Type 1 Nef Sequence Motifs Important for Its Immune Evasion
Function
In Vivo
Martha J. Lewis,a,bPatricia Lee,a,bHwee L. Ng,a,b,cand Otto O. Yanga,b,c
Department of Medicine, Division of Infectious Diseases,a
UCLA AIDS Institute,b
and Department of Microbiology, Immunology, and Medical Genetics,c
Geffen School of Medicine at UCLA, Los Angeles, California, USA
Human immunodeficiency virus type 1 (HIV-1) Nef downregulates major histocompatibility complex class I (MHC-I), impair-ing the clearance of infected cells by CD8ⴙcytotoxic T lymphocytes (CTLs). While sequence motifs mediating this function have been determined byin vitromutagenesis studies of laboratory-adapted HIV-1 molecular clones, it is unclear whether the highly variable Nef sequences of primary isolatesin vivorely on the same sequence motifs. To address this issue,nefquasispecies from nine chronically HIV-1-infected persons were examined for sequence evolution and altered MHC-I downregulatory function under Gag-specific CTL immune pressurein vitro. This selection resulted in decreasednefdiversity and strong purifying selec-tion. Site-by-site analysis identified 13 codons undergoing purifying selection and 1 undergoing positive selecselec-tion. Of the for-mer, only 6 have been reported to have roles in Nef function, including 4 associated with MHC-I downregulation. Functional testing of naturally occurringin vivopolymorphisms at the 7 sites with no previously known functional role revealed 3 muta-tions (A84D, Y135F, and G140R) that ablated MHC-I downregulation and 3 (N52A, S169I, and V180E) that partially impaired MHC-I downregulation. Globally, the CTL pressurein vitroselected functional Nef from thein vivoquasispecies mixtures that predominately lacked MHC-I downregulatory function at the baseline. Overall, these data demonstrate that CTL pressure exerts a strong purifying selective pressure for MHC-I downregulation and identifies novel functional motifs present in Nef sequences
in vivo.
T
he human immunodeficiency virus type 1 (HIV-1)/ simian immunodeficiency virus (SIV) accessory protein Nef, an abundantly expressed 27-kDa myristoylated protein, is not essen-tial for viral replication but is central to pathogenesis (reviewed in references21and48). This protein plays a key role in viral persis-tence and virulence. In humans, infection with Nef-defective HIV-1 has been associated with low-to-undetectable levels of viremia with vigorous antiviral immunity and delayed disease progression (14,18,19,31,32,34, 44). Similarly, experimental infection of rhesus macaques with SIV in which Nef has been deleted (SIV239⌬nef) results in low-to-undetectable levels of viremia, asymptomatic infection, and protection from subsequent challenge with wild-type virus (17). This model system has been considered the gold standard for a disease-attenuating vaccine model.Although numerous functions have been attributed to Nef, the mechanisms whereby Nef exerts these dramatic clinical effects ap-pear to involve its ability to direct immune evasion. While Nef initially was misunderstood as a negative transcriptional activator (2,45), further work has shown that it contributes to viral patho-genesis through multiple functions that enhance viral infectivity, such as downregulation of CD4 on the surface of infected cells (24,
37) and modulation of cellular activation (8,9,56,58,61). Fur-thermore, it is well established that Nef downregulates major his-tocompatibility complex class I (MHC-I) cell surface proteins (12,
13,60).In vitromodels demonstrate that Nef-mediated MHC-I downregulation impairs cytotoxic T lymphocyte (CTL) recogni-tion and clearance of infected cells (1,13,63,68), suggesting that it plays a central role in immune evasion.
In vivoevidence also suggests that this function is important for immune evasion. Rhesus macaques infected with SIV containing
Nef rendered specifically defective in MHC-I downregulation function via difficult-to-revert mutations showed trends for higher CTL levels and lower viremia in the first 14 weeks of infec-tion followed by viral rebound accompanied by a striking pattern of Nef evolution to reconstitute this function via new sequence motifs resembling those in HIV-1 (62). In chronically HIV-1-infected humans, Nef has been shown to lose function in persons with severely depressed cellular immunity due to very young age (25, 65) or late-stage AIDS (11, 33), and more specifically, its MHC-I downregulatory function correlates to the breadth of the HIV-1-specific CTL response during chronic infection (40). These data strongly suggest the importance of this function in the immunopathogenesis of infection by reducing the clearance of virus-infected cells by CTLs. Moreover, the variability of Nef func-tion during chronic infecfunc-tion suggests that it evolves to optimize its balance of different functions to maximize viral persistence in the face of changing selective pressuresin vivo(40).
