0022-538X/81/050593-13$02.00/0
Identification and Mapping of Two Polypeptides Encoded
Within
the Herpes Simplex Virus Type 1
Thymidine Kinase
Gene
Sequences
CHRISM. PRESTON* AND DUNCAN J. McGEOCH MedicalResearch Council Virology
Unit,
Glasgow, Gll5JR ScotlandReceived 7 November1980/Accepted 11 February 1981
mRNA's
homologous to the herpes
simplex
virus type
1 DNA restriction
endonuclease
fragment
BamHI p,
which contains the thymidine kinase gene,
have been identified
and mapped by hybrid-arrested translation and mRNA
selection. Such
mRNA's, when translated in vitro, directed the synthesis of
polypeptides of apparent molecular weights 43,000
(VI43) and 39,000
(VI39).
mRNA for
enzymatically active thymidine kinase was enriched by more than
20-fold after selection. Mapping was carried out with restriction endonuclease
fragments of BamHIp, and locations of the 5' and 3' termini of
VI43
mRNA
were
deduced.
Analysis
of nucleotide sequences around the 5' terminus revealed several
consensus
sequences
commonly found at the start of eucaryotic mRNA's and
which are
presumably involved in initiation of transcription by RNA polymerase
II. Translation of mRNA's for VI43, VI39, and the thymidine kinase enzyme was
arrested
only by a 1,170-base-pair region of BamHI p. Since this region is
insufficient
for adjacent genes, coding sequences for VI43 and VI39 must overlap;
the
possible
relationship
of these two polypeptides is discussed. A virus-induced
product equivalent to VI39 was detected in infected
cells.
An
important
method of
mapping and
puri-fying
specific mRNA's is
tocombine
nucleic acid
hybridization with in vitro
translation.
In such
experiments,
anmRNA
preparation is
hybrid-ized with a
purified fraction of genome DNA or
RNA.
Subsequent
analysis involves
either
recov-ery
from the
hybrids
and translation of selected
RNAs
ordetermination of the
polypeptides
whose
synthesis
is inhibited
(termed
"hybrid-arrested
translation").
This
approach
has
al-lowed mRNA
mapping
in
myxoviruses
(16,
20),
papovaviruses (42),
adenoviruses
(12,
22, 23,
30,
38), and
poxviruses
(8).
The
techniques
should
be
similarly applicable
toherpesvirus mRNA's,
since
herpes
simplex
virus
(HSV)-specific
RNA
can
be translated after
hybridization
toviral
DNA
(14) and mRNA's have been enriched
by
hybridization
tocellulose-bound DNA
frag-ments
(1, 2).
A
region
of the HSV type
1(HSV-1)
genome
whichcurrently
commandsspecial
interest isthe
BamHI
restrictionenzyme
fragment
BamHI
p. This
fragment,
located atapproximately
29to32
map units (46), contains the information for
expression of viral
thymidine
(pyrimidine
deox-yribonucleoside)
kinase(TK)
when used tobio-chemically
transform
TK-cells
to aTK+
phe-notype
(4, 7, 28, 29, 32, 37, 45,
47).
Furthermore,
cells transformed with BamHI p increase HSV
TK
lvels after
superinfection with TK- virus
mutants
(17, 21, 47),
showing that the
fragment
also contains sequences involved in activation
by
viral
polypeptides
(24, 35, 36).
Hybrid
plas-mids
containing
BamHI p have
recently
been
constructed, thereby
providing
a sourceof
large
amounts
of pure BamHI p
DNA,
and
detailed
restriction
maps have been
produced
(7, 10, 47).
The limits
of sequences necessary for biological
activity have been defined by the demonstration
that
cleavage
with
EcoRI, BglII
and
SstI abolish
transformation,
whereas
a2-kilobase
pair
(kbp)
Pvull fragment
retains full
activity
(7, 47).
Anal-ysis of
intertypic
recombinants has located TK
coding
regions
to 0.300 to 0.309genome
map
units
(13), and the direction of
transcription
of
TK
mRNA has also been determined
(41).
With this
background,
studies
toinvestigate
the
arrangement of mRNA
coding
regions within
BamHI p
wereinitiated.
Apart
from its
biologi-cal
relevance,
thisfragment appeared
tobesuit-able
for such anapproach
because
enzymatically
active
TK can besynthesized
in vitro(9, 34)
andthe
putative
TKpolypeptide, VI43,
isthe
mostabundant
product
upon translation
of RNAex-tracted from cells at
early
times of infection(33,
35).
We have beenable
toidentify
andsubstan-tially
purify
mRNA's
homologous
toBamHI
p,
to
map within fine limits the
coding
sequences
593
on November 10, 2019 by guest
http://jvi.asm.org/
of
TKmRNA,
andtoobtain nucleotidesequencedata for the controlregionsaroundthemRNA 5' terminus. The existence oftwo
polypeptides,
encodedby overlapping
sequences within the TK gene, has also been established.MATERIALS AND METHODS RNA isolation.
garly
RNA was extracted from BHK C13 cellsat 5hafter infectionat31°C with 20 PFU of HSV-1 strain17per cell(33).Late RNAwasobtained from BS-C-1 cellsat 6h afterinfection at
370C.
Plasmid DNA. A stock of Escherichia coli HB101 containingplasmid pTK1 (HSV-1
BamHIp
fragmentinserted into pAT153[44])wasinitiallyprovidedby
P. Sanders. PlasmidpTK6wasobtainedbycleavage
ofpTK1withBgllI, followed bypartial digestionwith BamHI. Theproduct, consistingofpAT153plusthe smallBgJI-BamHIfragment (see Fig. 10),was reli-gated and transfected into E. coliHB101.The struc-tureof theplasmidin theresulting ampicillin-resistant
colonieswasconfirmedbyrestriction enzymeanalysis,
andonecolonywaspurified and usedasthesourceof pTK6 DNA.
Restriction endonucleasemappingofpTK1.A number of approaches, all based on recleavage of known DNAfragments,wereused torefine existing
mapsofpTK1.(SeeFig. 4, 7, and10for therestriction enzymesites usedtogeneratethe DNAfragmentsfor this study.) Fragments are described here by their approximate coordinates, inkilobasepairs, from the BamHI site upstream from TK mRNA (41). The correctedpositions of AluI sitesdefiningfragmentsA, E, and F differ from previousresults (47).
Isolation of DNA from polyacrylamide gels. Gels ofvarious polyacrylamide concentrations (5 to
10%) were cast and run in 50 mMTris-borate (pH 8.3)-1.25 mM EDTA. After staining with ethidium bromide and visualization underlong-waveUVlight,
therelevant bandswerecut outandhomogenized in
10ml of TE buffer (20 mM Tris-hydrochloride [pH 7.5]and2mMEDTA). The macerated gelfragments
weregently mixed for10 to16 hat4°C,polyacrylamide
wasremoved bycentrifugation, and thesupernatant
was passed through a glass-fiber filter disk. Sodium phosphate buffer (pH 6.8) wasadded to 0.1 M and DNA adsorbed to hydroxylapatite. After thorough washingat60°C with0.2Msodiumphosphate buffer, DNAwaseluted with 0.5 M sodium phosphate buffer.
E. coli rRNA(20,ug)wasadded, and the eluate was dialyzed against TNE buffer (TE buffer plus 0.1 M
NaCI)andprecipitatedinethanol.
Portions of isolatedDNAfragments were electro-phoresed, and negatives of theUV-illuminated,
ethid-iumbromide-stained bands were prepared. The nega-tives werescanned, using aJoyce-Loebl
microdensi-tometer, to assess the purity and concentrations of
DNA fragments. None ofthefragments used in the experiments reported here contained detectable con-taminationby others.
