The N Terminus of Andes Virus L Protein Suppresses mRNA and
Protein Expression in Mammalian Cells
Patrick Heinemann, Jonas Schmidt-Chanasit, Stephan Günther
Department of Virology, Bernhard-Nocht-Institute for Tropical Medicine, Hamburg, Germany
Little is known about the structure and function of the 250-kDa L protein of hantaviruses, although it plays a central role in virus genome transcription and replication. When attempting to study Andes virus (ANDV) L protein in mammalian cells, we encoun-tered difficulties. Even in a strong overexpression system, ANDV L protein could not be detected by immunoblotting. Deletion analysis revealed that the 534 N-terminal amino acid residues determine the low-expression phenotype. Inhibition of translation due to RNA secondary structures around the start codon, rapid proteasomal degradation, and reduced half-life time were ex-cluded. However, ANDV L protein expression could be rescued upon mutation of the catalytic PD-E-K motif and further con-served residues of the putative endonuclease at the N terminus of the protein. In addition, wild-type ANDV L rather than ex-pressible L mutants suppressed the level of L mRNA, as well as reporter mRNAs. Wild-type L protein also reduced the synthesis of cellular proteins in the high-molecular-weight range. Using expressible ANDV L mutants as a tool for localization studies, we show that L protein colocalizes with ANDV N and NSs but not Gc protein. A fraction of L protein also colocalized with the cellu-lar processing (P) body component DCP1a. Overall, these data suggest that ANDV L protein possesses a highly active endonu-clease at the N terminus suppressing the level of its own as well as heterologous mRNAs upon recombinant expression in mam-malian cells.
A
ndes virus (ANDV) is a member of the genus Hantaviruswithin the familyBunyaviridae. The main reservoir host for ANDV is the long-tailed pygmy rice rat (Oligoryzomys longicauda-tus). Upon transmission to humans, ANDV causes a respiratory disease called hantavirus cardiopulmonary syndrome (HCPS) that is associated with a case fatality of up to 40%. ANDV is the only hantavirus that is further transmissible from human to hu-man. The virus is endemic in Argentina and Chile, but related HCPS-causing hantaviruses are found throughout the Americas (1,2).
The hantavirus genome consists of three antisense RNA seg-ments designated L (large), M (medium), and S (small) (3). The L segment encodes the 250-kDa L protein, which presumably pos-sesses several enzymatic functions involved in RNA transcription and replication (4,5). Expression of the M segment results in the glycoprotein precursor (GPC), which is cleaved posttranslation-ally into the envelope proteins Gn and Gc. The S segment encodes the nucleoprotein (N), which encapsidates the viral genome. Some hantaviruses, including ANDV, additionally encode on the S segment a nonstructural protein (NSs), which may play a role in modulating the interferon response of the infected cell (6,7).
The hantavirus L protein has been poorly characterized thus far. A 250-kDa L protein was detected in Tula hantavirus-infected cells, indicating that the majority of the protein is not processed posttranslationally (5). Homologies to polymerase proteins of other RNA viruses revealed a RNA-dependent RNA polymerase (RdRp) domain in the center of the protein (4). Hantavirus mRNAs contain heterologous nucleotides at the 5=end, suggest-ing the virus removes cap structures from cellular mRNAs to prime its own mRNA synthesis, a process called cap-snatching (8, 9). The prototype of cap-snatching viruses is influenza A virus (IAV; familyOrthomyxoviridae), which employs an RNA endonu-clease in the N terminus of PA protein for cleavage (10,11). Re-cently, endonucleases were also identified in the N termini of the L proteins of La Crosse virus (LACV; genusOrthobunyavirus, family
Bunyaviridae) and lymphocytic choriomeningitis virus (family
Arenaviridae) (12,13). An endonuclease motif is also found in the N terminus of hantavirus L protein, suggesting that it harbors an endonuclease as well (4,12,13). In addition, it is assumed that the N protein of hantaviruses is involved in cap-snatching due to its ability to bind cap structures and its association with processing (P) bodies (14,15). P bodies play a role in cellular mRNA turnover (16,17), and there is evidence that N protein binds to and protects 5=-terminal nucleotides of capped mRNA from degradation in P bodies (14). These capped RNA fragments may be transferred to L protein, further trimmed in length by the endonuclease and used by the RdRp to initiate viral mRNA synthesis.
Here, we studied the recombinant expression and subcellular localization of ANDV L protein in mammalian cells. Like other researchers (18), we encountered difficulties in expressing hanta-virus L protein in these cells. Several mechanisms possibly under-lying the low-expression phenotype of L protein were experimen-tally approached. By mutagenesis, we were able to overcome the low-expression phenotype of L protein and map regions and res-idues determining it. ANDV L mutants showing a high-expression phenotype were used as a tool to study the localization of the protein in the cell.
MATERIALS AND METHODS
Construction of plasmids for protein expression.Vero cells in 75-cm2
tissue culture flasks were inoculated with ANDV strain Chile-9717869
Received5 January 2013 Accepted4 April 2013
Published ahead of print10 April 2013
Address correspondence to Stephan Günther, guenther@bni.uni-hamburg.de.
J.S.-C. and S.G. contributed equally to this article.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.00043-13
on November 7, 2019 by guest
http://jvi.asm.org/
(kindly provided by Heinz Feldmann, Winnipeg, Canada), Sin Nombre virus (SNV) strain Convict Creek 107, Hantaan virus (HTNV) strain 76-118, or Puumala virus (PUUV) strain Sotkamo. After 8 to 10 days, the supernatant was cleared by low-speed centrifugation, and virus was pel-leted from the cleared material by ultracentrifugation. The pellet was re-suspended in water, and virus RNA was purified by using TRIzol LS Re-agent (Invitrogen). Purified RNA was reverse transcribed by using Superscript III reverse transcriptase (Invitrogen) with virus-specific primers, and the cDNA was amplified with the Phusion High-Fidelity DNA polymerase (New England BioLabs). ANDV genes were cloned into expression vector pCITE-2a (Novagen) containing a T7 RNA polymerase promoter, an internal ribosomal entry site (IRES), and a favorable Kozak sequence (pCITE-ANDV-L, pCITE-ANDV-N, pCITE-ANDV-GPC, and pCITE-ANDV-NSs). For immunoblotting and immunoprecipitation studies, hemagglutinin (HA), or FLAG tags were fused to the C termini of the genes. ANDV, HTNV, PUUV, and SNV L gene fragments (amino acid residues 1 to 534) were cloned into pCITE-2a and fused to a C-terminal HA tag (pCITE-LN constructs). The sequence of cloned genes
corre-sponded to published sequences (GenBank accession no. for ANDV L, AF291704; ANDV GPC, AF291703; ANDV N, AF291702; HTNV L, X55901; PUUV L, Z66548; and SNV L, AF425256) except the following differences which have been confirmed by direct sequencing of our virus isolates (numbering corresponds to nucleotide and amino acid position in the gene): ANDV L, G541A leading to amino acid exchange V181I; ANDV GPC, C2580T silent mutation and G3405T leading to K1135N; HTNV L, C47A leading to P16H; and SNV L, C124A leading to L42M and G6343A leading to V2115I. The construct for expression of Lassa virus L protein (pCITE-Las-L) has been described previously (19).
L protein mutants were generated via a classical two-step PCR mu-tagenesis approach as described previously (20) or QuikChange mutagen-esis (Stratagene) using pCITE-L or LNas a template. The resulting
frag-ments were cloned or used for transfection without prior cloning as described previously (20). Codon-modified L gene fragments (nucleotide positions 1 to 135) were chemically synthesized (GeneArt) and exchanged with the corresponding wild-type fragment in pCITE-ANDV-LN. The
presence of a fusion, deletion, or artificial point mutation was ascertained by sequencing.
