Copyright ©)1976 American Society for Microbiology PrintedinU.SA.
Deletion
Mapping of Moloney Type C Virus:
Polypeptide
and
Nucleic Acid Expression
in
Different
Transforming
Virus
Isolates
WADE P. PARKS,* RICHARD S. HOWK, ANTHONY ANISOWICZ, AND EDWARD M. SCOLNICK
Laboratory of Tumor Virus Genetics, National Cancer Institute, Bethesda, Maryland 20014,* andMeloy
Laboratories,
Rockville,
Maryland
20850Received for publication12December 1975
The viral
polypeptides and viral RNA present in cells transformed by various
replication-defective type C viruses derived from Moloney
murine leukemia
virus were
examined. Different portions of the Moloney type C viral genome
were
retained
inthe different transforming viruses, thus providing an
opportu-nity
for deletion mapping of the Moloney type C genome. DNA
transcripts were
prepared that are complementary to three distinct nonoverlapping portions of
the Moloney
viral genome. Based on an analysis of the polypeptides produced in
the different transformed cells, one complementary DNA apparently represents
sequences
coding for Moloney gp7O; one complementary DNA represents a
region
of the
Moloney genome common
toall of the transforming viruses
examined, and one complementary DNA represents the sequences for
p30, p15,
p10,12.
A
partial map of the
different
replication-defective transforming viruses
is
suggested.
Mammalian type C
virusesthat are
capable
of fibroblast transformation
intissue
culture
(FT>)
have been
isolated
from
murine,
feline,
and primate hosts (13,
14,16,
41,45).
Among
the
FT+
murineviruses, three
have been
de-rived
by passage of the Moloney
murineleuke-mia
virus(Mo-MuLV)
invarious
rodents. First,
the
Harvey sarcoma
virus(Ha-SV)
wasisolated
by
passage
of Mo-MuLV
in rats(18).
Second,
the
Moloney
sarcoma virus(Mo-SV)
wasiso-lated
by passage of the Mo-MuLV
inBALB/c
mice (27).
Third,
the
Abelson
virus wasisolated
by passage of Mo-MuLV
in aBALB/c
mousetreated with
aglucocorticoid
(4).
All
mammalian
FT+
virusesexamined
todate
have been shown
tobe defective for
repli-cation;
thus, they
canbe
isolated and studied
intransformed nonproducer
cells free of
replicat-ing
helper
virus(2, 5,
6). Nucleic
acid
hybridi-zation
experiments
utilizing
such
nonproducer
cells have
indicated that both Mo-SV and
Ha-SV
arerecombinants
composed
of
twodistinct
classes of
nucleic
acid sequences
(35).
Mo-SV
isarecombinant between
part
of Mo-MuLV andadditional
sarcoma-specific
sequences
(36).
Similarly,
Ha-SV iscomposed
ofa setof
Mo-MuLVsequences
and
anadditional
setof
sar-coma-specific
sequencesderived
fromrats(38);
the
sarcomavirus-specific
sequences
inMo-SV
and Ha-SV
aredistinct from each other.
Al-though several important
biological
studies
have been reported (32, 33, 40), analysis of the
Abelson viral genome and associated proteins
has been more
preliminary. Abelson virus also
consists
partly of Mo-MuLV genetic
informa-tion, but
additional sequences not present in
Mo-MuLV
have not yet been
demonstrated in
the
Abelson-transforming virus. Nevertheless,
neither the
sarcoma-specific sequence of Mo-SV
or
Ha-SV
iscontained
inAbelson
virus-trans-formed cells
(36).In current
studies,
wehave
focused
not onthe
sarcoma-specific
sequences
of
each of these
FT+ viruses
but
onthe Mo-MuLV
portionof
each virus. Since each of the genomes
responsi-ble for fibroblast transformation appears
tobe
arecombinant with part of the genome
Mo-MuLV,
weundertook experiments
to attempt tocorrelate
the Mo-MuLV nucleic
acid
se-quences remaining
ineach of these FT+ isolates
with
the Mo-MuLV
structural
polypeptides
synthesized
intransformed
heterologous
non-producer cells. To
measurethe Mo-MuLV
nu-cleic
acid sequences, viral
complementary
DNA
(cDNA)
wasfractionated
by
hybridization
toRNA
from different
nonproducer
cells.
Our
re-sults
suggestthat
differentpolypeptides
and
different portions of the
Mo-MuLVgenome
arepresent
intransformed
nonproducer cells,
thatdeletion
mapping of Mo-MuLV
ispossible,
and
that, by correlating
theproteins
synthesized
with
the
RNAexpressed,
cDNA
probes
repre-491
on November 10, 2019 by guest
http://jvi.asm.org/
senting different
portions
of theMo-MuLV ge-nome canbe
prepared.
MATERIALS AND METHODS
Cells and viruses. All cells were grown in the Dulbecco-Vogt modificationofEagleminimal
essen-tial medium with high glucose containing either
10%calfserum(Colorado Serum Co., Denver,Colo.) or10%fetal calfserum(Gibco, Long Island, N.Y.).
Cells were grown in humidified Wedco incubators (Wedco, Silver Spring, Md.) in an atmosphere of
10%CO2orindisposable glass bottles(Bellco Glass,
Inc., Vineland, N.J.) on a roller deck apparatus. Viruses were recovered from supernatant fluids
from roller bottle cultures and purified in 20-liter amounts by continuous flow centrifugation in su-crosedensitygradients. The cells andvirus strains employed in the present studyand their relevant
characteristics are provided in Table 1. The cells
used for thepropagation of these viruses were as
follows: NIH 3T3 cells (21) and normal ratkidney (NRK) cells (12) from George Todaro, Bethesda, Md.; mink lung fibroblasts (CCL64), rabbitcornea
cells (CCL60), and a canine kidney cell, MDCK
(CCL34), from theAmericanType Culture
Collec-tion; and the Sc-i wild mouse cell permissive for
replication of N-orB-tropic murine type C viruses (17)from Janet Hartley, Bethesda, Md.
Synthesis of viral-specific [3H]DNA. The
endoge-nous reverse transcriptase reaction from sucrose
densitygradient-banded viruseswasusedto synthe-sizethe[3H]deoxycytidine-labeled cDNA's; the
con-ditions have been fully detailedinearlier publica-tions (37, 39). Optimal detergent concentrations
wereexplored for eachviruspreparation for
synthe-sis ofprobes and variedfrom 0.01 to 0.02% Triton
X-100.The specific activityof thecDNA'swas
approxi-mately2x107 counts/minperjug.The cDNA homol-ogoustoMoloney leukemia viruswaspreparedfrom
Mo-MuLV grown inNIH 3T3 cells. The cDNA
ho-mologoustoMo-SVwasprepared fromavirus
popu-lation containing RD-114 and the D56(S+L-) strain ofMo-SV replicating in a canine kidney cell (31;
unpublished data). The portions of theMoloney leu-kemia virusgenome represented in the Mo-MuLV
cDNAand the fractionsthereofaredetailed below.
Isolation of viral and total cellular RNA and cellular DNA. Total cellular RNA wasisolated by
the method of Glison et al. using cesium chloride centrifugation (15). Concentrations of RNAwere
de-terminedby their absorbanciesat260rn,and puri-fied RNAsolutionswerestoredat-20 C. 32P-labeled
60-70S MoloneytypeC viral RNAwasisolated by
velocity sedimentation in linear 10to30% sucrose
gradients. Cellular DNAwasprepared essentially asdescribed (10).
Hybridization procedures. The procedures for DNA:RNA and DNA:DNAhybridization have been described and are provided in the legends to the figures and tables herein. Hybridization was
ana-lyzed by S1 nuclease digestion or cesium sulfate
centrifugation as previously described (8). Cesium
sulfatewaspurchased from Henley Co., New York.
AllCrt values have beencalculated by the method of Birnstieletal. (9) and correctedto0.18M
monova-lentsalt concentration (10).
Cot
valueswerecalcu-latedasdescribed (10).
