0022-538X/78/0026-0126$02.00/0
Copyright ©1978 AmericanSocietyforMicrobiology PrintedinU.S.A.
Altered
E2
Glycoprotein
of
Sindbis Virus
and Its
Use
in
Complementation Studies
MOSHE BRACHAt ANDMILTON J.SCHLESINGER*
WashingtonUniversity Schoolof Medicine, Department of Microbiology and Immunology, St. Louis, Missouri63110
Received forpublication14November1977
We have detected a Sindbis virus variant that contains a
smaller-molecular-weight form of the viral glycoprotein E2. The molecular weight of the PE2
precursor and the glycosylation pattern of the smaller E2 are normal, thus indicating that this E2 is formed by an aberrant proteolytic cleavage. Thealtered E2 was detected in an
RNA'
temperature-sensitivemutantthat wasdefective inproteolytic cleavage, but the abnormal PE2-to-E2 reaction could be separated
fromthe tsmutationand is notitselfatemperature-sensitivedefect. We used the variant E2 as a marker to monitor the complementation reaction between an
RNA'
and an RNA- mutant and discovered that complementation was notreciprocal; the RNA defectwas corrected by theRNA' mutantgene products
but theRNA'defect was notcomplemented by any RNA- gene products.Other studies have shown that the smaller E2 is notpreferentially selected duringviral
maturation and budding. No significant changes have been detected in the
biological activity of virions with this altered E2 protein. Comparison of the
electrophoretic migrationof the El andE2Sindbis viralglycoproteinsin a
two-dimensional polyacrylamideslabgelsystemthat was first run inthe absence of
sulfhydryl-reducingreagent and then
with,B-mercaptoethanol
indicated that themobilityofEl,butnotthat ofE2,wassignificantly altered by reduction.
Two majorglycoproteins
(designated
Eland the E2 viralglycoprotein
from aglycosylated
E2) arefoundonthesurface ofSindbis virus,a precursorcalled
PE2,
and it is believedtooccursmallRNA-enveloped virus of the Togaviridae closetothe cell'splasmamembraneduring
bud-family (24).Theyareformedinthe infectedcell dingofvirus from the hostcell(26).Someofthe
from larger-molecular-weight polypeptide pre- temperature-sensitive (ts) mutants isolated by
cursors viaaseries ofposttranslational proteo- BurgeandPfefferkorn (3) have been found
de-lytic cleavages and glycosylation steps (16, 22, fective in theseproteolyticevents, and precursor
23, 25, 26, 28,29).Aninitialproteolytic activity
polypeptides
accumulate in cells infected withisbelievedtooccuronthepolyribosome of the the mutants atthenonpermissive temperature
infected cellaftertranslation of the viralstruc- (2, 13). During a survey ofviral proteins
pro-turalproteins has been initiatednearthe5'end ducedincells infectedwiththese different
mu-of the
viral-specific
26SmRNA, and this activity tants, we detected an alteration in theE2gly-releases viral
capsid
polypeptides thataggregatecoprotein
formedby
one of themutants whenwith viral 42S mRNA to form
nucleocapsids.
grown atthepermissive
temperature. Wecon-Continued translation of the 26S mRNAyields sidered the possibility that thisphenotype
re-the twoproteins destined tobecome the enve- sulted from the same genetic lesion that
pro-lope glycoproteins.A secondproteolyticactivity duced the ts condition and proceeded to study
occursbeforeorshortlyaftersynthesisofthese thenatureandoriginofthealtered E2 glycopro-membranepolypeptideshas beencompletedon tein. In thisreport,wedescribehowthevariant
thepolyribosome,butpriortoglycosylation (8, E2
glycoprotein
appears, show that it is not16). Failure to effect the initialcleavageresults related to the ts
defect,
andutilize the alteredin a protein of about 140,000 daltons (22, 23), E2asamarker for
following
eventsduringcom-andfailure of the later one leads to an accumu- plementation betweentwo tsmutants. lation of a polypeptide of 110,000 daltons (18,
23). A third proteaseactivityisrequiredtoform MATERIALS
AND
METHODSVirus and cell culture. Sindbists2 mutant and
t Present address: Biochemistry Department, Tel Aviv the parental wild-type heat-resistant HR strain (3)
University,TelAviv,Israel. wereobtained from E.Pfefferkorn(Dartmouth Med-126
on November 10, 2019 by guest
http://jvi.asm.org/
ical School, Hanover, N.H.). Mutant ts5 was obtained beled glycopeptides from E2 glycoprotein.
