RNA Replication and Membrane Modification Require the Same
Functions of Alphavirus Nonstructural Proteins
Katri Kallio,aKirsi Hellström,aEija Jokitalo,bTero Aholaa
Department of Food and Environmental Sciencesaand Institute of Biotechnology,bUniversity of Helsinki, Helsinki, Finland
The alphaviruses induce membrane invaginations known as spherules as their RNA replication sites. Here, we show that inacti-vation of any function (polymerase, helicase, protease, or membrane association) essential for RNA synthesis also prevents the generation of spherule structures in a Semliki Forest virustrans-replication system. Mutants capable of negative-strand synthe-sis, including those defective in RNA capping, gave rise to spherules. Recruitment of RNA to membranes in the absence of spher-ule formation was not detected.
A
lphaviruses are a well-studied group of enveloped viruses with a single-stranded, positive-sense RNA genome and in-clude several human and animal pathogens (1). The replication of all positive-strand RNA viruses takes place in association with cellular membranes which are heavily modified or reorganized to act as efficient platforms for genome amplification (2,3). It seems that there are two main classes of such membrane modifications: the formation of small invaginations (termed spherules) on mem-branes and the generation of double-membrane vesicles (4). The alphaviruses and related alphavirus-like plant viruses, as well as nodaviruses, flaviviruses, and tombusviruses, induce the forma-tion of spherule-type structures (4).Alphavirus replication is catalyzed by four virus-encoded non-structural proteins, nsP1 to nsP4 (reviewed in reference5), which arise from a precursor polyprotein termed P1234. nsP1 is respon-sible for the capping of the viral genome through its methyltrans-ferase and guanylyltransmethyltrans-ferase activities. nsP1 also attaches the replication complexes to membranes via an amphipathic alpha helix and covalent palmitoylation of the protein. nsP2 contains an N-terminal helicase domain, which also acts as RNA triphospha-tase, and a C-terminal protease domain responsible for polypro-tein processing. The role of nsP3 is poorly understood compared to the roles of the other nsPs, but it is heavily phosphorylated and has important roles in virus-host interactions. nsP4 is the catalytic RNA-dependent RNA polymerase (RdRp) subunit acting in RNA synthesis.
We have recently constructed a plasmid-basedtrans -replica-tion system for Semliki Forest virus (SFV) which allows high-level expression and analysis of replicase protein mutants that would be lethal in the context of virus infection (6, 7). We previously showed that the presence of both an RNA template and a func-tional polymerase is required for the formation of spherules (7). We have now introduced to thetrans-replication system multiple point mutations of nonstructural proteins which destroy enzy-matic activities or posttranslational modifications in order to study the requirements for RNA replication and for the formation of membranous spherule structures. The mutations (Table 1) were transferred from prior constructs using standard methods, followed by sequence verification.
The polyprotein, together with a short template (Fig. 1A) which yields the highest luciferase levels, was transfected to T7 polymerase-expressing BSR T7/5 cells using Lipofectamine LTX (Invitrogen) as previously described (6,7). Initially,Renilla
lucif-erase expression from the template was used to assess the effi-ciency of replication at 16 h posttransfection. Since most of the mutations were completely inactivating for basic functions of the proteins, it was not surprising that only two showed a low-level increase of luciferase activity above the background (Table 1): the
⌬50 mutant, in which the phosphorylated region of nsP3 was deleted, and the P1^2^3Z4 mutant, in which the cleavage sites between nsP1, nsP2, and nsP3 were inactivated.
Received28 September 2015Accepted12 November 2015
Accepted manuscript posted online18 November 2015
CitationKallio K, Hellström K, Jokitalo E, Ahola T. 2016. RNA replication and membrane modification require the same functions of alphavirus nonstructural proteins. J Virol 90:1687–1692.doi:10.1128/JVI.02484-15.
Editor:A. Simon
Address correspondence to Tero Ahola, [email protected].