Mutational studies of Nef in laboratory strains of HIV-1 have defined key amino acid sites and functional domains involved in the downmodulation of MHC-I (reviewed in references26and
47). However, the sequence of Nef is highly variable in primary isolates of HIV-1. It is likely that Nef can adapt to downregulate MHC-I through altered or distinct motifs, depending on its
se-Received9 April 2012Accepted21 April 2012
Published ahead of print2 May 2012
Address correspondence to Martha J. Lewis, [email protected].
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.00878-12
on November 7, 2019 by guest
http://jvi.asm.org/
quence context, as seen in the SIV model (62). However, few stud-ies have addressed the ability of Nef from primary isolates of HIV-1 to downregulate MHC-I (46), and there is almost no infor-mation about whether the functional motifs of primary isolates of Nef match those identified by mutagenesis of laboratory-adapted strains of HIV-1.
To address this issue, we investigated the interplay between the MHC-I downregulatory function of primary isolate quasispecies Nef proteins and sequence evolution under experimentally im-posed selective pressure to evade Gag-specific CTLs. This selective pressure caused a clear pattern of purifying selection coincident with the optimization of MHC-I downregulation to allow viral persistence in the presence of CTL selective pressure. Sequence analysis of this adaptive evolution identified key amino acid sites important for Nef-mediated immune evasion in primary HIV-1 isolates, demonstrating the close reciprocal relationship between Nef and CTL-mediated immunity in pathogenesis and suggesting vulnerable regions that could be targeted beneficially by vaccines or pharmacologic blockade.
MATERIALS AND METHODS
Isolation of plasmanefquasispecies and insertion into recombinant reporter viruses.Thenefgene was amplified from the plasma of 9 chron-ically HIV-1-infected subjects and cloned into an NL4-3-based reporter virus as previously published (40). Briefly, cDNA was made from viral RNA using the gene-specific primer Nef 9589R (5=TAGTTAGCCAGAG AGCTCCCA). Thennefwas amplified using primers Nef 9589R (5=TAG TTAGCCAGAGAGCTCCCA), Nef 8670F (5=AATGCCACAGCCATAG CAGTG), Nef 8675F (5=GCAGTAGCTGAGGGGACAGATAGG), Nef 8687F (5=GTAGCTCAAGGGACAGATAGGGTTA), and Nef 8736F (5= AGAGCTATTCGCCACATACC). A nested PCR was performed with primers Nef 8787 XbaIF (5= GCTCTAGAATGGGTGGCAAGTGCT CAA) and Nef 9495R (5=TTATATGCAGCATCTGAGGGC). Following amplification, overhanging A’s were added to the ends of the PCR prod-ucts, which were then cloned in bulk by the TA method into the pCR2.1-TOPO vector (Invitrogen). Ligation mixtures were grown in liquid cul-ture and not subjected to individual colony selection on solid medium in order to preserve the quasispecies mixture of cloned PCR products. Plas-mid DNA was digested with XbaI and BspEI (New England BioLabs) and subsequently subcloned into thenefposition of the half-genome con-struct p83-10 (4). Ten micrograms of each half-genome plasmid, p83-10 withnefand the reporter p83-2-HSAxVpr (4), was digested with EcoRI (New England BioLabs). Both plasmids electroporated into 10 million T1 (174⫻CEM.T1) cells (57) using a GenePulser Electroporator (Bio-Rad). Recombinant reporter virus stocks were collected in the supernatant 7 to 10 days after electroporation. Control viruses carrying the Nef mutant M20A unable to downregulate MHC-I (3) or standard NL4-3 Nef were made in parallel.
