Wealso sawinthiscase anincrease inthe number oftetrads that gave 2X:2X-o meiotic
segrega-tion. Thus, these results confirmed that the MAK genes
belongingtogroup 2 are
required
forXmaintenance,
aswell as forM1,
and that in both cases theserequirements
are bypassed by the cytoplasmic element [B] carried on some L-Amolecules (Uemura andWickner,
inpress).Effectofskimutations on X copy number. SKI genes are repressors ofL-A,L-BC,andM1dsRNA
replication
sothatrecessive ski- mutants show an increase in copy number
compared with
SKI'
strains.Thecopy numberofindividualdsRNA species also depends onthe otherdsRNAs present
inthe samecell.For
example,
M1lowers the copy numberof L-A, andthiseffectiseven stronger inski- strains.The presence ofX in strain 64-4B
(a ski2-2)
lowered the copy numberofL-A(Fig. 1); however,
this straincontained smallamountsofL-BCaswell. To betterstudy
theeffectofaskimutationonXand L-A copy
number,
weused theclo-chromosomaldefect that makes strains L-BC-o, and so the
only L dsRNApresentis L-A. Thecross RE558(ahis4leul clokarl-i L-AL-BC-o X)x RE516 (aargi ski2-2 clo- L-o)
was carried out at 20°C because L-BC dependence onclois temperature dependent (18). Total nucleic acids were
ex-tracted from anumber of tetrads and analyzedon agarose gels (Fig. 6). Wefound that X dsRNA is favored in ski2-2 meiotic segregants, and its high copy number results inan extra decrease in L-A dsRNA as compared with
SKI'
clones. This is similartothe results withM1 and L-A (1). When meiotic segregants ofthe same cross were
incu-batedatlowtemperature(8°C) for15days, we observed no
difference in the pattern of growth between the ski- and
SKI'
clones of each tetrad, although M1-containingski-mutants showed cold sensitivity for growth at the same temperature (8).
Sequence ofthe ends of X dsRNA. X dsRNA extracted from purified X VLPs of strain RE455 (Fig. 3) was 3' end labeled with
cytidine[3',5'-32P]diphosphate
as described in Materials and Methods. Positiveandnegative strands wereseparated ina polyacrylamide gel, recovered from the gel, and usedtodetermine the terminalnucleotide sequenceby partial digestionswithbase-specificRNases.
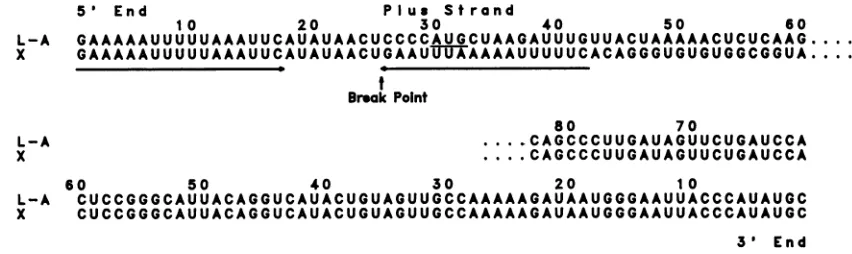
Figure7shows thesequencesof both5'and3'endsofthe
positive X ssRNA. For comparison, the 5'- and 3'-end
sequencesofpositive L-AssRNA arealso included(13). For convenience, the 5'-endsequenceof theX
positive
strand is shown, although direct RNA sequencing data gave us the 3'-end sequenceof thecomplementary negative strand.Both the5'and3'endsof L-AarepresentinX.Inthecase
ofthe 5'end, onlythefirst25bp ofL-A arethe same asthe first25bp ofX.Thebreakpoint occurred only5bpupstream
of the firstputativeopenreading frame initiation codon AUG
atposition30inL-A(underlined in Fig. 7). Beginning with
1 2
3 4
5
6 7 8 9101112
A
B C
DABC
DAB C
D
DNA
L-A
x
ski
_
+.
+
_
+
FIG. 6. Effect of theski2-2mutationonXand L-Acopynumber. Clonesfrom three differenttetrads,dissected from the
diploid
strain RE558 x RE516(a/a
ski2-21+ clo-Iclo- L-BC-o L-A X), weregrownonYPADmedium
plates
at20°C
for4days,
and their nucleic acidswereextracted,treatedwithRNase A in the presenceof 0.5 MNaClas described in Materials and Methods, and
analyzed
ona1.5%agarose
gel.