Sec-from B. M. Sefton (Salk Institute, San Diego, Calif.). ondary CEF monolayers in 100-mm dishes were in-Preparations of primary chicken embryo fibroblasts fected at an MOI of 100 with either the HR or the (CEF) and titration of virus have been described (2). revertant Rts2a strain of Sindbis virus at 37°C. Six Isolation of ts2 revertant strains.Astock of ts2 dishes were used for each strain. After 1 h of adsorp-that had been reisolated from a single plaque was tion, MEM containing 10% the normal amou.it of titered at either 30 or 40°C, and plaques were devel- glucose and 3%FCS was added. A total of 30,Ciof oped after2days. The ratio of PFU at 40°C to PFU D-[1-_4C]glucosamine (55 mCi/mmol; New England at 30°C was 5.5 x 106. Single plaques were picked Nuclear Corp., Boston, Mass.) was added to HR-in-from theplates incubated at 40°C and transferred to fected cells and 300
,Ci
ofD-[6-3H(N)]glucosamine (21ml ofminimal essential medium (MEM) containing Ci/mmol, New England Nuclear) to Rts2a-infected 3% fetal calf serum (FCS). Samples were diluted 10- cells after the adsorption period. Media were collected fold into phosphate-buffered saline containing 1% FCS after 12 h, cell debris wasremoved by brief centrifu-andretitered at 30 and 400C.Theratios of PFU at gation, and the virus sample was layered onto sucrose
400CtoPFU at 30°C for four plaque isolates were 0.6, gradients that consisted of 2 ml of 60% sucrose, 10ml 0.5, 0.47,and 0.82, respectively. Samples (0.2 ml) were of alinear gradient of 50 to 30% sucrose, and 12ml of diluted into2ml of phosphate-buffered saline plus 1% 10% sucrose. Sucrose solutions contained TNE and FCSand adsorbed to monolayers of CEF in 75-cm2 T 0.1% bovine serum albumin. After centrifugation at flasks. After 1h, the inoculum was replaced with 20 25,000 rpmfor3h at4°C inanSB110 rotor (Interna-ml of MEM containing 3% FCS, and thecells were tional EquipmentCo., Needham Heights, Mass.), frac-incubatedat37°C for21h.The media containing the tions werecollected, and radioactivity was measured revertantviral strains were stored at -70°. in smallsamples. The fractions containing the peak of
Labelingofviral-infectedcelis.Primary CEF in virus banding at 38%sucrose were pooled, diluted with
60-mmdishes were infected with either ts2 or ts5 virus twovolumes of TNE containing a sample of
nonradio-at amultiplicity of infection (MOI) of 200 and with ts2 active carrier virus (2x1011 PFU),and centrifuged at revertantsat anMOI of 100. After 1 h of adsorption, 50,000 rpm for90 mininaSpinco65 rotor at4°C. The
5ml of MEM containing 3% FCS was added to the pellets of virusweresuspended in0.4ml of 1%SDSin
plates, and the cellswereincubated at 30°C. At 9 h 0.5 MTris-hydrochloride (pH 9.0), reduced, and al-postinfection the medium was replaced with MEM kylated with iodoacetamide and the E2glycoproteins lacking amino acids butcontaining 3% FCS. For each isolated after electrophoresis on 10%polyacrylamide
virussample,twoplateswerekeptat300Candathird gels by the methods described previously (24). The platewasshiftedto40°C. After2h, the medium was eluted E2proteinsamplewaslyophilized, resuspended
replacedwith 1ml of MEMlacking amino acids and inasmallvolume of water, andprecipitated with 10
containing10,uCiofL-[3S]methionine (330 Ci/mmol; volumes of cold acetone. Bovineimmunoglobulin(200 Amersham/Searle, Arlington Heights, Ill.). After 20 ug)wasaddedascarrier materialpriortoadditionof minatthe abovetemperatures, mediumwasremoved, acetone.The driedproteinwassuspended in 0.5 ml of andoneplateat300Cwastreated with0.5ml of 2% 0.1MTris-hydrochloride (pH 8.0) containing 0.01 M sodium dodecyl sulfate (SDS) containing 1 M Tris CaCl2. Samples (25
pl)
of a 10-mg/ml solution of (pH 9.0). To the other twoplates was added 2 ml of Pronase (Calbiochem, LaJolla, Calif.,A grade), pre-MEM containing 3%FCSand four times the standard pared in thesamebuffer andpreincubated for 2 hatamountof aminoacids. Theplateswereincubated for 37°C,wereaddedat24-h intervals overa3-day
incu-an additional 1 h, and cellswerelysed with SDSas bation periodat37°C. The entire sample wasloaded
above. ontoaBio-Gel P-6 column(0.9 by 120cm) and eluted
Preparation of
[3S]methionine-labeled
virus. according to the method described by Sefton and PrimaryCEF intworoller bottleswereinfected with Keegstra(28).Atotalof70fractions(1 ml each)wereeither HRorts2virus in10mlofphosphate-buffered collected, and0.8mlwasaddedto8.0 ml ofaTriton saline and 1% FCS at an MOI of100. After 1 hof X-114-xylene(1:2 vol/vol) scintillation fluid. adsorption, each roller bottle received20ml of MEM Inhibition ofglycosylation withglucosamine.
lacking amino acids but containing 3%FCS. The HR PrimaryCEFwereinfected with HRorts2at anMOI bottlewaskeptat370Candthe ts2 bottleat300C. At of 100. At 7 hpostinfection at300C,the mediumwas 3 hpostinfection, the mediumwassupplementedwith replacedwithonelackingamino acids butcontaining
10%of thestandard amount of aminoacids and 100 3%FCSand,inone of eachpairofplates,20mM D-,uCi of[35S]methionine.Afteranadditional incubation (+)-glucosamine(SigmaChemical Co., St. Louis, Mo.). for16h,the mediumwasharvested and clarifiedby At 9hpostinfection, mediaonthe treatedplateswere
centrifugation atlow speedfor 20min. The volume replaced with 1 ml of MEM containing 20 mM D-wasreducedto 1mlbydialysisat40Cagainsta20% (+)-glucosamine, 10 MCi of [3S]methionine, and 106 solution ofpolyethylene glycol 6000 (Carbowax) in M unlabeledL-methionine. The control cells received
0.05MTris (pH 7.5),0.1MNaCl,and0.001MEDTA thesamemediumlackingD-(+)-glucosamine. After1 (TNE).The virus waspurified bycentrifugationover- h,the cellswerelysedwith SDSasdescribed above. night through a composite velocityand equilibrium Mixed infection withts mutants at30°C. Sec-gradientasdescribedbyScheele andPfefferkorn(22). ondary CEF in 60-mm plateswereinfected atanMOI
Viruswaspelletedfrom thegradientfractionsbythe of 100 with ts2 alone, ts6 alone, or with an equal
water-dilutionmethoddescribedbySefton andKeeg- mixture of ts2 and ts6 virus. After 1 h ofadsorption,
stra(28). theplateswereincubated for 7 h at300C with 5 ml of
Preparation and analysis ofglucosamine-la- MEM containing 3% FCS and then with the same