Fractionation of [3H]DNA probestoproduce
spe-cific DNA fragments. The fractionation of DNA transcriptshas been describedinarecent publica-tion (36)and is similartoprocedures also used for the preparationof cDNA's fromRoussarcomavirus
(D.Stehelin, J. M. Bishop, and H. Varmus,,personal
TABLE 1. Virus strains and cells employed
Virus Relevantproperties Sourceand/or
refer-ence Producer cells
Mo-MuLV/NIH NIH 3T3 cell line producingMoloney leukemia virus 2, 21 Mo-MuLV/NRK NRK cell line producing Moloney leukemia virus 36, 12 X-MuLV/Sirc Rabbit cornea cell line producing a xenotropic type C 7, 11
virusfrom BALB/c 3T3 cell line
G-MuLV/Sc-1 (N-tropic) Wildmouse cell line(Sc-i)producing Gross MuLV with 17; J. Hartley,
N-tropic host range personal
com-munication
G-MuLV/Sc-i
(B-tropic)Sc-i
producing Gross MuLV with a B-tropic host rangeV-NRK NRK cell line producingendogenous rat type C viruses 37,11, 12
Nonproducer cells
D56(S+L-) NRK NRK cell line transformed by D56(S+L-) strain of 1, 5, 6, 12
Mo-SV
Mo-NRKp- NRK cell line transformed byHT-1 strain of Mo-SV 20, 12
Ha-NRK NRK cell line transformed byHa-SV 38, 12
Ab-NRK NRK cell line transformed byAbelson virus 36, 12
Ki-Mink Mink cell line (CCL 64) transformed by Kirsten sar- 22, 19 comavirus
Ann-1 NIH 3T3 cell line transformed by Abelson virus 34, 21
Ha-MDCK Caninekidney cell line transformed by Ha-SV Unpublished data
D56(S+L-)MDCK Canine kidney cell line transformed by D56 strain of 30
Mo-SV
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.503.72.466.423.655.2]communication). The general method is detailed herein and specific details are given in the appropri-atelegends. [3H]cDNAtranscripts werehybridized tototal cellular RNA from the variousnonproducer cells to aCrt of5 x 103 to 1 x 104 mol/s per liter. The hybridization reactions were then processed by hy-droxylapatite chromatography in the following manner. A
water-jacketed
hydroxylapatite column wasprepared with a bed volume of 1 g of hydroxyl-apatite resin permilligram of RNA in the hybridiza-tionreaction. Thehybridization mixture was loaded onto acolumn at 24 C in buffercontaining 0.025 M sodiumphosphate (pH 6.8)-0.6 M NaCl. After wash-ing with 5 to 10 column volumes of the starting buffer, the temperature was raised to 64 C and the column was washed with asecond buffercontaining 0.14 Msodium phosphate, pH 6.8, with 0.6 M NaCl. Thesecond buffer wash was monitored by radioac-tivity measurements on fractions eluted from the column with thiswash until less than 1% of the peak of radioactivity was eluted. The remaining radioac-tivity waseluted from the column with 0.46 M so-diumphosphate (pH 6.8)-0.6 M NaCl. The cDNA in the 0.14 and 0.46 M sodium phosphate washes was dialyzed for2to 3 hagainst 0.01 M Tris-hydrochlo-ride, pH 7.2, treated with 0.5 N NaOH for 5 h at37 C, dialyzed against 0.01 M Tris-hydrochloride, pH 7.2,andlyophilized prior to use.
Further purification of the fractionated cDNA transcripts was carried out to select for the cDNA portionsthat formed more stable hybrids. Hydrox-ylapatite-cycled cDNA transcripts were hybridized to saturating levels of appropriate nonproducer total cell RNA and, at the completion of the reac-tion,the reaction mixture was then treated with S1 nuclease to digest unhybridized cDNA. After S1
digestion, the hybridization reaction was extracted
with a 1:1 mixture of redistilled phenol (buffered with 0.1 M Tris-hydrochloride, pH 8.0) and
chloro-form-isoamyl alcohol (24:1). After hydrolysis, the
deproteinized hybrid solution was dialyzed to
re-move phenol and was again treated with 0.5 N NaOH for 5 h at 37 C. After alkali hydrol-ysis, the cDNA fractions were dialyzed exten-sively against 0.01 M Tris-hydrochloride, pH 7.2,
and lyophilized prior to use. These steps were
as-sociated with some loss of cDNA and recoveries are given below.
As a third fractionation procedure, particularly forthe final purification of thesarcoma virus-spe-cific sequences containedinMoloney sarcoma virus, cesiumsulfate densitygradient centrifugation was
employed. The sarcoma virus-specific cDNA
ob-tained by procedures previously described using hydroxylapatitechromatography washybridizedto
the RNA of a NRK nonproducer cell
nonproduc-tively transformed by a strain ofMo-SV isolated from the HT-1 hamster cell (20) and referred to herein as the HT-1 strain ofMo-SV, or Mo-SV p-(36) (see Table 1). After S1 nucleasedigestionofthe
probe, as detailedabove, theprobe wasstill found
tocontainapproximately 5to 10%ofsequences
ho-mologous to Mo-MuLV.To remove theseMo-MuLV sequences, the cDNAwashybridized to Mo-MuLV 60-70S viralRNAandfractionated intohybridized
and nonhybridized regions by cesium sulfate
den-sity gradient centrifugation. Details of the frac-tionation are given in theresults. After purification in cesium sulfate, thesarcoma-specific cDNA was dialyzed andlyophilized prior to use.
Immunological assays.Immunoassays of the pro-teins from Moloneyleukemia virus were performed by double-antibody competition radioimmunoas-says. The major viral glycoprotein, gp7O, was iso-lated using a slight modification of the procedures of Strand and August (42). Briefly, purified, con-centrated viruspreparations (approximately 25 mg) were made1% in Triton X-100 in 0.2 M KC1 contain-ing 0.01 MTris-hydrochloride, pH 7.8, with 0.001 M EDTA. After 30 minofincubation at 24 C with occa-sional shaking, the solubilized virus was centri-fuged at 104,000 xg for 1 h. The supernatant was removed, extracted three times with 10 volumes of diethyl ether to remove the detergent. The extracted solubilized viral protein was then dialyzed against two changes of 0.01 M BES (Sigma Chemical Co., St. Louis, Mo.), pH 6.5, with 0.001 M EDTA. After dialysis at 4 C, the virus was applied to a column (1 by 15 cm) ofphosphocellulose p-11 (Whatman)
equilibratedin0.01 MBES, pH 6.5. Following
appli-cationof the sample at4C, the column waswashed until no detectablematerial at an absorbance at 280 nm was detected. A linear 200-ml gradient of 0 to
0.5 MKClinBES was thenapplied; 2-ml fractions were collected and monitored by conductivity, ab-sorbance at 280 nm, and, where indicated, by
so-diumdodecylsulfate-polyacrylamide gel
electropho-resis. A periodic acid-Schiff-staining polypeptide
with amolecular weight of 70,000 elutedat 0.15M
KCl. A polypeptide(s) with comparable staining
andmolecularweight characteristicswasalso noted in the phosphocellulose flow-through fractions at
concentrations4to 10 timeshigherthan thateluted
at0.15MKCl.Onlyasingle bandwasnotedeluting
at0.15MKClincontrasttothe doublebandtermed gp69,71byStrand and August (42).
Thegp7Owas furtherpurified bySephadexG-150
chromatography. Briefly, fractions from
phospho-cellulose chromatography were
pooled, dialyzed
against 0.01 M
Tris-hydrochloride, pH
7.6, con-centrated 10-fold by lyophilization, anddialyzed
again. This material
(approximately
0.5 mg) wasappliedto acolumn (3.0 by 90cm)
equilibrated
in0.01 M Tris-hydrochloride, pH 7.6, with 0.001 M
EDTA, and run at 16 ml/h. Fractions were moni-tored by sodium dodecyl
sulfate-polyacrylamide
gels;fractionscontaining
gp7O
and free of otherde-tectablepolypeptidebandswere
pooled,
lyophilized,
and used forimmunological assays. The
Sephadex
G-150-purified
gp7O
migrates as apredominantly
70,000polypeptide, althoughsome
lower-molecular-weight bandsarenoted
(unpublished
data). Isolatedgp7O, iodinated with chloramineT, wasprecipitable with immune but not with control
se-rum and demonstrated
predominantly
group- andtype-specific reactivities. Thedetails ofthe
immu-noassayproceduresasgiveninthe
legend
toFig. 1 aresimilartothosepreviously
described fromthislaboratory (29). Theprocedures for the
immunoas-sayof theMoloney
p30
andtheMoloneylow-molecu-lar-weightpolypeptides
(p10,12
andp15)
have beendescribed in detail in a previous
publication
(29).on November 10, 2019 by guest
http://jvi.asm.org/
494 PARKS ET AL.
In some cases, precipitate counts were determined inan LKB gamma counter equipped with a beryl-lium crystal, which gives a 70% efficiency for 125I determinations. A single pool (IS-166) ofpolyvalent goat anti-Mo-MuLV supplied by Roger Wilsnack, Baltimore, Md., wasused forall assays at the fol-lowing concentrations: gp70, 1:1,600; p30, 1:6,400; p15, 1:200; andp10,12, 1:400.