Cells and transfections.BSR-T7/5 cells stably expressing T7 RNA polymerase (21) were grown in Glasgow’s minimal essential medium (Gibco) supplemented with 5% fetal calf serum (FCS). Every second pas-sage, 1 mg of Geneticin (Gibco) per ml of medium was added to the cells. BHK-21 cells were grown in Dulbecco modified Eagle medium (DMEM; PAA Laboratories) supplemented with 10% FCS. Cells were seeded at a density of 1⫻105or 5⫻105cells per well of a 24- or 6-well plate,
respectively, at 1 day prior to transfection. Cells were inoculated with modified vaccinia virus Ankara expressing T7 RNA polymerase (MVA-T7) (22) for 1 h before transfection. All transfections were performed with 3l of Lipofectamine 2000 (Invitrogen) perg of DNA in cell culture medium without supplements. Medium was replaced at 4 h after trans-fection by fresh medium complemented with FCS.
Immunoblot analysis.BSR-T7/5 or BHK-21 cells in a well of a 24-well plate were infected with MVA-T7 and transfected with 250 to 500 ng of expression construct (plasmids or PCR products). One day after transfec-tion, cells were lysed in 100l of 1⫻NuPAGE LDS sample buffer (Invit-rogen) complemented with dithiothreitol (DTT) and then heated to 95°C for 5 min. Proteins were separated by sodium dodecyl sulfate polyacryl-amide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membrane (Carl Roth). Membranes were blocked in 1⫻Roti-Block (Carl Roth) for 1 h and incubated with rabbit anti-HA (1:10,000; H6908; Sigma-Aldrich) for 1 h and peroxidase-conjugated secondary antibody (1: 50,000; 111-035-003; Jackson ImmunoResearch) in phosphate-buffered saline (PBS)– 0.2⫻Roti-Block at room temperature for 1 h. After washing, protein bands were visualized by chemiluminescence using SuperSignal West Pico substrate (Pierce) and X-ray film (Kodak) or the Fusion SL image acqui-sition system (Vilber Lourmat).
Confocal immunofluorescence microscopy. BHK-21 cells were grown on coverslips in 24-well plates, infected with MVA-T7, and trans-fected with 50 to 100 ng of plasmid DNA per well. Cells were washed with PBS at 10 h after transfection and fixed with 4% paraformaldehyde– 4% sucrose in PBS for 15 min. Cells were permeabilized with 0.2% saponin in PBS for 10 min, washed with PBS, blocked with 1% bovine serum albumin (BSA) in PBS at 37°C for 30 min, and incubated with mouse monoclonal anti-HA F-7 (1:100; sc-7392; Santa Cruz), rabbit anti-FLAG (1:2,000; A190-102A; Bethyl), mouse monoclonal anti-FLAG M2 (1:100; F1804; Sigma-Aldrich), rabbit anti-HA (1:250; H6908; Sigma-Aldrich), mouse monoclonal DCP1a (1:100; ab57654; Abcam), and/or rabbit anti-ANDV-N serum (1:500) in 0.2% BSA– 0.2% saponin in PBS at 37°C for 1 h. After the cells were washed, they were incubated with 4= ,6-diamidino-2-phenylindole (DAPI; 1:10,000; Boehringer Ingelheim), anti-mouse IgG coupled to rhodamine (1:100; 115-025-072; Jackson ImmunoResearch), and anti-rabbit IgG coupled to fluorescein isothiocyanate (1:250; PA1104; SIFIN) in 0.2% BSA– 0.2% saponin in PBS at 37°C for 1 h. Cells were washed, air dried, and covered with ProLong Gold antifade reagent (In-vitrogen). Confocal images were captured at⫻100 magnification using a FluoView 1000 microscope (Olympus) and processed with the ImageJ program (http://rsb.info.nih.gov/ij/).
Quantification of mRNA.BHK-21 cells in a well of a 24-well plate were infected with MVA-T7 and transfected with 250 ng of pCITE-ANDV-L, 10 ng of pCITE-FF expressing firefly luciferase, and 10 ng of pCMV-RenLuc expressingRenillaluciferase. Total RNA was purified with the RNeasy kit including DNase I treatment (Qiagen) and quantified spec-trophotometrically. ANDV L mRNA, firefly luciferase mRNA, andRenilla
luciferase mRNA were quantified by real-time reverse transcription-PCR (RT-PCR) on the LightCycler 480 II system (Roche) using 3l of purified RNA, the QuantiTect SYBR green RT-PCR Kit (Qiagen), and primer combinations ANDV-L-fwd/ANDV-L-rev (CTTGCACCTGGAACGG TATC and TGTCCAGAAGGGCTATTTGG), Firefly-fwd/Firefly-rev (A GATCGTGGATTACGACGCCAG and TGCGTCGAGTTTTCCGGTA AG), and Renilla-fwd/Renilla-rev (GACAAGATCAAGGCCATCGTCC and GCACCATTTTCTCGCCCTCTTC), respectively. The specificity of the reaction was verified by melting-curve analysis. Relative mRNA levels were calculated using standard curves and normalized with the total RNA amount added to the RT-PCR assay.
Proteasome inhibition.Transfected BHK-21 cells were treated at 4 h posttransfection with 0 to 100M of the proteasome inhibitor carbo-benzoxy-L-leucyl-L-leucyl-L-leucinal (MG-132; Merck Millipore) diluted in GMEM–10% FCS. One day posttransfection, cells in a well of a 24-well plate were lysed with 100l of passive lysis buffer (Promega), and nuclei were pelleted by centrifugation. Proteins in cytoplasmic lysate were sepa-rated by SDS-PAGE and detected by immunoblot. Proteasome inhibition was monitored in parallel in a 20S proteasome activity assay: 10l of lysate was incubated with 100l of substrate solution containing 100M
N-succinyl-Leu-Leu-Val-Tyr-7-amino-4-methylcoumarin (Suc-LLVY-AMC) for 1 h at 37°C in black microtiter plates. The proteolytic activity was measured by release of the fluorogenic AMC group (excitation 360 nm, emission 460 nm) with a Synergy Reader (BioTek).
Pulse-chase labeling and immunoprecipitation.BHK-21 cells in a well of a six-well plate were infected with MVA-T7 and transfected with 2.5g of pCITE-ANDV-LN. Cells were first incubated in regular DMEM for 4 h, then in DMEM lacking methionine and cysteine for 1 h, and finally in DMEM containing [35S]methionine and [35S]cysteine (Hartmann
An-alytic) for 2 h (pulse). The radioactive medium was replaced with regular DMEM, and cells were incubated for an additional 1 and 2 h, respectively (chase). Pulsed and pulse-chased cells were lysed in 500l of lysis buffer (50 mM Tris-HCl [pH 7.5], 200 mM NaCl, 1 mM EDTA, 0.5% Triton X-100, 40l of complete protease inhibitor cocktail/ml [Roche]). Nuclei were pelleted by centrifugation; 450l of supernatant was mixed with 1l of anti-HA antibody solution (0.5 to 0.7 mg/ml; H6908; Sigma-Aldrich), and the mixture was incubated at 4°C overnight with gentle agitation. Protein A-Sepharose 4 Fast Flow solution (40l, as supplied by GE
on November 7, 2019 by guest
http://jvi.asm.org/
Healthcare) was added, and the mixture was incubated at 4°C with gentle agitation for 2 h. Sepharose-coupled antibody-protein complexes were precipitated by centrifugation and washed twice with lysis buffer and twice with 50 mM Tris-HCl (pH 7.5). Precipitated proteins or lysate were mixed with 1⫻NuPAGE LDS sample buffer (Invitrogen) and DTT, heated at 95°C for 5 min, and separated by SDS-PAGE. Radioactive signals were detected with a FLA-7000 phosphorimager (Fuji) and quantified using Multi Gauge software (Fuji). Half-life time was calculated from the signal intensities by fitting an exponential curve to the data using Excel software (Microsoft).