on November 10, 2019 by guest
http://jvi.asm.org/
128 BRACHA AND SCHLESINGER J. VIROL.
medium lacking amino acids for an additional 2 h. E1 E2 *E2
This mediumwasreplacedwith 1 ml of MEMlacking amino acidsbutcontaining25MCiofL-[35S]methionine 9
and 106 M unlabeled methionine. After 1 h, the
medium wasremoved, and the cellswere lysed with 8
SDSasdescribedabove. 11
Mixedinfection with ts mutants at 40°C. Pri- 7 l mary CEFin 75-cm2T flaskswereinfected at400C
,-withts2, ts6, orboth,asdescribed above. Between3 6
and 4 h postinfection, the cells werewashed exten- § l
sively severaltimeswith medium andincubatedwith 5S
4ml of MEMcontaining10%of thenormalamounts c I
of amino acids, 1% FCS, and50MCi ofL-[35S]methio- 4 nine. After an additional 5 h, the medium was re- 3
moved, andvirus waspurifiedasdescribed above.
SDS-polyacrylamide gel electrophoresis. Cell 2
extracts were reduced andalkylated priortoelectro- l
phoresis
(24).Purified viruspreparations weremixed 1 -with a loading bufferconsisting of 20 mM Tris (pH7.4), 2% SDS,5%2-mercaptoethanol,10% sucrose, and 2
bromophenolblue. Themixtureswereboiled for 5 min DISTANCE
andappliedto discontinuous slabgels prepared with
DiStAnCE
7.5or10%acrylamideaspreviously described(24).In FIG. 1. Electrophoretic pattern ofproteins from
general, 20,000 to 40,000 cpm wasadded togel slots. purified virions of ts2 and its parental HR strain. Two-dimensionalgel electrophoresis. Samples Samples of35S-labeled virus were denatured and
(20jl) ofpurified 35S-labeledHR and ts2 virus were subjected to electrophoresis in SDS-polyacrylamide mixed with an equal volume of the loading buffer slab gelsasdescribed in the text. Autoradiogramsof
described above butlacking2-mercaptoethanol. The the dried gelswerescanned inaGilford
spectropho-samplewereboiled for4minandsubjectedtoelectro- tometer at 600nm.( ) HR;
W---)
ts2.phoresisin a 7.5%acrylamide discontinuous slab gel
at 20Vfor13h.Strips containing viral proteinswere
immediatelycut from the slab gelandplaced across be expressed at 30°C as an aberrant cleavage thetop of a 10%acrylamide slab gel apparatus that thatproduces the E2 protein. To test this
model,
contained only the "running" gel. A layer of 7.5% we isolated temperature-resistant revertants ofacrylamide "stacking" gel containing 10%2-mercap- we andas nledanother ts mutant om
toethanoland 1%agarosewaspouredto fillthespace ts2 and also analyzed another ts mutant from
(1 cm) betweentherunning gel andthestrips. After complementation groupC. Thepatternsof
viral-electrophoresis at 120 V for 3 h, the gel was dried specific proteinsformedbytwo ofthe revertants under vacuum and exposed toKodak No-Screen X- showed that the E2 phenotype was retained
rayfilm. while thep140 protein, which ischaracteristicof
groupC mutants grownat40°C,disappeared in
RESULTS
cells
infected with the revertants (Fig. 2).Cells
Identification ofanaltered E2 glycopro- infected with ts5, another group C mutant, made
tein and its relationto the tsmutation. In p140 at the higher temperature and formed a
the process ofstudyingthe formation ofviral- normal E2 protein at permissive temperatures
specific proteins in cells infected with Sindbis (Fig.2). Thus, the mutation affecting
tempera-virus ts mutants, weobserved that virions pro- turesensitivitywasdistinctand separablefrom ducedbythe ts2mutant atpermissivetemper- thatleadingtothe E2phenotype.
aturesdisplayedaprotein pattern distinct from Origin of *E2. Despite an easily detectable that of theheat-resistantparental strain (HR). difference inelectrophoretic mobility of *E2, its The ts2 virions contained an E2 glycoprotein immediate precursor, PE2, displayed a mobility
(designated*E2) withasignificantly greater mo- in SDS-polyacrylamide gels identical to that of
bility inSDS-polyacrylamidegel electrophoresis thewild-type virus (Fig. 2 and 3). It is known, than was observed for the HR strain (Fig. 1). however, that glycoproteins have atypical mo-The ts2 strain belongs to complementation bilities inSDS-gelelectrophoresis (12, 30); there-group C mutants (4) and is defective in the fore weexamined the mobilities of the
nongly-posttranslationalprocessing of a high-molecular- cosylated viral membrane proteins. When
glu-weight protein (designated pl40) that contains cosaxnine was used to inhibit glycosylation, the
the sequences of the three viral structural pro- patternsofproteins from the ts2 and wild-type teins(23). At40°C,almostnoproteolytic cleav- viral-infectedcells were identical (Fig. 3, lanes 2 ageofp140isdetectedincellsinfected with ts2, and 4). Conversion of PE2 to E2 also involves
andwepostulatedthat thesamemutationmight changesinglycosylation (28), and it was possible
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.271.462.68.266.2]WT
ts
2
REVERTANTS
t
5
WT
ts2
~A
B
t
P 140
E
E2
9 ,*-...%. e,
a
b c
a
b
c
a
b c
a b
cFIG. 2. Viral-specific proteinsin cells infected with mutants ts2 and ts5 and revertants ofts2.Procedures
forelectrophoresis of samplesinSDS-polyacrylamideslab gelsand preparation of autoradiograms are in the text. (a)20-minpulseat30°C;(b) pulsefollowedby1-h chase at30°C;(c) 20-min pulse and1-h chase at 40°C.
that E2wasdeficient inglycosylgroups,thereby esis that *E2 is ashorterpolypeptide chainas a
increasingitselectrophoretic mobility. result of an aberrantproteolytic cleavage from
Glycopeptidesfrom E2and *E2 wereanalyzed anormal-sizedPE2 precursor.
after Pronasedigestion of theglycoproteins that Mobilities of viral glycoproteins in the
werepurifiedfrom virionsgrownin cellslabeled nonreduced conformation. The
electropho-with
["C]-
or[3H]glucosamine.