DNA sequencing. DNA sequences around the 5' terminus of the TK genewere determined by using thechemical degradation system of Maxam and Gil-bert(26, 27), and also by cloning fragments into phage
M13derivedvectors(11)andcopying the recombinant
single-stranded phageDNA in the presence ofspecific
chain terminators(39, 40;F.Sanger,A. R.Coulson, B. G.Barrell,A. J. H.Smith,and B. A.Roe, J. Mol.Biol.,
inpress).
For the chemicaldegradationsystem,pTK1 DNA
wascleaved with EcoRIorwithBglll,labeledatthe 5'termini withpolynucleotidekinase and
[y-32P]ATP,
andthen cleaved withHinfl. Theresulting single end-labeledfragments wereseparatedbyelectrophoresis
in polyacrylamide gels, and the appropriate bands
were recovered. All sequencing gels contained 9 M
urea.
EcoRI-cleaved fragments of pTK1 DNA were
cloned into the EcoRI site of the phage vector M13mp2am and propagated in E. coli JM101 (11).
Sequencing was carried out as described by Sangeret
al. (J. Mol. Biol., in press) with the short universal
primerofAndersonetal.(3).
Hybridarrest.Plasmid DNAs were cleaved with EcoRI, extracted with phenol, andprecipitated with ethanol. Twomicrograms of restricted plasmidDNA,
orthe molarequivalent amount of a particular isolated
fragment, wasdenatured in 85
pl
of 95%formamide (deionized) containing 60 mM PIPES [piperazine-N,N-bis(2-ethanesulfonic acid)] (pH 7.4) at 95°C for 5
min. HSV-1-infected cell cytoplasmic RNA (6
pl;
8mg/ml)wasadded, followed by addition of 8
p1
of 5 MNaCl,and thehybridization mixture wasrapidly trans-ferredtoa58°Cbath. Afterincubation for 1 h, 1 ml of HSB (500 mM KCl; 20 mM Tris-hydrochloride [pH 7.5]; 1 mM EDTA) was added. Polyadenylic acid-containing RNAwasselectedon asmall oligodeoxy-thymidylic acid-cellulose column and precipitated withethanol afteraddition of E. coli rRNA.Pelleted RNAwasdried, dissolved in deionized water, and used for in vitrotranslation either directly or after denatur-ationby heatingat95°C for 2min.
mRNAselection.Thetechnique of Ricciardi et al. (38) was modified for selection of pTK1-specific mRNA's. A nitrocellulose filter (13-mm diameter; Schleicher and Schuell, Dassel, West Germany), loaded with 100ytgofplasmidDNA,wascutinto small pieces and incubated for2hat550C withearly or late cytoplasmic RNA, dissolved in hybridization buffer (40%o formamide [deionized], 600mM NaCl, 50 mM PIPES [pH 7.4], and1mMEDTA). The hybridization mixturewasthen removed andprecipitated after ad-dition of 1 volume of water and 2.5volumes of ethanol
toyield thenonhybridized fraction. The filter pieces were incubated at550C for 10 min inhybridization buffer, then washed 10timesat60°C with SSC (0.15 M NaCl and0.015Msodiumcitrate) containing 0.5% sodiumdodecylsulfate, followed by2washes at600C withSSC containing2mMEDTA. The pieces were
then washed for two 5-min periods in SSC-2 mM EDTA andoncewith colddeionized water.A
100-pl
amount of 95%formamide-50 mM PIPES (pH 7.4)wasadded,andincubationwascontinued at 60°C for
5 min. The formamide fraction was stored, and the filter pieces were incubated with 300 ,l of deionized waterfor2min at95°C. This fraction wascombined withthe 95% formamide fraction, E. colirRNA (15
,ug)
and NaCl (0.2 M) were added, and RNA wasprecipitated after addition of2.5 volumes of ethanol
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
(selectedRNA fraction). The RNA was washed with ethanol,dried, dissolved in water, and used for in vitro translation. In someexperiments, the selected RNA fraction was repurified on a small oligodeoxythymi-dylicacid-cellulosecolumn, as describedabove.
In vitro protein synthesis. RNA samples were translated in a micrococcal nuclease-treated fraction-atedreticulocyte system (31, 33, 35). In vitro-synthe-sized TK enzyme wasassayed as described previously
(34).
Labeling of infected cell polypeptides. BHK
celiswere pulse-labeled at 37°C for 15 min in phos-phate-buffered saline containing 500uCiof [3S]meth-ionine (The Radiochemical Centre, Amersham, Eng-land) per ml.Cell monolayers were then washed with ice-cold phosphate-bufferedsaline,harvested, and pre-pared for analysis by sodium dodecyl sulfate-poly-acrylamide gel electrophoresis (25).
Polypeptide nomenclature. As described previ-ously (25, 33) polypeptides synthesized in vitro and in vivo areclassified according to their apparent weights in thousands and areprefixed with VI or Vmw, re-spectively.
Biologicalcontainment. Allmanipulations with bacteriacontaining plasmid pTK1 or pTK6 or phage M13mp2am were conducted in a category II labora-tory, asrecommendedby GMAG.
RESULTS
Selection of mRNA's which
hybridize
topTKI
DNA. mRNA's
wereselected
by
incuba-tionof late RNA from infected BS-C-1 cellswith
nitrocellulose
filtersloaded with
plasmid DNA.
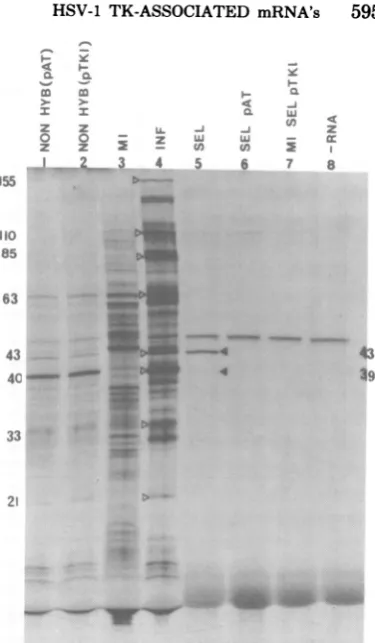
After the filters were thoroughly washed, hy-bridized RNA was eluted and translated. Fig. 1 shows that thisprocedure selected mRNA's
cod-ing for
amajor (VI43) and minor
(VI39) species
from
the array of late virus-inducedpolypep-tides.
These mRNA's were not selected frominfected cell RNA
by pAT153
DNA filters orfrom uninfected
cell RNAby pTK1 DNA,
inwhich
casesonly endogenous
reticulocyte bands
were
observed. The
specificity of the
procedure
was
further demonstrated
by
thefacts that V143was not
the
mostabundantly represented
spe-cies intranslation
products.
of theoriginal
RNAand VI39
wasundetetable before
mRNAselec-tion.
To
investigate whether mRNA for active TK
enzyme was
also
enriched
by
hybridization
topTK1
DNA, RNA from BHK
cellsat5h after
infection
at31°C,
atime when TK mRNA levelsare
maximal,
wasused. Selected andnonhybri-dized
RNAs wereexamined for
theability
tostimulate
incorporation
of14C-amino
acid or todirect
synthesis
of TK.Fig.
2(A
andB)
showsthat
selected
RNAwasless active instimulating
protein
synthesis,
but hadmarginally
greaterTK mRNA content.The measured
enrichment
of TK mRNA(TK
activity
per unit of amino acidincorporation
of selected
RNAcompared
_ _
a _
0 0
z s
F-C')
i
'U
- 'U
A A K. a 7
4c
z
FIG. 1. Selection of pTKI-specific mRNA.