RESULTS
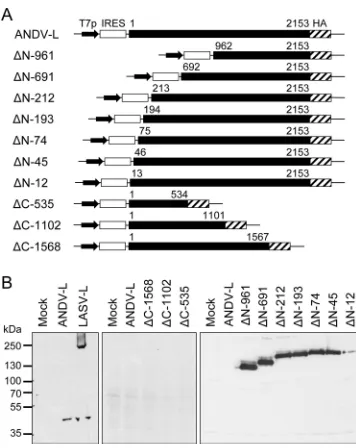
Cloning and expression of ANDV L protein.Virus RNA was iso-lated from ANDV-infected cells, converted to cDNA, and ampli-fied by PCR using various combinations of L gene-specific prim-ers. A complete L gene was assembled from several fragments and cloned into an expression vector under the control of T7 RNA polymerase promoter and IRES (pCITE-ANDV-L). A C-terminal HA tag was fused to the L protein to facilitate detection in immu-noblots. BSR-T7/5 cells, which constitutively express T7 RNA polymerase, were transfected with pCITE-ANDV-L. To maximize protein expression, the cells were additionally infected with re-combinant vaccinia virus expressing T7 RNA polymerase (MVA-T7) before transfection. However, despite several attempts, ANDV L protein could not be detected in immunoblot, while arenavirus L protein (Lassa virus) expressed in parallel was readily detected (Fig. 1B, left). Therefore, we assumed that ANDV L pro-tein might be poorly translated or instable due to specific sequence elements. To identify these elements, ANDV L was truncated at N and C terminus (Fig. 1A). C-terminally truncated ANDV L
pro-tein showed a wild-type phenotype, i.e., it was not detectable (Fig. 1B, middle). In contrast, ANDV L protein truncated at the N terminus was clearly detectable (Fig. 1B, right). Removal of the first 12 amino acid residues already led to a measurable increase in expression level, although deletion of 45 or more N-terminal res-idues resulted in strong signals in immunoblot as observed for the arenavirus L protein. In conclusion, ANDV L protein is poorly expressed after transfection, which appears to be linked to regula-tory RNA or protein elements residing in the N-terminal 135 nu-cleotides or 45 amino acid residues, respectively.
Potential sequence elements interfering with ANDV L pro-tein translation.To facilitate mutagenesis and expression analy-sis, further experiments were conducted using a C-terminally truncated version of ANDV L protein (ANDV LN, 55 kDa, amino
acid residues 1 to 534 plus HA tag), which shares the low-expres-sion phenotype with full-length L protein. In the context of ANDV LN, we first tested whether the 5=end of the L gene contains
sequence elements, codons, or RNA secondary structures which might reduce translation efficacy (23,24). To this end, the nucle-otide sequence of the L gene (positions 1 to 135) was modified without changing the amino acid sequence. Two different L genes were constructed in which 29 and 35%, respectively, of the 5= -terminal nucleotides were exchanged; one was codon optimized for expression inHomo sapienscells, and the other was randomly codon modified (Fig. 2A). Potential sequence or RNA secondary structure elements should have been inactivated or at least af-fected by the high number of nucleotide exchanges. However, no FIG 1Expression of L protein in mammalian cells. (A) Schematic
represen-tation of constructs for expression of ANDV L proteins with N- and C-termi-nal deletions. The constructs contain a T7 RNA polymerase promoter (T7p) and IRES to facilitate cap-independent translation. L gene amino acid posi-tions are indicated with the constructs. (B) Immunoblot analysis of wild-type L protein of ANDV (ANDV-L), wild-type L protein of Lassa virus (LASV-L), and deletion mutants of ANDV L protein. BSR-T7/5 cells were infected with MVA-T7 and transfected with expression plasmids or empty vector as a neg-ative control (Mock). L proteins were separated by SDS-PAGE, blotted, and detected with anti-HA antibody via the HA tag.
FIG 2Influence of multiple nucleotide exchanges in the 5=end of L gene on expression of ANDV L protein. (A) The L gene sequence (nucleotide positions 1 to 135) in the context of ANDV LNwas modified without changing the amino acid sequence. In one construct, codons were optimized forHomo sapienscells (Mut-1, 29% mutations) and in the other construct, each codon was randomly modified (Mut-2, 35% mutations). (B) BSR-T7/5 cells were infected with MVA-T7 and transfected with expression plasmids or empty vector as a neg-ative control (Mock). The expressible L mutant⌬N-45 lacking nucleotides 1 to 135 served as a positive control. L proteins were separated by SDS-PAGE, blotted, and detected with anti-HA antibody via the HA tag.
Andes Virus L Protein
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.73.251.64.288.2] [image:3.585.318.526.66.307.2]change in protein level was observed when ANDV L was expressed from these constructs (Fig. 2B). Thus, RNA elements interfering with translation are unlikely to account for the low expression level.
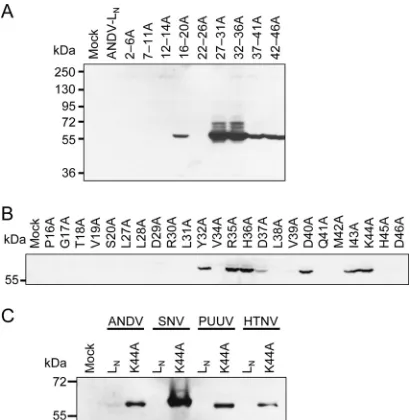
Mapping of amino acid residues associated with downregu-lation of protein expression.To map amino acid residues that might be involved in the low-expression phenotype, clusters of 3 to 5 amino acids were replaced by alanine between positions 2 and 46 of ANDV LN. Five of the nine alanine cluster mutants were
detectable in immunoblot (Fig. 3A), suggesting that amino acid residues within these clusters are involved in the low expression. To narrow down the sequences, single amino acid residues were exchanged within these clusters. Seven mutants were identified by this approach (Y32A, R35A, H36A, D37A, D40A, I43A, and K44A) showing detectable protein expression (Fig. 3B). However, no change in phenotype was observed when introducing single amino acid substitutions between residues 16 to 20 and residues 27 to 31, although the corresponding alanine cluster mutants were expressed at high levels. Probably, changes in the local protein structure inducible only by substituting two or more residues are needed to change the phenotype here. Taken together, individual amino acid residues as well as structural features in the N terminus appear to be involved in the low-expression phenotype of ANDV L protein.
Expression analysis of Sin Nombre (SNV), Puumala (PUUV), and Hantaan virus (HTNV) L proteins.The residues Y32, R35, H36, D37, D40, I43, and K44 associated with the low expression of ANDV L protein are highly conserved within the hantavirus genus (4). To investigate whether the expression
phe-notype of SNV, PUUV, and HTNV L proteins is similar to that of ANDV L protein, the N-terminal residues 1 to 534 were cloned into pCITE-2a. The partial genes corresponding in length to ANDV LNwere fused to a HA tag. In addition, constructs for
expression of SNV, PUUV, and HTNV LNproteins with the K44A
mutation were generated. Cells were infected with MVA-T7 and transfected with the L protein constructs. None of the wild-type L proteins of SNV, PUUV, and HTNV was detectable in immuno-blots, whereas all K44A mutants produced clearly visible signals (Fig. 3C). Thus, the L proteins of ANDV, SNV, PUUV, and HTNV share the low-expression phenotype and expression may be res-cued by exchanging the conserved residue K44.