K.K. and K.H. contributed equally to this article.
[image:1.585.299.541.281.467.2]Copyright © 2016, American Society for Microbiology. All Rights Reserved. TABLE 1The luciferase levels produced by the mutants duringtrans -replication, together with the sources and known effects of the mutations
Mutanta
%
replicationb Effect
P123Z4 100 None (wild-type protein with ZsGreen fusion in nsP3)
H38A 0.7 Inactivates guanylyltransferase (8) D64A 0.4 Inactivates methyltransferase (8) Y249A 0.9 Inactivates methyltransferase and possibly
prevents membrane association (8,9,10) R253E 0.8 Prevents membrane association (9,11) W259A 0.2 Prevents membrane association (9,11) C418-420A 0.6 Removes palmitoylation site (12,13) GNS 0.2 Inactivates helicase, NTPase, and RNA
triphosphatase (14,15) CA 0.4 Inactivates protease (16,17) ⌬50 5.0 Removes phosphorylation sites (18) GAA 0.4 Inactivates polymerase (19)
P1^2^3Z4 2.6 Destroys nsP1/2 and nsP2/3 cleavage sites (20,21) P123Z 0.5 Produces P123Z without nsP4
a
The nsP1 mutants are named based on the amino acid position, whereas the others have individual names (see alsoFig. 1A).
b
Data represent percentRenillaluciferase activity compared to wild-type P123Z4.
crossmark
on November 7, 2019 by guest
http://jvi.asm.org/
The expression levels of nsP1 and nsP3-ZsG fusion were veri-fied by Western blotting (6) for those mutants that had normal polyprotein processing (Fig. 1B), and the presence or absence of nsP4 was verified for the relevant mutants (Fig. 1C). The levels
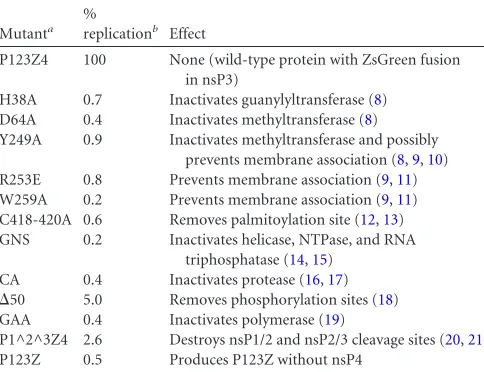
mostly corresponded to those of the wild-type protein, although some exceptions were noted; e.g., nsP1 was specifically present at lower levels when its ability to interact with membranes was com-promised (mutants R253E and W259A). Mutants perturbing the FIG 1Expression of and RNA replication by the protein mutants. (A) Scheme of the polyprotein construct and the short template (Tshort). The T7 promoter drives the transcription of the constructs. The expression of the polyprotein is enhanced by the presence of an internal ribosome entry site (IRES), and it contains the fluorescent protein ZsGreen (ZsG) fused with nsP3. The different mutations constructed are marked in the polyprotein. The template contains viral 5=-untranslated-region (5=UTR) and 3=UTR sequences as well as the subgenomic promoter (SGP), and its termination after the poly(A) sequence is mediated by hepatitis delta antigenomic ribozyme (Rz). The only gene expressed from Tshort isRenillaluciferase (Rluc). aa, amino acid. (B to D) Expression of the nonstructural proteins studied by Western blotting using antibodies against nsP1 and nsP3 (B), nsP4 (C), or nsP3 (D), all shown in red, at 16 h posttransfection for the indicated mutants. The template was always cotransfected with the indicated polyprotein, and-actin (green) was used as a loading control. (E) The amounts of positive- and negative-strand RNAs present at 16 h posttransfection for each polyprotein mutant as analyzed by Northern blotting. The migration positions of Tshort are indicated by arrows. The larger bands in the (⫹)RNA blot result from the failure of T7 to terminate properly (6). The amount of the correctly sized RNA band was quantified as a percentage compared to wild-type (wt) P123Z4⫾standard deviation based on the results of three biological-replicate experiments.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:2.585.123.459.64.573.2]processing of the polyprotein (CA and P1^2^3Z4) gave rise to P123Z4 and P123Z (arrows inFig. 1D), respectively, but also to anomalous smaller fragments.