In vitroimmune selection.One million T1 (HLA-A*02-positive) lymphocytes (see above) were infected with a virus stock containing 12.5 ng of p24. This is equivalent to a multiplicity of infection of approximately 0.05 to 0.1 based on previous titers. After infection for 4 h at 37°C, cells were washed and split into two wells of 0.5⫻106each. Then either 0.5 ml
of RPMI supplemented with 10% fetal calf serum and 50 U/ml interleu-kin-2 (IL-2; R10-50) or 0.5 ml of R10-50 with an HLA-A*02-restricted CTL clone specific for the p17 Gag epitope SL9 was added to the infected cells at an effector-to-target ratio of 1:4. Culture supernatant was collected on days 5 and 7 postinfection, and virus growth was quantified by p24 antigen enzyme-linked immunosorbent assay (ELISA). These p24 levels were used to set up a second round of infections again with 12.5 ng of p24 and 1⫻106fresh T1 cells, and selection was performed as before with the
same CTL clone. The first-round virus cultured in the presence of the CTL clone was again cultured with the clone, and as a control for genetic drift, the viruses cultured without CTL selection were also cultured again
with-out CTL selection. Again, culture supernatants containing the selected quasispecies were collected on days 5 and 7 postinfection and quantified by p24 ELISA to confirm viral growth.
RNA isolation, reverse transcription-PCR, andnefsequencing.Viral RNA was isolated from either the viral stock (i.e., the input virus) or culture supernatant after two rounds of culture with or without the CTL clone. RNA was isolated using the QIAamp viral RNA minikit (Qiagen) according to the manufacturer’s protocol and then used as a template for cDNA synthesis using SuperScript III reverse transcriptase (Invitrogen) and the gene-specific primer Nef 9589R (5=TAGTTAGCCAGAGAGCT CCCA). The resulting cDNA was used as the template fornef amplifica-tion using the high-fidelity polymerase Phusion (New England BioLabs) and primers Nef 8787 XbaIF (5=GCTCTAGAATGGGTGGCAAGTGCT CAA) and Nef 9495R (5=TTATATGCAGCATCTGAGGGC). PCRs were carried out under the following conditions: 5 min at 98°C; 35 cycles of 98°C for 10s, 57°C for 30s, 72°C for 30s; and a final extension at 72°C for 10 min. A 20-min incubation at 72°C with standardTaqpolymerase (New England BioLabs) and deoxynucleoside triphosphates added the neces-sary overhanging A’s, and the PCR products were then cloned in bulk by the TA method into the pCR2.1-TOPO vector (Invitrogen). A minimum of 10nefclones per subject were selected for sequencing using standard vector primers M13F and M13R and BigDye Terminator Reaction kit 3.1 (Applied Biosystems). Cycle sequencing products were run on an ABI 3130 Genetic Analyzer (Applied Biosystems).
Sequence analysis.Nucleotide sequences were translated into amino acid sequences and manually edited using the program BioEdit and then aligned along with NL4-3 and the Los Alamos HIV-1 database clade B consensusnef using CLUSTAL X. A neighbor-joining tree was con-structed using the DNADist and Neighbor programs of PHYLIP 3.64 (22). The tree was statistically evaluated with 1,000 bootstrap replicates. The sequences were then divided into three separate populations—input, with CTL, and without CTL selection—for the subsequent analyses. Sequence diversity within the quasispecies swarm and overall divergence from clade B consensus sequence were determined using the program SENDBS with the Hasegawa plus gamma model, and standard errors were estimated from 500 bootstrap replicates. Changes in diversity and divergence were calculated by taking the value for the with-CTL population minus the value for the no-CTL population. Differences between control and se-lected sequences were evaluated with a two-tailedttest. Divergent se-quences were examined for G-to-A hypermutation using Hypermut 2.0 from the Los Alamos National Laboratory (LANL) HIV-1 database tools. Sequences with nonintact reading frames due to frameshift or nonsense mutations were counted and excluded prior to the analysis for adaptive evolution. Differences in the number of sequences containing stop codons between control and selected sequences were evaluated by a two-tailed2
test. All of the following analyses were performed using HyPhy (50). The program MODELTEST (52) was used to determine that the best-fitting model for the data was HKY85. The global ratio of the rate of nonsynony-mous-to-synonymous changes (dN/dS ratio) and its 95% confidence in-terval (CI) were estimated after the maximum-likelihood function for each of the three data sets had been built and optimized. Individual amino acid positions with evidence of adaptive evolution were identified by three separate methods, ancestor counting (SLAC), random-effects likelihood (REL), and fixed-effects likelihood (FEL). A site was considered to be adapting under CTL selective pressure if that site was identified by at least two of three methods with a significance level of at least 95% and was only identified in the data set with CTL and not in the without-CTL data set. Additionally, only those sites with a dN/dS ratio significantly⬎1 or⬍1 were considered positive. Selected sites were highlighted on the composite crystal structure of Nef kindly provided by Art F. Y. Poon (Vancouver, British Columbia, Canada) using the program RasMolhttp://www.umass .edu/microbio/rasmol/. Conservation of the selected sites was determined by compiling an amino acid alignment of all of the complete, nonrecom-binant Nef sequences submitted to the LANL HIV-1 Sequence Database
on November 7, 2019 by guest
http://jvi.asm.org/
through 2010 (n⫽2,114), including genotypes A to K. The probability of each amino acid at the selected sites was plotted using WebLogo3 (16).