Mock-infected (MI)orlateinfected(INF) BS-C-I cellRNA
washybridized tofilters containingpTKI DNA or
pAT153 DNA. Both nonselected and selected mRNA preparations were translated in vitro. RNA prepa-rationsused were the following: INFcellRNA not selected by pAT153 DNA (track 1), INFcellRNAnot
selectedbypTKIDNA(track 2),originalMIBS-C-I cellRNA (track 3), originalINFcellRNA(track4), INFcellRNA selected(SEL) bypTKI DNA (track 5),INF
cell
RNA selectedby pATi
53DNA(track 6), MI cellRNA selectedbypTKIDNA(track7), andnoadded RNA (track 8). Forreference, the molecular weights (in thousands) ofsomeprominent virus-in-ducedpolypeptidespresentareshown in track4(<J).
Selectedpolypeptides arelabeled in track 5
(4).
Amore thorough categorization oflate virus-induced
polypeptides synthesizedin vitro isgivenelsewhere
(32,34).
with
nonhybridized RNA) varied from
20- to50-fold in
repeated experiments.
An accuratevalueis
difficult
toobtain because thestimulation
of"C-amino
acidincorporation by
selected RNA was sosmall.
Translation of relevant RNA fractions
(Fig.
2C) revealed
that thepolypeptides
directedby
selected mRNA were
VI43
andVI39,
as found with late RNA(Fig. 1).
Thegreater abundanceof
VI43 and
VI39
mRNA wasreflected
in aon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.500.256.444.63.385.2]596
PRESTON ANDB
ProteinSynthesis
0 1 2 3 4
C
IL
. 2 A I
RNA(pl)
FIG. 2. SelectionofTKmRNA. Earlyinfected (INF)BHKcellRNAwashybridizedtofilters containing pTK1 DNA. The original (0), nonselected (U), and selected (0) fractions were translated in vitro, and
productswereanalyzedeitherbyassayofTKenzyme(A)orincorporation of
["4Clamino
acidmixture intoprotein (B).All RNApreparationswereat 1.2mg/ml,thisbeingachievedbyadditionofE. colirRNA to the selected RNApreparation. (C) Autoradiogram of ['5S]methionine-labeled polypeptidesdirectedbythe three
RNApreparations, togetherwithmock-infected (MI)BHK cell RNA andnoadded RNA.
greateryieldofselectedmRNA,asindicatedby thehigher intensities of these polypeptides, as compared with the majorendogenousband (mo-lecularweight,ca.50,000).
The selection experiments, therefore, show that themajor mRNA encoded by BamHIpat
both early and late times of infection specifies VI43. Another polypeptide,
VI39,
was alsosyn-thesized, and TK mRNAwasenriched, strongly suggesting that TK activity resides in either VI43orVI39.
Hybrid-arrested translation with pTK1 DNA.AnalternativewayofmappingmRNA's
is by hybrid-arrested translation (16, 30). This methodcanbe used withsmallDNAfragments and might give more precise locations than mRNA selection, as it has been reported that DNAfragments shorter than300bp donotbind reproduciblytonitrocellulose filters (38). Figure
3 (left) shows translation of early RNA after incubation with pTK1 or pAT153 DNA under conditions which favor annealing of RNA to
DNA. With both plasmids, three polypeptide bands
(VI136, VI129,
andVI88)
disappeared afterhybridization, andone(VI85) was synthe-sized in reduced amounts. These returned, tosomeextent,after denaturation ofhybrids, but clearly their arrest cannot be attributed to
BamHIp sequences. The effect is presumably due tosomeunusual structural featuresofthe mRNA's, sinceitcanalsobeobtainedby mock-hybridization without addition of DNA (data
notshown).
Hybridization with pTK1 DNA,ascompared with that of pAT153 DNA, additionally pre-ventedsynthesis of VI43 and
VI39.
mRNA's for these polypeptides regained activity uponde-naturation ofhybrids, thusdemonstrating that theyareencodedby BamHIp.Figure3 (right) shows that thetwo arrested bands correspond
toselected VI43 and VI39. It shouldbe noted, however, that VI39 comigrates withaminorcell
polypeptide, and that RNA preparations
con-taining high levels of its mRNA (i.e., early RNA)
must be used for efficientdetection in
hybrid-arrestexperiments.
Analysis ofsequences atthe 5' terminus ofTKmRNA. Initialexperimentswere aimed
atlocating those BamHIp sequenceswhichare represented in the mRNA's for VI43 and VI39. FragmentsofpTK1 generated by SmaIorAluI
were used in hybrid-arrested translation, and
A
TK
0 X
E
0. u
4
0 x
E
0. u
2
RNA(l)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.500.62.456.64.336.2]z pAT pTK I
sr- IL
I Z H D H D
I 2 a A A a 7
43
39
Al
z MA
0) H D
.3
[image:5.500.111.398.76.364.2]39
FIG. 3. Hybridarrestoftranslation by pTKI DNA. Earlyinfected(INF)BHKceURNA washybridized to
pATl53DNAorpTKIDNA anddirectly translated (tracks labeled H) or denatured before translation (tracks labeledD). RNA preparation usedwereno addedRNA (tracks I and 8), mock-infectedceURNA (track 2), early INFcellRNA (track 3), hybridized to pAT153 DNA (track 4), track 4 sampledenatured (track 5), hybridizedtopTK1 DNA (track 6), track 6 sample denatured (track 7). From a separate experiment, the productsofpTK1-selected RNA (track 9) werecompared (tracks 8 through 11) with those ofpTKI-arrested RNA(track10).(Track11showstrack 10sampledenatured.)Symbols: , known virus-inducedpolypeptides; (0) polypeptideswhosesynthesis is arrested byincubationwitheither pATl53 orpTK1 DNA.
these
experiments
showed that such
sequenceslay within the 2.02-kbp PvuII fragment
(0.50 to2.52
kbp from the 5' BamHI site) previously
reported
topossess
full
transforming activity
(7)(data
notshown).
Further definition
wasachieved
by
useof small DNA
fragments
gen-erated
by double
digests
orby recleavage of
larger
fragments followed by
asecond
cycle of
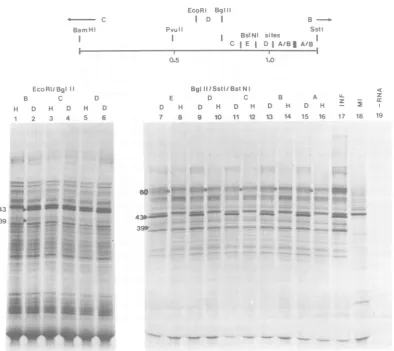
electrophoresis and purification. Figure
4shows
hybrid-arrested translation in the region which
should contain the mRNA
5'terminus and
un-translated
sequencesof the
TK gene(42).
All
five BstNI
fragments,
derivedby
cleavage
of thepTK1
BgII-SstI fragment (0.74
to1.24),
ar-rested
translation
ofV143
andVI39.
Fragments
C
(-0.38
to0.61)
and
D(0.61
to0.74)
from theEcoRI-BglII
double
digest failed
toarrest trans-lationunder standard
conditions,
whereasfrag-mentB
(0.74
to3.0)
of thesamedigest
wasfully
active.This
experiment, therefore,
shows thatafragment
assmall
as78bp (the
estimated size of
BglII-SstI-BstNI fragment
E)
canefficiently
ar-rest
translation.
Inspection of
Fig.
4,tracks
7through
16, shows
a
minor
polypeptide
of
an apparentmolecular
weight of 60,000 whose
synthesis
wasarrested
by
all
BglII-SstI-BstNI fragments.
This
species
has
notbeen
resolved
soclearly
in anyother
autoradiograms, and it
istherefore
notpossible
to
determine whether the effect
isdue
toBamHI
p
DNA
ortothe
self-arrest described above.