The levels of positive-strand and negative-strand RNAs pres-ent were analyzed by Northern blotting (6) (Fig. 1E). Only the wild-type polyprotein catalyzed a large excess of positive-strand synthesis compared to the template-only control. Interestingly, the P1^2^3Z4 processing-defective mutant made as much nega-tive-strand RNA as the wild-type protein. Additionally, the nsP1 mutants H38A, D64A, and Y249A each catalyzed significant (30% to 40%) negative-strand synthesis (all quantifications were made with the program ImageJ 1.47v).
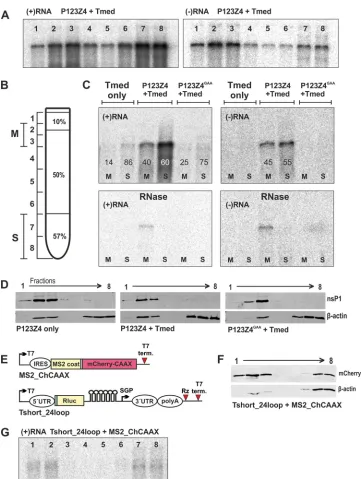
The association of RNA and proteins with membranes was studied by flotation in a discontinuous sucrose gradient prepared in 150 mM NaCl–50 mM Tris-HCl (pH 7.5) in 5-ml tubes, as previously described (11), with a slightly modified sucrose com-position (Fig. 2). The template Tmed (7) was used to achieve ex-actly comparable conditions in subsequent experiments. In the presence of wild-type P123Z4 (full replication), the negative strands were predominantly localized with membranes in floating fractions 2 and 3. Significant amounts of the positive strands were also membrane associated (Fig. 2AtoC). In contrast, when the RNA was expressed alone in the absence of replicase, it did not associate with membranes. This situation did not change when the polymerase-defective mutant GAA was coexpressed with the RNA (Fig. 2C, upper panels). Thus, we did not detect significant re-cruitment of positive-strand RNA to the membrane fraction in the absence of replication. When the fractions were treated with 100
g/ml RNase A for 1 h at room temperature, only the membrane fraction prepared under full-replication conditions showed pro-tection of both RNA strands (Fig. 2C, lower panels), indicating the presence of protective structures. There were significant quanti-ties of negative strands in the soluble fraction, and these were vulnerable to RNase treatment, suggesting that some spherules might break down during the experiment. The membrane-an-choring protein nsP1 was always quantitatively associated with membranes, irrespective of the presence or absence of RNA or the polymerase mutation within the replicase (Fig. 2D). For a positive control, we were able to induce⬃50% RNA recruitment to mem-branes artificially by expression of a strong RNA binding protein modified to bind membranes and by inclusion of the correspond-ing RNA bindcorrespond-ing sites in the template (Fig. 2EtoG).
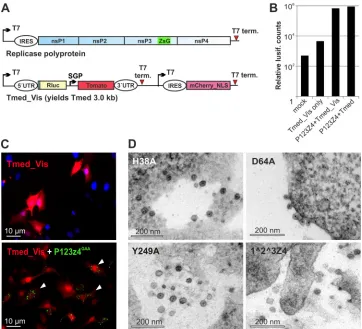
Since many of the mutants did not replicate, it was not possible to visualize the presence of the template using the previous con-structs, which had a red fluorescent marker under the control of the viral subgenomic promoter (7). Therefore, a new version of Tmed, Tmed_Vis, which contains an additional reading frame for the marker, independently driven by a second T7 promoter, was created (Fig. 3A). The sequence encoding fluorescent protein mCherry with a nuclear localization signal (PKKKRKV) added to its C terminus was first cloned in vector pTM1 (22), followed by PCR amplification of the entire new cassette from the T7 pro-moter to the terminator, which was then cloned to the SacI site of Tmed (7). The derived Tmed_Vis template replicated as well as the original Tmed, based on luciferase activity (Fig. 3B). It also showed strong red fluorescence in transfected cells that was mostly concentrated in the nucleus (Fig. 3C). The nsP3-ZsG fusion was localized in cytoplasmic granules/aggregates (Fig. 3C), which are prominent for nsP3 both during SFV replication and when the protein is expressed alone (23,24). Thus, cells containing both the
replicase and the template could easily be visualized for the repli-cation-defective mutants.