Creation ofnefmutants by site-directed mutagenesis.The seven sites undergoing purifying selection with no previously known associa-tion with Nef funcassocia-tion were selected for site-directed mutagenesis. The following eight mutations were created individually within the NL4-3-based p83-10 plasmid using the appropriate primers and the QuikChange XL-II kit (Stratagene): N52A, N52S, A84D, Y135F, G140R, S169I, H171A, and V180E. The amino acid changes selected were based on mutations observed at these sites in the primary isolates, except H171A. All muta-tions were confirmed by sequencing. Recombinant reporter viruses were created by coelectroporation with p83-2 HSAxvpr as detailed above.
Assessment of MHC-I downregulation by Nef.Levels of HLA-A*02 on the surface of cells infected with Nef recombinant reporter viruses were determined as previously described (40). Briefly, T1 cells were infected with either the input virus stock or the supernatant containing the quasi-species surviving after two rounds of CTL selection. All nine input viruses were tested, and five of nine samples after two rounds of culture with CTL yielded adequate samples for testing. Similarly, T1 cells were also infected with the eight Nef mutants. On day 5 postinfection, cells were stained with fluorescein isothiocyanate-conjugated anti-murine CD24 (HSA) (BD) to detect reporter-positive infected cells and phycoerythrin-conjugated anti-human HLA-A*02 (ProImmune). At least 2⫻104live cells were counted
using a FACScan flow cytometer, and data were analyzed using CellQuest software (BD). Maximum levels of HLA-A*02 were determined using the mean fluorescent intensity (MFI) of the M20A Nef mutant, which is de-fective in MHC-I downregulation or Delta Nef virus. Percent HLA-A*02 downregulation was calculated using the MFI of M20ANef-infected cells as the maximum and the MFI of isotype stained cells as the minimum. Infections and flow measurements were repeated at least three times, ex-cept for the passaged viruses, for which only one sample was available. A two-tailedttest was used to determine differences between NL4-3 and mutant viruses.
Nucleotide sequence accession number.The sequences determined in this study have been deposited with GenBank with the accession num-bersJQ927866toJQ928096.
RESULTS
Isolation ofin vivoHIV-1nefquasispecies.As previously de-scribed (40), full-lengthnefsequences were isolated from plasma of nine persons with chronic, untreated HIV-1 infection. All sub-jects had detectable viremia ranging from 400 to⬎750,000 RNA copies/ml and peripheral blood CD4⫹T lymphocyte counts rang-ing from 0 to 900/mm3(data not shown). The bulknef
quasispe-cies from each subject (input sequences) were cloned into a repli-cation-competent NL4-3-based reporter virus for subsequent selection experiments.
Genetic evolution of primarynefquasispecies under experi-mental selection by Gag-specific CTLs.The influence on Nef of immune pressure against HIV-1 was assessed by subjecting the recombinant viruses to experimental selection by HIV-1 Gag-spe-cific CTLs. The recombinant viruses containing primarynef qua-sispecies were cultured either alone as a control for random ge-netic drift (control) or in combination with CTLs recognizing the Gag epitope SLYNTVATL (selected) for two passages of 7 days each, followed by clonalnefsequence analysis of the resulting vi-ruses. These control and selected sequences were aligned with the inputnefsequences (n⫽231) to create a neighbor-joining phy-logenetic tree that was statistically evaluated with 1,000 bootstrap replicates (Fig. 1). Sequences from each subject clustered indepen-dently (⬎99% bootstrap support), with the exception of those from subjects 00035 and 00039, who previously were identified to have related viruses, suggesting a common infection source (40). FIG 1Phylogeny ofnefquasispecies in the absence and presence of immune selection by Gag-specific CTLs. Plasmanefsequences from nine subjects (input sequences,n⫽94, blue circles),nefsequences passaged in the absence of CTL selection (control sequences,n⫽71, green squares), andnefsequences passaged under Gag-specific CTL selection (selected sequences,n⫽67, red triangles) were aligned with NL4-3nefto create a neighbor-joining phylogenetic tree. Independent clusters for each subject had⬎99% bootstrap support, with the exception of subjects 00035 and 00039, whose sequences were previously found to be related (40). Significant clustering of CTL-selected sequences (bootstrap values of⬎70%) is indicated by asterisks.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.123.464.66.317.2]A few highly divergent sequences were observed in the control quasispecies of subjects 00034, 00039, and 00041, although only the sequence from 00034 had evidence of G-to-A hypermutation (P⫽0.02). Generally, however, the persistingnefsequences after immune selection were intermingled with the input and control sequences. The CTL-selected sequences formed phylogenetically distinct clusters (bootstrap values of⬎70%) in two of nine sub-jects (00022 and 00034). In both of these cases, these sequences converged toward the clade B consensus sequence, suggesting evolution toward a more fit sequence.