Nucleotide
sequencesof the
potentially
inter-esting
regions around the
BglII
site
weredeter-minedby the Maxam-Gilbert method (26) and
by
the use ofdideoxytriphosphates
incopying
single-stranded fragments
cloned inphage
M13mp2.
The440-bp
region depicted
inFig.
5combines
data fromfragments end-labeled
atthe
EcoRIsite
ortheBglII
site and fromEcoRI
fragments
cloned
inM13mp2.
Five
ATG
codons,
potential protein
synthesis
initiators,
canbe detected. The first
four of
on November 10, 2019 by guest
http://jvi.asm.org/
598 PRESTON AND McGEOCH
- C
BamHl
I
Pvull
0.5
EcoRI Bgl11
I D I B-.
Sstl
I BstNI sites C I E
I
D IA/BI
A/B1.0
EcoRlIBgI11
B C D
D H D H D
Bgl11ISstIl/ BstN I
E D C
D H D H D H
7 a a 44U11 12
B A U.z i cZ
D H D H - a
13 14 15 16 17 18 19
4
3
FIG. 4. Hybrid-arrested translation with TK mRNA 5'sequences. Fragments
of pTK1
from
anEcoRI-BglIIdoubledigest(tracks1 through 6)or aBstNIdigest of theBglII-SstI fragment(tracks7through16)
wereused. RNAwastranslateddirectly(H)orafterdenaturation(D).Theorigins ofthefragmentsareshown
inthemap(BglII-SstI-BstNI fragmentsAand B havenotbeenordered). Track17,translationproducts of
the original RNA; track 18, BHKcell RNA; and track 19, endogenousproducts of the cell-free system.
Symbols:0,anadditionalpolypeptide (60) which is arrestedbyallBglII-SstI-BstNIfragments.
these, all upstreamfrom the BglII site, are in
closed reading frames since protein synthesis
termination codons follow closely in the same
phase, whereas the fifth, located 57-bp
down-streamfromtheBglII site, isin anopenframe
asfarastheanalysiscandetect.Provided
splic-ing doesnotoccurinthe region analyzed,this is the only candidate site for protein synthesis
initiation.
Upstream from the BgIII site, various
se-quences previously recognized in the initiation regions ofmanyeucaryoticgenes canbe
identi-fied (5, 6). Moststriking is the AT-richregion, whichis verysimilar toa Hogness box
(GTA-TAATAG), atpositions 197 through207. This
sequence is usually approximately 30 bp
up-stream from an mRNA 5' terminus, and the
sequence CGAATAC (224 through 230) in the
appropriate position resembles the consensus
sequences found at the mRNA 5' capping site (6). At5and12bpupstreamfromthe Hogness box are two GGTCCsequence, whichare very
similar totheGATCCconsensus (6). Twoother regions further upstream from the mRNA
ter-minusareatpositions 144through152 (the-70 to -80homology) and at positions 83 through 92,anAT-richsequencecommonly found130to
150bpupstreamfromthemRNA terminus(5). Consideration of theorganization ofsequences
therefore suggests that an mRNA is initiated
approximately 50 bp upstream from the BglII
site,aconclusionwhich iscompatible with
ini-tiation ofprotein synthesis 57 bp downstream
from theBglIIsite.
H 1
43
39
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.500.63.458.76.427.2]SEQUENCES AT THE 5' TERMINUS OF THE HSV-1 TK GENE
PstI PvulI
5'GACTGC____AGCTTCAGGGA_TGGCG_
CTTCATC_CGTGGCCCGTTGCTCGCGV_TGCTGGCGGTGTCCrCGGGAA__TAAT_TGCQI5TCT
20 40 60 80 100
EcoRI Sau96I
TTAGTTCTATGATGACACAAACCCCGCCCAGCGTCTTGTCATTGGCG GAACACGCAGATGCAG
GGGGCGGCGGC
dCIACTTCGCA
120 146 GGYCAATCT 160 GATCC GATCC GTA 200
PstI Bgl II
TATTAAGfiTGACGCGTGTGG,CCTCGAATACCGAGCGACCC
eCGACCCGCTTAACAG,GTCAACAGCGTGCCGCAbA,TCTTGGTGGCGTGAAACTCC
TAATAG 220 YCATTCR MRNA 260
280
300BstNI
CGCACCTCTTCGGCCAGCGiC,CTTGTAGAAGCGCGTATGGC,TTCGTACCCCTGCCATCAM0ACGCGTCTGCGTTCGAC
CTGCGCGTTCTCGCGGCCA320 - 340 360 380 400
TAACAACCGACGTACGGCGT,1CGCCCTCG~CCGG4RCAA, 3'
420
440
FIG. 5. Sequenceof440bpatthe 5'terminus of the TK gene. Previously recognized consensus sequences
areshownatappropriatepositions, apartfromthe-140AT-richregion, which has a broken underline. The proposed initiation sitefor protein synthesis has a solid double underline; other ATG codons (all in a different, closedreadingframe)have broken double underlines. Restriction sites are given solid underlines, andtwohigh GC regions which may be important in interpreting hybrid-arrested translation are boxed. Y, Pyrimidine;R, purine.
In
view of the
implications from
studyof
theTK
gene 5'sequences, theeffects
of the region upstreamfrom the
BglII site
onmRNA selectionand
hybrid-arrested translation
wasinvestigatedfurther, since
no arrestcould be obtained by
EcoRI-BglII fragments from this region (Fig.
4).To
facilitate these
experiments, plasmid pTK6,
which contains
only the small
BamHI-BglII
fragment of pTK1 (0.0
to0.74),
wasconstructed
(see
Fig. 10). This
plasmid
DNA wasable
toselect mRNA for VI43 and VI39
apparently
asefficiently
aspTK1 DNA (Fig. 6, tracks
1through 6), showing that
sequencesof
these
mRNA's extend
upstreamfrom the
BglII
site.
When
hybrid-arrested
translation
wasinvesti-gated further,
it wasfound that reduction in
hybridization
temperature to540C
and
afivefold
increase in
equivalent
DNAconcentration
al-lowed
detectable
arrestof
VI43 and VI39
(Fig.
6,
tracks
7through 14).
Presumably
the
require-ment
for lower
temperaturereflects
thelower
GC
contentof the
region
immediately
upstream fromthe
BglII
site.
Afurther
interesting
point
is thattranslation
wasarrested
moreefficiently
when
Sau96I-cleaved
pTK6 DNA,
asopposed
toEcoRI-cleaved DNA,wasused. Theenzyme
Sau96I cleaves
adjacent
toaveryhigh
GC
region
that is17 to 34
bp
downstream from the EcoRI site. Thisregion
should reassociatestrongly
un-der the
hybridization
conditions,
and mayre-duce the
availability
of downstream sequencesfor
hybridization
with mRNA.The
conclusion
from theseexperiments
isthatV143
andVI39 mRNA sequencesextending
fromthe
BglII
site
canbe detected
by
RNAselection
and
hybrid-arrested translation under
appropri-ate
conditions. The altered characteristics of
hybrid-arrested translation
suggestthat the
mRNA
sequencesdo
notextend
entirely
through the
EcoRI-BglII
fragment
D(0.61
to0.74), since
BglII-SstI-BstNI fragment
E,
afrag-ment
with
similar
overall
nucleotide
composi-tion
including
aregion of
veryhigh GC, inhibited
translation under standard conditions
(Fig. 4).
Itis
mostlikely
that the mRNA
5'terminus
is 20to30
bp downstream from
theHogness
box andthat the
protein
synthesis initiation codon
is 57bp downstream from the
BglII
site.
