0022-538X/79/02-0716/10$02.00/0
Interruption-Deficient
Mutants of
Bacteriophage T5
I. Isolation and
General
Propertiest
STEPHEN G. ROGERS,t ELIZABETH A. GODWIN, ELIZABETH S. SHINOSKY, AND MARC RHOADES*
Departmentof Biology, The Johns Hopkins University, Baltimore, Maryland 21218
Receivedfor publication 15 August 1978
Mutants ofbacteriophage T5 were isolated which lack one or more ofthe
natural single-chain interruptions that occur in the mature DNA of this virus.
Interruption-deficient mutantswere detected by screening survivors of
hydrox-ylaminemutagenesis for altered DNA structure by electrophoresis in agarose slab
gels. Over60independentmutantswere isolated fromasurvey ofapproximately
800phageparticles.Allof themutants wereviable andcould be grouped intotwo
classes. Mutants inoneclass lackedoneof the localized siteswhere interruptions
occur in T5 DNA. To date, mutants that affect five different sites have been
obtained. Mutants inthe otherclasswereessentially free from interruptions or
hadareducedfrequency of interruptions throughoutthe genome. The members
of this class included several ambermutants. Complementation tests indicated
thatatleasttwo genes arerequired for thepresence ofinterruptions in mature
T5 DNA.
The nonpermuted genome of bacteriophage
T5 is contained within a single linear,
double-stranded DNA molecule (27). T5 DNA has a
molecular weight ofapproximately80 x 106(14,
24) and containsan 8.3% terminal redundancy
(20). T5DNA isdistinguished from the DNA of
mostothercoliphages by the presence ofasmall
number ofsingle-chain interruptions (1). These
sitescanberepairedbytheaction ofT4 DNA
ligase andarelargely confinedto onestrand of
theduplex molecule (12).Althoughthe
interrup-tions appeartooccur atfixedsites,thefrequency
ofinterruptions at the different sites is highly
variable (9, 24). The positions of the fourmost
frequent interruptions, which occur in nearly
everymolecule, have been determinedby
elec-tronmicroscopy (2, 3, 13, 23, 24) and agarose gel
electrophoresis (8, 19). These sitesoccur at
ap-proximately 8, 18, 32, and 65% from what is
arbitrarily designated asthe left end of the
ge-nome. Some of the less frequent interruptions
have also been mapped (19, 24). The highest
concentrationofthese sites occurs at the leftend
of theDNA, within the terminal repetition.
Thesignificance ofthesingle-chain
interrup-tions in T5 DNAisnotknown. Although ithas
beensuggested thatone or moreof the
interrup-tions could influence genetic recombination or
DNAinjection (1, 2),there is no directevidence
t Contribution no. 972 from the DepartmentofBiology,
TheJohnsHopkinsUniversity,Baltimore,Md.
tPresent address:Department of Microbiology, The Johns
Hopkins UniversitySchool ofMedicine,Baltimore,MD 21205.
linking these sites to any specific process. To
investigate this question, we initiated a search
formutantsof T5 that lack the normal
comple-mentofinterruptions. This searchwasaidedby
thedevelopmentofanelectrophoreticassay that
allows the genomes ofmutagenized phagetobe
rapidlyanalyzed for structural alterations. Use
of thisscreening procedure resulted in the
iso-lation of two classes of mutants with altered
patterns ofsingle-chain interruptions. Mutants
inoneclasslackedoneof the sites where
inter-ruptions normally occur. Mutants in the other
class had reduced levels of interruptions
throughoutthe genome. Some members of this
classappearedtobecompletelyfree from
inter-ruptions.
The present report describes thegeneral
prop-erties of interruption-deficient mutants of T5
and the methods used for their isolation. The
accompanying paper (21) describes the
proper-ties of a mutantthatspecificallylacks the
inter-ruption located at 8% from the left end of T5
DNA.
MATERIALS AND METHODS Bacteriaandbacteriophage. T5st(+)isthe wild-typestrain of Abelson and Thomas(1). T5st(102) isa
heat-stable deletion mutant (25). Escherichia coli B40sul, B40su2, andB40su3 wereobtained from H. Berger.E. coliF,afast-adsorbing strainforT5,was
obtained from D. Botstein. E.coli B23 wasobtained from M. Bessman. All bacteriophage weregrownby confluentlysisasdescribedbyRhoadesandRhoades (20).
716
on November 10, 2019 by guest
http://jvi.asm.org/
Isolationofmutants.T5st(102)wastreatedwith hydroxylaminefor72h at370Casdescribed by Tess-man (26) to afinalsurvival of106. During the same period, a control incubated without hydroxylamine showed a 50% loss in titer. Survivors were plated on E. coliB40sul at 30°C. Individual plaques were sus-pended in 10 mM Tris-hydrochloride (pH 8.0) and replated on E.coli B40sul to eliminate heterozygotes. Single plaques were then transferred with sterile toothpicks to 1-cm2 grids on soft-agar overlays (6) seeded with E. coliB40sul. After 6to 8 h at 30 or
370C,thesoft-agar grids were removed from theplates
with aspatula andplaced in 0.1 ml of 10 mM Tris-hydrochloride (pH 8.0). After 30 min at250C,a 0.03-mlsample of the resuspended phage was mixed with 5,Aof 2% sodiumdodecylsulfate-0.1 M EDTA (pH 9.0) and heatedfor 5 min at 60°C. Ten minutes before analysis of the DNA, 5
pl
of afreshly prepared solution of1 MNaOH was added to denature the DNA. The denaturedsamples were then subjected to electropho-resis inhorizontal 0.7% agarose slab gels as described by Rogers and Rhoades (22).Amber mutants wereobtained byplating survivors ofhydroxylaminetreatmentonthe su- strain E. coli B23. Single plaqueswerethen directly tested for in-terruption-deficient phenotypes as described above. Phage which were either completely free from inter-ruptionsorwhichexhibited reducedlevels of interrup-tionsthroughout the genome were then retested on E. coliB40sul, B40su2, and B40su3.