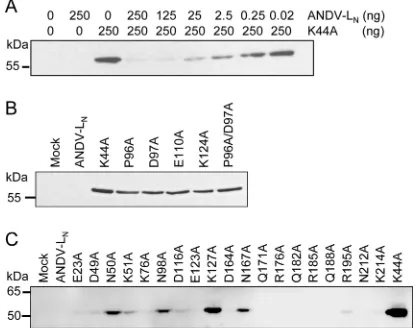
Proteasomal degradation and half-life time of ANDV L pro-tein.The experimental data obtained thus far may be consistent with a low half-life time of ANDV L protein, for example, due to rapid proteasomal degradation. Other hantavirus proteins have been shown to be subject of enhanced proteasomal degradation (25). Proteins are usually targeted for degradation by conjugation with ubiquitin molecules at lysine residues (26). Since K44 is highly conserved among all hantavirus L proteins and alanine sub-stitution enhances the protein level, we hypothesized that this res-idue might be a target for ubiquitinylation. If this is true, replace-ment of K44 by any other side chain should enhance the protein level. Indeed, all K44 mutants generated and tested (K44A, K44F, K44P, K44S, K44Q, K44E, and K44R) were expressed, suggesting that low expression depends on a lysine residue at position 44 (Fig. 4A). Two sets of experiments were conducted to test the hypothesis of enhanced degradation. First, the proteasome activ-ity was inhibited with MG-132. The inhibitor was added at differ-ent concdiffer-entrations (0 to 100M) to cells transfected with con-structs for expression of wild-type ANDV LNor the K44A mutant.
No increase in L protein level was observed (Fig. 4B), although the proteasome activity was nearly completely inhibited at 100M MG-132 (Fig. 4C).
Second, the half-life time of both proteins was determined in a pulse-chase experiment. Cells were transfected with constructs for expression of wild-type ANDV LNor the K44A mutant and
met-abolically pulse-labeled with [35S]methionine and [35S]cysteine. The35S-labeled ANDV L
Nproteins were immunoprecipitated at
0, 1, and 2 h after the pulse and separated by PAGE, and their levels were measured by autoradiography. Three independent experi-ments were performed. Weak and strong signals for wild-type and mutant proteins, respectively, were visible after radioactive label-ing (Fig. 4D), facilitatlabel-ing calculation of the half-life times for both proteins. However, no difference in half-life time between wild-type LNprotein and the K44A mutant were observed; it is⬃1 h for
both (Fig. 4E). In conclusion, inhibition of proteasomal degrada-tion and half-life time measurement indicates that the low steady-state level of wild-type ANDV L protein is not due to enhanced protein degradation or instability.
Influence of mutations in the putative endonuclease of ANDV L protein on the expression level.Having largely excluded RNA sequence elements repressing translation or enhanced pro-tein degradation, we assumed the low-expression phenotype is mediated at posttranscriptional level. However, the existence of RNA-destabilizing elements within the 5=end of L gene is hardly compatible with the observation that substitution of individual amino acid residues (Fig. 3) rather than multiple synonymous nucleotide changes (Fig. 2) can rescue ANDV L protein expres-sion. A solution is to assume that ANDV L protein is both target FIG 3Mapping of amino acid residues within the N terminus of L protein
mediating the low-expression phenotype. BSR-T7/5 cells were infected with MVA-T7 and transfected with expression plasmids or empty vector as a neg-ative control (Mock). L proteins were separated by SDS-PAGE, blotted, and detected with anti-HA antibody via the HA tag. (A) Three to five amino acid residues in a row were replaced by alanine in the context of ANDV LN. The position of the mutated stretch is indicated above the immunoblot (first–last residue). (B) Individual amino acid residues were substituted by alanine in the stretches from 16 to 20 and from 27 to 46 in the context of ANDV LN. (C) Expression of SNV, PUUV, and HTNV LNproteins (wild-type and K44A mutant), corresponding in length to ANDV LN, was analyzed in immunoblots.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.61.266.66.276.2]and effector. To test this, decreasing amounts of wild-type ANDV LNconstruct were cotransfected with a constant amount of
ex-pression construct for the K44A mutant. Cotransfection of wild-type LNconstruct clearly affected the expression of the K44A
mu-tant; even when both constructs were transfected in a 1:100 ratio, the level of the K44A mutant was significantly reduced (Fig. 5A). This dominant-negativetranseffect suggests that ANDV L protein plays an active role in downregulating protein expression and is not just a target. The activity would have to be located in the N terminus of L protein.
Recently, structural and functional studies revealed an endo-nuclease of the PD-(D/E)-K superfamily at the N terminus of L proteins of arena- and orthobunyaviruses (12, 13,27). Primary and secondary structure alignment suggests that the N terminus of hantavirus L protein also harbors an endonuclease. The PD-(D/ E)-K motif of hantavirus endonuclease probably corresponds to the conserved residues P96, D97, E110, and K124 (see the align-ment inFig. 11). These residues were exchanged to alanine in the background of the ANDV LNconstruct and expression of the
mu-tants was tested by transfection. All catalytic site mumu-tants were clearly expressed as tested in immunoblot (Fig. 5B). To identify further residues important for the low-expression phenotype, 19 conserved E, D, Q, N, K, and R residues within the putative endo-nuclease domain (residues 1 to 220) were replaced by alanine. Four mutants (N50A, N98A, K127A, and N167A) were found to be expressed, although to a lower level than K44A (Fig. 5C). In conclusion, ANDV L protein expression was enhanced upon mu-tation of the PD-E-K motif and further conserved residues within the putative endonuclease, suggesting a role for this enzymatic activity in downregulating L protein expression.
Influence of ANDV L protein on the mRNA level.Since the putative endonuclease is supposed to act on mRNA level
(consis-tent with the finding that the low L protein level is not due to increased protein degradation), the influence of various muta-tions in this domain on the L mRNA level was studied. In addition, the influence of L protein on the level of heterologous mRNAs was measured, since the dominant-negativetranseffect suggests that FIG 4Testing the stability of L protein. (A) Lysine on position 44, which might be a target for ubiquitinylation and a mediator of proteasomal degradation, was substituted by residues with related or different chemical properties. BHK-21 cells were infected with MVA-T7 and transfected with expression plasmids or empty vector as a negative control (Mock). L proteins were separated by SDS-PAGE, blotted, and detected with anti-HA antibody via the HA tag. (B) Cells were transfected with wild-type LN(ANDV-LN) or the K44A mutant and treated with the proteasome inhibitor MG-132. L protein in cytoplasmic lysate was detected in immunoblot via the HA tag. (C) Control experiment demonstrating inhibition of the proteasome in the above experiment. Proteasome activity in cytoplasmic lysate was measured using a fluorogenic substrate. (D) Pulse-chase labeling of ANDV LNand K44A mutant. BHK-21 cells were transfected with the correspond-ing constructs, metabolically labeled with [35S]methionine and [35S]cysteine, and harvested 0, 1, and 2 h after the labeling. L proteins were immunoprecipitated via the HA tag, separated by SDS-PAGE, and detected by autoradiography. (E) The half-life time of ANDV LNand K44A mutant was calculated from autoradiography data of three independent pulse-chase experiments.
FIG 5Relevance of the putative endonuclease in the N terminus of L protein for the low-expression phenotype. BSR-T7/5 cells were infected with MVA-T7 and transfected with expression plasmids or empty vector as a negative control (Mock). L proteins were separated by SDS-PAGE, blotted, and detected with anti-HA antibody via the HA tag. (A) The expressible LNK44Amutant was coexpressed with the nonexpressible wild-type ANDV LN. Different ratios of the corresponding expression plasmids were transfected. (B) Expression anal-ysis of ANDV LNmutants with mutations of the putative catalytic site of the endonuclease. (C) Expression analysis of ANDV LNmutants with substitu-tions of additional conserved and charged residues in the endonuclease do-main.