Inotheragarosegels
(not shown),weanalyzed
thesamples
without RNasedigestion
and used the 26S rRNA as aninternal standardtonormalizetherelative concentration of dsRNA
in each
sample.
A, B, C, and Darethe four sporeclones of eachtetrad. The
symbols
- and + representski2-2 andSKIwild-typesegregants,
respectively.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.319.560.438.624.2]5' End Plus Strand
10 20 30 40 50 60
L-A GAAAAAUUUUUAAAUUCAUAUAACUCCCCAUGCUAAGAUUUGUUACUAAAAACUCUCAAG....
X GAAAAAUUUUUAAAUUCAUAUAACUGAAUTUTAAAAAUUUUUCACAGGGUGUGUGGCGGUA.... Break Point
80 70
L-A ... .CAGCCCUUGAUAGUUCUGAUCCA
x ....CAGCCCUUGAUAGUUCUGAUCCA
60 50 40 30 20 10
L-A CUCCGGGCAUUACAGGUCAUACUGUAGUUGCCAAAAAGAUAAUGGGAAUUACCCAUAUGC
X CUCCGGGCAUUACAGGUCAUACUGUAGUUGCCAAAAAGAUAAUGGGAAUUACCCAUAUGC
[image:7.612.91.517.80.207.2]3' End
FIG. 7. Comparison of the5'-and3'-end sequences ofL-AandXpositivestrands.The L-A sequencewastaken from the datapublished byThiele et al. (13). The sequence of theX5'-endpositive strandwasdeducedfrom the 3'-end sequence of thenegativestrand.The3'-end sequenceofXwasdirectly determined. The breakpoint indicates thebeginningof different sequences forL-Aand X 5'ends. Thetwo17-bp inverted repeats in theX 5'end (between bases1to 17and26 to42)areunderlined andmarkedwitharrows.Theunderlined AUG in the 5' endof the L-A sequence is the first AUG and is thuspresumedtobe theinitiation codonof thecoatproteingene.
thebreakpointatbp 25, there is a stretch of 17bpthat is a perfect inverted repeat of the first 17 bp of the RNA
molecule (both inverted repeats areunderlined and marked witharrows).Thesecondoneendsatbp42.Wedonotknow
whether the second invertedrepeatfound in X wasalready
presentsomewhereinL-A orwhetherthistypeofstructure wasgenerated duringthe processofdeletion. Inthecaseof
the 3' ends,bothL-Aand X areidentical throughthe 110bp sequenced (Fig. 7).
DISCUSSION
We describeXdsRNA, a 0.52-kb molecule that has lost most of the internal regions of L-A and retains the ends.
Since it is encapsidated, transcribed, and replicated, X
delimits the sequences of L-A needed in cis for these processes.
As in thecaseofthe S dsRNAdeletionderivatives of M1,
theretained5'segmentofX(5' withrespect tothemessage
strand) is much shorter than the retained 3' segment.
Al-though the small number ofindependent isolates examined makes this observationstatisticallytenuous,thismayreflect
therequirement forsome 3' subterminal siteon both mole-cules.
Thesignificance of the5'invertedrepeatisnotclear. This could beanaccident ofthemechanism of thegeneration of
Xor anessential feature.
Whereas deletion mutants ofM1 dsRNA(S dsRNAs)are
oftenisolated, this isthe first described deletion mutantof
L-A. L-AdsRNA doesnotgive cells arecognizable pheno-type, so mutation ofL-Acan only be detected genetically
through its effect on M. Given sufficient time, S mutants
completely eliminate (exclude) M from all cells, and so S mutants aredetected as nonkiller mutants, whereas X is only
incompatible with
M1.
A significantportion of the progeny of a cell initially carrying X andM1
loses X and retains M1. Thus, the phenotype of an X mutant would not be asclear-cut as that of an S mutant. Moreover, S mutants can
completely exclude their parent
M1
and still survive to befound, since they depend only on L-A, not on M1. In contrast, if an X mutant completely excludes L-A, it will be
lost itself. This may set some limits on the type of L-A
deletion mutant that can be isolated and thus make X mutants rarerthan S mutants.