Procedurefor crosses. E. coli B40sul or E. coli Fwasgrown to 2x108 cells/mlinT-MGM (15) with aeration at370C. Aftercentrifugation, the cells were resuspendedat109/mlin TMB (T-MGM without glu-cose or NH4C1) containing 1 mM CaC12 and were starved for 20min at 370C with aeration. The cells were then diluted with an equal volume of TMB containing1 mMCaC12andwereinfected at a multi-plicity of 7.5 of each parent. After 20 min at 37°C without aeration, the infected cells were centrifuged to remove unadsorbedphage andweregently resus-pendedat 5x 108/mlinprewarmed T-MGM contain-ing 1 mM CaCl2.After 5 min at370C, aeration was started and continued for 60 to 90 min. Lysis was completed by addition ofCHC13.Progeny wereplated on an appropriate indicator, and the nature of the DNA of individual plaques was tested as described above.
In crosses where one parent carried the st(102) deletion mutation, the progeny were fractionated either by plating on E. coli F, where the st(102) mutation confers a small-plaque phenotype, or by banding in CsCldensity gradients. In the latter case, aportion of thelysate wasmade 45% (wt/wt) CsCl andcentrifugedin aBeckmanSW50.1rotor at25,000 rpm for16 h at40C.
Complementation analysis and dominance tests.Lysates ofmixedly infected bacteria were pre-paredasdescribedabove, except thatE.coli B23was used for all experiments involving amber mutants. Electrophoresiswascarried outdirectly on the prog-enypresentin thelysates.In somecases,particularly if oneparent carried thest(102) deletion, the progeny werefirstfractionatedby centrifugation in CsCl den-sity gradients. After dialysis against 10mM
Tris-hy-drochloride (pH8.0)-i mMMgCl2-1 mM CaC12, the CsCl-purified phage particleswere analyzed by elec-trophoresis.
RESULTS
Isolation of interruption-deficient mu-tants. The presence of single-chain
interrup-tions in T5 DNA is not known to confer any
phenotypicpropertiesuponthe virus.
Interrup-tion-deficient mutants were therefore obtained
bydirect screening ofmutagenized phage
prep-arations. To survey rapidly a large number of
potential mutants, we used a procedure that
permits theDNA presentinasingle T5 plaque
-to be analyzed, after alkaline denaturation, by
agarosegel electrophoresis. Since bacterial DNA
isefficiently degraded during the earlystagesof
T5infection (5),virtually all of the DNA inaT5
plaque consists ofphage DNA, either packaged
orunpackaged.The DNAcontentofawild-type
T5 plaque issufficientlyhigh (0.01to0.1 jug) to
permitelectrophoretic analysiswith the use of
ethidiumbromidestaining.Inpractice,however,
more satisfactory results are obtained if each
plaque isreplicaplatedtoform a 1-cm2area of
lysisasdescribed in Materials and Methods.
Mutagenesis was carried out by treating
T5st(102) phage particles with hydroxylamine
until thesurviving fractionwas 10-6. T5st(102),
aheat-stable deletionmutantthatlacks 10% of
thewild-typegenome(25), waschosento
mini-mizeanylossof viablephagenotassociated with
the action of hydroxylamine. The surviving
phage particles were analyzed by two
proce-dures. In theinitialstagesof thisinvestigation,
survivorswereplatedat
300C
onE. coli B40sulto avoid loss of conditionally lethal mutants.
Heterozygotes wereeliminatedbyreplating
in-dividualplaquesunder identical conditions.
Sev-eral plaques derived from each originalisolate
werethenanalyzedforloss ofinterruptions.
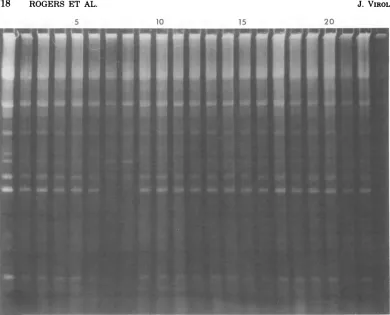
Theresults ofa typical analysisof22
prepa-rations, derived from 10 independentsurvivors
ofhydroxylaminemutagenesis,areshowninFig.
1.With theexceptionof slot23, which received
aninsufficientamount ofDNA, all of the
sam-plesexhibiteda distinctpattern ofsingle-chain
fragments.A mutantpatternwasclearlyevident
in slots 7and 8. Both of these samples, which
were derived from the same isolate, HA99,
lacked two of theprominent single-chain
frag-ments that were present in all of the other
samplesand inpurifiedT5st(102)DNA(slot 1).
Asdescribedbelow, thisparticularmutant
pat-tern indicates lossof the interruption at18.5%.