RESULTS
Viral polypeptide expression in
nonpro-ducer cells. By appropriate isolation
proce-dures it is possible
toinfect and transformnon-mouse cells
with
different transforming viral
genomes
derived from the Moloney
leukemiavirus (35). It is thus
possible
toevaluate mousetype
C viral
polypeptide expression
insuch
cells inthe absence of expression of endogenous
mouse type
C virion
polypeptides
(28).
Initially,
a
series of cells infected
witheither Mo-MuLV orthe
replication-defective
transforming
viralgenomes was
assayed
for the differentMoloney
polypeptides. The results
areshown
inFig.
1.As shown
inFig. 1A, a gp7O was detected
at alevel of
5 to 10gg
of
cellular protein
in NRKcells infected with Mo-MuLV; complete
compe-tition
with the labeled gp7O from
the
Mo-MuLV-infected NKR cells
wasobtained
at 100 [image:4.503.269.459.54.339.2]lig
of protein.
D56(S+L-)
NRK,
evenat
100ug
of protein,
was notfound
to containdetectable
Mo-MuLV
gp7O. However, since D56(S+L-)
NRK cells contain
10to 20times
less viral RNA
and other viral proteins
(Table
2)
than
Mo-MuLV
inNRK cells, the
D56(S+L-)
canine cell,
which
has p30
levels comparable to Mo-MuLV,
was
also used
to assaygp7O expression. When
cell
extractsfrom the
D56(S+L-)
canine
cellwere
used
ascompeting protein,
nosignificant
displacement
of
labeled
gp7O
viral
polypeptide
was
noted,
even atconcentrations
of
protein
that exceeded those required for detection of
gp7O
inproducer levels by 50-fold. Also,
nosignificant
displacement
wasnoted with
Mo-NRK p-,
Ab-NRK,
or inNRK
cells
producing
endogenous
ratviruses,
V/NRK.
Ha-NRK (not
shown)
wasalso negative.
Addition of
purified
gp7O
tonegative
extractsresulted
incomplete
displacement
curves, suggesting that
the
nega-tiveresults
were notdue
tointerfering
sub-stances
(unpublished data).
We,therefore,
con-clude that
noneof
these four strains of
trans-forming
virusexpresses significant amounts of
Mo-MuLV
gp7O
innonproductively
trans-formed
heterologous cells.
In
Fig. 1B,
p30
competition
immunoassays
with
the
same extracts areshown
in anassay
which
specifically
reactswith
mousetype C
p30
and
will not measure rattype C
p30.
As haspreviously
beenreported,
D56(S+L-) NRK (not
R90
~90
90
50-10D.
001 0.01 01 0 0 00
MICROGRAMS PROTEIN PER ASSAY
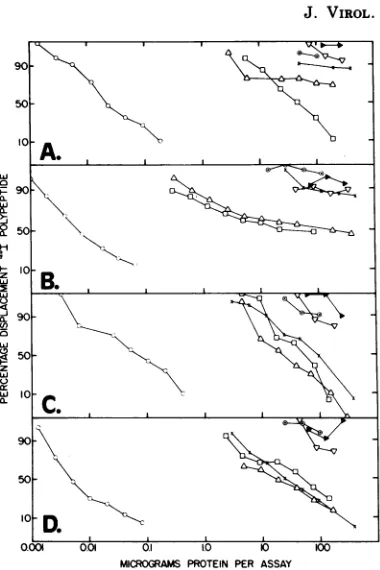
FIG. 1. Competition radioimmunoassay analysis of Mo-MuLV polypeptides in producer and nonpro-ducer cell extracts. Extracts (20%, vollvol) of
Mo-MuLVINRK(LO), D56(S+L-) MCDK (A), Mo-NRK
p- (V), Ab-NRK (x), uninfected NRK (-), and MCDK (0D) were prepared for immunoassay by scraping cells from monolayer cultures, pelleting by centrifugationat 1,000 x g for 5min,and solubiliza-tion in 0.1 MNaCl-0.01 M Tris-hydrochloride, pH 7.8, containing 1.0% Triton X-100. Protein concen-tration was determined by the method ofLowry et al. (24) using bovine serum albumin as a standard. Antigen extracts were tested indouble-antibody
imn-munoassay reactions as previously described (28), except that in the assays forp10O,12andp15 approxi-mately 50,000 counts per assay were employed and precipitate counts after one wash in 0.1 M NaCl-0.01 M Tris-hydrochloride, pH 7.8, with 0.5% normal rabbit serum were measured as describedin the text. In A through D, displacement curves with purified unlabeled polypeptides (0) are shown to indicate assaysensitivity. (A) gp7O. The specific activity of labeled antigen was 6,uCi of
12C'I/pg
of protein. (B) p30. (C) p15, Moloney. (D) p10,12, Moloney. The specific activity of the latter three labeled polypep-tides waspreviously reported.shown) by complement fixation and
radioim-munoassays contain detectable levels of MuLV
p30 (1). In contrast, no detectable murine p30
could be detected in either Mo-NRK
p-,
Ha-NRK cells, or Ab-Ha-NRK. The absence of p30 was not previously reported in Abelson
on November 10, 2019 by guest
http://jvi.asm.org/
DELETION MAPPING OF MOLONEY TYPE C VIRUS 495
TABLE 2. Comparison of levels of Moloney leukemia virus RNA and polypeptides in producer and
nonproducer NRK
ceilsa
Cellstudied RNA levels Immunoreactiveproteins'
1/2
Crt Finaltentex-plo,
12 p15 p30 gp7OMo-MuLV/NRK 23 100 625 580 2,400 360
D56(S+L-)NRK 200 44 75 90 75 <20
Mo-NRKp- 27 28 <5 <5 <2 <20
Ab-NRK 160 36 550 500 <2 <20
Ha-NRK 18 16 <5 <5 <2 <20
a For RNAand protein levels in nonproducer cells, each hybridization reaction was incubated at 66 C and
contained in 0.05 ml: 0.02 MTris-hydrochloride,pH7.2, 0.6 MNaCl,0.05%sodium dodecyl sulfate, 5 x 10-5 EDTA, and approximately 3,000trichloroaceticacidcounts of [3H]cDNA perminfrom Mo-MuLV/NRK. The final extent of the reaction was at a
Crt
of104 mol/s per liter. The100% value with the Mo-MuLV/NRK RNA was approximately 2,600counts/min. Reactions were analyzed withS1nuclease.bThe levelsof immunoreactive proteins expressed as nanograms per milligram of total cellular protein weredetermined as described in the legend to Fig. 1.
ducers
because heretofore heterologous
nonpro-ducer
cells
have
not
been available, an
impor-tant
prerequisite since
normal
mousecells
con-tain a
significant basal level of immunoreactive
p30 that can be measured by
radioimmunoas-say
(28).
Results
of competition immunoassays, for
p10,12 and p15
areshown
in Fig.iD and
C,respectively. Importantly, Ab-NRK reacts at
protein levels
comparable
to cells
infected with
and
producing Mo-MuLV. In other studies
notshown,
the genetic
stability
of
this expression
has
been
demonstrated by showing that NIH
cells also transformed by the same strain of
Abelson
virus, Ann-1 (34)
(provided by C.
Scher, Boston, Mass.), had
similar
expression
of the
type-specific
low-molecular-weight
Molo-ney
polypeptides,
whereas Ha-SV-transformed
NIH
3T3 cells were negative.
D56(S+L-)
canine
cells alsoexpress
bothp10,12 and p15,
consistent
with
earlier studies
which this genome has been
reported
tocontain
sequences
for
p12
and
p30 expression
(43).
Thus, it is clear that Abelson virus and D56
(S+L-)
Mo-SV both express the
Moloney
low-molecular-weight polypeptides
but
that
Abel-son
virus-transformed
nonproducer
cells fail
toproduce immunoreactive
p30.
No
detectable
re-action in
either the
p1O,12
orthe
p15
assay
wereobserved
with either
Ha-NRK
orMo-NRK
p-,
indicating that they do
notcontain
significant
levels of
gp7O,
p30, p10,12,
orp15.