The findingthat X isinVLPs and that X VLPs can have
fromoneto eightX dsRNA molecules per VLP shows that an L-A replicon, like an M1 replicon, can have multiple
genomes perparticleifonly it is small. X dsRNAsynthesis
in vitro was observed in particles with various densities.
This confirms our head-full replication model
originally
postulated for M1. This model is, ofcourse, distinct from
Streisinger's originalhead-fullpackaging model,in which T4
DNA replicates outside the particle and is stuffed into the
head until the head is full (11). In the case ofX and M1
dsRNAs, one positive single strand is encapsidated in a VLP, andthisreplicatesinside theparticleuntil theparticle
is full. Once the particles are full, all of the newly
synthe-sized positive strands are extruded from theparticles. The
capacitylimit observed for Xparticles (eightperparticle)is justthe value expected (4.5/0.52 = 8.6) if one L-AdsRNA (4.5 kb) perparticle is all the RNA that theseparticles can
hold.
AlthoughX is an L-Areplicon, X surprisingly replicates likeM1. Xrequiresallofthe nine MAK genes tested thatare dispensable for L-A but
required
for M1. Inaddition,
therequirements of X for several MAK genes is apparently suppressed by ski- mutationsorby [B],asisthecaseforM1
(16).XandM1 are
incompatible,
unlikeXand L-AorM1andL-A. Thesefacts indicate thatXand M1sharemany aspects
oftheir
replication
andpackaging
mechanisms.Why doesadeletionmutantofL-Aneedmany extra MAK genes like M1? X and M1 have in common their small size andtheir need toborrowcoatproteinfrom L-A.Theirsmall
size may impose special requirements at the packaging
stage, while their need to borrow coat protein from L-A
might require some help. Clearly, an in vitro packaging
systemwould be useful inexamining these problems further.
The findingthat [B], asuppressorofmutations in oneclass ofMAKgenes, is avariantofL-A (and thuspresumably a
variant ofthe coatprotein) (Uemura and Wickner, inpress) alsopointstothepackagingprocess ortheVLPs themselves asthe site of action ofthis classofMAKgeneproducts.
Xarosein a strain carryingonly L-A-HNB (Uemura and
Wickner,inpress);thatis,Xisderivedfrom an L-Acarrying [B]. Nevertheless, in theabsence of anintact L-A-HNB, X
apparently requires the MAK genes that are bypassed by
[B]. This indicates that the [B] function has been deleted from X. X carries all of the sites for transcription,
replica-tion, and packaging. Thus, [B] is probably a property of a
protein product of L-A.
X, likeM1or
M2,
lowers L-A copynumber, especially in ski- strains. Thus, whereas both X and SKI products lower L-A, removing a SKIproduct by a ski- mutation actuallylowers L-A copy number if X is present because X copy
on November 10, 2019 by guest
http://jvi.asm.org/
number is raised. The same effect is seen with M1 (1).
Whereasski- M1 strains are cold sensitive for growth (8)or evengrowslowly at normal growthtemnperaturesif another cytoplasmic element called
[D]
is also present (Esteban andWickner,Genetics, in press), ski- X
[DI
strains donotshow this effect. The explanation of these phenomena will await further insight into the mechanism of action of SKIgene products and the details of X and M replication.LITERATURE CITED
1. Ball, S. G., C. Tirtiaux, and R. B. Wickner. 1984. Genetic control of L-A and L-(BC) dsRNA copy number in the killer systems ofSaccharomyces cerevisiae. Genetics 107:199-217. 2. Conde, J., and G. R. Fink. 1976. A mutantof Saccharomyces
cerevisiae defective for nuclearfusion. Proc. Natl. Acad. Sci. USA 73:3651-3655.
3. El-Sherbeini, M., and K. A. Bostian. 1987. Viruses infungi: infectionof yeast with the Kl and K2killer virus. Proc. Natl. Acad. Sci. USA 84:4293-4297.
4. Esteban, R., and R. B. Wickner. 1986. Three different M1 RNA-containing viruslike particle types in Saccharomyces cerevisiae: in vitro M1 double-stranded RNA synthesis. Mol. Cell. Biol. 6:1552-1561.