Ofapproximately 150survivorsof hydroxyla-mine treatment that were screened in this man-ner, 16 were found to contain atleastone mu-tation which caused an interruption-deficient 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
5 10 15
T v _ _
N
-
.
t..H.... s~~
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.Ai
~
.ia.
A1ll s 1 I I I |~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~F 1
FIG. 1. Exampleof isolation of interruption-deficient T5 mutants. The selection shown contained two or three descendants of isolates HA92through HA101 from hydroxylamine mutagenesis of T5st(102). Both descendants ofHA99 showed a mutant single-chainpattern in slots 7and 8. Slot 1 contained purified T5st(1 02) DNA. The other22sampleswereprepared for electrophoresis from small-platelysates as described in Materials and Methods. The directionof electrophoresis in this and all other figures is from top to bottom.
phenotype. In one instance, HA104, two
differ-entmutantphenotypessegregatedupon
replat-ing. Three different mutant phenotypes were
recovered from a second isolate, HA73, by
re-combination (see Table 1).
Interruption-deficientmutantsof T5werealso
obtained by a more rapid procedure in which
survivors of the hydroxylamine treatment
de-scribed above weretested directly, without
re-plating,for loss ofinterruptions.E. coliB23, an
su host,wasemployedin theseexperimentsin
order to allow isolation of amber
interruption-deficient mutants. Approximately 650 plaques
werescreened in thismanner, with the isolation of 45 additional mutants. The phenotypes of four of these mutants were found to be sup-pressed by growth on su+ strains.
Members of each of the categories of mutants obtained (see Table 1) were crossed with the wild type in an attempt to eliminate extraneous
mutations. Theoriginalisolates,whichcarry the
st(102) deletion,werecrossed withT5st(+),and
interruption-deficient st(+) recombinants were
obtained. The interruption-deficient mutation
was then crossed back and forth between
T5st(+) andT5st(102)for,in some cases, atotal
of five backcrosses. Wheneverpossible,any
de-tectable conditionallethal, plaque morphology,
orhostrangemutationswereeliminatedby this
procedure.However,as aresult of thehigh level
ofmutagenesisemployedinthisstudy, it islikely
that none of the strains are isogenic with the
wild type.
Electrophoreticanalysis.Thevarious
mu-tantpatternsofsingle-chain fragmentsthatwere
obtainedareshown inFig.2and3.For
conven-ience, all of the phage shown carrythe st(102)
deletion. A map showing the positions of the
single-chain interruptions and fragments that
are relevant for this study is shown in Fig. 4.
The fragments are numbered as described by
Rhoades(19).
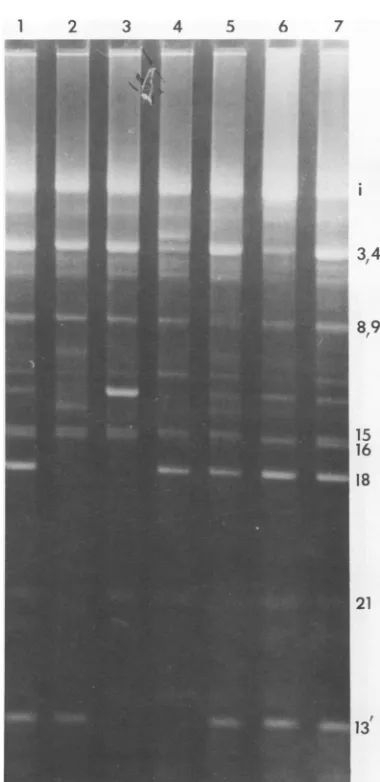
The fragment patterns of the five mutants
shown inFig.2all differedfromT5st(102)DNA in onlya few instances. The simplest
interpre-tation of each of these patterns is that they
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.68.458.66.381.2]2 3
rw
0t _4
* * * _~~~"
_ _$
4 5 6 7 is probably fragment 4 inT5HA65 (slot 4) sug-gests loss of the interruption at 32.6%. A new fragment that presumably representsthe fusion
of fragments 4 and 13' is visible just behind
fragment3.Theinterpretationof T5HA214 (slot
6) is lessclear; however,thediminished intensity
of the band where fragments 3 and 4 nonnally comigrate and the absence offragment 15 sug-gestthatthe
interruption
at64.8% has been lost.The faint band remaining at the position of
fragments 3 and 4 probably represents a
frag-3 4 mentthatextends from 45%to an interruption
1 2 3 4
e!w
---nr.-
ew 8,915 16
18
21
[image:4.505.56.246.72.463.2]13
FIG. 2. Electrophoreticpatternsof five single-site interruption-deficientmutants.All slots contained1
pg of alkali-denatured DNA from CsCl-purified phage particles.Slots 1 and7, T5st(1 02);slot2,HA4; slot3, HA626;slot4, HA65;slot5, HA73-16; slot6, HA214. The locationsofthe numberedfragmentsare
shown inFig.4. Thesymbol"i"designatesthe intact
strandofT5 DNA.
reflect loss ofoneof the siteswhereinterruptions normally occur in T5 DNA. In the case of T5HA4 (slot 2),the absence offragment 18 and
some of the smaller fragmentssuggests loss of
theinterruptionat7.9%.Moreover,severalnew
fragments which span the site of the missing
7.9% interruption arevisible (21). The absence
offragments13' and 18 in T5HA626(slot 3),and the increased intensity of fragment 7', both in-dicate loss ofthe interruption at 18.5%. Simi-larly, the absence of fragments 13', 16, and what
&-.:
I 4
18
21
13
FIG. 3. Electrophoreticpatterns of four mutants that exhibited reduced levels of interruptions throughoutthegenome.Allslots contained0.53 gof alkali-denatured DNAfromCsCl-purifiedphage
par-ticles. Slots 1 and6, T5st(102); slot2, HA23; slot3, HA104S;slot4,HA90;slot5, HA229.