Andes Virus L Protein
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.112.472.65.260.2] [image:5.585.317.525.456.620.2]any mRNA might be a target. To this end, two different reporter constructs were cotransfected with the L protein expression con-structs: (i) a construct expressing firefly luciferase (FF) under the control of a T7 RNA polymerase promoter and IRES (pCITE-FF) and (ii) a construct for expression ofRenillaluciferase (RenLuc) under the control of a polymerase II promoter (pCMV-RenLuc). To verify that the use of the truncated version of L protein (LN) in
the above experiments did not lead to artificial results, selected mutations that enhanced the L protein level were introduced into the full-length L protein (mutants Y32V, R35H, H36R, D37A, D40E, I43A, K44A, N50A, P96A, D97E, N98A, E110A, K124A, K127A, and N167A; the more conservative substitutions at posi-tions 32, 35, 36, 40, and 97 enhanced expression in the context of ANDV LN[data not shown] just as the alanine substitutions had
done [Fig. 3Band5B]). Cells were infected with MVA-T7 and transfected with L protein expression constructs, as well as with pCITE-FF and pCMV-RenLuc. The L protein level was analyzed in immunoblots, whereas the levels of L mRNA, FF mRNA, and RenLuc mRNA were measured by quantitative real-time RT-PCR. The expression data obtained with the full-length L proteins confirmed the results with the truncated version of L protein. All mutants were expressed, although at different levels like the anal-ogous LNmutants (Fig. 6A). Corresponding with the protein data,
the L mRNA levels of the mutants were higher than the mRNA level of wild-type L protein (Fig. 6B, upper diagram). Further-more, there was a rough correlation between L protein and L mRNA levels. Consistent with the dominant-negativetranseffect, the L protein level also correlated with the FF mRNA and RenLuc mRNA levels (Fig. 6B, middle and lower diagram). The higher the expression level of a specific mutant, the higher the level of the heterologous mRNAs. These experiments are consistent with the hypothesis that wild-type ANDV L protein downregu-lates the level of its own mRNA, as well as the level of heterol-ogous mRNAs. The mutational analysis strongly suggests that this effect is mediated by an endonuclease activity in the N terminus of the L protein.
Influence of ANDV L protein on cellular protein level.Since L protein reduces the level of mRNAs expressed from reporter con-structs, we assumed that it also reduces the cellular mRNA and thus protein levels. Cells were transfected with constructs for ex-pression of wild-type ANDV LNor the K44A mutant and
meta-bolically pulse labeled with [35S]methionine and [35S]cysteine. The35S-labeled cytoplasmic proteins were extracted at 0, 1, and 2
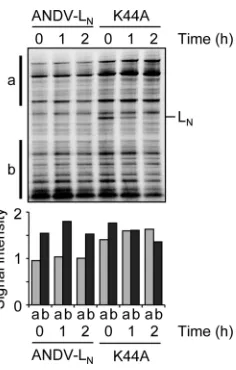
h after the pulse, corresponding to 7, 8, and 9 h, posttransfection, respectively, separated by SDS-PAGE, and their level was mea-sured by autoradiography. At all time points, the steady-state lev-els of high-molecular-weight proteins was ca. 50% higher in cells expressing the K44A mutant than in cells expressing ANDV LN
(Fig. 7). No difference was seen in the low-molecular-weight range. The signal intensities in the high-molecular-weight range did not decrease over time, indicating that the reduced steady-state level in ANDV LN-expressing cells was not due to reduced
half-life time of the proteins. We conclude from these data that the expression of ANDV LNreduces the cellular protein level
specifi-cally in the high-molecular-weight range and that this effect is not mediated by enhanced protein degradation.
Localization studies with ANDV L, N, Gc, and NSs proteins.
The availability of L protein mutants with single amino acid ex-changes in the N terminus, which can be readily expressed and visualized in mammalian cells, enabled us to study the subcellular
localization of L protein in the context of other virus and cellular proteins. Cells were infected with MVA-T7 and transfected with various combinations of expression plasmids for ANDV L, N, GPC, and NSs proteins containing HA or FLAG tags. The local-ization of the proteins was examined by confocal immunofluores-cence microscopy using antibodies directed against the tag se-quences.
The FLAG-tagged full-length K44A L mutant (L-K44A) was coexpressed with HA-tagged ANDV N and GPC protein, respec-tively. As the latter is cleaved posttranslationally into Gn and Gc, only Gc carrying the C-terminal HA tag is detected. L-K44A and N proteins are distributed diffusely and in granular and punctate FIG 6Expression analysis of full-length L protein mutants and influence of mutations on mRNA levels. Mutations, which enhanced the expression level in the context of ANDV LN, were tested in the context of full-length L protein. BHK-21 cells were infected with MVA-T7 and transfected with L protein ex-pression constructs, pCITE-FF, and pCMV-RenLuc. Three wells were trans-fected in parallel. (A) Cells in one well were lysed, L proteins were separated by SDS-PAGE, blotted, and detected with anti-HA antibody via the HA tag. (B) RNA was extracted from cells in the second and third well and the levels of L mRNA, FF mRNA, and RenLuc mRNA were measured by quantitative real-time RT-PCR independently for each well. The relative mRNA levels are shown with or without normalization with the total RNA amount added to the RT-PCR assay (dark and light gray bars, respectively). Mean and range for the two measurements are shown. The corresponding level of the L protein mu-tants is shown above the diagrams. The immunoblot bands from panel A were rearranged according to signal intensity.
on November 7, 2019 by guest
http://jvi.asm.org/
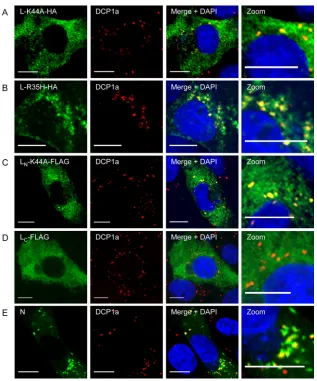
[image:6.585.331.510.66.426.2]structures in the cytoplasm (Fig. 8A). Both proteins colocalize to a large extent (Fig. 8B). No colocalization with Gc protein was ob-served (Fig. 8C).
The HA-tagged NSs protein preferentially localizes in the nu-cleus when expressed alone (Fig. 9A). In contrast, coexpression with L-K44A relocalizes NSs to the cytoplasm and a high level of colocalization with L protein was observed (Fig. 9B). Cytoplasmic colocalization of NSs protein was also seen with N-terminal (res-idues 1 to 534) and C-terminal (res(res-idues 962 to 2153) fragments of L protein, although the relocalization of NSs to the cytoplasm was more pronounced when coexpressed with the C-terminal L frag-ment (Fig. 9CandD). No colocalization was observed between NSs and N protein; they mainly localized in nucleus and cyto-plasm, respectively (Fig. 9E).
It has previously been demonstrated that hantavirus N protein colocalizes with P bodies in the cytoplasm of infected cells (14). P bodies are compartments for storage and degradation of cellular mRNAs. To visualize P bodies, a monoclonal antibody against the P body component DCP1a (decapping enzyme) was used (16,17). A potential colocalization of L protein with DCP1a was studied using two different mutants, L-K44A and L-R35H. Both showed the typical granular to diffuse cytoplasmic distribution, whereas DCP1a was localized to a few discrete cytoplasmic dots (Fig. 10A andB). Although most of the L protein does not colocalize with the DCP1a dots, some of the dense granular L protein structures overlap with these dots. This moderate colocalization could be reproduced with the N-terminal (Fig. 10C), but not C-terminal fragment of L protein (Fig. 10D), suggesting that the N terminus containing the putative endonuclease determines the colocaliza-tion with DCP1a. The negative result obtained with the C termi-nus also provides evidence for the specificity of the L protein-DCP1a colocalization. In agreement with previous data (14), a fraction of ANDV-N protein colocalizes with DCP1a (Fig. 10E). Taken together, ANDV L protein colocalizes with N but not Gc
protein and relocates NSs protein from the nucleus to the cyto-plasm where it colocalizes with NSs. In addition, a certain fraction of L protein colocalizes with the P body component DCP1a.