Theelutionpro- retic mobilities offully reduced and denaturedfiles ofglycopeptidesfromaBio-Gel P-6column proteins in SDS-gels can be used to calculate
werevirtually identical forE2and *E2(Fig. 4). molecularweights (9,31);weestimated that*E2
The three peaks eluted first (designated else- is smallerthan E2by 3,000 to4,000 daltons. A where as
Si,
S2, and S3) representthecomplex loss of about 30amino acids might haveasig-oligosaccharide
with varying amounts ofsialic nificanteffectonvirusstructure,but thereistheacid (14), and the last fraction isthe
high-man-
possibility that *E2 retains this fragmentinitsnose"simple" oligosaccharide.The
slight
differ- native state through a disulfide linkage. Weence inrelativeamountsof
Si
and S2 in *E2 as couldtestthismodel bycomparing theelectro-compared
with E2 could represent more sialic phoretic mobilities of E2 in its reduced andacidin *E2. However,therelativemobilities of nonreducedstatesandatvaryingconcentrations
the E2 and *E2
proteins
ingel
electrophoresis
ofacrylamide. Here,wedescribe the results ob-were notaltered aftertreating
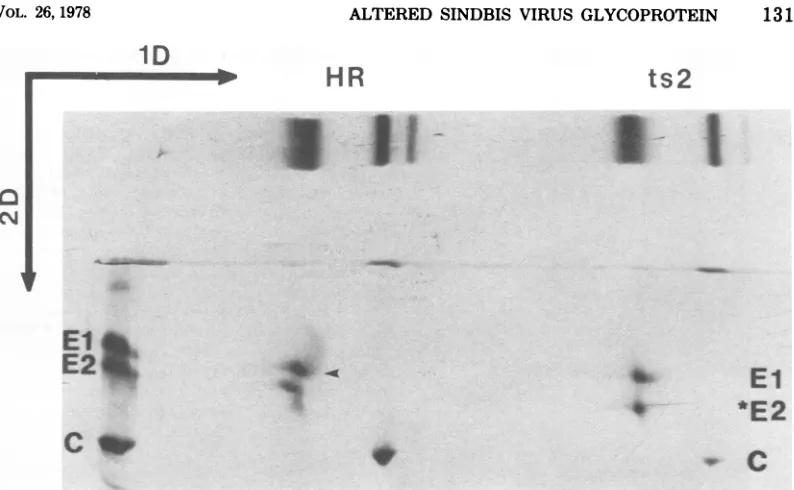
virionswithneur- tained witha two-dimensionalgelsystem (Fig.aminidase, even though all bands showed a 5). In their nonreduced forms, there is poor
slightly increased mobility as a result of the resolution between Sindbis virus glycoproteins
neuraminidase treatment(datanotshown).We Eland E2 of the HR strain(Fig.5,1D),and the
alsoassayedtherelativeamountsof
galactose
in second dimension of thegel
shows that thethe glycoproteins of the variant and wild-type
faster-moving
component of thepartially
re-virions. The E2 protein contained 45% of the solved
glycoproteins
in anonreducedgel
has atotal
galactose
recoveredin ElandE2,andthis slowermobility
after reduction(Fig.
5,arrow).
value was obtained for both E2 and *E2proteins. Thus,for the normal viral
glycoproteins
in the Allofthese dataareconsistentwith thehypoth- nonreducedstate, El hasafastermobility
thanon November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.98.379.77.342.2]130 BRACHA AND SCHLESINGER J. VIROL.
2
3N
4
' cal gels andrerunning the leading andtrailing
edges of the band after reduction with 2-mercap-toethanol.The Elglycoproteinapparently takes
on adifferent conformation after
reduction,
andthe native nonreduced form of El may be a
highlycompactdisulfide-linkedstructure.
The mobilities of nonreduced El and *E2
fromthets2strain arevirtually identical (right
portion ofFig. 5), butareclearly resolved in the
second dimension after reduction. From these
dataweconclude thatE2and*E2donot
change
their conformation significantly between the
nonreduced and reduced states and that *E2
doesnotretainthe cleaved
polypeptide
fragment
in a covalent form.
The aberrantputativeproteolytic
cleavage
ofPE2inthets2andrevertantstrainscould arise
byamutational
change
inthecleavage
site inPE2,oritmight be the resultof a
mutationally
altered
viral-specific
protease. On the basis ofresults described
below,
we consider thelatterexplanationunlikely.
2 Useof *E2as amarker in
complementa-tion. The *E2
phenotype
hasprovided
auseful,.-
_=~
marker formeasuring the expression ofts2viralE
I structural genesduring
acomplementation
re-2 action between ts2 and other ts mutants. For
*---
E
thisstudy
wechosets6,
anRNA-tsmutant(3)
____* E that
complements
verywelltheRNA'
ts2 defect*E
, (4). We first examined the viral proteins fromcellscoinfected with
equal
numbers ofthemu-tant virions and incubated at the permissive
2000- -200
500 I'I' 15..1
S 6
T-f
000o
oo0
FIG. 3. Effect of glucosamine on
electrophoretic
/Oo
ispatternofviralproteins in cellsinfectedwith wild-typeorts2 virus.Experimentaldetailsarein thetext.