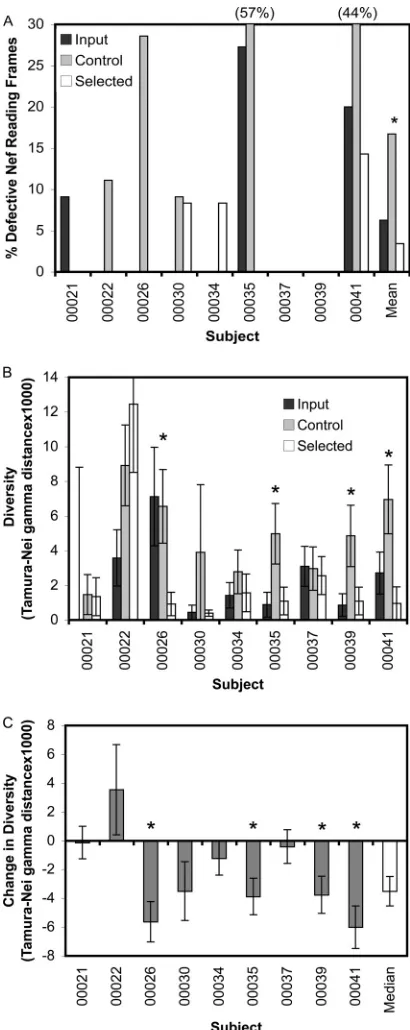
Increased maintenance of thenefreading frame as a result of CTL selection.The nucleotide alignments were translated into amino acid sequences to examine the status of the reading frame
(Fig. 2A). At the baseline, 6.5% (7/107) of the input sequences
from plasma contained nonsense mutations, including both pre-mature stop codons and frameshift mutations. The control pas-saged population cultured without CTL exhibited an increase to a nonsense mutations frequency of 14.8% (12/81), consistent with genetic drift in a setting where changes in Nef have little or no fitness cost, i.e.,in vitroculture in immortalized T cells (29). In contrast, the CTL-selected quasispecies had a significantly lower-than-expected nonsense mutation frequency of 4.9% (4/81) (2, P⫽0.0351). Overall, the increase in reading frame preservation with CTL selection versus the decrease in the absence of CTLs suggests that CTLs exert selective pressure on Nef to increase viral persistence.
Reduced diversity of primarynefquasispecies after CTL se-lection.The change in variability within thenefquasispecies pop-ulation in response to immune selective pressure was assessed for each subject individually and across all subjects by calculating changes in diversity and divergence from the clade B consensus sequence in the absence and presence of selection by the Gag-specific CTLs. As mentioned above, the quasispecies from subjects 00022 and 00034 with immune selection clearly converged on the clade B consensus sequence (Fig. 1). For all other subjects, whether analyzed individually or grouped, there was no signifi-cant change in sequence divergence with immune selection (data not shown). However, for four of nine subjects, there was a signif-icant (ttest,P⬍0.05) decrease in the diversity of the quasispecies after CTL selection (Fig. 2BandC). The decrease in diversity and the convergence toward the consensus sequence in the presence of CTL suggest that CTL selection places constraints on the evolu-tion of thenefreading frame.