5 6
29,1979
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.270.436.210.609.2]0.8 7.9 18.5 32 6
21 8 13_
I 1--
-1-22 7
I l
64.8 77.9
45.3
4
[image:5.505.111.413.69.148.2]16 9
FIG. 4. Maps of single-chaininterruptionsandsingle-chainfragments ofT5 DNA. The locationsofseven interruptions, expressedas apercentageof thewild-type length from the left end of themolecule,aregivenon the top line. The locationsof themoreprominent fragmentsof T5 DNA that define these sitesaregiven in the lower partofthefigure. Onlythe sites andfragmentsthat arerelevantforthisstudyare shown. A more detailed map is available in the accompanying paper (21). The st(102) deletion extendsfrom21.6to32.0%o. Fragments7' and 13'areshortenedversionsoffragments7and13thatoccurinT5st(+) DNA.
thatoccurswithlowfrequencyat77%(19).The
newfragment formedby the fusion of fragments
3 and 4 would not be resolved fromthe intact
strand under these conditions. Thesefour
mu-tantsthusrepresent,individually, loss of each of
the four principal single-chain interruptions in
T5 DNA.
The fifthmutantshown inFig. 2,T5HA73-16
(slot 5), was isolated by recombination from
T5HA73 (see Table 1). Although HA73-16 has
the fourprincipal interruptions, the absenceof
fragment 16 suggests loss of the interruption
thatnormallyoccurs at45%. Loss offragment9,
which would also be expected inHA73-16, was
difflculttodetect sincefragments8and9
comi-grateunder these conditions.
Additional electrophoretic studies, involving
analogous sets of duplex fragments (19), were
performed onthe fivemutants shown in Fig. 2.
In eachcase,theinterpretationgiven abovewas
confirmed. The results of this kind ofanalysis
with T5HA4 are given in the accompanying
paper (21).
The mutant patterns shown in Fig. 3reflect
reduced levels of interruptions throughout the
genome. In the case of T5HA23 (slot 2), the
absence of detectable fragments smaller than
the intact strand indicates that this genome is
virtually free from interruptions. T5HA104S
(slot 3) appears to retain some interruptions
although the overall level is greatly reduced.
The patternsof T5HA90 and T5HA229 (slots4 and 5) both suggestthat mostofthe principal
interruptions are present, but thatthe less
fre-quent secondary interruptions are either
com-pletely absent (HA90) or greatly reduced
(HA229). The most prominent feature of
T5HA229 is the increasedintensityoffragment
21.The newbands that canbeseeninT5HA90
andT5HA104Sareprobably derived from
mol-ecules that lack either orboth of the principal
interruptionsat7.9and 18.5%.
The patterns ofsuppressionof thefouramber
interruption-deficientmutantsareshown inFig.
5. Two of these mutants, T5amHA676 and
1 2 3 4 S 6 7 8 9 10 11 12
LIlUIX 4i.
FIG. 5. Electrophoretic analysis of amber inter-ruption-deficient mutants after growth on su- and su+strains. Slots1 to 4containedT5amHA676 grown on E.coliB23, B40sul, B40su2, and B40su3, respec-tively. Slots5 to8containedT5amHA911 grown on thesamehosts.Slots9 and10 containedT5amHA346 grown onB23 and B40sul. Slots 11 and 12 contained T5amHA910 grown on B23 and B40sul. All slots contained alkali-denatured DNAfromCsCl-purified phage particles.
T5amHA911,werefree frominterruptionswhen
grown on E. coli B23. After growth onE. coli
B40sul andB40su3, amHA676wassuppressed
tothe wildtype(slots2and4); however,growth
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.274.465.217.522.2]on B40su2 resulted in a pattern similar to that ofT5HA90 (see Fig. 3) in which only the prin-cipal interruptions are present (slot 3). In con-trast,amHA911 was fully suppressed only by E. coli B40su2 (slot 7). Growth on B40sul resulted in a reduced level of interruptions (similar to T5HA104S), whereas no suppression occurred on B40su3. The two other amber mutants, T5amHA346 and T5amHA910, exhibited a re-duced level ofinterruptions on E. coli B23 (slots 9and11) that resembles HA104S. Both of these mutants weresuppressed tothe wildtype onE. coli B40sul(slots10and12) and on the su2 and su3 strains (datanotshown). In addition, all of
the mutants that had a phenotype on E. coli
B40sulthat was intermediate between the wild
type and completely interruption deficient (like HA90 andHA104S) were tested on E. coli B23. None ofthepatterns werefoundtochange.
A list of the categories of
interruption-defi-cient T5 mutantsobtained in thisstudyisgiven in Table 1.