However,
from the results
presented here,
the
possibility
that
sequences betweenthe PvuII site
and the
very
high GC
region (0.50
to0.63)
specify
ashort
5'
leader, which
isspliced
onto5'
noncoding
sequences, cannot
be
excluded.
Sequences
atTKmRNA 3'
terminus.
Hy-brid-arrested translation with
fragments
derived
from
the
3'end
of the TK geneis
shown inFig.
7.
Three
fragments within SmaI
fragment
A(0.25
to1.91) inhibited
synthesis
of
V143 and
VI39.
Additionally,
truncated
polypeptides,
pre-sumably
resulting
from ribosome terminationatthe
duplexed site,
could bedetected,
and theseare
labeled
in tracks2,
4 and 6 ofFig.
7. Theintensities of these
bands,
ascompared
tothose ofVI43 afterdenaturation,
werelower thanex-pected
andinsufficient
for confidentalignment
of
restriction
sites withcoding
sequences. The16,000-molecular-weight
species
formedanovelband,
but the29,000-
andon November 10, 2019 by guest
http://jvi.asm.org/
[image:7.500.48.451.69.284.2]600
PRESTON AND McGEOCHSau961
58 .54
pTKIpTK6 pAT -RNA
EcoRI
58 54
H D H D H D H D
2 3 4 5 6
3
FIG. 6. RNA selectionandhybrid-arrestedtranslation withpTK6DNA. Shownaretranslationproducts ofmRNA'sselectedby pTK1, pTK6, andpAT153DNAfilters (tracks 3, 4,and5, respectively); products of mock-infected (MI)BHKcellRNA(track1),infected (INF)cellRNA(track 2), endogenous proteins (track 6);
andhybrid-arrestedtranslationwithpTK6 DNA,cleaved with Sau96I(tracks7through 10)orEcoRI(tracks
11through 14),at54°Cor58°C.RNAwastranslateddirectly (H)orafterdenaturation (D).
weight bandswere onlydetectable as enhance-ments ofotherpolypeptides. Aproductof mo-lecularweight 35,000has beenproduced by
ar-restofpTK1-selectedmRNA with
SmaI-Hinfl-AvaI fragmentA(1.68to1.91) (datanotshown), and thereforeappearstobeagenuinetruncated
polypeptide.
The SmaI-AluI-DdeI fragment B (2.31 to
2.55) didnot arresttranslationof V143 and
VI39,
but fragmet A from this digest (1.91 to 2.31) partially inhibited synthesis of both polypep-tides(compare theintensities of actin andVI43 inFig. 7, track 8). This curious effect could bea real property ofsequences in SmaI-AluI-DdeIfragment A, orcould be due to contamination by degradation products of SmaI fragment A
(0.25to1.91) during electrophoresis. Hybrid-ar-rested translationwastherefore carriedoutwith increasing concentrations ofa
BglII-SstI
frag-ment (0.74 to 1.24), SmaI fragment B (1.91 to
2.69) isolated from aSmaI digest of pTK1, or SmaIfragment B, isolated froma SmaI-BglII-SstI-HinfIdigestofpTK1, which reduced SmaI fragment A to fragments smaller than SmaI fragment B and should eliminate any possible
contamination. Figure 8 shows that maximum
inhibition ofVI43 synthesiswasachieved with
approximately 1 ug equivalent (i.e., derived from 1
,tg
ofpTK1 DNA) of each fragment, and that increasing the concentration ofSmaI fragment B afurthersixfold didnotgiveagreatereffect. Similar results were obtained with whichever digest wasused togenerate SmaIfragment B, and, therefore, the partialarrestisa character-istic of thisfragment.The size of the35,000-molecular-weight trun-cated polypeptide produced by arrest with SmaI-Hinfl-AvaIfragment Asuggests thatthe codingsequencesofVI43 endatapproximately 230bp downstream, i.e.,veryclosetotheSmaI site, provided thereare nointrons in this region. Partial arrest with SmaI fragment B might therefore resultfrom hybridization with the 3' untranslatedregionorfromanoverlap into cod-ingsequencesproducing, atreduced efficiency, atruncatedpolypeptide whose reduced molec-ularweightcould notbedetected. Since it has been shown thathybridization of plasmid DNA
to the entire 3' noncoding region of 8-globin mRNA doesnotaffect translation in vitro (19),
U. z
1
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.500.111.401.73.354.2]Sat
AIl
HinfI Aval
1 1
Alul
E I
Smai
A I
1.5
Ddel
A I
2.0
Pvull
Alul
8
I
Smai2-5
Alul SmaiHinf I/AvaI
E a A
D H D H D H
1 2 3 4 5 6
SmaUAlul/DdeI
A B M
D H D H Z
[image:9.500.109.399.71.474.2]7 8 9 10 11
FIG. 7. Hybrid-arrested translation with 3' fragments of TKgenesequences.Five fragmentsofpTKI DNA,
indicated in the map, were used in hybrid-arrested translation (tracks 1 through 10). RNA was either
translateddirectly(H)orafter denaturation(D). Shownaretranslationproducts of infected (INF)cellRNA
(track 11), mock-infected (MI) BHK ceU RNA (track 12) and endogenous proteins (track 13). The shorter
exposureonthelowerpaneldemonstratespartialinhibitionof VI43synthesis by SmaI-AluI-DdeI fragment
Amoreclearly(track 8). Symbols:<,putativetruncated
polypeptides.
the latter interpretation appears most likely. The 3' limit ofcoding sequences for VI43 and
V139 thereforeprobablylieswithin SmaI-AluI-DdeIfragmentA,nomorethan 30bp from the SmaI site.
Hybrid-arrestofTKsynthesis.Thecoding
sequencesforTKenzymewerealsoanalyzed by hybrid-arrested translation, and Table 1 shows thatfragmentswhich inhibitedsynthesisof TK
mapped in thesameregionasVI43andVI39.It
isinterestingtonotethat theputativetruncated
polypeptide generated by SmaI fragment B
re-tained TKactivity.
Synthesisinvivoofapolypeptide
equiv-alentto V139. CodingsequencesforVI43 and
V139 lie withinthe same 1,170 bp between the
BgM
(0.74) andSmaI (1.91) sites and presum-ablyhave thesamepolarity,sincethey behave inthesameaberrantwaytohybridarrestwith fragmentsfrom the mRNA termini. Sincethereis insufficientcoding capacity for twoadjacent polypeptidesof thesesizes,theirsequencesmust
overlap. It wasconsidered importantto deter-mine whether both polypeptides were
synthe-_z
12 13
on November 10, 2019 by guest
http://jvi.asm.org/
602
PRESTON AND McGEOCHo100
(A
~80
c60
d>
40*
&20-0 2 4 6
DNA(pg
equiv)
FIG. 8. Hybrid-arrestofVI43 with SmaIfragment
B and BglII-SstIDNA. Various concentrations of
[image:10.500.89.221.61.257.2]fragments ofBglII-SstI (a), SmaIfragmentB iso-latedfrom aSmaIdigest (0), orSmaIfragmentB isolated from a SmaI-BglII-SstI-Hinfl digest (0) were used.Autoradiograms werescanned, andthe radioactivity in VI43wascompared with neighbor-ing,unaffected VI37.5.