Cells cotransfected with the mutant replicases and Tmed_Vis were then subjected to correlative light and electron microscopy (CLEM) analysis as described previously (7). In each case, only cells expressing both markers were selected for visualization at the EM level. Spherule structures were clearly found in cells with four mutants: H38A, D64A, Y249A, and P1^2^3Z4 (Fig. 3D). The re-sults seen with the⌬50 mutant were inconclusive (see next sec-tion), and in spite of an extensive search, spherules were not found for any of the other mutants. Thus, there was an absolute correla-tion with the ability to synthesize RNA in the generacorrela-tion of spher-ules. The helicase and protease activities of nsP2 (14,25), the poly-merase activity of nsP4 (19), and the membrane binding ability of nsP1 (9) were each essential for all replication steps, since neither negative- nor positive-strand RNA was detected (Fig. 1E). The P1^2^3Z4 mutant synthesized negative strands at wild-type lev-els, but few positive strands, fully recapitulating previous results indicating a specific role in negative-strand synthesis for the par-tially processed polyprotein (26–29).
RNA capping activity was not needed for negative-strand syn-thesis or for spherule biogenesis (Fig. 3D). We think it is likely that negative-strand synthesis is sufficient for spherule formation. However, similarly to the P1^2^3Z4 mutant, the capping mu-tants may be able to make some positive-sense RNA, which for the nsP1 mutants would be uncapped and therefore unstable and poorly translated. InSaccharomyces cerevisiaeexpressing similar Brome mosaic virus (BMV) RNA capping mutants, newly made positive-strand RNAs could also be detected when the major RNA degradation pathway in the yeast was disrupted (30). The current results also indicate that the main function of residue Y249, which is highly conserved in the alphavirus-like capping enzymes (31,
32), crucially contributes to the enzymatic activity of nsP1, since the mutant phenotype is similar to those of the other active site mutants and not to those of the membrane association mutants, with which Y249A has also been grouped (9). The⌬50 mutant gave increased luciferase activity and therefore should have pro-duced some new negative strands, which we failed to detect, and it also produced few if any spherules. However, it may be that the actual level of RNA synthesis is very low. Luciferase activity greatly amplifies the replication signal, probably also due to the capping and efficient translation of RNAs made by the SFV replicase com-pared to the uncapped RNAs made by T7 polymerase. Thus, it seems that the⌬50 mutant has a specific defect intrans -replica-tion, since⌬50 mutant viruses replicate quite well (18), producing prominent spherules (33).
For the polymerase mutant (Fig. 2C), or for P123Z in the ab-sence of nsP4 (data not shown), we did not find evidence for the recruitment of positive-strand RNA to membrane fractions. Also, we did not observe significantly reduced luciferase translation from the template RNAs when any of the replication-defective polyproteins were coexpressed (data not shown), which could serve as another indication for viral RNA-protein interactions prior to replication. Viral RNA recruitment to membranes has been studied and prominently detected in the presence of BMV and Flock House nodavirus (FHV) replicase proteins (34–36). For BMV, spherules are formed by protein 1a also in the absence of replication, and it seems that RNA is mainly stabilized inside the spherules (34). In contrast, FHV, similarly to SFV, requires RNA replication for spherule biogenesis (37). Nevertheless, a polymer-Alphavirus Replication and Membrane Modification
on November 7, 2019 by guest
http://jvi.asm.org/
ase-inactive FHV mutant can still fully recruit viral RNA to a membrane-associated state (35). Several domains of the FHV rep-licase protein A, including the polymerase domain itself, are re-quired for membrane recruitment (35), but no other point mu-tants beside polymerase inactivation have been studied to date.