DISCUSSION
In this study, we demonstrate that expression of recombinant ANDV L protein in mammalian cells is severely impaired and that this phenotype is determined by the 534 N-terminal amino acid residues. Evidence is presented that the low expression level is not due to translational inhibition, codon usage, rapid proteasomal degradation, or reduced half-life time. High-level expression can be rescued by mutation of catalytic site and other conserved resi-dues of the putative endonuclease in the N terminus of ANDV L protein. These mutations also increased the levels of L mRNA, as well as coexpressed heterologous mRNAs. Overall, these data are consistent with the hypothesis that wild-type ANDV L protein possesses an endonuclease activity, which downregulates the level of its own and other mRNAs. Using expressible ANDV L mutants as a tool for localization studies, we show that L protein colocalizes with N and NSs protein, and at least partially with the P body component DCP1a.
Although our study has been mainly conducted with ANDV L protein, the experiments with SNV, PUUV, and HTNV suggest that hantavirus L proteins are generally difficult to express. Three studies have investigated recombinant L protein expression thus far (5,18,28). In agreement with our findings, either no L protein could be detected, or the immunoblot and fluorescence signals were much less intense than the signals obtained here with L pro-tein mutants. The inability to express sufficient quantities of han-tavirus L protein and the negative effect on coexpressed proteins also have consequences for the development of hantavirus repli-con and reverse genetics systems. Compared to the achievements with orthobunyaviruses (29), phleboviruses (30), nairoviruses (31), arenaviruses (32), and orthomyxoviruses (33), only minor progress has been made with the hantaviruses. Replicon sys-FIG 7Influence of L protein on the cellular protein level. BHK-21 cells were
infected with MVA-T7, transfected with constructs for expression of wild-type LN(ANDV-LN) or the K44A mutant, metabolically labeled with [
35 S]methio-nine and [35S]cysteine, and harvested 0, 1, and 2 h after the labeling. Cytoplas-mic lysate was separated by SDS-PAGE, and proteins were detected by auto-radiography (top). The intensities of autoauto-radiography signals in regions “a” and “b” indicated to the left of the blot were measured and are shown in the diagram (bottom).
FIG 8Intracellular localization of ANDV L, N, and Gc protein. MVA-T7-infected BHK-21 cells were transfected with constructs for expression of L-K44A-FLAG, N, and/or GPC-HA. The FLAG tag was detected with mouse monoclonal anti-FLAG M2, the HA tag with rabbit anti-HA, and N protein with rabbit anti-ANDV-N serum. Nuclei were stained with DAPI. Images were captured by confocal microscopy. Scale bars, 10m. (A) Localization of N, Gc, and L-K44A expressed alone. (B) Localization of coexpressed N and L-K44A. (C) Localization of coexpressed Gc and L-K44A.
Andes Virus L Protein
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.107.225.63.248.2] [image:7.585.300.541.64.240.2]tems—two for HTNV and one for ANDV— have been described (18,28,34); a reverse genetics system is still not published. At least one group reported problems with reliability and reproducibility of minigenome rescue despite thorough efforts to optimize the system (18). These authors could not at all or only at very low level detect L protein expression and assumed this was a major ob-stacle to successful establishment of a reliable replicon system as well as rescue of recombinant hantavirus. Our findings may help designing experimental strategies to overcome this meth-odological hurdle.
X-ray crystallographic and functional studies revealed the presence of an endonuclease in the N terminus of arena- and orthobunyavirus L proteins (12,13,27), as well as the N terminus of IAV PA protein (11). The N terminus of hantavirus L protein shows significant homology to these endonucleases (Fig. 11) and host cell mRNA-derived nucleotides are found at the 5=end of hantavirus mRNA (8, 9). Both suggest that the N terminus of hantavirus L protein also harbors a cap-snatching endonuclease.
Here, we provide further evidence for this hypothesis. Although we cannot directly prove that downregulation of L protein expres-sion is mediated by the endonuclease activity, the observations leave little room for alternative explanations. Several other possi-ble mechanisms have largely been excluded. The key finding sup-porting the existence of an endonuclease activity is the increase in steady-state levels of L protein and various mRNAs upon muta-tion of the predicted catalytic residues of the endonuclease, that is, D97, E110, and K124.
Accepting the hypothesis that the endonuclease activity of L protein underlies our observations, one may infer some features of this enzyme. First, in addition to the catalytic site residues, the mutational analysis identified further residues that seem to be essential for enzymatic activity: Y32, R35, H36, D37, D40, I43, K44, N50, P96, N98, K127, and N167 (Fig. 11). Several of them are located in a conserved region between amino acid position 30 and 45, which is predicted to form an alpha helix corresponding to helix 2 of LACV and helix 3 of IAV. Residue H36 in the center of FIG 9Intracellular localization of ANDV L, NSs, and N protein. MVA-T7-infected BHK-21 cells were transfected with constructs for expression of L-K44A-FLAG, LN-K44A-FLAG (residues 1 to 534), LC-FLAG (residues 962 to 2153), NSs-HA, and/or N protein. The FLAG-tag was detected with mouse monoclonal anti-FLAG M2 and the HA tag with rabbit anti-HA (A to D); N protein was detected with rabbit anti-ANDV-N serum and the HA tag with mouse monoclonal anti-HA (E). Nuclei were stained with DAPI. Images were captured by confocal microscopy. Scale bars, 10m. (A) Localization of NSs expressed alone. (B) Localization of coexpressed NSs and L-K44A. (C) Localization of coexpressed NSs and N terminus of L-K44A. (D) Localization of coexpressed NSs and C terminus of L protein. (E) Localization of coexpressed NSs and N protein.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.134.451.64.448.2]this helix is completely conserved among bunya- and orthomyxo-viruses, and the available endonuclease structures indicate that it contributes to the active site. The putative homologs in LACV and IAV of residue K127 are assumed to contribute to RNA binding (11,13) (Fig. 11).
Second, the nuclease appears to be quite active. As the wild-type endonuclease downregulates its own expression to undetect-able levels (a strong negative-feedback loop), it is only a very low quantity of L protein molecules mediating the effects.
Third, the endonuclease cleaves mRNAs irrespective of se-quence and cytoplasmic or nuclear origin. This would predict that cellular mRNAs are degraded by L protein as well, which would eventually downregulate host cell protein synthesis as demon-strated here by metabolic labeling. The endonuclease may also play a role in regulating virus protein levels. However, in the con-text of a virus infection the properties of the L protein might be different due to sequestration or interaction with other virus pro-teins or RNA. Just coexpression of ANDV N, G, or NSs propro-teins
alone or in combination did not change the low-expression phe-notype (data not shown).
Fourth, the properties of the hantavirus endonuclease are dif-ferent from that of arena- and orthobunyaviruses. L proteins of the latter viruses can be readily expressed in mammalian cells (20, 35) and mutations in the arenavirus endonuclease do not enhance the expression level of L protein or coexpressed proteins (27). Thus, the endonuclease activity of hantaviruses appears to be less tightly regulated than that of arena- and orthobunyaviruses.