Correlation
of levelsof
Moloney
viruspoly-peptides and
Moloney
viral-specific
RNA indifferent NRK cell.
To further evaluate the
patterns of
polypeptide
expression
inNRK cells
infected
with thevariousMo-SV
strains,
cellu-lar RNA from the transformed
nonproducer
cells and from cells
producing
Moloney
leuke-mia virus were
examined for levels of
viral-specific Moloney RNA. The kinetics of reaction
between the cell RNA and
acDNA prepared
from
Moloney leukemia virus
aresummarized
in
Table 2. NRK
cells
producing
Moloney
leu-kemia virus had
a1/2
Crt of
23mol/s per
liter,
avalue that was almost identical
tothe 1/2 Crt
levels
inthe
Ha-NRK cells
and the Mo-NRK
p-cells. Thus,
inthe
caseof
the Mo-NRK
paand
Ha-SV NRK-transformed
cells,
eventhough
noMoloney structural proteins are detected
insuch cells in the
competition
radioimmunoas-says,
the concentrations of RNA
homologous
toMoloney
leukemia virus
arecomparable
tothe
RNA
found
inthe NRK cells
producing
Molo-ney
leukemia virus. The difference noted is
inthe final
extentof the reaction with the
Molo-ney
cDNA
probe
and
notthe concentrations of
RNA
homologous
tothe
Moloney
leukemia
ge-nome. In
fact, the
D56(S+L-)
NRKand
Abel-son
NRK cells had somewhat lower absolute
amounts
of the
Moloney
leukemia
virus-specific
RNA,
eventhough
the viral
polypeptides
dis-cussed
inthe
preceding
section
werereadily
demonstrated.
Thus,
the
inability
todetect
anyviral structural
proteins
orthevarying pattern
of expression
seen invarious
transformed
non-producer
cells does
notcorrelate
simply
with
adecrease in the absolute levels ofRNA
homolo-gous
toMoloney
leukemia virus. These
results,
however,
do
notexclude
translational
interfer-ence
with
polypeptide synthesis.
Fractionation of DNA
representing
variousparts
of the Mo-MuLV genome. To further
assess the differing polypeptide patterns
ob-served
inthe
nonproducer
transformed cells
and
todetermine whether the
apparent
absence
of
polypeptide
expression
incertain
Mo-SV
strains was
due
totranscriptional
ortransla-tional
controls,
Mo-MuLV cDNA
wasfraction-ated
by
hybridizing
itwith the RNA contained
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.57.449.88.177.2]in
the
NRK cell transformed by the
D56(S+L-)
strain
of Mo-SV (fraction
2A,
Table3). The
portion
that hybridized
to theS+L--positive
RNA
wasfurther
hybridized
a second time tothe
sameRNA and
was treated withS1
nu-clease,
asdetailed
above. Thisconstitutes,
asshown in
Table
3,fraction
2B of theMo-MuLV
genome.
This fraction (2B) of
theMo-MuLV
cDNA
which did hybridize
to the S+L- genome wasfurther
subfractionated into
aportion
thatwascontained in the Mo-NRK
p-cells andaportion
that
was notcontained
inthe
RNA ofthose
cells.
This fractionation
wasperformed with
hydroxylapatite
chromatography,
and
these
portions
representfractions
4 and 5 oftheMo-MuLV
genome(Table 3).
In
addition, fraction
6 wasprepared
by
hy-bridizing the whole Mo-MuLV
cDNAdirectly
tothe Mo-NRK
p-RNA and
treating
the reactionwith
S1 nuclease.
Thehybridization properties
of
the six fractions
of the Mo-MuLV cDNAaresummarized
inTable
4. Allhybridization
re-sults indicate the final
extentof thehybridiza-tion
reactionatCrt
values
of 104mol/sperliterand
arenormalized
to100%,
based on theex-tent
of
hybridization
withMo-MuLV.
Fraction
2B
hybridized
well
tothe
RNAs from all of the
NRK nonproducer cells
transformed by the
dif-ferent
strains oftransforming virus.
In contrastto
whole
Moloney cDNA (fraction 1), the final
extent
of
the reaction with each
nonproducer
RNA
is
higher with fraction 2B relative
tothe
final extent with
the
RNA fromMo-MuLV
NRK
producer cells. As controls, the final
ex-tentof reaction
relative
toMo-MuLV
wascom-parable with
the RNA of either
axenotropic
MuLV
virusgrowing
in
Sirc
cells
or anN-tropic
or
B-tropic
strain of
Gross virus
growing
in
SC-1cells.
Fraction
3,
which
represents
aportion
of
Mo-MuLV
genomenotcontained in the
S+L-
NRK
RNA, hybridizes
appreciably
tothe
Mo-NRK
p-RNA.
Fractions
4
and
5,
which
represent the
further
subfractionation
of
fraction
2B,
also
re-veal
someimportant
differences
notseenwith
either the
Mo-MuLV fraction 1
or2B
probes.
The results with fraction 4 indicate that this
portion
of
the
Mo-MuLV
genomeis
homologous
toboth the S+L- D56 and
virtually
equally
homologous
tothe
Mo-NRK
pa
RNA. With
frac-tion
4,
ahigher
final
extentof
hybridization
with each
nonproducer
RNA
relative
tothat
with
the
whole
Mo-MuLV
probe
is
detected in
the
Harvey
NRK and Abelson NRK cells.
Equally important,
the
Ki-Mink
nonproducer
cell
(Table 1) gives
the
highest
final
extentof
hybridization
with this
specific portion
of the
Mo-MuLV
genome.Fraction
5,
which
representsthe portion of
the
Mo-MuLV
genomethat is contained in
S+L- NRK but
notcontained
inMo-NRK
p-,reveals
animportant finding
that
correlates
well with the
patternof
protein expression
pre-viously
shown in
Table
2.Fraction
5hybridizes
appreciably
tothe
Abelson-transformed NRK
RNA but
not toeither the
Mo-NRK
p- orHarvey
NRK RNA. As
acontrol
for
fraction
4,
fraction
-5failed
tohybridize
tothe Ki-Mink
cellular
RNA. As
afurther control,
fraction
6,
which
representsthe
cDNA
fragment
homolo-gous to
Mo-NRK
p-RNA, gives
a patternof
hybridization
similar
tothe results with
frac-tion 3
cycled cDNA probes. The results
indicate
TABLE
3.
Summary ofpurification of
Mo-MuLV
cDNA
Fractional
Trichloroa-probedesig- Properties of cDNA Method ofpurification cetic acid
mint
nation
counts/min
1 Homologoustowhole Mo-MuLV 10 x 106 100
2A HomologoustoD56(S+L-)NRK Fraction 1 cDNA + D56 2.6 x 106 26
RNA -.
hydroxylapa-tite,0.46 Msalt wash
2B Homologousto
D56(S+L-)
NRK Fraction 2A + D56 RNA, 1.3 x 106 13S1 nuclease resistant
3 NothomologoustoD56(S+L-)NRK Fraction 1 cDNA + D56 2.4 x 106 24
RNA -+
hydroxylapa-tite,0.14 Msaltwash
4 cDNAcommon toD56(S+L-)NRK and Fraction 2B + Mo-NRK 0.2 x 106 2
Mo-NRKp- p- RNA--
hydroxylap-atite, 0.46 M salt wash
5 cDNAhomologoustoD56(S+L-)NRK, Fraction 2B + Mo-NRK 0.3 x 106 3
nothomologoustoMo-NRKp- p- RNA-. hydroxylap-atite, 0.14 M saltwash
6 HomologoustoMo-NRKp- Fraction 1 cDNA + Mo- 1.7 x 106 17
NRK p- RNA, S1
nu-clease resistant
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.60.454.457.656.2]DELETION VIRUS 497
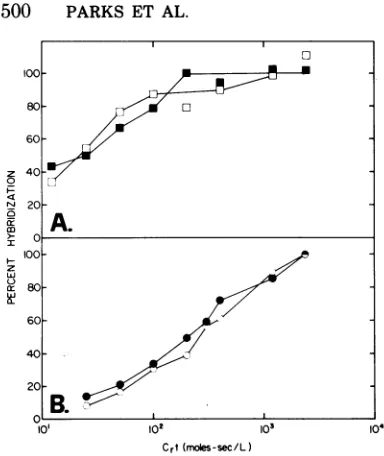
TABLE 4. Hybridization with different fractions ofMo-MuLVcDNA withcellularRNAsa
Finalextents of hybridization of fraction: RNAsource
1 2B 3 4 5 6
Mo-MuLV/NRK 100 100 100 100 100 100
D56(S+L-)NRK 44 100 6 97 91 61
Mo-NRK p- 28 40 22 94 10 91
Ha-NRK 16 26 4 49 5 52
Ab-NRK 26 52 3 67 31
X-MuLV/Sirc 40 40 41 29 36
G-MuLV/Sc-1 (N-tropic) 38 38 39 25 30
G-MuLV/Sc-1 (B-tropic) 32 38 38 25 28
Ki-Mink 5 10 3 19 3
V/NRK 7 3 3 5 3
Mink,uninfected 2 3 3 3 3
aForhybridization of cDNA fractions to total cellular RNAs, reactions were carried out in a volume of
0.05 ml at 66 C to a Crt value of 104 mol/s per liter with an input of approximately 1,000 to 3,000 trichloroacetic acid counts of cDNA per min and 250 ,ug of total cellular RNA. The conditions of hybridiza-tion are asdescribed in footnoteatoTable 2. Mo/MuLV NRK total cellular RNAhybridized maximally to all the cDNA's tested and therefore was considered 100%; all other values are normalized to this. The actual final extents ofhybridization were as follows: whole Mo-MuLV cDNA hybridized66%;S+L--positive cDNA fraction 2Bhybridized 74%; S+L--negative cDNA fraction 3 hybridized 53%; S+L--positive, Mo-NRK-positive cDNA fraction 4 hybridized 76%; S+L--positive, Mo-NRK-negative cDNA fraction 5 hybridized 68%; Mo-NRK p- fraction 6 cDNA hybridized 78%. Each value represents the average of three determinations that reached the final extentof hybridization. The average standard deviation is 5% of the value given (range, 1to10%). Blanksindicate values not determined.