Although SFVtrans-replication is efficient overall (6), it may be that the RNA recruitment was inefficient or readily reversible under the conditions used, and in future work we will explore alternative methods, such as RNA-protein cross-linking to detect the potential recruitment intermediate, which should exist at least FIG 2Membrane association of RNA. (A) The ability of positive- and negative-strand RNA to float with membranes, when replication is ongoing. (B) Scheme of the sucrose gradient used for flotation and the fractions (M, membrane; S, soluble) collected. The sample was initially present in 57% (wt/wt) sucrose, and membranes floated through the 50% layer to the interface with the 10% layer. For subsequent analysis, fractions were combined as shown. (C) The prominent floating fractions (marked “M” for membrane) and soluble fractions (“S”) were combined for easier comparison between the wild type and the mutants. The presence of RNA strands was quantified in Northern blots, taking the combined amount of M plus S as 100%. For the bottom panels, these fractions were treated with RNase A before Northern analysis was performed. (D) nsP1 and the-actin control present in the fractions after the indicated plasmids had been transfected to cells. A minor fraction of actin was associated with the membranes. (E) Template Tshort_24loop contains 24 repeats of MS2 bacteriophage coat binding loop (38,39) and was coexpressed with MS2 coat protein fused with the fluorescent protein mCherry, containing a C-terminal CAAX sequence inducing efficient prenylation and membrane association (40,41). (F and G) Ability of the MS2 coat fusion protein to interact with membranes (F) and to induce the flotation of the Tshort_24loop template RNA (G). All samples used in these experiments were collected at 16 h posttransfection. Each of the experiments was repeated 3 to 5 times with similar results.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.111.472.60.539.2]transiently. In summary, we have established a tight link between RNA replication and spherule biogenesis for multiple mutants of SFV replicase and have developed Tmed_Vis as a tool to detect the presence of template RNA in individual cells.
ACKNOWLEDGMENTS
We thank Ilkka Fagerlund for performing some Northern blotting exper-iments and Mervi Lindman and Arja Strandell for excellent technical as-sistance in EM.
This study was supported by the Academy of Finland (grant 265997). K.K. was supported in part by a fellowship from the Helsinki Graduate Program in Biotechnology and Molecular Biology and K.H. by an Acad-emy of Finland postdoctoral fellowship.
FUNDING INFORMATION
Suomen Akatemia (Academy of Finland) provided funding under grant number 265997.
The funders had no role in study design, data collection and interpreta-tion, or the decision to submit the work for publication.
REFERENCES
1.Griffin DE.2013. Alphaviruses, p 651– 686.InKnipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B
(ed), Fields virology, 6th ed, vol 1. Lippincott Williams & Wilkins, Phila-delphia, PA.
2.Salonen A, Ahola T, Kaariainen L.2005. Viral RNA replication in association with cellular membranes. Curr Top Microbiol Immunol 285:139 –173.
3.Belov GA, van Kuppeveld FJ. 2012. (⫹)RNA viruses rewire cellular pathways to build replication organelles. Curr Opin Virol2:740 –747.
http://dx.doi.org/10.1016/j.coviro.2012.09.006.
4.Paul D, Bartenschlager R. 2013. Architecture and biogenesis of plus-strand RNA virus replication factories. World J Virol2:32– 48.http://dx .doi.org/10.5501/wjv.v2.i2.32.
5.Kääriäinen L, Ahola T.2002. Functions of alphavirus nonstructural pro-teins in RNA replication. Prog Nucleic Acid Res Mol Biol71:187–222.
http://dx.doi.org/10.1016/S0079-6603(02)71044-1.