Dominancetests.Dominancetestswere
per-TABLE 1. Categoriesof interruption-deficient mutants
Mutantphenotype' Nlateidb Representativeallelesc
8- 7(2) HA4,HA73-17, HA336
18- 11 (1) HA99, HA626, HA682
32- 16(4) HA65, HA77, HA435
45- 2 (1) HA73-16, HA772
65- 5(3) HA83, HA214, HA392
Interruption-free 9(4) HA23, HA34, HA54, HA104L, HA363, HA663, amHA676, HA883, amHA911 Reducedinterrup- 9(2) HA73, HA104S,
tions amHA346, HA645,
HA771, HA776,
HA896, amHA910, HA935
Principal inter- 3(1) HA90, HA599, HA835 ruptions only
Reduced second- 1 (1) HA229 ary
interrup-tions
aThedesignations "8-," "18-," etc., refer to phage specifically lacking interruptionsat8%, 18%, etc., from the left end of T5 DNA. The last four categories describe mutantswithgenerallyreduced levels of in-terruptions throughoutthegenome,as shown inFig. 3.
b The values inparenthesesarethenumber of mu-tantsobtained inthe initialphase ofthisstudy when each isolate was replated before testing. All allele numbers below HA300aremembersof thisgroup.
cMutantsHA73-17andHA73-16wereisolatedfrom HA73 by recombination. HA104L and HA104S are segregants of isolate HA104. Ambermutants are de-notedby theprefix"am."
formed by analyzing the single-chain fragments present inthe unreplicated progeny of mixedly infected bacteria. In thecaseof themutantsthat lackedasingle interruption, mixed infection with the wild type gave bursts thatappeared to con-tain both mutant andwild-type genomes. The results obtained for mixed infection with T5st(+)HA4, which lacks the interruption at 7.9%, and T5st(102) are shown in Fig. 6. In
1 2 3 4
131
[image:6.505.277.422.182.586.2]18 ? - _ _ _ _ _
FIG. 6. Electrophoreticanalysisoftheprogenyof mixedinfectionwithT5st(+)andT5st(102)HA4. The progeny werefractionated in CsCl to separate the st(+) and st(102) phage before analysis. Slot 1, T5st(+) DNA; slot2, T5st(102) DNA; slot 3, mixed infectionprogenycarryingthest(102)deletion;slot4, T5st(102)HA4DNA. Thesymbols"a" and "b" denote two newfragments that appeared in T5st(102)HA4 DNA.
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
722
addition to lacking fragment 18, T5HA4
con-tainedseveralnewfragmentsthatdistinguish it
from thewildtype (Fig. 6,slot4). As described
in the accompanying paper (21), the new frag-ments extended between interruptions located
closetotheleft end ofgenomeandthe
interrup-tionat18.5%. Thepresence of bothmutantand
wild-typegenomes wastherefore demonstrated
by the presence of both fragment 18 and the
newfragments in the mixed infection progeny.
Similar results were obtained for T5HA99,
which lacks the interruption at 18.5%, and for
T5HA65,whichlacks the interruption at32.6%.
Analysisof themutantsthat lack interruptions
at45.3and64.8%wasmoredifficult since, under
the electrophoretic conditions employed here,
neither of these mutants possesses fragments
thatareabsent inthe wildtype.
These results indicate that loss of a single
interruption isnotduetothe absence ofa
dif-fusiblegeneproduct.Instead, itappears thatin
eachmutant oneofthe sites where interruptions
normallyoccur has beenalteredsothatthesite
isno longerrecognized by the mechanism that
specifies the interruptions.
In contrast,all of the mutations that resultin
areducedlevel ofinterruptions throughout the
genome appear to be recessive to wild type.
These tests werecarriedout onthe 21mutants
listed in Table 1 thatresemble eitherT5HA23
(no interruptions) or T5HA104S and T5HA90
(reducedlevels ofinterruptions).Inallcases,the
progenyof mixedly infected bacteriawerefound
topossessnormalpatterns ofsingle-chain
frag-ments. The results of mixed infection with
T5st(+)HA23 and T5st(102)areshownin Fig. 7.
T5HA229alsoappears tobe recessive;however,
the phenotype of this mutant is so similar to
that ofthe wild type (see Fig. 3) that an
inter-mediate result would be difficult to recognize.
Complementation analysis. The results
presented above suggest that absence ofall or
most of the single-chain interruptions in T5
DNA canbeduetothelack ofadiffusiblegene
product. To determine the number ofgenes
in-volved in this process, we carried out comple-mentation tests between various pairs of
inter-ruption-free andreduced-interruption mutants.
Theresultsof theseexperiments,which are
sum-marized inTable2, can be stated asfollows.In
all of the tests involving pairs of
interruption-free mutants, no complementation was ob-served.The progeny, like the parents, were free
frominterruptions. Severalofthe mutants that
had reduced levels of interruptions (HA346, HA645, HA771, amHA910, and HA935) also failed tocomplementtheinterruption-free mu-tants.In thesecases, theprogeny resembled the
1 2
I..