that,
by the fractionation
procedure outlined,
the Mo-MuLV
cDNA
canbe
purified
into
atleast three distinct
nonoverlapping
portions
(fractions 3, 4,
5)and that
hybridization with
these fractions reveals various
typesof RNA
intransformed
nonproducer
cells that
cannotbe
detected
with
the whole Mo-MuLV cDNA
probe.
Percentage
of Mo-MuLV
genomerepre-sented
by
different
portions
of the
cDNA.
To
evaluate what
proportion
of the Mo-MuLV
ge-nome was
represented
by
the various cDNAfractions,
anexperiment
wasperformed
inwhich
increasing
amountsof
the
fractionated
[3H]cDNA's
werehybridized
tolimiting
amounts
of
32P-labeled
60-70S Mo-MuLV viral
RNA. The
results
areshown
inFig.
2.With
increasing
quantities
of the
[3H]cDNA
fraction
1, up to 57%
of the
input 32p
RNA
waspro-tected. The shape of the
curveindicates that the
whole
Mo-MuLV
genome is notrepresented
uniformly
in the cDNA transcript. The 57%hybridization
valuewasachieved withatwo-tothreefold
molar excess of cDNA. Whenfrac-tions 2B
and
3werehybridized
totheMo-MuLV60-70S
RNA,
each
fractionclearly
represented
less of the
Mo-MuLV
genomethan the
starting
fraction
1.At
approximately
atwofold
excessineach
case,about
25%of the
genome waspro-tected
fromnuclease
digestion.
Again,
when
fractions
4and
5werehybridized
in asimilar
fashion,
a two- tothreefold
excessprotected
roughly
15 to 18%of
the
32Pviral RNA. Because
of limited quantities of the
fractionated
[3H]cDNA
probes
and the fact that only
a two-tothreefold molar
excesscould be
added, final
extents
of
hybridization could
notbe
achieved.
However,
the results
areconsistent
with the
results
obtained with
[3H]DNA
and
excessRNA experiments cited above
and
indicate that
the
differing cDNA fractions
representsmaller
fractions
of Mo-MuLV
genomethan
repre-sented
inthe whole cDNA
probe.
Relative
concentrations of
sarcoma-specific
and
MuLV-specific
RNA in
transformed
non-producer
cells. To
assessthe
relative levels of
MuLV-specific and
sarcoma-specific RNA
in twostrains
of
Mo-SV,
it wasfirst
necessarytofurther
purify
the
sarcoma-specific
cDNA (36)
to
free
itcompletely
of MuLV
sequences.To
do
this,
the
sarcoma-specific
DNA (as
outlined
above)
washybridized
toMo-MuLV
60-70S
RNA
and the
hybridization
reaction
wasfrac-tionated
in cesiumsulfate
asshown
inFig.
3.A
small
peak
of
hybridized
cDNA
wasobtained
inthe region of the
gradient
withabuoyant
den-sity of
approximately
1.62g/cm3.
Fractions 16to22were
pooled
toobtain
a"sarc"
cDNA
homolo-gousto
Moloney
sarcomavirus, which
wascom-pletely free of Mo-MuLV
sequences.This
sar-coma-specific cDNA and
aMo-MuLV cDNA
(fraction
1,Table
3) werehybridized
either
tocellular
RNA ofD56(S+L-)
NRKcellsortothe
RNA of Mo-NRK
p-cells,
and the results
wereon November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.46.444.80.225.2]4~u
C
0.5 10
ngcDNA
FIG. 2. 32p protection studies with fractionated
cDNA probes. Each 0.05-mi reaction mixture was
incubated at 66 C for 36 h andcontained 250
trichlo-roaceticacid counts Of32p~60-70S RNA permmnwith a specific activity of 5 x 105 trichioroacetic acid
counts/mmnper pg. Eachreaction mixture contained ionic components as indicated in footnote a toTabke2 and was hybridized to increasing amounts of
f3H]cDNA fractionated as indicated in Table 3 and
4. The specific activity of the wholestarting DNA
probe was 2 x 107trichioroacetic acidcounts/mmnper pg. Thus, for an input of 250 trichioroacetic acid
counts of32p~per min, one would expect 10,000 3H counts to protectall of the 32p counts if the cDNA
transcriptcontained a 1:1 distribution of cDNA.
Hy-bridization was analyzed with the use of Si nuclease
aspreviously described (8). Symbols: *, whole Mo-MuLV cDNA; U,fraction 2B; O.fraction 3; *, frac-tion 4; A, fraction 5; fraction 6 (not shown) was
similar to hybridization pattern noted with fractions
4and 5.
analyzed by RNA association kinetics. The re-suits are shown in Fig. 4. In Fig. 4A, the reasso-ciation of the Mo-NRKpa RNA is shown with the sarcoma-specific cDNA and the Mo-MuLV cDNA. The results indicate that the rate of hybridization with each cDNA was equal with the two cDNA fractions; the 1/2 Crt for each probe was approximately 3 x 102 mol! s per liter. In Fig. 4B, the similar experiment was performed with the D56 S+L- strain of NRK RNA. The rate of association was slower with this cellular RNA (as shown in Table 2) but again was identical with both cDNA probes; the 1/2 Crt value with each probe was approxi-mately2 x 102 mol/s per liter. Thus, in both the Mo-NRK pa RNA and the S+L- NRK RNA, although the absolute values for the viral RNA differ in the two cells, the kinetics of the reac-tion for the MuLV portion and the
sarcoma-specific portion
areidentical in each
casefor the
twosarcomavirus
strains.
DNA:DNA
hybridization
withpurified
cDNA
fractions.
Employing fractions 1,
2B,
and
3,
itwaspossible
todirectly
testwhether
athigh
Cot
values
heterologous nonproducer cells
contained
asmuch
Mo-MuLV DNA
asproducer
cultures. The results shown
inTable
5indicate
that both
fraction
1and
2Bhybridize
toboth
Mo-MuLV/NRK
DNA and
D56(S+L-) NRK
DNA. In
contrast,fraction
3hybridizes
toMo-MuLV NRK DNA but
nottoD56(S+L-) NRK
DNA.
Thus, the
samepatternof results
isob-tained
with DNA:
[3H]DNA studies
asnoted
inRNA:
[3H]DNA studies. Thus, the results with
these
twofractions of the Mo-MuLV probe
areconsistent with the
hypothesis
that
S+L-
Mo-SVrepresentsadeletionmutantof part of
the
Mo-MuLV
genome.Expression of MuLV cDNA fractions
in sar-comavirus
nonproducer cells
superinfected
with
heterologous helper viruses. To
assesswith
yetanother
experimental
approach
whether the
differing
patternsof RNA
expres-sion
seenin
sarcomavirus-transformed
nonpro-ducer cells
representedin
parttranscriptional
blocks
toexpression, various
sarcomavirus-transformed nonproducer
cells weresuperin-fected with
heterologous helper
typeC
viruses
torescue
the
sarcomaviruses contained inthe
cells.