(1)Nornal cellsinfected with wild-type virus;(2) cells
treated with 20 mMglucosamine and infectedwith
wild-type virus; (3) normal cells infected with ts2 40 50 60 70
virus; (4)glucosamine-treatedcells infectedwith ts2 FRACTION NUMBER
virus. Thenonglycosylated proteins are located be- FIG. 4.
Profile of glycopeptides from
E2 and *E2tweenthe normalglycosylatedEl and E2.
glycoproteins.
Pronase-digested
sampleswere sepa-rated on a Bio-Gel P-6 column according to theprocedure of Sefton and Keegstra (28). Details of E2,but Elisslower than E2 afterreduction.
labeling
the proteins with[14C]-
and[3H]glucosa-We have confirmed this property ofEl by mine are in the text. E2 was labeled with
'4C-
*E2elutingthepoorlyresolvedbandsafterelectro- was labeled with
[3HJglucosamine.
About 10 timesphoresisof thenonreducedproteins in cylindri- more 3H counts were used than 14C in the analysis.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.66.258.56.530.2] [image:5.504.271.460.405.588.2]1D
HR
ts2
04
El
E2
~~~~~~~~~~El
*~*E
CCM
c~~~~~~~~~~~~~~~~~~
go
FIG. 5. Autoradiogram ofa two-dimensionalSDS-polyacrylamide gel electropherogram containing
'S-labeled virionproteins. Preparation of samplesandproceduresforthisgelsystemaredetailed in thetext.
Thefirst dimension(ID)contains nonreducedproteins. The second dimension (2D)wassubjectedto
electro-phoresisin thepresenceof mercaptoethanol. On thefar leftisthepattern forasetofreduced andalkylated
viralproteins.
temperature.
Equivalent
amounts ofboth viral Thecis-dominance andnonpreferentialselec-structuralgene
products
werefound(lane
c,Fig.
tion in viral budding of the *E2 gene product6), while cells infected with
only
one of the allowedus toanalyze
the expression of the ts2mutants revealed the
appropriate
phenotype
genomein cells coinfected withcomplementing
(lanesaand
b, Fig.
6). Therewasnorestriction mutantsatthenonpermissive
temperature.La-on ts2 or ts6viralRNA
expression
inthemixedly
beled virusparticles
wereisolated from theme-infected cells
but,
moreimportantly,
the aber- diumof the coinfectedcells, and thepattern ofrant
cleavage
reactionyielding
the *E2was co- viral proteins was displayed inSDS-polyacryl-dominant with that
producing
E2. Ifachange
amide gels(Fig.7).Of thelabel foundinthetwohad occurredin a
viral-specific
protease coded viral E2proteins,
morethan 90%appearedas aby the ts2 genome,we
might
haveexpected
allnornal
migrating
E2band(lane
c,Fig.
7).
In theE2
protein
to be altered in thecoinfected cell. coinfectedcells at400C,
more than 80% oftheThus *E2acts as acis-dominant trait that
prob-
labelwasfound inp140 (data
notshown),
indi-ably
reflects achange
in ts2 PE2protein
se- cating that thets2-specific
mRNA wastrans-quence. In
addition,
we have examined cells latedat400C.
Thus, themostlikely explanationcoinfected with the HR and Rts2a Sindbis virus for the failuretodetect
larger
amountsof*E2isstrains todetermine whether there was a
pref-
that thecomplementation
between ts2 and ts6erential selectionforthe E2or*E2
glycoproteins
isnonreciprocal.
The ts2 genome canprovide
during
viral maturation andbudding.
Coinfected the functionmissing
in ts6(possibly
an RNACEF
monolayers
at370C
werelabeled with[35S]
polymerase),
butts6cannotcorrectthetemper-methionine a 4 h
postinfection,
and virus and ature-sensitive defect in the ts2genome;other-cellswereharvested4hlater. Proteins extracted wise,the structuralproteinsencoded
by
ts2(ex-from the cells and from
purified
virions were hibited by *E2) would have been found in theseparated
by
SDS-polyacrylamide gel
electro- virions secreted from the coinfected cells(Fig.
phoresis,anddensitometricscanswere
prepared
8).
The small amount of *E2 is attributed tofrom
autoradiograms
of the slabgel electropher-
leakage
ofthetsmutation. ograms. The ratio oflabeled E2 to *E2 in the DISCUSSION virions wasalmost identicaltothatmeasured inthecells(0.51and
0.53,
respectively).
These data Wehave described here anaberrant form of showthat neither E2nor*E2waspreferentially
the Sindbis virus E2glycoprotein
that differsutilized
during
thebudding
process. fromthenormal
protein
by
anincreasedon November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.44.441.54.299.2]132 BRACHA AND SCHLESINGER J. VIROL.
ridechains (28). Our data on the glycopeptides in E2 and *E2 indicate that both contain the same amount of the Aand B type oligosaccha-rides.
.n4 Theglycosylated and nonglycosylated
precur-P
14'
sorsto*E2 in thets2strainhadelectrophoretic
mobilities identicaltothose of theparentalHR strain. These data leadustoconclude that *E2 results fromamutation that has altered the site for proteaseactivity onthePE2 precursor.The mutation is distinct from the one that conferstemperaturesensitivity to thereplication ofts2
virus, eventhough the ts mutation itself involves
adefect in another
proteolytic cleavage step
inviral structural-protein formation (22). The
other product of the PE2 cleavage has been
designated asE3 for the closelyrelated Semliki
Forest virus (11), but has not been detected in
Sindbis virions. It appears in the medium of
aU
:<
1E-1
~muII
FIG. 6. Viralproteins incells cointectedwith ts2 andts6 virus at 30°C. Details ofcelllabeling and
preparation of the autoradiogram after
SDS-poly-acrylamide gel electrophoresis are in the text. (a)
Infection withts2alone; (b) ts6alone; and (c) both
together.
phoretic mobility in SDS-polyacrylamide gel.