5. Fried, H. M., and G. R. Fink. 1978. Electron microscopic heteroduplex analysis of "killer"double-strandedRNAspecies from yeast. Proc. Natl. Acad. Sci. USA 75:4224-4228. 6. Lee,M., D. F. Pietras, M. E. Nemeroff, B. J. Corstanje, L.J.
Field, andJ.A. Bruenn. 1986. Conserved regions in defective interfering viral double-stranded RNA from a yeast virus. J. Virol. 58:402-407.
7. Mortimer,R.K.,and D.C. Hawthorne. 1975. Geneticmapping in yeast. Methods Cell Biol. 11:221-233.
8. Ridley,S.P., S. S.Sommer, and R. B. Wickner. 1984. Super-killer mutations inSaccharomyces cerevisiae suppress exclu-sion of M2double-stranded RNAby L-A-HN and confercold sensitivity in the presence of M and L-A-HN. Mol. Cell. Biol. 4:761-770.
9. Ridley,S.P.,and R. B. Wickner.1983.Defective interference in thekiller system ofSaccharomycescerevisiae.J.Virol. 45:800-812.
10. Somers,J.M. 1973. Isolation ofsuppressivemutantsfrom killer andneutral strains of Saccharomyces cerevisiae. Genetics74: 571-579.
11. Streisinger, G., J. Emrich,and M. M.Stahl.1967.Chromosome structure in phage T4. III. Terminal redundancy and length determination. Proc.Natl. Acad. Sci. USA 57:292-295. 12. Thiele,D.J., E. M. Hannig, and M. J.Leibowitz.1984.Genome
structure andexpression ofadefective interfering mutant ofthe killer virusof yeast. Virology137:20-31.
13. Thiele,D.J., E.M.Hannig,andM.J. Leibowitz. 1984.Multiple Ldouble-stranded RNA speciesofSaccharomycescerevisiae: evidence for separateencapsidation. Mol. Cell. Biol. 4:92-100. 14. Thiele,D.J., andM. J. Leibowitz. 1982. Structural and func-tional analysis of separated strands of killerdouble-stranded RNA of yeast.Nucleic Acids Res. 10:6903-6918.
15. Toh-e, A.,P. Guerry, and R. B. Wickner. 1978.Chromosomal superkillermutantsofSaccharomyces cerevisiae. J.Bacteriol. 136:1002-1007.
16. Toh-e, A.,and R. B. Wickner. 1980. "Superkiller" mutations suppress chromosomal mutations affecting double-stranded RNAkiller plasmid replication in Saccharomyces cerevisiae. Proc.Natl. Acad. Sci. USA 77:527-530.
17. Welsh,J. D., M. J. Leibowitz, and R. B. Wickner. 1980. Virion DNA-independent RNA polymerase from Saccharomyces cere-visiae. Nucleic Acids Res.$:2349-2363.
18. Wesolowski, M., and R. B. Wickner. 1984. Two new double-stranded RNA molecules showing non-Mendelian inheritance andheatinducibility in Saccharomyces cerevisiae. Mol. Cell. Biol.4:181-187.
19. Whitaker-Dowling, P., and J. S. Youngner. 1987. Viral interfer-ence-dominance of mutant viruses over wild-type virus inmjxed infections. Microbiol. Rev. 51:179-191.
20. Wickner, R. B. 1978.Twenty-sixchromosomal genes neededto maintain the killer double-stranded RNA plasmid of Saccha-romyces cerevisiae. Genetics 88:419-425.
21. Wickner,R. B.1986.Double-stranded RNAreplication in yeast: thekiller system. Annu. Rev. Biochem.55:373-395.
22. Wickner,R. B., A.Hinnebusch, A. M.Lanmbowitz, I. C. Gun-salus, and A. Hollaender (ed.). 1986. Extrachromosomal genetic elements in lowereukaryotes. Plenum Publishing Corp., New York.

![Fig. 3wasparticles (five peaks can be seen in the figure, and a sixth peak present in fraction 26 [not shown])](https://thumb-us.123doks.com/thumbv2/123dok_us/1342780.87985/4.612.63.301.77.365/fig-wasparticles-peaks-figure-sixth-present-fraction-shown.webp)