6.Kallio K, Hellstrom K, Balistreri G, Spuul P, Jokitalo E, Ahola T.2013. Template RNA length determines the size of replication complex spher-ules for Semliki Forest virus. J Virol87:9125–9134.http://dx.doi.org/10 .1128/JVI.00660-13.
7.Spuul P, Balistreri G, Hellstrom K, Golubtsov AV, Jokitalo E, Ahola T. 2011. Assembly of alphavirus replication complexes from RNA and pro-tein components in a novel trans-replication system in mammalian cells. J Virol85:4739 – 4751.http://dx.doi.org/10.1128/JVI.00085-11.
8.Ahola T, Laakkonen P, Vihinen H, Kaariainen L.1997. Critical residues of Semliki Forest virus RNA capping enzyme involved in methyltrans-ferase and guanylyltransmethyltrans-ferase-like activities. J Virol71:392–397. 9.Spuul P, Salonen A, Merits A, Jokitalo E, Kaariainen L, Ahola T.2007.
Role of the amphipathic peptide of Semliki forest virus replicase protein FIG 3Formation of spherules. (A) Scheme of the polyprotein and template Tmed_Vis used for CLEM analysis of the mutants. Abbreviations are as defined in theFig. 1Alegend. mCherry_NLS denotes the red fluorescent protein mCherry containing a nuclear localization signal. (B) Luciferase (lusif.) activity produced by Tmed_Vis compared with the previous Tmed template. The indicated combinations were transfected, and activity was measured at 16 h posttransfection. (C) Visualization of the presence of both template and polyprotein constructs in the same cells, when a polymerase-inactivated mutant was used. The upper panel shows the transfection of Tmed_Vis only. In the lower panel, cells containing both the template (red signal mainly in the nucleus) and replicase, as indicated (nsP3-ZsG [green puncta]), are marked with arrowheads. (D) Spherules produced by the indicated polyprotein mutants during template replication.
Alphavirus Replication and Membrane Modification
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.111.473.66.395.2]nsP1 in membrane association and virus replication. J Virol81:872– 883.
http://dx.doi.org/10.1128/JVI.01785-06.
10. Wang HL, O’Rear J, Stollar V.1996. Mutagenesis of the Sindbis virus nsP1 protein: effects on methyltransferase activity and viral infectivity. Virology217:527–531.http://dx.doi.org/10.1006/viro.1996.0147. 11. Ahola T, Lampio A, Auvinen P, Kaariainen L.1999. Semliki Forest virus
mRNA capping enzyme requires association with anionic membrane phospholipids for activity. EMBO J18:3164 –3172.http://dx.doi.org/10 .1093/emboj/18.11.3164.
12. Laakkonen P, Ahola T, Kaariainen L.1996. The effects of palmitoyl-ation on membrane associpalmitoyl-ation of Semliki forest virus RNA capping enzyme. J Biol Chem271:28567–28571.http://dx.doi.org/10.1074/jbc .271.45.28567.
13. Zusinaite E, Tints K, Kiiver K, Spuul P, Karo-Astover L, Merits A, Sarand I.2007. Mutations at the palmitoylation site of non-structural protein nsP1 of Semliki Forest virus attenuate virus replication and cause accumulation of compensatory mutations. J Gen Virol88:1977–1985.
http://dx.doi.org/10.1099/vir.0.82865-0.
14. Das PK, Merits A, Lulla A.2014. Functional crosstalk between distant domains of Chikungunya virus non-structural protein 2 is decisive for its RNA-modulating activity. J Biol Chem289:5635–5653.http://dx.doi.org /10.1074/jbc.M113.503433.
15. Vasiljeva L, Merits A, Auvinen P, Kääriäinen L.2000. Identification of a novel function of the alphavirus capping apparatus. RNA 5= -triphosphatase activity of Nsp2. J Biol Chem275:17281–17287. 16. Merits A, Vasiljeva L, Ahola T, Kaariainen L, Auvinen P.2001.