In contrast to the latter viruses, there are striking parallels be-tween the IAV endonuclease encoded by the PA gene and hanta-virus L protein. A repressive effect due to expression of the PA gene on the steady-state level of other coexpressed proteins, as well as PA itself—which corresponds to the observations reported here— had been described several years ago, but mistakenly as-cribed to a reduced half-life time of the proteins (36,37); an op-tion we have excluded for the hantavirus L protein. Only recently, it has been shown that this repressive effect is mediated by the FIG 10Intracellular localization of ANDV L and N protein in relation to P body component DCP1a. MVA-T7-infected BHK-21 cells were transfected with constructs for expression of L-R35H-HA, L-K44A-HA, LN-K44A-FLAG (residues 1 to 534), LC-FLAG (residues 962 to 2153), or N protein. The HA tag was detected with rabbit anti-HA, the FLAG tag with rabbit anti-FLAG, N protein with rabbit anti-ANDV-N serum, and P bodies with monoclonal mouse anti-DCP1a. Nuclei were stained with DAPI. Images were captured by confocal microscopy. Scale bars, 10m. (A) Localization of L-K44A and DCP1a. (B) Localization of L-R35H and DCP1a. (C) Localization of N terminus of L-K44A and DCP1a. (D) Localization of C terminus of L and DCP1a. (E) Localization of N protein and DCP1a.
Andes Virus L Protein
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.134.451.63.444.2]so-called PA-X protein, which is expressed via ribosomal frame-shifting from the PA gene (38). PA-X comprises the N-terminal endonuclease domain of PA fused to the X peptide encoded in the
⫹1 frame. Although PA-X is expressed only at a very low level from the PA gene, it efficiently downregulates the level of coex-pressed protein and corresponding mRNA. As shown here for ANDV, mutation of catalytic site residues of the influenza virus endonuclease abolished this effect. Because the repressive effect is mediated by PA-X rather than full-length PA, it is tempting to speculate that in hantaviruses the effect might also be due to ex-pression of a truncated L protein comprising only the endonu-clease domain. However, we could neither identify within the hantavirus L gene a frameshifting motif as described for PA-X nor identify a conserved⫹1 ORF corresponding to the X ORF. Nev-ertheless, this does not exclude the possibility that a truncated L protein, expressed at a low level via any other mechanism, is re-sponsible for the effects described here.
The expressible hantavirus L protein mutants, although pre-dictably endonuclease deficient, may be a useful tool to study pro-tein localization or interaction. However, some influence of the mutations on both aspects may not be fully excluded. Like L pro-teins of arenaviruses and other bunyavirus genera (35,39,40), the hantavirus L protein is localized in granular to punctate cytoplas-mic structures. In agreement with previous studies (5), it some-times accumulated in the perinuclear area. Evidence for expres-sion of NSs protein during ANDV infection has been provided only recently, and its function is still unknown (7). Our data sug-gest that this small protein might interact with L protein, as indi-cated by its relocalization from nucleus to cytoplasm upon the
coexpression of L protein. A small fraction of L protein also colo-calizes with P bodies, mediated by the N-terminal part of the pro-tein harboring the putative endonuclease. This finding is consis-tent with the colocalization of L and N protein, since N protein also colocalizes with P bodies as shown here for ANDV and pre-viously for SNV (14). This supports the proposed mechanism for hantavirus cap-snatching involving the protection by N protein of 5=-terminal nucleotides of capped mRNA in P bodies and their transfer to L protein for endonucleolytic cleavage and initiation of transcription (14). The colocalization of L protein with P bodies in the absence of N protein indicates that both virus proteins target this structure independent of each other.
ACKNOWLEDGMENTS
We thank Romy Kerber, Michaela Lelke, Carola Busch, and Alexandra Bialonski for assistance with experiments and Ulrike Kuckelkorn for per-forming the proteasome activity assay.
This study was supported by grant GU 883/1-1 from the German Research Foundation (DFG). The Department of Virology of the Bern-hard-Nocht-Institute is a WHO Collaborating Centre for Arbovirus and Haemorrhagic Fever Reference and Research (DEU-115).
REFERENCES
1.Macneil A, Nichol ST, Spiropoulou CF.2011. Hantavirus pulmonary syndrome. Virus Res.162:138 –147.
2.Jonsson CB, Figueiredo LT, Vapalahti O.2010. A global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev.23: 412– 441.
3.Hepojoki J, Strandin T, Lankinen H, Vaheri A.2012. Hantavirus struc-ture-molecular interactions behind the scene. J. Gen. Virol.93:1631–1644. FIG 11Sequence comparison of the putative endonuclease of hantaviruses with known endonucleases of orthobunyaviruses and orthomyxoviruses. L protein sequences of ANDV, SNV, HTNV, and PUUV were aligned with the L protein of LACV and the PA protein of IAV using CLUSTAL W (41) and MacVector (MacVector). Amino acid residues are colored according to chemical type. The N termini of LACV L protein and IAV PA harbor endonucleases (11,13), and only those parts of the hantavirus L protein showing significant homology to these enzymes are shown (residues 1 to 51 and residues 95 to 167). The secondary structure (below the sequence alignment) of ANDV L protein was predicted using Jpred (42), while the secondary structure data for LACV and IAV were obtained from the PDB structures 2XI5 and 2W69, respectively. Residues, which have been tested in our study, are indicated with a gray triangle and those, which are important for the low-expression phenotype, are indicated with a black triangle. Residues implicated in the catalytic process of LACV and IAV endonucleases are marked with a filled red triangle; those implicated in RNA binding by an open red triangle. The putative catalytic and RNA-binding residues in hantaviruses— H36, D97, E110, K124, and K127— correspond to H34, D79, D92, K94, and K108 in LACV (13) and H41, D108, E119, K134, and K137 in IAV (11), respectively. The hantavirus sites perfectly match with the putative homologous residues of IAV in the alignment. The match to LACV is not as perfect as to IAV, as the position of the catalytic lysine of LACV (K94, marked with a red triangle) maps to T112 rather than K124 in hantaviruses. Thus, the arrangement of the catalytic site in hantaviruses might be more related to orthomyxoviruses than to orthobunyaviruses.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:10.585.135.454.66.266.2]4. Kukkonen SK, Vaheri A, Plyusnin A. 2005. L protein, the RNA-dependent RNA polymerase of hantaviruses. Arch. Virol.150:533–556. 5.Kukkonen SK, Vaheri A, Plyusnin A.2004. Tula hantavirus L protein is
a 250-kDa perinuclear membrane-associated protein. J. Gen. Virol.85: 1181–1189.
6.Jaaskelainen KM, Kaukinen P, Minskaya ES, Plyusnina A, Vapalahti O, Elliott RM, Weber F, Vaheri A, Plyusnin A.2007. Tula and Puumala hantavirus NSs ORFs are functional and the products inhibit activation of the interferon-beta promoter. J. Med. Virol.79:1527–1536.
7.Vera-Otarola J, Solis L, Soto-Rifo R, Ricci EP, Pino K, Tischler ND, Ohlmann T, Darlix JL, Lopez-Lastra M.2012. The Andes hantavirus NSs protein is expressed from the viral small mRNA by a leaky scanning mech-anism. J. Virol.86:2176 –2187.
8.Garcin D, Lezzi M, Dobbs M, Elliott RM, Schmaljohn C, Kang CY, Kolakofsky D.1995. The 5=ends of Hantaan virus (Bunyaviridae) RNAs suggest a prime-and-realign mechanism for the initiation of RNA synthe-sis. J. Virol.69:5754 –5762.
9.Cheng E, Mir MA. 2012. Signatures of host mRNA 5= terminus for efficient hantavirus cap snatching. J. Virol.86:10173–10185.