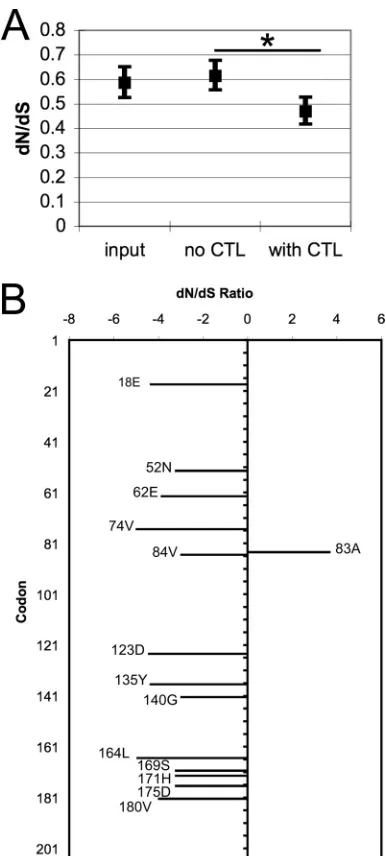
Global adaptive evolution ofneffor viral persistence in the setting of CTL immune selective pressure.The subset ofnef se-quences with intact reading frames was codon aligned and used to calculate the dN/dS ratio for the entire coding region for each of three sequence groups: input plasma sequences (n⫽94), CTL-selected sequences (n⫽71), and control passaged sequences (n⫽ 67) (Fig. 3A). The dN/dS ratio of the input plasmanefsequences demonstrated purifying selection at the baselinein vivo(dN/dS ratio⫽0.59; 95% CI, 0.53 to 0.65), similar to previously reported data (39). Control sequences passaged without CTL selection had a ratio similar to that of the input sequences (dN/dS ratio⫽0.61; 95% CI, 0.56 to 0.68). However, the CTL-selectednefsequences had significantly greater purifying selection (dN/dS ratio⫽0.47; 95% CI, 0.42 to 0.53) than to control sequences, as demonstrated by the nonoveralpping 95% CIs of the control and selected dN/dS ratio estimates. These results demonstrate that CTLs exerted
se-FIG 2CTL selection exerts evolutionary pressure on thenefquasispecies. Thenefquasispecies sequences were examined for changes resulting from selection by the Gag-specific CTLs, comparing the input, control, and se-lected sequences. (A) The percentages of sequences with nonsense muta-tions (frameshift and/or early stop mutamuta-tions) are plotted for each group across all subjects. (B) Fornefsequences from each subject, pairwise diver-sity (calculated for each group using 500 bootstrap replicates to give the standard error of the mean) is plotted for each group. (C) For each subject, the change innefdiversity due to CTL selection (comparing the control and selected groups) is plotted, and the median across all subjects is indicated. An asterisk indicates aPvalue⬍0.05 for the difference between control and selected sequences.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.318.523.65.580.2]lective pressure for maintenance of Nef through a functional con-straint.
Amino acids in Nef undergoing selection lie in important functional domains.To identify key sites within Nef that were un-dergoing selection, dN/dS ratios were calculated for each codon by the SLAC, REL, and FEL methods (50). Codons were considered to be under significant selection if they reached aPvalue of⬍0.05 by at least two of these three methods for the CTL selection and not the control sequences. Site-by-site analysis identified 13 sites subject to purifying selection and 1 site undergoing positive selection (Fig. 3B
andTable 1). Of these 14 sites, 7 were previously reported to be
asso-ciated with motifs important for Nef function, of which 5 were linked specifically to MHC-I downregulation (Table 1).
The identified sites are located in key domains of Nef (Fig. 4A), such as the N-terminal␣-helix (E18) and unstructured loops that bind cellular proteins (E62, L164, and D175) (27,38). Notably, site E62 lies within the EEEE acidic domain and site V74 lies at the position within the PxP motif, and both motifs are known to be required for MHC-I downregulation, although V74 has not been tested specifically for its effect on downregulation indepen-dently of the prolines (43,49,56,67). Site D123 is required for dimerization of Nef and therefore all its functions (7, 41,67), including MHC-I downregulation. Site E18 is the X within the RXR motif important for-COP binding and necessary for max-imal MHC-I downregulation, although previously only the argi-nines within this motif have been tested specifically (59,67). Site L164 lies within the dileucine motif required for CD4 downregu-lation by Nef and is also important for infectivity and replication in peripheral blood mononuclear cells (15,26,54). Sites V74, A83, and D175 lie within motifs implicated in the modulation of cell signaling pathways by Nef (20). While site S169 has no previously identified role in Nef function, a recent analysis showed that this site is coevolving with N157 and is therefore likely to have some functional role (51). The remaining six other sites under purifying selection (N52, A84, Y135, G140, H171, and V180) have no pre-viously defined associations with known functions of Nef.