A
FIG. 7. Electrophoretic analysis oftheprogenyof
mixedinfectionwithT5st(+)HA23andT5st(102).The progeny werefractionated in CsCl to separate the st(+) andst(102)phage before analysis.Slot1, mixed-infectionprogeny of st(102) density; slot 2, mixed-infectionprogenyof st(+)density.
reduced-interruption parent.However, threeof
the reduced-interruption mutants (HA73,
HA104S, and HA896) were found to partially
complement the interruption-freemutants and
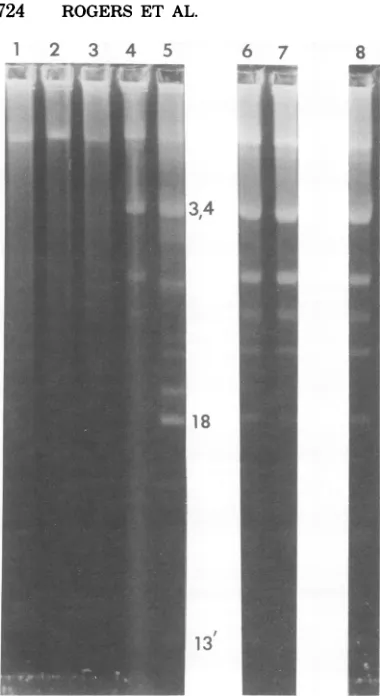
theotherreduced-interruptionmutants.The re-sults of mixed infections with T5HA104S and
several of the interruption-free mutants are
shown inFig.8.In eachcase, themixed-infection
progeny hadall of the major single-chain
frag-ments, but the minor fragments were barely
visible. These twogroups of mutants can,
there-fore, cooperate to produce progeny that have
mostbut notallof theinterruptions present in the wild type. Oneinterpretationof this result is
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.505.339.387.76.462.2]TABLE 2. Complementation analysis ofinterruption-deficientmutantsa
Allele Allele
HA23 HA34b HA363' amHA676 HA73 HA104S amHA346c HA896
HA896 +
amHA346c - +
HA104S + + + + -
-HA73 +
amHA676
-HA363C
-HA34b - _
HA23
-a+, Positivecomplementationas
shown
inFig. 8; -,nocomplementation.bResults identical to that shown for HA34 have been obtained with HA54 and HA104L.
All
pairwisetests within this group werenegative.cResults identicalto that shown for amHA346 have been obtained with HA645, HA771, amHA910, and HA935. Results identical to that shownfor HA363 have been obtained for HA663, HA883,and amHA911. Pairwise tests within these two groups have not been done.
that T5HA23 and T5HA104S define two genes whose functions are required for wild-type levels ofinterruptions. We have tentatively designated
the gene represented by HA23 as sciA
(single-chaininterruption) and thegenerepresentedby HA104S as sciB.
Otherproperties. Thegrowth properties of
T5HA23, which is free from interruptions, were
tested for any detectable differences from the
wild type. In addition to normal plaque
mor-phology, the latent period and burst size of
T5HA23 were indistinguishable from the wild
type.The absence ofinterruptions also did not
alter the survival of either T5st(+)HA23 or
T5st(102)HA23 after heatingto60°C incitrate
buffer(11).
DISCUSSION
This report describes the isolation of two
classesof viable interruption-deficient mutants
ofbacteriophage T5.In one class,the
interrup-tion deficiency appears to be limited, in each
mutant, to a single site. Since these mutations
are neither dominantnor recessive tothe wild
type, theypresumably represent alterations of
criticalsequences thatspecifytheinterruptions.
Mutants in the second class are
interruption-free or have reduced levels of interruptions
throughout the genome. These mutations are
recessivetowildtypeandappeartodefinetwo
genes, sciA andsciB,whoseproductsparticipate
informationoftheinterruptionsin T5 DNA.
Bothclassesofinterruption-deficientmutants
wereisolatedfrom a stock ofT5st(102)that had
beenheavily mutagenizedwithhydroxylamine.
Althoughthe resulting highfrequencyof
inter-ruption-deficientmutants wasadvantageousfor
this study, it is very unlikely that any of the
surviving phage were free from additional
hy-droxylamine-induced mutations. We have
at-temptedto reduce the levelof extraneous
mu-tations by backcrossing selected mutants with
the wildtype.Thest(+) and st(102) alleleswere
employedassecond markersinthese crosses to
insure that the mutant genomes were able to
recombine. Since this procedure may neverbe
completely successful, it is fortunate that
mul-tiple alleleswereobtainedformostof themutant
categories. Future studies of each
interruption-deficient phenotype will therefore not be
de-pendent upon the possibly aberrant properties
ofsinglemutants.
Theoccurrenceof both classes of
interruption-deficientmutants atnearly equal frequenciesis
of interest since thesingle-sitemutants
presum-ablyrepresentalterations ofsequencesthatare
much smaller thanagene.Hydroxylamine,
how-ever, is known to mutate preferentially
single-stranded DNA (7).Since the regions
surround-ing interruptions are more likely to be
tran-siently single stranded than other regions
(es-peciallyinaphage particle), hydroxylaminemay preferentiallymutatethese sites in T5 DNA. If correct, thishypothesis offersa possible
expla-nation for the observationthat,with one
appar-entexception (M. Rhoades, unpublished data),
no mutants were obtained with new
interrup-tions.