The
RNA from such aproductively
in-fected cell
wasthen tested
against the different
portions of the Mo-MuLV
cDNA's
to seewhether
sequences would beexpressed
inthe
rescued
genome notcontained in
the
nonpro-ducer cell.
Correspondingly,
proteins ofthe
Mo-MuLV virus
werealso analyzed
inthe
nonpro-ducer cell superinfected
withthe
heterologous
helper
viruses to seewhether other
proteinswould also be expressed. In each case, FT+
viruses
wereisolated and the
ratio ofFT+
virusto
helper
wasdetermined.
RD114-pseudotyped
FTviruses
had a 1:1 ratio or afourfold FT+
virus excess.
The
Wo-LV-pseudotyped
FT+vi-ruses
had
ahelpertoFT+ virusexcessof five-totenfold
(unpublished data). The results of the
hybridization
andpolypeptide
analysis fromthese cells
are shown inTable
6. The resultsindicate that the
final
extent ofhybridization
with the
various
cDNA's does
notchange
innonproducer cells
after infection withheterolo-gous
helper
virusin thepresenceof
theheterol-ogous
helper
virus.Similarly,
the proteinsde-tected by
radioimmunoassay
areidentical
tothose
profiles detected
inthe nonproducer cells,asindicated in
Fig.
1.The
heterologous
virusesused in
these
experimentsdonot cross-reactinthe nucleic
acid
hybridization
orimmunoassaysused.
The
results indicate
that the absence ofI
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.503.68.264.56.263.2]600-
1.6~
E
14
4 8 12 16 20 24 28
Bottom FRACTION NUMBER Top
FIG. 3. Cesium sulfatepurification of the Mo-SV-specific[3H]DNA.Partially purified Moloney sarcoma-specific [3H]DNA (200,000 trichloroacetic acidcounts/min) washybridized with 60-70S Moloney leukemia virusRNA to aCrtof4.3x101mol/perliter usingreaction conditions described previously (36). The hybrids wereseparated from the sarcoma-specific [3H]DNA byCs2SO4density gradient centrifugation in polyallomer tubes (8). One-half of the reaction mixture was mixed with 11.0 ml of
CsSO4
(p =1.52g/cm3)
(Gallard-Schlesinger,New York) in 0.01 MTris-hydrochloride(pH 7.2),0.001 M EDTA, and 0.01%(vol/vol)sodium
laurylsarcosinate. Thegradients were centrifuged in a Beckman 65 angle rotor for 72 h at 15 C at 44,000
rpm. Gradients were collected by puncturing the bottom of the tube and collecting 20drops/fraction. The buoyant density was calculatedfromtherefractive index of every
fifth
fraction. Recovery ofcounts per minute wasgreater than 95%.expression
of the RNA
inthe various
nonpro-ducer cells is
notdue
to ablock
inthe
transcrip-tion
of part of the MuLV genome
contained
inthe
transformed cell.
DISCUSSION
The
availability
of three different
fibroblast
transforming
(FT+)
viruses
derived
from
Mo-MuLV
provides
the
opportunity
for
ananalysis
of both the
sarcoma-specific
sequences
associ-ated with fibroblast
transformation
aswell
asthe sequences
responsible
for
polypeptide
expression.
We
recently
compared
the
sarcoma-specific
sequences
of
Mo-SV,
Ha-SV,
and
Abel-son
virus
(36),
and the
currentstudies
wereundertaken
tocorrelate Mo-MuLV nucleic
acid
sequences
and the
polypeptides
expressed
incells
transformed
by
three
different FT+
vi-ruses.
The
different FT+
viruseshave
been
found
toproduce
different Mo-MuLV
polypep-tides
and toexpress
differentportions of
theMo-MuLV
genome
inassociation with their
re-spective
transforming
functions.
Fromacom-bined
analysis
of the
expression
of
polypeptides
and nucleic acids
inthe
different
FT+viruses,
certain
facts have
emerged. First,
replication-defective,
transforming
virusesproduce
neither
Mo-MuLV gp70,
asshown
herein,
nor,
asprevi-ously
shown,
viral
reversetranscriptase
(23,
31).
Thus,
the
expression of these
twoproteins
would seem
tobe
least
closely
linked
tothe
expression
of
transforming functions
and/or
tohave
beendeleted. Second, the
p10,12
and
p15
can
be
expressed
inthe absence
orpresence
of
p3O
inthe
different
FT+
isolates.
Thus,
expres-sion
of
low-molecular-weight
polypeptides
ap-pears
tobe
moreclosely
linked
toexpression
of
the
transforming
functions than
p30
and
moredistantly
linked
tothe
expression
of the
gp7O
and
reversetranscriptase. These results with
the
low-molecular-weight
proteins
areconsist-ent
with the data
on variouswoolly monkey
sarcoma
virus-transformed
nonproducers
re-cently
reported by
Aaronson
etal. (3).
Third,
in
some
strains
of
FTP+
viruses
(Mo-NRK
p-
and
Ha-SV),
noneof the known structural
proteins
of Mo-MuLV is
produced.
Because
of the differing patterns of
polypep-tide
expression,
weanalyzed the transformed
nonproducer cells for
RNAhomologous
toMo-MuLV
and,
in correlation with theprotein
data, noted that different portions
of the Mo-MuLVgenome
wereexpressed
in thesecells.
Thus,
wewere abletoprepare
cDNAfractions
by
hybridizing
the whole Mo-MuLVcDNA
with the RNA from the cells
transformed by
499
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.503.102.393.50.253.2]10 a
z
a:)
m
z
6 4
10
6
4
2
lo' I0z
Crt(mfdes-sec/L)
10'
FIG. 4. Hybridization ofwhole Mo-MuLV cDNA andpurifiedsarccDNA to Mo-NRK-p-and
S+L--NRKtotal cellular RNA. Thereactionswerecarried outinavolumeof0.05 ml withaninputof
approxi-mately 4,000trichloroaceticacid countsofMo-MuLV cDNAperminor1,000trichloroaceticacid countsof
cDNAsarcperminand 50pgoftotal cellularRNA. The conditionsof hybridization areasdescribed in thetext.(A) HybridizationtoMo-NRKp-total
cellu-larRNAwith wholeMo-MuLV cDNA(E)andcycled
"sarc" A cDNA(U).At thefinalextentof hybridiza-tionthe Mo-MuLV cDNAhybridizedto1,150counts! minand the cDNA "sarc"hybridizedto875 counts! min. These valuesaretakenas100%forcalculation ofCrt value. (B) Hybridization to S+L--NRK total cellularRNAwith whole Mo-MuLV cDNA(0)and
cycled sarc "A" DNA (O). At the final extent of hybridization, the Mo-MuLV cDNA hybridized to 2,000trichloroacetic acidcounts/minand the cDNA
sarc hybridized to 878 trichloroacetic acid counts! min.These valuesaretakenas100%forcalculation ofCt values.
different FT+ viruses.
Hybridization
with theseprobes
provided
several interestingobserva-tions. First,
a portion of the murine type C genome ispresent in all thereplication-defec-tive murine FT+ viruses,evenwhennomurine
typeC polypeptidesareproducedinsuch cells.
This
commonregionseemstobecloselylinkedtothe
transforming
functions andrepresentsasaminimum estimateatleast15%of theMuLV
genome. The
results
suggest that therecom-binationeventleadingtothe formation of FT+
viruses
is occurringatacommonregionof themurinetypeC viralgenome, even
though
thisevent
has ledtotheacquisition ofheterologous
sarcoma-specific
sequencesderived
fromratsor mice.Secondly,
anexcellent correlationwasfoundbetween the final
extentsof
Mo-MuLV RNA
expressed
and the number of
structuralpoly-peptides
produced.For example,
Abelson-transformed
cells contained RNAsequencesnot present in Mo-NRK p- cell RNA butfewer
RNA
sequencesthan
werecontained in D56
(S+L-)
NRK cells.Thiscorrelated well
withthe
fact that Ab-NRK contained
low-molecular-weight MuLV proteins, Mo-NRK
p-contained
no
MuLV
structuralproteins,
and S+L-NRK
contained
low-molecular-weight
proteinsplus
MuLV
p30.