This trait isgenerallyattributed to a decrease in the size ofapolypeptide, althoughit could also indicate alower amount ofcarbohydrate in the
glycoprotein.Based onthe difference in
mobili-ties, *E2 was estimated to be 3,000 to 4,000 daltons smaller than E2. Can thisdifference be attributed to only a change in carbohydrate? Sindbis virus glycoproteins deficient in carbo-hydrateby virtue of growth incellsaltered in a
glycosyl transferase activity (25) or treated with ja b
C.
inhibitors ofglycosylation activities (8, 16) showincreasedmobilities,and in the former case the FIG. 7. Structuralproteinsfromvirionsformed by alteredproteinwasestimated todiffer in molec- cells coinfected with ts2 and ts6 virus at40°C. Refer
ularweight by800. The totalamount ofcarbo- to the text for the preparation of these 35S-labeled
hydratein eachglycoproteinofSindbis virus has samples and the gel electropherogram. (a) HR
paren-been estimated to be 4,000 daltons (28), and tal
virus;
(b) ts2virus
formed at30°C; (c) virus fromanalyseshaveshownthtvs
gcoinfection.
Plaque assays gave thefollowing virusanalyses have shown that virus grown in chick titers after 10 h of
infection
at40°C: ts2, 4.2 x10i
cellshas E2 glycoproteins that contain a"com- PFU/ml; ts6, 7.5 x
10i
PFU/ml; ts2 +ts6,
1.4 x 109plex" A type and "simple" B type oligosaccha-
PFU/ml.
on November 10, 2019 by guest
http://jvi.asm.org/
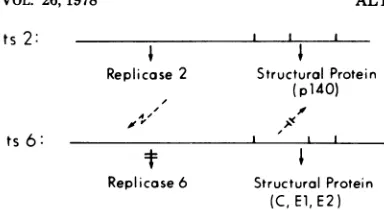
[image:7.504.88.230.70.389.2] [image:7.504.267.458.273.574.2]ts 2: ' ' ' which led to noninfectious, atypical, and
heter-i
StI
ogeneous particles (21). Results from trypticReplicase 2
StructuralPr(4O)
peptide
analyses
of the alteredproteins
estab-p lished that the ts defect caused an incorrect
le A cleavage ofthePR76 precursorprotein, but the
ts 6: I I precise nature of the mutation was not
deter-t
+ mined.Replicase 6 Structural Protein Variations in the
posttranslational
cleavage
(C, El, E2) products have also been
reported
fortsmutantsFIG. 8. Modelfornonreciprocalcomplementation.
of
poliovirus
(6). A putative alteration in apro-The ts2 virus supplies polymerase complex for both teolytic cleavage site was found in reovirus ts ts2 and ts6 26S RNA production, but ts6 cannot mutants, but the
genetically
stable defect pro-correct the ts2defect in cleavage.ducing
anelectrophoretically
altered,2 viralstructural protein could be geneticallyseparated
from the ts mutation inseveral of the reovirus infected cells (D. Brown, personal communica- mutantclassesandshoweditselfnot to be a ts
tion).Wehavenotdetectedthepredicted larger defect(7).Thisnondefective alterationprovided
E3from themutantstrain. Crossand Fields(7) witha thirdgenetic marker
The shortened *E2 glycoprotein does not for evaluating interactions between gene
seg-seem tohave grossly affected the structure or ments during reovirus
replication.
Analyses of function of the virion. ts2virus wasreportedto thetemperature-resistant"recombinant" clonesbe slightlymore thermolabilethan ts5 (3), and for the,u2 phenotype provedthatreassortment
wefoundnodifferenceinsensitivitytorepeated of RNA segments during reovirus replication
freeze-thawingin ts2 ascomparedwith theHR was arandomevent.Further studies located the
orwild-type strains
(unpublished
data).Asingle- ,u2 changeto the RNA A segment, but noaddi-step growth cycle ofts2 was identical to other tional information has been obtained on the
Sindbis strains when tested at
300C
(unpub- biochemical nature of the defect. Anotherreo-lished data).Wehavenotcarefully analyzedthe virusmutant had an
electrophoretically
alteredhost range or antigenic behavior of the *E2 viral protein,
X3.
Interestingly, bothX3
and,u2variant, but no differenceswere seen inplaque aremajor surfacecomponents ofthevirion,and
size on CEF cells. BHKcells infected with ts2 these
capsid polypeptides
mayalso differamongproducedan*E2 thatwasindistinguishablefrom thereovirus serotypes(20).