Informationonthesequencessurrounding the
interruptions in T5DNAhasrecentlybeen
ob-tainedbyNichols and Donelson(17,18). Astudy
of the 5' sequences, done on intactDNA, indi-catesthatpGpCpGpC-occurs atallof the
inter-ruptions and suggests that the octanucleotide
pGpCpGpCpGpGpTpG- occurs at most orallof
theprincipal interruptions.Thesequencesatthe
3' side are more diverse. Studies on isolated
single-chain fragmentshaveprovidedextensive
29,
on November 10, 2019 by guest
http://jvi.asm.org/
724 AL.
1 2 3 4 5
vL av~
-6 7
-'.-i
Pvapt
3,4
zl
;18
13
8
[image:9.505.61.251.64.412.2]VW.';
FIG. 8. Electrophoretic analysis of complementa-tiontestsbetweenT5HA104S(reduced interruptions) and several interruption-freemutants. Slot1, dena-tured T5st(102)HA54 DNA; slot 2, denatured T5st(102)HA23DNA;slot3,denaturedT5st(+)HA23 DNA;slot4,denaturedT5st(102)HA104S DNA;slot 5, denatured T5st(102) DNA; slot 6,progeny from mixed infection with T5st(102)HA104S and T5st(102)HA54; slot 7, st(102) densityprogeny from mixed infection with T5st(102)HA104S and
T5st(+)HA34; slot 8, st(102) densityprogeny from mixed infection with T5st(102)HA104S and T5st(+)HA23.
3' sequences at two sites, 8 and 18%, where interruption-deficient mutants have been ob-tained. At the 18%sitefourofthe first six base pairsonthe 3' sideareGC,whereasat8%oneof
thefirst six isaGCpair. Therearethus numer-oussites inthe immediatevicinity of thesetwo interruptions where hydroxylamine, which is specific for C, could act. Thehigher frequency ofGC pairsat 18%, however, doesnotseemto have significantly increased the frequency of mutations atthis siteascomparedto8% (Table 1). It is possible, as suggested by Nichols and
Donelson (18), that the critical 3' sequences
occurfarther fromtheinterruptions inaregion
whereoverlapping segmentswithtwofold
rota-tionalsymmetry arefound.
The existence ofviable T5mutants that are
apparently free frominterruptions raises
ques-tionsastowhetherthese sites playanessential
role in the viral life cycle. The DNA of these
mutants,however, hasonly been showntolack
interruptions inmaturevirus particles. The
in-tracellular forms of the DNAof the
interruption-deficientmutantshavenotbeenexamined. It is
possible that thedefects seenin these mutants
affect only the terminalstagesof viral
develop-ment. Therefore, atpresent, theonly firm
con-clusions are that intact T5 phage heads can
contain DNA that hasnointerruptions and that
such DNA is infective.
Theconclusion thatatleasttwogenescontrol
the formation of interruptions in mature T5
DNAwasbasedonthepartialcomplementation
observed after mixed infection with certain pairs ofmutants.Sinceafullwild-type phenotypewas never observed, these results might be due to
intragenic complementation. However, all
com-plementing pairs invariably produced patterns
similartothose shown inFig.3,including
infec-tionson su- hosts in which oneparent carried
anamber mutation. Thus, althoughthereason
for partial complementation is notknown, the
presentresults favoratwo-genehypothesis.
Theavailability ofinterruption-deficient
mu-tantsof T5should greatly facilitate further
anal-ysesof the origin and function of the
interrup-tions in T5 DNA. In particular, the single-site
mutantsshouldpermitevaluation of the role of
interruptions inprocessessuchasgenetic
recom-bination, transcription control, and DNA
trans-fer. As described in the accompanying paper
(21), T5HA4, which lacks the interruption at
8%, isnotdeficient in thetwo-steptransfer
proc-ess that is uniquetoT5. Itshould also be
pos-sibletorecombine the varioussingle-site
muta-tions to produce multiple mutants. Since the
defects in the single-site mutants appearto be
permanent, the construction of recombinants that lackallof theprincipal interruptions should
provide additional insights into the possible
functions of these sites.
Analysis of the interruption-free mutants
shouldprovidesomeinsightinto the mechanism
that generatesinterruptionsin T5 DNA. Little
is known about the nature of the T5 sciA and
sciB genes, although bacteria infected with an sciA mutant, HA23, are not deficient for four
previously isolated (22)site-specific
endonucle-asesthat are inducedbyT5 (S.Rogers,
unpub-lished data). The availability of amber
muta-tions in thesciA gene shouldpermit identifica-tion and isolaidentifica-tion of its geneproduct.Knowledge J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
of thelocation ofthescigenes onthe T5genetic map may also provide some indications as to
how, or when, theyareexpressed. Preliminary
results (B.Lange-Gustafson, unpublisheddata)
havelocated the HA23 mutationin alategene
region atthe extremeright of the nonrepeated
portion of the T5genome.None of the mutations
in the sciB gene has been mapped; however,
recombinationbetween HA23 andHA104S
ap-pears tobeinfrequent.
The T5 D15gene, anessentialearlygenethat
specifies a 5' exonuclease (10), has been
impli-cated ingeneratinginterruptions in intracellular
T5 DNA. Although D15 mutants are defective
ininitiatinglategeneexpression (4), Moyer and
Rothe(16) have obtained evidence that the D15
gene product directly participates in forming
interruptions. These authors have also
discov-ered that thepurifiedD15 proteinhas
endonu-cleolyticactivity, although it is unableto
intro-duce single-chain interruptions into duplex
DNA. Itis alsonotknown whether the breaks
generated by the D15 nucleasearethosepresent
in mature phage particles. Nevertheless, these
resultssuggestthat theprocess orprocessesthat
place interruptions into T5 DNA are complex
and that theirunderstanding willrequire
anal-ysisofanumber ofgenes.
ACKNOWLEDGMENTS
Thisresearchwassupported bygrantPCM76-24036from theNational Science Foundation. S.G.R.wassupported bya training grant (HD-139) from the National Institutes of
Health.