10. Plotch SJ, Bouloy M, Ulmanen I, Krug RM. 1981. A unique
cap(m7GpppXm)-dependent influenza virion endonuclease cleaves capped RNAs to generate the primers that initiate viral RNA transcription. Cell23:847– 858.
11. Dias A, Bouvier D, Crepin T, McCarthy AA, Hart DJ, Baudin F, Cusack S, Ruigrok RW.2009. The cap-snatching endonuclease of influenza virus polymerase resides in the PA subunit. Nature458:914 –918.
12. Morin B, Coutard B, Lelke M, Ferron F, Kerber R, Jamal S, Frangeul A, Baronti C, Charrel R, de Lamballerie X, Vonrhein C, Lescar J, Bricogne G, Gunther S, Canard B.2010. The N-terminal domain of the arenavirus L protein is an RNA endonuclease essential in mRNA transcription. PLoS Pathog.6:e1001038. doi:10.1371/journal.ppat.1001038.
13. Reguera J, Weber F, Cusack S.2010. Bunyaviridae RNA polymerases (L-protein) have an N-terminal, influenza-like endonuclease domain, es-sential for viral cap-dependent transcription. PLoS Pathog.6:e1001101. doi:10.1371/journal.ppat.1001101.
14. Mir MA, Duran WA, Hjelle BL, Ye C, Panganiban AT.2008. Storage of cellular 5=mRNA caps in P bodies for viral cap-snatching. Proc. Natl. Acad. Sci. U. S. A.105:19294 –19299.
15. Mir MA, Panganiban AT.2008. A protein that replaces the entire cellular eIF4F complex. EMBO J.27:3129 –3139.
16. Meyer S, Temme C, Wahle E.2004. Messenger RNA turnover in eu-karyotes: pathways and enzymes. Crit. Rev. Biochem. Mol. Biol.39:197– 216.
17. Parker R, Sheth U.2007. P bodies and the control of mRNA translation and degradation. Mol. Cell25:635– 646.
18. Brown KS, Ebihara H, Feldmann H.2012. Development of a minige-nome system for Andes virus, a New World hantavirus. Arch. Virol.157: 2227–2233.
19. Hass M, Golnitz U, Muller S, Becker-Ziaja B, Gunther S.2004. Replicon system for Lassa virus. J. Virol.78:13793–13803.
20. Hass M, Lelke M, Busch C, Becker-Ziaja B, Gunther S.2008. Mutational evidence for a structural model of the Lassa virus RNA polymerase domain and identification of two residues, Gly1394 and Asp1395, that are critical for transcription but not replication of the genome. J. Virol.82:10207– 10217.
21. Buchholz UJ, Finke S, Conzelmann KK. 1999. Generation of bovine respiratory syncytial virus (BRSV) from cDNA: BRSV NS2 is not essential for virus replication in tissue culture, and the human RSV leader region acts as a functional BRSV genome promoter. J. Virol.73:251–259. 22. Sutter G, Ohlmann M, Erfle V.1995. Non-replicating vaccinia vector
efficiently expresses bacteriophage T7 RNA polymerase. FEBS Lett.371: 9 –12.
23. Gu W, Zhou T, Wilke CO.2010. A universal trend of reduced mRNA stability near the translation-initiation site in prokaryotes and eukaryotes. PLoS Comput. Biol.6:e1000664. doi:10.1371/journal.pcbi.1000664. 24. Gingold H, Pilpel Y.2011. Determinants of translation efficiency and
accuracy. Mol. Syst. Biol.7:481. doi:10.1038/msb.2011.14.
25. Geimonen E, Fernandez I, Gavrilovskaya IN, Mackow ER.2003. Ty-rosine residues direct the ubiquitination and degradation of the NY-1 hantavirus G1 cytoplasmic tail. J. Virol.77:10760 –10868.
26. Kravtsova-Ivantsiv Y, Ciechanover A.2012. Non-canonical ubiquitin-based signals for proteasomal degradation. J. Cell Sci.125:539 –548. 27. Lelke M, Brunotte L, Busch C, Gunther S.2010. An N-terminal region
of Lassa virus L protein plays a critical role in transcription but not repli-cation of the virus genome. J. Virol.84:1934 –1944.
28. Zhang Y, Li XH, Jiang H, Huang CX, Wang PZ, Mou DL, Sun L, Xu Z, Wei X, Bai XF.2008. Expression of L protein of Hantaan virus 84FLi strain and its application for recovery of minigenomes. APMIS116:1089 – 1096.
29. Bridgen A, Elliott RM.1996. Rescue of a segmented negative-strand RNA virus entirely from cloned complementary DNAs. Proc. Natl. Acad. Sci. U. S. A.93:15400 –15404.
30. Ikegami T, Won S, Peters CJ, Makino S.2006. Rescue of infectious rift valley fever virus entirely from cDNA, analysis of virus lacking the NSs gene, and expression of a foreign gene. J. Virol.80:2933–2940.
31. Bergeron E, Albarino CG, Khristova ML, Nichol ST.2010. Crimean-Congo hemorrhagic fever virus-encoded ovarian tumor protease activity is dispensable for virus RNA polymerase function. J. Virol.84:216 –226. 32. Flatz L, Bergthaler A, de la Torre JC, Pinschewer DD.2006. Recovery of
an arenavirus entirely from RNA polymerase I/II-driven cDNA. Proc. Natl. Acad. Sci. U. S. A.103:4663– 4668.
33. Neumann G, Watanabe T, Ito H, Watanabe S, Goto H, Gao P, Hughes M, Perez DR, Donis R, Hoffmann E, Hobom G, Kawaoka Y. 1999. Generation of influenza A viruses entirely from cloned cDNAs. Proc. Natl. Acad. Sci. U. S. A.96:9345–9350.
34. Flick K, Hooper JW, Schmaljohn CS, Pettersson RF, Feldmann H, Flick R.2003. Rescue of Hantaan virus minigenomes. Virology306:219 –224. 35. Shi X, Elliott RM.2009. Generation and analysis of recombinant
Buny-amwera orthobunyaviruses expressing V5 epitope-tagged L proteins. J. Gen. Virol.90:297–306.
36. Sanz-Ezquerro JJ, Zurcher T, de la Luna S, Ortin J, Nieto A.1996. The amino-terminal one-third of the influenza virus PA protein is responsible for the induction of proteolysis. J. Virol.70:1905–1911.
37. Sanz-Ezquerro JJ, de la Luna S, Ortin J, Nieto A. 1995. Individual expression of influenza virus PA protein induces degradation of coex-pressed proteins. J. Virol.69:2420 –2426.
38. Jagger BW, Wise HM, Kash JC, Walters KA, Wills NM, Xiao YL, Dunfee RL, Schwartzman LM, Ozinsky A, Bell GL, Dalton RM, Lo A, Efstathiou S, Atkins JF, Firth AE, Taubenberger JK, Digard P.2012. An overlapping protein-coding region in influenza A virus segment 3 modu-lates the host response. Science337:199 –204.
39. Brunotte L, Lelke M, Hass M, Kleinsteuber K, Becker-Ziaja B, Gunther S.2011. Domain structure of Lassa virus L protein. J. Virol.85:324 –333. 40. Brennan B, Li P, Elliott RM.2011. Generation and characterization of a recombinant Rift Valley fever virus expressing a V5 epitope-tagged RNA-dependent RNA polymerase. J. Gen. Virol.92:2906 –2913.
41. Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG.2007. CLUSTAL W and CLUSTAL X version 2.0. Bioinformatics23:2947–2948.
42. Cole C, Barber JD, Barton GJ.2008. The Jpred3 secondary structure prediction server. Nucleic Acids Res.36:W197–W201.
Andes Virus L Protein