Proteo-lytic processing of Semliki Forest virus-specific non-structural polypro-tein by nsP2 protease. J Gen Virol82:765–773.http://dx.doi.org/10.1099 /0022-1317-82-4-765.
17. Strauss EG, De Groot RJ, Levinson R, Strauss JH.1992. Identification of the active site residues in the nsP2 proteinase of Sindbis virus. Virology 191:932–940.http://dx.doi.org/10.1016/0042-6822(92)90268-T. 18. Vihinen H, Ahola T, Tuittila M, Merits A, Kaariainen L.2001.
Elimi-nation of phosphorylation sites of Semliki Forest virus replicase protein nsP3. J Biol Chem 276:5745–5752. http://dx.doi.org/10.1074/jbc .M006077200.
19. Rubach JK, Wasik BR, Rupp JC, Kuhn RJ, Hardy RW, Smith JL.2009. Characterization of purified Sindbis virus nsP4 RNA-dependent RNA polymerase activity in vitro. Virology384:201–208.http://dx.doi.org/10 .1016/j.virol.2008.10.030.
20. Vasiljeva L, Merits A, Golubtsov A, Sizemskaja V, Kaariainen L, Ahola T.2003. Regulation of the sequential processing of Semliki Forest virus replicase polyprotein. J Biol Chem278:41636 – 41645.http://dx.doi.org /10.1074/jbc.M307481200.
21. Lemm JA, Rice CM.1993. Roles of nonstructural polyproteins and cleav-age products in regulating Sindbis virus RNA replication and transcrip-tion. J Virol67:1916 –1926.
22. Moss B, Elroy-Stein O, Mizukami T, Alexander WA, Fuerst TR.1990. Product review. New mammalian expression vectors. Nature348:91–92. 23. Neuvonen M, Kazlauskas A, Martikainen M, Hinkkanen A, Ahola T, Saksela K. 2011. SH3 domain-mediated recruitment of host cell am-phiphysins by alphavirus nsP3 promotes viral RNA replication. PLoS Pat-hog7:e1002383.http://dx.doi.org/10.1371/journal.ppat.1002383. 24. Salonen A, Vasiljeva L, Merits A, Magden J, Jokitalo E, Kaariainen L.
2003. Properly folded nonstructural polyprotein directs the Semliki Forest virus replication complex to the endosomal compartment. J Virol77: 1691–1702.http://dx.doi.org/10.1128/JVI.77.3.1691-1702.2003. 25. Hardy WR, Strauss JH.1989. Processing the nonstructural polyproteins
of Sindbis virus: nonstructural proteinase is in the C-terminal half of nsP2 and functions both in cis and in trans. J Virol63:4653– 4664.
26. Kim KH, Rumenapf T, Strauss EG, Strauss JH.2004. Regulation of Semliki Forest virus RNA replication: a model for the control of alphavirus
pathogenesis in invertebrate hosts. Virology323:153–163.http://dx.doi .org/10.1016/j.virol.2004.03.009.
27. Lemm JA, Rumenapf T, Strauss EG, Strauss JH, Rice CM.1994. Poly-peptide requirements for assembly of functional Sindbis virus replication complexes: a model for the temporal regulation of minus- and plus-strand RNA synthesis. EMBO J13:2925–2934.
28. Sawicki DL, Sawicki SG.1994. Alphavirus positive and negative strand RNA synthesis and the role of polyproteins in formation of viral replica-tion complexes. Arch Virol Suppl9:393– 405.
29. Wang YF, Sawicki SG, Sawicki DL.1994. Alphavirus nsP3 functions to form replication complexes transcribing negative-strand RNA. J Virol68: 6466 – 6475.