TABLE 1. Hybrid-arrestofTKsynthesizedin vitro
[3H]thymidine phosphorylated DNAfragment used in Location (cpmx103)
hybrid-arrest (kbp)'
Hybrid- Dena-ized tured
pTK1 0.0 72.4
pAT153 78.9 59.4
SmaI-B 1.91-2.69 15.3 83.8
Hinfl-SmaI 1.50-1.91 1.1 91.9
AluI-E 1.24-1.58 1.0 52.3
BglII-SstI-BstNI-A| 0.0 68.2
BgIII-SstI-BstNI-B 1.01-1.24 0.0 67.6
BglII-SstI-BstNI-D 0.92-1.01 0.5 58.0
BglII-SstI-BstNI-C 0.82-0.92 0.0 66.4
BglII-SstI-BstNI-E 0.74-0.82 0.0 48.8
pTK6,54oCb 0.0-0.74 5.0 72.6
pTK6,58oCb 0.0-0.74 53.6 72.4
EcoRI-BglII-C -0.38-0.61 52.0 47.2 'Location offragmentsisgiveninkbp downstream from the BamHIsite.
bHybrid-arrestwithpTK6 used Sau96I-cleaved DNAat
fivefold higherconcentration at the temperatureindicated.
sized ininfected
cells
aswell
asin vitro. Figure 9 showspulse-labeled polypeptides
from BHKcells
infected withwild-type
HSV-1 or theTK-mutant
dPyKV-7,
which fails tosynthesize
V143
(33).Comparison
of tracks4, 5, 6, and 7 of Fig.9
reveals
that a bandcomigrating
with VI39andof
approximately
thesamerelativeintensity
wasdetected in
wild-type
HSV-1-infectedcells,
but notinmock-infected ordPyKV-'-infected
cells.
The
probable
reasonwhy
the V139equivalent
has not
previously
beendetected
ininfected cells
is
that theintense band below it
israpidly
proc-essed
to aslower
migrating form which obscures
it. A short
labeling
period (15
min wasused
toobtain the results shown
inFig. 9) is
necessarytoprevent this masking effect.
DISCUSSION
Many
of the
techniques described here have
not
previously been applied
toherpesviruses,
and it
istherefore worthwhile
todiscuss them
with
reference
topossible problems
arising fromthe
high
GC
contentof HSV
DNA.mRNA
selection
onnitrocellulose
filters wasvery
specific,
asshown
inFig.
1through
3.It waspossible
toenrich BamHI
p-specific
RNAs withno
detectable
contamination
from othermRNA's. The
efficiency of hybridization
tofil-ter-bound DNA is
relatively low,
withonly
5toIN VITRO I IN VIVO
-RNA Ml SEL INF WT MI -7 WT -7
1 2 3 4 5 a 7 a a
FIG. 9. SynthesisofVI43 and VI39 invitro and in
vivo.Shownareendogenous products ofthe transla-tionsystem(track 1),polypeptidesdirected by
mock-infected (MI) BHK cellRNA, pTK1-selected (SEL) RNA, andinfected (INF) BHK cell RNA (tracks 2
through 4, respectively), polypeptides synthesizedin
BHK cellsat 5hafter infection at31°Cwith
wild-type(WT) HSV-1ordPyKj7(tracks 5, 7, 8, and 9),or MIBHKcells(track 6).
I I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.500.271.459.284.575.2] [image:10.500.64.256.356.519.2]10%
of the original mRNA activity recovered,
and high RNA concentrations
aretherefore
re-quired. The high GC
contentdoes
notseriously
affect
hybridization of RNA
todenatured,
im-mobilized pTK1 DNA, and
thefinding
thatpTK6 DNA
selects mRNA shows that only
asmall region of sequence homology
isnecessary.
The
washing
procedure described for adenovirus
mRNA's (39)
removesnonspecifically bound
HSV mRNA's.
Hybrid-arrested
translation is
apotentially
more
sensitive method of mRNA
mapping and
the
studies
presented
here have used the
small-est
fragments
yetreported.
Since
both
DNA-DNA and DNA-DNA-RNA interactions
can occur,temperature
control
tofavor DNA-RNA
reac-tions
mustcompensatefor
GC
content(15, 43).
This
results
inthe
useof relatively high
hybrid-ization temperatures for HSV mRNA's.
Hybrid-arrested translation could
notdetect
mRNA's
for minor
polypeptides
which
comigrate
with
major cellular
orviral
bands,
but otherwise the
specificity
seemsequal
tothat of mRNA
selec-tion.
Some
mRNA's, however,
cannotbe
mapped
since
they
areinactivated
by
mock-hy-bridization, emphasizing
the
importance of
in-cluding
appropriate
controls. DNA
fragments
assmall
as78bp
efficiently
arresttranslation,
pro-vided they
hybridize entirely
tomRNA
coding
regions. When the
areaof
overlap
includes
mRNA
ends, the results become
moredifficult
to
interpret and further
investigation
is
neces-sary.
Production of truncated
polypeptides
wasless
efficient than
recently
described for
influ-enza
mRNA's in the wheat germ
system(20).
Possibly, ribosome release
atthe
abnormal site
is
inefficient, and the
resulting decreased
syn-thesis of
truncated
products
is
moremarked in
the
reticulocyte
lysate,
asit
supports morerounds of
translation
than the wheat
germsys-tem.
Hybrid-arrest and selection
experiments
with
pTK6 DNA
suggestthat sequences
represented
in VI43
and VI39 mRNA end within
90bp
(the
site of the
veryhigh GC region)
upstream fromthe
BglII site.
Analysis
of the nucleotide
se-quence in
this
region
shows
manyfeatures
char-acteristic of
eucaryotic
mRNA
5'termini.
Thepositions of the
consensus sequencesfound
inother
genesimply
that
anmRNA initiation site
exists at
approximately
50bp
from theBgII
site.
Recently, S1 nuclease
mapping
has identi-fied amajor
mRNA 5'terminus
atthisposition
(48),
reinforcing
theconclusions
made here. Ifthe AT-rich
region
80bp
further upstream is necessary forinitiation
oftranscription,
the fail-ureof
EcoRI-cleaved
pTK1
DNAtotransform
TK-
cells
canreadily
beinterpreted.
Location
of
the 3' limit ofcoding
sequencesclose
tothe
SmaI
site (1.91) is based on theproperties of hybrid-arrest
withSmaI
fragment
B and thesizes of
truncated
polypeptides
gen-erated
byfragments hybridized
tothe 3' end ofVI43
mRNA.
Thefinding
that theputative
shortened polypeptide produced
byhybridiza-tion with
SmaI
fragment
B has TK activitymight
explain the ability
ofSmaI-cleaved
plas-mids
totransform
cells
(7), and it would beinteresting
toexamine TK mRNA in suchcells.
The experiments reported
here have deriveda
precise location
ofcoding regions
forVI43
andVI39 and
havetherefore enabled
thefunction
ofother sequences
necessary for TK geneexpres-sion
tobe understood.
Sincegenomic
codingsequences
for VI43 spanapproximately
1,110 bp,and assuming the location
ofproteinsynthesis
initiation
iscorrect,
it isclear
thatintrons
ofsignificant
sizecannot
exist in this gene.Fur-thermore,
the absence in adjacent sequences
ofother
mRNA's
withsimilar
abundance
orkinet-ics
of
synthesis indicates that the
TK gene iscontrolled
as anindependent
unitrather
thanone
of
a groupof coordinately expressed
prod-ucts.Despite the high GC content
andrequire-ment
for
aviral
polypeptide
ininfected
cells (24,35,
36), the TK
geneexhibits all
thepostulated
signals
for RNA polymerase
binding andinitia-tion of
transcription.
An
important finding from mapping
BamHIp-specific mRNA's
isthe identification of
twopolypeptides
whosemRNA's
share codingse-quences. Whether VI43 and VI39
areproduced
from
the
sametranslational
reading framecan-not
be
determined until further analysis of these
polypeptides
has been
performed.