*E2 from the chick fibroblasts. On the virus In amanner somewhat analogous to the
reo-surface, E2 glycoprotein appears to be more viruswork noted above, we have used the
elec-exposed than El (29). Our analyses of thenon- trophoretically distinct *E2 mutation as a
phe-reduced forms of these glycoproteins suggest notypic marker in examining gene expression
thatEl maybein a
"tighter"
conformationinduring
thecomplementation
reaction betweenits native state, possibly the resultofdisulfide two tsSindbis virus mutants.Ourresultsshowed
loops. that complementation wasnonreciprocal-that
Abnornalproteolyticcleavages have beende- is, theRNA'ts2 mutant couldcorrect the defect
tectedinthereplicationofseveral animal virus of the RNA- ts6, but ts6 gene products were
systems, and
apparently they
are widespread. unable to correct the RNA' defect. OnlytraceThe ambiguities detected in posttranslational amounts of the ts2 viral gene products were
cleavageof
picornavirus polyprotein
precursors detected in virions formedduring
complemen-may accountfor differencesinserotyping of the tation at the
nonpermissive
temperature. Our viruses. For example, type 1 Brunhildepolio-
analysesofcellscontaining
both thets2and ts6virus is distinguished from type 2 strain by a viralgenes showed that ts2mRNAwas
function-differenceinthe sizesof
capsid proteins
VP2 and ingandproducing the 140,000-daltonprecursor.VP3(1). These
proteins
areproducts
ofacleav- Thus, thetsdefectisacis-dominant trait.Iftheage in precursor3a, andtype1iscleavedto
give
ts2temperature-sensitive
defectwere inaviralasmaller VP3 anda
concomitantly larger
VP2 codedprotease,wemight
haveexpected
thetraitthanfoundinthetype2virus. Alterationswere to be recessive in a trans
configuration,
andaalsodetected in those
cleavages
thatyield
non- nondefectivets6protease should have been ablestructural
polypeptides
in thetype2strain. Var- to cleave the ts2 precursor. We have made aniations have been found in the
pathways
of analogousargument for the *E2 mutation sinceposttranslational processing of rhinovirus spe- thisdefectwasretainedin amixedinfection of
cificproteins(17).A tsmutantof aviansarcoma twomutants, oneof which carriedanormal E2. virus produced a different set of viral "core" cis-dominant mutations here are most
easily
proteins at the nonpermissive temperature,
explained by
alteration ofsequence andon November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.48.240.58.163.2]134 BRACHA AND SCHLESINGER J. VIROL.
tureof precursor
polypeptides.
Our results differ and evidence foracomplex ofPE2-E1 viralglycopro-fromthose
recently
published by Scupham
etal. teins.Virologyr
74:441-449.3.Burge, B.W., and E.
R.
Pfefferkorn.1966.
Isolation(27), who notedadisappearanceof the ts2pro- andcharacterization of conditionallethal mutantsof
tein incells coinfected with thecomplementing Sindbis virus.Virology30:204-213.
ts2O mutantand
suggested
that aviral-specific
4. Burge, B. W., and E. R. Pfefferkorn.1966.Comple-protease from the ts2O was acting on the ts2 mentation between temperature-sensitive mutantsof
polypeptid.Wewouldarguethat theirputative 5. Sindbis virus.Virology30:214-223.
polypeptide. wewoula argue tnat tnelr putatlve 5. Cancedda, R., R. Swanson, and M. J.Schlesinger. proteaseis unableto convert ats2precursorto 1974. Viral proteins formed in a cell-free rabbit
reticu-properly
sized viral membraneproteins. For the locytesystemprogrammed with RNAfromatempera-ts2 temperature-sensitive defect, the cis-domi- ture-sensitive mutant of Sindbis virus. J. Virol.
nantcaracteisticsconsstent ith th dem- 14:664-671.
nant characteristic is consistent with the dem- 6. Cooper, P. D., D. F.
Summers,
and J. V.Maizel.
1970.onstration that the mutation was expressed in Evidence for ambiguity in post translational cleavage of
an in vitro protein-synthesizing system (5, 32). poliovirusproteins.Virology41:408-418.
However, one intriguing model that our data 7. Cross, R. K., and B. N. Fields. 1976. Use of anaberrant
exclude is a cis-dominant sf polypeptideas amarker in three-factorcrosses:further
cannot totally evidence for independent reassortment as the
mecha-proteolytic activity expressed by the virus capsid nism of recombination between temperature-sensitive polypeptide as it is translated from the 26S mutantsof reovirus type 3.Virology 74:345-362.
mRNA. If thistype ofactivitywere altered in 8. Duda, E., and M. J.Schlesinger. 1975. Alterationsin
,thenafunctional self-protease in ts6 capsid
Sindbis
viralenvelope proteins bytreatingBHKcells
ts2, tne a nclonl slrproeas mtsocapla withglucosamine. J. Virol.15:416-419.
mightactpoorlyifatall withthenascentpoly- 9. Dunbar, A.K., andR.R.Rueckert.1969. Observations
peptide translated from the ts2 mRNA. There onmolecularweightdeterminationsonpolyacrylamide
are now several reports
suggesting
that post- gels. J.Biol. Chem. 244:5074-5080.translational cleavage products ofvirus genes 10
Fennell, RI,
andB. A.Phillips.1974.Polypeptidecom-position ofurea-andheat-resistantmutants of
polio-haveproteolytic activity (15, 33). virus types 1 and 2. J. Virol.14:821-833.
The nonreciprocal contribution of the differ- 11.Garoff,HI,K.Simons,and0.Renkonen. 1974.
Isola-ent genomesdiscovered herecanofferanexpla- tion andcharacterizationof the membraneproteinsof
nation foraresult described
by
Pfefferkor and 12. Grefrath, S. P., and J. A.Semliki Forest virus.VirologyReynolds. 1974.61:493-504.
Themolec-Burge (19)intheir initial studiesoncomplemen- ular weight of the major glycoprotein from the human
tation with the tsmutants.Theyobservedthat erythrocytemembrane.Proc. Natl.Acad. Sci. U.S.A.
the yield of virus obtained fromamixed infec- 71:3913-3916.
tinofRNA'and RNA- mutantwasverysen- 13. Jones,K.J.,M. R. F.Waite,and H. R.Bose. 1974.
tion of RNA andRNA mutant was very sen- Cleavageofaviralenvelope
precursor
duringthemor-sitive to the MOI of the RNA- but insensitive phogenesis of Sindbis virus. J. Virol.13:809-817.
tothe MOI of theRNA'.Fromourresults, only 14. Keegstra,K., B.Sefton,and D. Burke.1975.Sindbis the RNA- isprovidingthestructuralgeneprod- virus glycoproteins: effect of the hostcellonthe
oligo-ucts, and alimitation onthe amount of these
saccharides.
J.Virol.
16:613-620.