30. Ahola T, den Boon JA, Ahlquist P. 2000. Helicase and capping enzyme active site mutations in brome mosaic virus protein 1a cause defects in template recruitment, negative-strand RNA synthesis, and viral RNA capping. J Virol74:8803– 8811.http://dx.doi.org/10.1128 /JVI.74.19.8803-8811.2000.
31. Rozanov MN, Koonin EV, Gorbalenya AE.1992. Conservation of the putative methyltransferase domain: a hallmark of the ‘Sindbis-like’ super-group of positive-strand RNA viruses. J Gen Virol73(Pt 8):2129 –2134.
http://dx.doi.org/10.1099/0022-1317-73-8-2129.
32. Ahola T, Karlin DG.2015. Sequence analysis reveals a conserved exten-sion in the capping enzyme of the alphavirus supergroup, and a homolo-gous domain in nodaviruses. Biol Direct10:16.http://dx.doi.org/10.1186 /s13062-015-0050-0.
33. Thaa B, Biasiotto R, Eng K, Neuvonen M, Gotte B, Rheinemann L, Mutso M, Utt A, Varghese F, Balistreri G, Merits A, Ahola T, McInerney GM. 2015. Differential phosphatidylinositol-3-kinase-Akt-mTOR activation by Semliki Forest and Chikungunya viruses is dependent on nsP3 and connected to replication complex internalization. J Virol89:11420 –11437.http://dx.doi .org/10.1128/JVI.01579-15.
34. Schwartz M, Chen J, Janda M, Sullivan M, den Boon J, Ahlquist P. 2002. A positive-strand RNA virus replication complex parallels form and function of retrovirus capsids. Mol Cell9:505–514.http://dx.doi.org/10 .1016/S1097-2765(02)00474-4.
35. Van Wynsberghe PM, Chen HR, Ahlquist P.2007. Nodavirus RNA replication protein a induces membrane association of genomic RNA. J Virol81:4633– 4644.http://dx.doi.org/10.1128/JVI.02267-06.
36. Wang X, Lee WM, Watanabe T, Schwartz M, Janda M, Ahlquist P. 2005. Brome mosaic virus 1a nucleoside triphosphatase/helicase domain plays crucial roles in recruiting RNA replication templates. J Virol79: 13747–13758.http://dx.doi.org/10.1128/JVI.79.21.13747-13758.2005. 37. Kopek BG, Settles EW, Friesen PD, Ahlquist P. 2010.
Nodavirus-induced membrane rearrangement in replication complex assembly re-quires replicase protein A, RNA templates, and polymerase activity. J Virol 84:12492–12503.http://dx.doi.org/10.1128/JVI.01495-10.
38. Boireau S, Maiuri P, Basyuk E, de la Mata M, Knezevich A, Pradet-Balade B, Backer V, Kornblihtt A, Marcello A, Bertrand E.2007. The transcriptional cycle of HIV-1 in real-time and live cells. J Cell Biol179: 291–304.http://dx.doi.org/10.1083/jcb.200706018.
39. Fusco D, Accornero N, Lavoie B, Shenoy SM, Blanchard JM, Singer RH, Bertrand E.2003. Single mRNA molecules demonstrate probabilistic movement in living mammalian cells. Curr Biol13:161–167.http://dx.doi .org/10.1016/S0960-9822(02)01436-7.
40. Huet G, Rajakyla EK, Viita T, Skarp KP, Crivaro M, Dopie J, Vartiainen MK.2013. Actin-regulated feedback loop based on Phactr4, PP1 and co-filin maintains the actin monomer pool. J Cell Sci126:497–507.http://dx .doi.org/10.1242/jcs.113241.
41. van Rheenen J, Song X, van Roosmalen W, Cammer M, Chen X, Desmarais V, Yip SC, Backer JM, Eddy RJ, Condeelis JS.2007. EGF-induced PIP2 hydrolysis releases and activates cofilin locally in carcinoma cells. J Cell Biol179:1247–1259.http://dx.doi.org/10.1083/jcb.200706206.