Nevertheless,
mechanisms by which VI39 might be related
toVI43
canbe
considered. The
possibility
of
pro-teolytic cleavage of VI43
waseliminated by the
finding that the relative proportions of the
twopolypeptides
did
notchange during
achase of
up to 20hin vitro
(data
notshown).
Premature
termination of
protein synthesis
also
seemsun-likely, since SmaI
fragment
Bpartially
inhibits
production
of both
V139
and
VI43,
eventhough
it cannot
overlap sufficient
coding
sequences tomask
apretermination
site. The other
possibili-ties
appear tobe
splicing
of
coding
sequencesfrom VI43 mRNA
orfalse initiation of
protein
synthesis
at asite
100 to 150bp
downstream
from theVI43 initiation codon.
The
evidence
currently
available doesnotsup-port the concept of
splicing
VI43 mRNA togenerate
VI39 mRNA. These mRNA's couldnotbe
separated
by
sucrosedensity
gradient
cen-trifugation
orpolyacrylamide gel electrophoresis
under
denaturing
conditions(data
notshown),
and no band 100 to 150
bp
smaller
thanV143
mRNA
could
bedetected
whenpTK1
DNAwason November 10, 2019 by guest
http://jvi.asm.org/
604
AND McGEOCHIngiotion
TK 43
3p
1
:mRNA
Hinf Alu
BmHlinf SaI I Eco BI
I
Term.
1K 29K 35K
Alu
Ss[t rif iU Snma
I1Ij
I IIkbP
I I I2kbp IPT
Kl
S
!AID!
pTI6L FT Bst NI AVGI Ddel
Sau961
FIG. 10. Summary ofmRNAmappingdata.Sequenceswhich arrest translation under standard conditions
areshown in solidlines;those whichrequirealtered conditionsorpartiallyarrestareshown in brokenlines. Points16,000, 29,000,and35,000molecularweightunitsfromtheproposed VI43initiation codonaremarked and should becomparedtotheAluI,HinflandAvaIsites whichgeneratedputativetruncatedpolypeptides ofthesesizes. Theproposedtermination sitefor VI43andVI39and initiation sitefor VI39arealsoindicated. Restriction sites above the linesarefrom completemapsofpTK1;those below have beenidentified onlyin
digests ofpTK1 subfragments.
used to probe fractionated RNA on Northern blots (J. B. Clementsand N. M.Wilkie, manu-scriptinpreparation). Such experiments might
not succeed because the expected mRNA size reduction is less thantheaveragelengthof the
heterogeneous polyadenylic acid tract. Any splice would be more than 270bp downstream from theBglII site,since removal of 100 to 150
bp in this region would prevent arrest ofVI39 synthesisbyone or moreof thesmall BglII-SstI-BstNI fragments C, D, or E. Alternatively, a situation in which ribosomes occasionallyread
through the usualinitiation codoniscompatible with the datapresentedhere, but rigorous dis-crimination between thepossibilitieswillrequire detailedpeptide mapping ofVI43 and the minor VI39 band. It should be noted that ribosome attachment for bothVI43andVI39occurs near the 5'terminus(18), since pTK6 DNAcanarrest
synthesisofbothpolypeptides.
Figure 10 summarizes the mapping of TK,
VI43,
and VI39 mRNA's, and indicates thepu-tative initiation codon forVI39.
AttherecentColdSpring HarborHerpesvirus Workshop, wherethis workwaspresented, the complete sequence ofthe HSV-1 TKgene was reported by M. Wagner, J. Sharp, and W. C. Summers, and by S. McKnight. Analysis of strain CL101 shows that the SmaI-B overlaps coding sequences by 18 bp, and that the next ATG codon from theproposed VI43initiator is
130 bp downstream and in phase (M. Wagner, personalcommunication).
ACKNOWLEDGMENTS
Wethank J.H.Subak-Sharpe forhis interest in this work, and critical readingof themanuscript. M. Dunlop and A. Dolangaveskilledtechnicalassistance.
LIrERATURE CITED
1.Anderson,K.P.,R. H.Costa,L E.Holland,and E. K.Wagner.1980.Characterization ofherpes simplex virustype 1RNApresentin the absence of de novo
protein synthesis.J.Virol.34:9-27.
2. Anderson,K.P.,L.E.Holland,B. H.Gaylord,and E. K.Wagner.1980.Isolation and translation of mRNA encodedbyaspecific regionof theherpessimplexvirus type1genome.J. Virol. 33:749-759.
3. Anderson,S.,M. J.Gait,L.Mayol,andI. G. Young. 1980. A shortprimerforsequencingDNAcloned in the
single-stranded phagevectorM13mp2. Nucleic Acids Res. 8:1731-1742.
4. Bacchetti,S.,and F. L.Graham. 1977. Transfer of the
geneforthymidinekinasetothymidinekinase-deficient
humancells by purified herpes simplex viral DNA. Proc.Natl.Acad.Sci.U.S.A. 74:1590-1594.
5. Benoist, C.,K.O'Hare,R.Breathnach,and P.
Cham-bon. 1980.The ovalbumin gene-sequenceofputative
controlregions.Nucleic Acids. Res.8:127-142.
6. Busslinger,M., R. Portmann,J. C.Irminger,and M. L.Birnstiel. 1980.Ubiquitousandgene-specific
regu-latory5'sequencesinaseaurchin histoneDNAclone
coding for histone protein variants. Nucleic Acids Res.
8:957-977.
7. Colbere-Garapin, F., S.Chousterman, F.
Horodni-ceanu,P.Kourilsky,and A-C.Garapin.1979.
Clon-ing of the active thymidine kinasegeneof herpes
sim-plex virustype1inEscherichia coli K-12. Proc. Natl.
Acad. Sci. U.S.A. 76:3755-3759.
8. Cooper, J.A., and B. Moss.1979.Translation of specific vaccinia virus RNAs purifiedasRNA-DNAhybridson
potassium iodide gradients.Nucleic Acids Res.
6:3599-3612.
9. Cremer, K.,M.Bodemer,and W. C.Summers. 1978. Characterization ofthe mRNA forherpessimplexvirus
thymidine kinase by cell-free synthesis of active en-zyme.NucleicAcids Res. 6:2333-2344.
10. Enquist, L.W.,G. F.VandeWoude, M. Wagner,J. R.
Smiley,and W.C.Summers.1979.Constructionand
characterisation ofarecombinantplasmid encoding the genefor thethymidinekinase ofherpessimplextype1
virus.Gene7:335-342.
11. Gronenborn, B.,and J.Messing.1978.Methylation of single-strandedDNAin vitro introducesnewrestriction endonucleasecleavagesites. Nature(London) 272:375-377.
12.Halbert,D.N.,D. J.Spector,and H. J.Raskas.1971. Alu
Sma
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.500.123.408.77.187.2]In vitrotranslation products specified by the transform-ingregion ofadenovirus type 2. J.Virol.31:621-629. 13. Haliburton, I. W., L. S. Morse, B. Roizman, and K.
E.Quinn.1980.Mapping of thethymidine kinase genes oftype 1 and type 2 herpes simplex viruses using intertypicrecombinants.J. Gen.Virol.49:235-253. 14.Holland,L. E., K. P. Anderson, C. Shipman, and E.
K.Wagner. 1980. Viral DNA synthesis is required for the efficientexpression of specific herpessimplexvirus type 1 mRNA species.Virology 101:10-24.
15.Holmes, D. S., R. H. Cohn, L H. Kedes, and N. Davidson.1977.Positions of seaurchin (stongylocen-trotuspurpuratus) histone genes relative to restriction endonucleasesites on thechimeric plasmids pSp2 and pSpl7. Biochemistry 16:1504-1514.