These results suggest that thecDNAprobes prepared
by
cycling againstdif-ferent nonproducer RNAs represent at
least
partof thesequences
coding
forp30 and/or thelow-molecular-weight
proteins.Thus,
infuturestudies it should be possible to compare the
cross-reaction between the cDNApresumed to
encode for p30 from other mammalian type
C
viruses.
Itshould also
now bepossible
toex-plore
the genetic basis for the apparentgroup-specific
reactivities of thep30
and thetype-specific
immunoreactivity
of thelow-molecular-weight proteins (29, 43) by comparison with other murine type C viruses.
The
thirdobservation
from these studies isthat the patterns of RNAor
protein expression
didnot
change
inthe thetransformednonpro-ducer cells, evenwhen
they
weresuperinfectedwith
heterologous helper
type C virusestopro-duce infectious
sarcomavirus.These
data,
cou-pled
tothe observations of Maiseletal.(25, 26)
that the
replication-defective
Ha-SV andMo-SV contain asmallergenomethan
Mo-MuLV,
indicate that these F+ viruses are deletion
mutantsand that the RNA andprotein
expres-sion patterns noted are due to
differing
dele-tions of the Mo-MuLVgenomeand not to
tran-TABLE 5. Hybridizationwithdifferent fractions of
Mo-MuLV cDNAwithcellular DNAa Finalextentsofhybridization
CellDNAsource Fraction Fraction Fraction
1 2B 3
Mo-MuLVNRK 100 100 100
D56(S+L-) NRK 20 29 <4
V/NRK <1 <1 <1
a For DNA:DNA reassociation, reactions were
carriedoutinavolume of0.05ml for100hat66Cto a Cot value of >2.4 x 104 mol/s perliterwith an
input of approximately 1,500 trichloroacetic acid
countsperminute of cDNA and 100,ug ofcellular
DNA. Final extent values arenormalized to 100% basedon Mo-MuLV/NRK. The actual final extent ofhybridizationwith Mo-MuLV/NRKwith fraction
1 was 45%, with fraction 2B was 44%, and with fraction3was24%.
zc
-,
io 0 )0
30
30
0 .
,-
-0-2,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.503.73.265.50.278.2] [image:10.503.276.464.486.556.2]DELETION MAPPING OF MOLONEY TYPE C VIRUS 501
TABLE 6. RNA and protein expression in nonproducer cellssuperinfectedwithheterologousviruses
Proteins'
Cell Fraction 2B Fraction 3 Fraction 5
p10,12 p30
Mo-NRK p- 40 22 10 <10 <2
+ Wo-LV 42 23 12 <10 <2
Ha-MDCK 26 4 5 <10 <2
+ RD-114 26 4 5 <10 <2
D56(S+L-)MDCK 100 6 91 2,600 4,000
+ RD-114 100 5 92 3,200 4,500
Ab-NRK 52 3 31 3,000 <2
+ Wo-LV 48 3 33 3,200 <2
Mo-MuLV/NRK 100 100 100 3,500 5,250
aHybridization reactions were
carried
out to a finalCAt
value of 104mol/s
per liter as described in the footnote to Table 4. Wo-LV indicates a type C virus isolated from a woolly monkey (45).b The levels of immunoreactive proteins expressed as nanograms per milligram of total cellular protein weredetermined as described in the legend to Fig. 1.
1 D56(S'L-)Mo-SV |
2 Mo-NRKpA
3. HarveyMo-SV _
4.Abelson Mo-SV ..xxxX ..XXXXXXXXX...X.. I "JNT" pl0,12p30
p15
FIG. 5. Proposedsequenceof different
Mo-MuLV-derivedfibroblast transforming viruses. The differ-entsarcoma-specific sequences ofD56(S+L-)Mo-SV
(-) and Harvey Mo-SV (000) are identified by
letter symbols. The postulated transforming se-quencesof Abelson Mo-SV (xxx) arealso shown. Indicatedlengths ofgenomesshownareapproximate although there is evidence that. Ha-SV and D56(S+L-) Mo-SV contain smaller RNA subunits than Mo-MuLV (26; unpublished data). For com-parative purposes, the gene sequence ofRous sar-comavirusproposed by Wongetal.(44) ispresented.
scriptional
or translational blocks.Thus,
theprotein
and
RNAexpression
patternsnoted
earlier
canbe
transposed
to apartial
genetic
map as
shown
inFig.
5. Fromourpresent dataan internal insertion of the "sarc" sequences
cannot be
excluded; further,
it is notpossible
totranspose the order of
the
defectivesarcomagenomestothe whole Mo-MuLV untilmore is
known
about
thespecific
mechanism ofrecom-bination involved in sarcomavirus formation.
However, Fig. 5summarizes earlier discussion
points and emphasizes: (i) the common joint
region, (ii) the relative
linkage
of the MuLVsequences with respect to transforming
func-tions, and (iii) the
heterogeneity
of the sarcsequences.
No
attempt is madeto order thesefunctions withrespect to either the 5' or 3' end
of
the
virus, as has beenelegantly
donere-cently by Wang et al. for avian sarcoma
vi-ruses (44).
However, if
the
mechanism
of formation ofall
fibroblast-transforming
isolates isthe sameand
ifthe
geneorder is similar for mammalianand avian
viruses,
then the 5'-3' order ofthelinear RNA genome
S+L-
would be5',
p30,p12,
p15, sarc, common-3'.
For
the Abelson andHT-1 strains
of Mo-MSV
and
Ha-SV,
increaseddeletions from the circular proviral
DNAwouldresult
inthe
sarcsequencesbecoming
closertothe
5'end of the
linear RNA sarcoma virusgenome. Since translation
begins
at the 5' endof
mRNA, the relevant
implications
of thismodel
isthat
invitro translation of thedefec-tive
sarcomavirus genomes would favortrans-lation
of the
transforming
protein.
ACKNOWLEDGMENT
This workwassupported byacontractfrom the Virus
CancerProgram.
LITERATURE CITED
1. Aaronson, S.A.,R.Bassin,and C. Weaver. 1972.
Com-parison of murinesarcoma viruses in nonproducer andS+L--transformed cells.J. Virol. 9:701-704. 2.Aaronson, S.A.,and W. P. Rowe. 1970. Nonproducer
clones of murinesarcomavirustransformedBalb/3T3 cells.Virology42:9-19.
3. Aaronson,S.A.,J. R.Stephenson,S.Hino,and S.R.
Tronick.1975.Differentialexpressionofhelperviral
structuralpolypeptidesincells transformedbyclonal
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.503.41.235.259.441.2]isolates of woolly monkey sarcomavirus. J. Virol. 16:1117-1123.
4. Abelson, H. T., and L. S. Rabstein. 1970.
Lymphosar-coma: virus-induced thymus-independent disease in
mice. Cancer Res. 30:2213-2222.
5. Bassin, R. H., L.Phillips, M. J. Dramer,D. K. Haa-pala, and P. T. Peebles. 1971. Transformation of
mouse3T3 cells by murinesarcomavirus: release of virus-like particles in the absence of replicating
mu-rineleukemia helper virus. Proc. Natl. Acad. Sci. U.S.A. 68:1520-1524.
6. Bassin, R. H., N.Tuttle, and P. Fischinger. 1970. Isola-tion of murine sarcoma virus transformed mouse
cells whicharenegative for leukemiavirus fromagar
suspensionculture. Int. J. Cancer 6:95-107.
7. Benveniste, R. E., M. M. Lieber, andG. J. Todaro.
1974.A distinct class of inducible murine type-C
vi-ruses which replicate in the rabbit SIRC cell line. Proc. Natl. Acad. Sci. U.S.A. 71:602-606.
8. Benveniste, R. E., and E. M. Scolnick. 1973. RNA in mammaliansarcomavirustransformed non-producer cells homologous to murine leukemia virus RNA. Virology 51:370-387.
9 Birnstiel, M. L., B. H. Sells, and I. F. Purdom. 1972. Kinetic complexity of RNA molecules. J. Mol. Biol. 63:21-39.
10. Britten, R. J., D. E. Graham, and B. R. Neufeld.1974.
Analysis ofrepeating DNAsequencesby reassocia-tion.Methods Enzymol. 29:363418.
11. Callahan, R., R. E.Benveniste, M. M. Lieber, and G. Todaro. 1974.Nucleic acid homology of murine
type-Cviralgenes.J. Virol. 14:1394-1403.
12. Duc-Nguyen, H., E. N. Rosenblum, and R. F. Zeigel. 1966. Persistent infection ofa rat kidney cell line withRauschermurineleukemia virus. J. Bacteriol. 92:1133-1140.