15.Lawrence,C., andR. E.Thach.1975.Identification of
structural proteins would be expected to limit a viral protein involved inpost-translationalmaturation
virion production. In contrast, RNA' mutants of theencephalomyocarditisviruscapsidprecursor.J. are
presumed
toprovide
geneproducts
thatact Virol.15:918-928.catalytically
during complementation.
16. Leavitt, R., S. Schlesinger, and S.Kornfeld.
1977.Tunicamycin
inhbits
glycosylation andmultiplication
More detailed structuralanalysesofthe *E2 of Sindbis and vesicular stomatitis viruses. J. Virol.
variant may allowus to identify the region in 21:375-385.
the
polypeptide
of normal E2 that has been 17. McLean, C., T. J. Matthews, andR. R.Rueckert.
1976.changed,
and may further help to identifythe Evidenceof ambiguous processing and selectivedegra-changed, dation in thenoncapsid proteinsofrhinovires 1A. J.
substrate
specificity
for theimportant
proteo-Virol.
19:903-914.lytic cleavage that is essential for budding of 18.Pfefferkorn,E. R., and M. K. Boyle. 1972.Selective
Sindbisvirus. inhibition of thesynthesisof Sindbisvirionproteinsby aninhibitorofchymotrypsin. J. Virol. 9:187-188. ACKNOWLEDGMENTS 19. Pfefferkorn, E.R., and B. W. Burge. 1967. Genetics We thank Sondra Schlesinger and Janice Brielmeier for and biochemistry of arbovirus temperature-sensitive assistanceinpreparation andanalysis of the glycopeptides. mutants, p. 403-426. In J. S. Colter and W.Paranchych Thisinvestigation wassupported by Public Health Service (ed.), The molecular biology of viruses. Academic Press grants CA-14311-04 and CA-16217-03fromtheNational Can- Inc., New York.
cerInstitute. 20. Ramig, R. F., R. K. Cross, and B. N. Fields. 1977. Genome RNAsandpolypeptides of reovirus serotypes LITERATURE CIMD 1, 2, and 3. J. Virol. 22:726-733.
21. Rohrschneider,J.M., H.Diggelmann, H. Ogura, R. 1. Beckman,L.D., L A.Caliguiri,andL.S.Lilly.1976. R.Friis,and H. Bauer. 1976. Selective cleavage of a Cleavage site alterations inpoliovirus-specific precur- precursor polypeptide in atemperature-sensitive mu-sorproteins.Virology73:216-227. tant ofavian sarcoma virus. Virology 75:177-187. 2. Bracha,M., and M. J. Schlesinger. 1976. Defects in 22. Scheele, C. M., and E. R. Pfefferkorn. 1970.
Virus-RNA'temperature-sensitive mutantsofSindbis virus specific proteins synthesized in cells infected with
on November 10, 2019 by guest
http://jvi.asm.org/
RNA' temperature-sensitive mutants of Sindbis virus. Sindbis virus:preliminary characterization of the oli-J. Virol.5:329-337. gosaccharides. J. Virol. 14:522-530.
23. Schlesinger, M. J., and S. Schlesinger. 1973. Large- 29. Sefton, B. M., G. G. Wickus, and B. W. Burge. 1973. molecular-weight precursors of Sindbis virus proteins. Enzymatic iodination of Sindbis virus proteins. J. Virol.
J.Virol.11:1013-1016. 11:730-735.
24. Schlesinger, M. J., S.Schlesinger, and B. W. Burge. 30. Segrest, J. P., R. LJackson, E. P.Andrews, and V. 1972.Identification of a second glycoprotein in Sindbis T.Marchesi. 1971. Human erythrocyte membrane gly-virus.Virology 47:534-541. coprotein: areevaluation of the molecular weight as 25. Schlesinger, S., C. Gottlieb, P. Feil, N. Gelb, and S. determined by SDSpolyacrylamide gel electrophoresis.
Kornfeld. 1976.Growth ofenveloped RNA virusesin Biochem.Biophys. Res. Commun. 44:390-395. alineof Chinesehamsterovarycells with deficient N- 31. Shapiro, A. C.,E.Vinuela, and J. V. Maizel. 1967. acetyl-glucosaminyltransferase activity. J. Virol. 17: Molecularweight estimation ofpolypeptide chainsby
239-246. electrophoresis inSDS-polyacrylamide gels. Biochem.
26. Schlesinger,S., and M. J. Schlesinger. 1972. Forma- Biophys. Res. Commun. 28:815-820.
tion ofSindbisvirusproteins: identification ofaprecur- 32. Simmons, D. T., and J. H. Strass. 1974. Translation of sor for one of the envelope proteins. J. Virol. Sindbis virus 26S RNA and 49S RNA in lysates of 10:925-932. rabbit reticulocytes. J. Mol. Biol. 86:397-409. 27. Scupham, R. K., K. J.Jones,B. P.Sagik,and H. R. 33. VonderHelm, K.1977.Cleavage of Rous sarcoma viral
Bose, Jr.1977.Virus-directedpost-translational cleav- polypeptide precursor into internal structural proteins agein Sindbis virus-infectedcells.J.Virol.22:568-571. in vitro involves viralproteinp. 15. Proc.Natl. Acad. 28. Sefton, B.M., and K. Keegstra.1974.Glycoproteinsof Sci. U.S.A.74:911-915.


![FIG.FRACTIONwasratedprocedureglycoproteins.morelabelingmine 4. Profile of glycopeptides from E2 and *E2 Pronase-digested samples were sepa- on a Bio-Gel P-6 column according to the the of proteins Sefton and with Keegstra [14C]- and (28)](https://thumb-us.123doks.com/thumbv2/123dok_us/1531549.105753/5.504.271.460.405.588/fractionwasratedprocedureglycoproteins-morelabelingmine-profile-glycopeptides-pronase-digested-according-keegstra.webp)