16. Inglis,S. C., D. J.McGeoch, and B. W. J. Mahy. 1977. Polypeptidesspecified by the influenza virus genome. 2.Assignmentofprotein codingfunctions to individual genome segmentsby in vitro translation. Virology 78: 522-536.
17. Kit,S.,andD. R.Dubbs.1977.Regulation of herpesvirus thymidine kinase activity in LM (TK-) cells trans-formed by ultraviolet light-irradiated herpes simplex virus.Virology76:331-340.
18.Kozak, M. 1978. How do eukaryotic ribosomes select initiation regionsin messenger RNA? Cell 15:1109-1123.
19. Kronenberg, H. M., B. E. Roberts, and A. Efstra-tiadis.1979.The 3'noncodingregion of,B-globin mRNA is notessentialfor in vitro translation. Nucleic Acids Res.6:153-166.
20. Lamb, R. A., P. W.Choppin,R. M.Chanock, and C.-J. Lai. 1980. Mappingof two overlapping genes for polypeptides NS,andNS2onRNAsegment 8 of influ-enzavirusgenome. Proc. Natl. Acad. Sci. U.S.A.77: 1857-1861.
21. Leiden, J. M., R. Buttyan, and P. G. Spear. 1976. Herpessimplexvirusgene expressionintransformed cells.I.Regulationof the viralthymidinekinasegene in transformedLcellsbyproductsofsuperinfectingvirus. J.Virol. 20:413-424.
22. Lewis, J. B., C. W. Anderson,andJ. F. Atkins. 1977. Furthermappingof late adenovirusgenesby cell-free translation of RNA selectedbyhybridizationtospecific DNAfragments.Cell 12:37-44.
23. Lewis, J.B., J. F. Atkins, C. W. Anderson, P. R. Baum,and R. F.Gesteland. 1975.Mappingof late adenovirus genesby cell-free translation of RNA
se-lected by hybridization to specific DNA fragments. Proc.Natl. Acad. Sci.U.S.A.72:1344-1348.
24. Leung,W.-C. 1978.Evidence foraherpes simplex virus-specific factorcontrolling thetranscription of deoxy-pyrimidine kinaseJ. Virol.27:269-274.
25. Marsden, H. S., I. K. Crombie, and J. H. Subak-Sharpe. 1976.Controlofprotein synthesisin
herpes-virus-infected cells:analysisof thepolypeptidesinduced by wild-type and sixteentemperature-sensitivemutants
ofstrain17.J. Gen. Virol. 31:347-371.
26. Maxam, A.M.,and W. Gilbert.1977. A newmethod for sequencing DNA. Proc. Natl.Acad.Sci.U.S.A. 74:560-564.
27. Maxam, A.M., and W. Gilbert 1980.Sequencing end-labelled DNA with base-specific chemical cleavage. MethodsEnzymol.65:499-560.
28. Minson, A.C.,P.Wildy,A.Buchan,and G.Darby. 1978.Introduction of theherpessimplexvirus
thymi-dine kinasegeneinto mousecellsusingvirusDNAor transformedcell DNA.Cell 13:581-587.
29. Munyon,W.,E.Kraiselburd,D.Davis,and J. Mann. 1971.Transfer ofthymidinekinsetothymidine
kinase-less L cells withultraviolet-irradiated herpessimplex
virus. J.Virol.7:813-820.
30. Paterson, B. M., B. E. Roberts, and E. L Kuff. 1977. Structural gene identification and mapping by DNA mRNAhybrid-arrested cell-free translation. Proc. Natl. Acad. Sci. U.S.A.74:4370-4374.
31. Pelham, H. R. B., and R. J.Jackson.1976. Anefficient mRNA-dependent tannslationsystem fromreticulocyte lysates. Eur. J.Biochem.67:247-256.
32. Pellicer,A., M. Wigler, R. Axel, and S.Silverstein.
1978. Thetransfer and stable integration of theHSV thymidine kinase gene into mouse cells. Cell 14:133-141.
33. Preston,C.M. 1977.Thecell-free synthesis of herpesvi-rus-induced polypeptides. Virology 78:344-353. 34. Preston,C. M. 1977.Cell-freesynthesis of herpes simplex
virus-codedpyrimidinedeoxyribonucleosidekinase en-zyme. J. Virol.23:455-460.
35. Preston, C. M. 1979. Control of herpessimplex type 1 mRNAsynthesis in cells infected with wild-type virus orthetemperature-sensitivemutant tsK. J. Virol.29: 275-284.
36.Preston,C. M. 1979.Abnormalproperties of an immedi-ateearlypolypeptideincellsinfected withthe herpes simplexvirus type1mutanttsK. J. Virol.32:357-369. 37. Rapp, F., N. Turner, and P.Schaffer. 1980.
Biochemi-caltransfonrnationbytemperature-sensitivemutants of herpessimplex virustype 1. J. Virol.34:704-710. 38. Ricciardi,R.P., J. S.Miller,and B. E. Roberts. 1979.
Purificationand mapping ofspecific mRNAs by
hybrid-ization-selection and cell-freetranslation. Proc. Natl. Acad. Sci.U.S.A.76:49274931.
39. Sanger,F., S.Nicklen,andA. R. Coulson. 1977. DNA sequencing with chain tenninating inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74:5463-5467.
40.Schreier,P.H., and R.Cortese.1979. Afast and simple method for sequencing DNA cloned in the single-strandedbacteriophageM13. J.Mol.Biol.129:169-172.
41.Smiley,J.R., M. J.Wagner, W. P. Summers, and W. C. Summers.1980.Geneticandphysicalevidence for thepolarity oftransciption of thethymidine kinase gene ofherpessimplexvirus.Virology102:83-93. 42. Smith,A. E.,S. T. Bayley,T.Wheeler, and W. F.
Mangel.1975.Cell-freesynthesisofpolyomaandSV40
viral proteins, p. 331-338. In A.-L. Haenni and, G. Beaud (ed.), Invitrotranscription and translation of viral genomes.INSERM,Paris.
43. Thomas, M., R. L. White, and R. W. Davis. 1976.
Hybridizationof RNA todouble-stranded DNA: for-mationofR-loops. Proc.Natl. Acad. Sci. U.S.A. 73:
2294-2298.
44. Twigg,A. J., and D. Sherratt. 1980.
Trans-comple-mentablecopy-numbermutantsofplasmidColEl. Na-ture(London)283:216-218.
45. Wigler, M.,A.Pellicer,S.Silverstein,and R.Axel. 1978. Biochemical transfer ofsingle-copy eucaryotic
genesusing totalcellularDNAasdonor.Cell 14:725-731.
46.Wilkie,N.M.,A.Davison,P.Chartrand,N. D.Stow,
V.G.Preston,and M. C.Timbury.1979. Recombi-nation inherpessimplexvirus:mappingof mutations and analysisofintertypic recombinants. Cold Spring
HarborSymp. Quant.Biol.43:827-840.
47. Wilkie,N.M.,J.B.Clements,W.Boll,N.Mantei,D.
Lonsdale,and C. Weissmann.1979.Hybridplasmnids containinganactivethymidinekinasegene ofherpes simplexvirus1.Nucleic Acids Res.7:859-877. 48.Wilkie,N.M.,R.P.Eglin,P.G.Sanders,andJ. B.
Clements. 1980. The association of herpes simplex
virus withsquamous carcinoma of thecervix,and
stud-iesonthethymidinekinasegene.Proc. R. Soc.London Ser.B210:411-421.





![TABLE 1. Hybrid-arrest of TK synthesized in vitro[3H]thymidine](https://thumb-us.123doks.com/thumbv2/123dok_us/1479984.100638/10.500.271.459.284.575/table-hybrid-arrest-of-tk-synthesized-vitro-thymidine.webp)