LITERATURE CMD
1.Abelson,J.,and C. A.Thomas,Jr.1966.Theanatomy of theT5bacteriophage DNAmolecule. J.Mol. Biol. 18:262-291.
2. Bujard,H. 1969. Location ofsingle-strand interruptions
inthe DNA ofbacteriophageT5+. Proc. Natl. Acad. Sci. U.S.A. 62:1167-1174.
3. Bujard,H., and H. E. Hendrickson. 1973. Structure
and function of the genome ofcoliphageT5. I. The
physicalstructureof the chromosome ofT5+.Eur. J. Biochem. 33:517-528.
4.Chinnadurai,G.,andD. J.McCorquodale. 1973.
Re-quirement ofa phage-induced 5'-exonuclease for the
expressionof late genesofbacteriophageT5. Proc.Natl. Acad.Sci.U.S.A.70:3502-3505.
5. Crawford,L. V.1959.Nucleic acid metabolism in Esch-erichiacoli infected withphageT5.Virology7:359-374. 6. Doermann,A.H.,andL Boehner.1970.The identifi-cation ofcomplex genotypes inbacteriophage T4. I. Methods. Genetics66:417-428.
7.Freese, E.,and H. B.Strack.1962.Induction of muta-tions intransforming DNAbyhydroxylamine. Proc. Natl. Acad.Sci. U.S.A.48:1796-1803.
8. Hayward,G.S.1974.Unique double-stranded fragments ofbacteriophage T5 DNA resulting frompreferential shear-induced breakage at nicks. Proc. Natl. Acad. Sci. U.S.A. 71:2108-2112.
9. Hayward, G. S., and M. G. Smith. 1972. The chromo-some ofbacteriophage T5. I. Analysis of the single-stranded DNAfragments by agarose gel electrophore-sis. J.Mol.Biol. 63:383-395.
10.Hendrickson, H.E., and D. J.McCorquodale. 1972. Geneticandphysiological studies of bacteriophage T5. III.Patternsofdeoxyribonucleic acid synthesis induced by mutants of T5 and the identification of genes influ-encing theappearance of phage-induced dihydrofolate reductase and deoxyribonuclease. J. Virol. 9:981-989. 11. Hertel, R., L. Marchi, and K. Muller. 1962. Density
mutants ofphage T5.Virology 18:576-581.
12. Jacquemin-Sablon, A., and C. C. Richardson. 1970. Analysis of the interruptions in bacteriophage T5 DNA. J.Mol.Biol. 47:477-493.
13.Johnston, J.V.,B. P.Nichols, and J. E. Donelson. 1977.Distributionof "minor" nicks in bacteriophage T5 DNA. J. Virol. 22:510-519.
14. Lang,D., A. R.Shaw,and D. J. McCorquodale. 1976. Molecular weights of DNA from bacteriophages T5,
T5st(0),BF23st(4). J.Virol. 17:296-297.
15.Moyer, R. W., A. S. Fu, and C. Szabo. 1972. Regulation ofbacteriophage T5 development by Col I factors. J. Virol. 9:804-812.
16. Moyer, R.W., andC.T. Rothe. 1977. Role of the T5 geneD15 nuclease in the generation of nicked bacterio-phage T5 DNA. J. Virol. 24:177-193.
17. Nichols, B. P., and J. E. Donelson. 1977. Sequence analysis of the nicks and termini of bacteriophage T5 DNA. J.Virol. 22:520-526.
18. Nichols,B.P., andJ. E. Donelson. 1977. The nucleotide sequence at the 3'-termini of three major T5 DNA
fragments.Virology83:396-403.
19.Rhoades,M. 1977.Localization of single-chain interrup-tions in bacteriophage T5 DNA. II. Electrophoretic studies. J. Virol. 23:737-750.
20.Rhoades, M.,and E. A. Rhoades. 1972.Terminal repe-tition in the DNA of bacteriophageT5.J. Mol.Biol.
69:187-200.
21. Rogers,S.G.,N. V.Hamlett, and M. Rhoades. 1979. Interruption-deficient mutants of bacteriophage T5. II.
Propertiesof a mutantlacking a specific interruption. J.Virol. 29:726-734.
22. Rogers,S.G., and M. Rhoades. 1976. Bacteriophage T5-induced endonucleases that introduce site-specific
single-chain interruptionsinduplex DNA. Proc. Natl. Acad. Sci. U.S.A. 73:1576-1580.
23. Saigo,K.1975.Denaturationmapping and chromosome structureinbacteriophageT5.Virology68:166-172.
24. Scheible,P.A.,E.A.Rhoades,and M. Rhoades. 1977. Localization ofsingle-chain interruptions in bacterio-phage T5 DNA. I. Electron microscopic studies. J. Virol. 23:725-736.
25. Scheible,P.A., andM.Rhoades. 1975.Heteroduplex
mappingof heat-resistant deletion mutants of bacterio-phage T5. J. Virol.15:1276-1280.
26.Tessman,I. 1968.Mutagenic treatmentof double- and
single-strandedDNAphagesT4and S13 with hydrox-ylamine. Virology35:330-333.
27. Thomas,C.A., Jr.,andI.Rubenstein.1964.The ar-rangements of nucleotide sequence in T2 and T5
bac-teriophageDNAmolecules.Biophys. J. 4:93-106.
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/