13. Finkel, M. P., B. Biskis, and P. Jinkins. 1966. Virus induction ofosteosarcomasinmice. Science 151:698-700.
14. Gardner, M. B., R. W. Rongey, P. Arnstein, J. 0.
Estes, P. Sarma, R. J. Huebner, and C. G. Rickard. 1970. Experimental transmission of feline
fibrosar-comato catsanddogs. Nature (London) 226:807-809.
15. Glison, V., R.Crkvenjakov, and C. Byers. 1974. Ribo-nucleic acid by cesium chloride centrifugation. Bio-chemistry 13:2633-2637.
16. Hartley, J. W., and W. P. Rowe. 1966. Production of altered cellfoci in tissue cultureby defective Moloney
sarcomavirusparticles. Proc. Natl.Acad. Sci. U.S.A.
55:700-786.
17. Hartley, J. W., and W. P. Rowe.1975.Clonal cell lines from a fetal mouse embryo which lack host-range restrictions for murine leukemia viruses. Virology 65:128-134.
18. Harvey, J. J. 1964. An unidentified virus whichcauses
therapid production oftumorsinmice.Nature (Lon-don) 204:1104-1105.
19. Henderson,I.C., M. M. Lieber, and G. J. Todaro.1974. Focusformation and thegeneration of"nonproducer" transformed celllines with murineand feline
sar-comaviruses.Virology 60:282-287.
20. Huebner, R. J., J. Hartley,and W. Rowe. 1966. Rescue ofthe defective genome of Moloney sarcoma virus
fromanoninfectioushamstertumorand the produc-tionofpseudotypesarcomaviruses with various
mu-rineleukemiaviruses.Proc.Natl. Acad. Sci. U.S.A.
56:1164-1169.
21. Jainchill, J. F., S. Aaronson, and G. Todaro. 1969. Murinesarcomaand leukemiaviruses:assayusing
clonal lines ofcontact-inhibitedmousecells.J.Virol.
4:549-553.
22. Kirsten, W. H.,and L. A. Mayer. 1967. Morphologic
responses to a murineerythroblastosis virus. J. Natl. Cancer Inst. 39.311-319.
23. Livingston, D. M., W. P. Parks, E. M. Scolnick, and J. Ross. 1972.Affinity chromatography of avian type C viral reverse transcriptase: studies with Rous sar-coma virus transformed rat cells. Virology 50:388-395.
24. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall. 1951. Protein measurement with the Folin phenolreagent. J.Biol. Chem. 193:265-275. 25. Maisel, J., V.Klement,M. M.-C.Lai, W. Ostertag, and
P. Duesberg. 1973. Ribonucleic acid components of murine sarcomaandleukemia viruses. Proc. Natl. Acad. Sci. U.S.A. 70:3536-3540.
26. Maisel, J., E. M. Scolnick, and P. Duesberg. 1975. Base sequencedifferences between the RNA components of Harvey sarcoma virus. J. Virol. 16:749-753. 27. Moloney, J. B. 1966. A virus-induced
rhabsomyosar-comaof mice. Natl. Cancer Inst.Monogr. 22:139-142. 28. Parks, W. P., D. M. Livingston, G. J. Todaro, R. E. Benveniste, and E. M. Scolnick. 1973. Radioimmu-noassay of mammaliantype-C viral proteins. III. De-tectioninnormal murine cells. J.Exp.Med. 137:622-635.
29. Parks, W. P., M. Noon,R. Gilden,and E. Scolnick. 1975. Serologicalstudies withlow-molecular-weight polypeptidesfrom theMoloney strain of murine leu-kemiavirus.J.Virol. 15:1385-1395.
30. Peebles, P. T., P. J. Fischinger, R. H.Bassin,and A. G. Papageorge. 1973. Isolation of human amnion cells transformedby rescuable murine sarcoma vi-rus.Nature(London) New Biol. 242:98-101. 31. Peebles,P.T.,B. I.Gerwin,A.G.Papageorge,and S.
G. Smith. 1975. Murine sarcoma virus defectiveness. Viralpolymerase expression in murine and nonmu-rinehost cellstransformedbyS+L- type murine
sar-comavirus.Virology 67:344-355.
32. Potter, M., M. Sklar, and W. P. Rowe. 1973. Rapid viral induction ofplasmacytomasinpristane-primed Balb/cmice.Science 132:592-594.
33. Rosenberg, N.,D.Baltimore,andC. D. Scher.1975.In vitro transformation oflymphoid cells byAbelson murineleukemiavirus. Proc.Natl.Acad. Sci. U.S.A. 72:1932-1936.
34. Scher,C.D., and R.Siegler. 1975.Direct transforma-tion of3T3 cells byAbelson murine leukemia viruses. Nature(London)253:729-731.
35. Scolnick,E.M., R.Goldberg,andW.Parks. 1975.A biochemical and genetic analysis of mammalian RNA-containingsarcomaviruses.ColdSpring Har-borSymp. Quant.Biol. 39:885-895.
36. Scolnick,E.M.,R.S. Howk, A. Anisowicz, P. Peebles, C. Scher.and W. Parks. 1975.Separation ofsarcoma virus-specificandleukemiavirus-specific genetic se-quencesofMoloneysarcoma virus. Proc.Natl.Acad. Sci. U.S.A. 72:4650-4654.
37. Scolnick,E.M.,J. M.Maryak,and W. P. Parks.1974.
Levelsofratcellular RNA homologous to either Kir-stensarcoma virus or rattype C virus in celllines derived fromOsborne-Mandel rats. J. Virol. 14:1435-1444.
38. Scolnick, E. M., and W. P. Parks. 1974. Harvey sar-coma virus: asecond murine type C sarcomavirus with rat geneticinformation. J. Virol. 13:1211-1219.
39. Scolnick,E.M., E. Rands, D. Williams, and W.Parks. 1973.Studies on the nucleic acid sequences of Kirsten sarcomavirus: amodelfor formation of a mammalian RNA-containingsarcoma virus. J. Virol.12:458-463.
40. Sklar, M. D.,B. J.White,and W. Rowe. 1974. Initia-tion of oncogenic transformaInitia-tion of mouse lympho-cytesinvitroby Abelson leukemia virus. Proc. Natl. Acad. Sci. U.S.A. 71:4077-4081.
on November 10, 2019 by guest
http://jvi.asm.org/
41. Snyder, S. P., and G. H. Theilin. 1969.Transmissible filinefibrosarcoma. Nature (London) 221:1074-1075. 42. Strand, M., and J. T. August. 1973. Structural proteins
ofoncogenic ribonucleic acid viruses. Interspec II, a new interspecies antigen. J. Biol. Chem.
248:5627-5633.
43. Tronick,S. R., J. R. Stephenson, and S. A. Aaronson. 1973.Immunological characterization ofalow
molec-ular weightpolypeptide of murineleukemia virus. Virology 54:199-206.
44. Wang, L., P.Duesberg, D. Beemon, and P. Vogt.1975.
Mapping RNase T,-resistant oligonucletodies of avian tumorvirus RNAs: sarcoma-specific oligonu-cleotidesare nearthepoly(A) end and
oligonucleo-tidescommontosarcomaand
transformation-defec-tive virusesareatthe poly(A) end. J. Virol.
16:1051-1070.
45. Wolfe, L. G., F. Dienhardt, G. H. Theilin, H. Rabin, T. Kawakami, and L. R. Bustad. 1971. Induction of
tumorsin marmosetmonkeys by simiansarcoma
vi-rus,typeI(lagothrix):apreliminaryreport.J.Natl. Cancer Inst. 47:1115-1120.


![FIG. hybridization pattern noted with4MuLVcounts4.countsbridizationtranscriptation wascounts/mmnionicandroaceticincubatedf3H]cDNApg](https://thumb-us.123doks.com/thumbv2/123dok_us/1558980.108508/8.503.68.264.56.263/hybridization-pattern-noted-mulvcounts-countsbridizationtranscriptation-wascounts-mmnionicandroaceticincubatedf-cdnapg.webp)
![FIG. 3.Schlesinger,specificrpm.lauryltubeswereviruswasbuoyant Cesium sulfate purification of the Mo-SV-specific [3H]DNA](https://thumb-us.123doks.com/thumbv2/123dok_us/1558980.108508/9.503.102.393.50.253/fig-schlesinger-specificrpm-lauryltubeswereviruswasbuoyant-cesium-sulfate-purification-specific.webp)