CTL-selected sites in Nef are highly conserved in primary isolates of all HIV-1 genotypes.To determine whether these se-lected sites in the cohort tested here are broadly important to Nef in general, all of the complete Nef sequences in the LANL HIV-1 sequence database were examined for amino acid sequence con-servation at these sites. A total of 2,114 complete, nonrecombi-nant Nef sequences representing genotypes A to K submitted through 2010 were aligned and translated into amino acid se-quences. The probability of each amino acid at each of the 13 sites under purifying selection was plotted (Fig. 4B). At 11 of the 13 sites, there was⬎90% conservation of the amino acid, with only Y135 and S169 showing significant variability. There was virtually 100% conservation of 7 of 13 sites (V74, A84, D123, G140, L164, H171, and D175) of which A84, G140, and H171 have no previous association with Nef function. In comparison, the LANL Nef
se-TABLE 1Nef residues under selection by Gag-specific CTLs
Nef residuea
dN/dS
ratio Known functional role; reference(s)
E18 ⫺4.37 X in RXR motif,2MHC-I,-COP binding;59,67
N52 ⫺3.24 None reported
E62 ⫺3.87 2MHC-I, PACS-1 binding;49,67
V74 ⫺5.00 in PxP motif,2MHC-I, cell signaling;43,56
A83 3.68 2MHC-I, cell signaling;43
A84 ⫺3.00 None reported
D123 ⫺4.45 2MHC-I, dimerization, thioesterase binding;41,67
Y135 ⫺4.38 None reported G140 ⫺3.00 None reported
L164 ⫺4.97 Cellular trafficking,2CD4;15
S169 ⫺3.24 None reported H171 ⫺3.24 None reported
D175 ⫺3.24 Cellular trafficking and signaling;28,30,42
V180 ⫺4.00 None reported
aHXB2 numbering. FIG 3Passaging of HIV-1 in the presence of Gag-specific CTLs results in
purifying selection ofnef. The input, control, and selected sequences were evaluated for evidence of selective pressure as reflected by the dN/dS ratios. (A) Maximum-likelihood estimates of the global dN/dS ratios with 95% CIs are plotted for the three groups of sequences. An asterisk indicates nonoverlap-ping CIs. (B) Site-by-site analysis for CTL selection was performed by multiple methods; shown here are results from the SLAC method. The plot shows the estimated dN/dS ratios for codons (numbered according to the HXB2 num-bering system) that demonstrated significant selection (P⬍0.05) by both the SLAC and FEL methods.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.67.260.64.492.2] [image:5.585.299.545.547.716.2]quences were also examined for conservation at other sites previ-ously known to be associated with MHC-I downregulation: R17, R19, M20, E62 to E65, P72, P75, and P78 (Fig. 4C). There was less conservation of these sites than of the 13 selected sites, with only the three prolines demonstrating nearly 100% conservation and R17, R19, and E62 showing⬎90% conservation (60 versus 85% showing⬎90% conservation and 30 versus 54% with nearly 100% conservation). There was significant variability at E63 to E65 and significant numbers of M20I and M20L isolates of unknown func-tional significance. These results highlight the amino acid residues of primary Nef isolates that are associated with a survival advan-tage, confirm previously identified motifs, and suggest novel res-idues that are important for Nef structure/function in the context of CTL pressure.
Functional testing of Nef polymorphisms at CTL-selected sites.In order to determine whether any of the newly identified sites under purifying selection affected Nef’s ability to down-regulate MHC-I, a panel of mutants was created. Site-directed mutagenesis of NL4-3 Nef was used to incorporate the follow-ing polymorphisms, all observed in one or more of the primary plasma sequences and removed by CTL purifying selection (ex-cept H171A): N52A, N52S, A84D, Y135F, G140R, S169I, H171A, and V180E. Cells infected with recombinant reporter viruses with these Nef polymorphisms were assessed for levels of MHC-I downregulation compared to control viruses (Fig. 5). Six of eight mutant proteins had significant reductions in MHC-I downregulation compared to wild-type NL4-3 Nef
(Fig. 5A). Nef with G140R had complete loss of function, and
Nef with A84D produced a phenotype comparable to that of Nef with M20A, a mutant known to be deficient in MHC-I downregulation (3) (Fig. 5A andB). Nef with Y135F had an intermediate phenotype, about 50% of the function of NL4-3 Nef, while Nef with N52A, S169I, or V180E had significant although more modest reductions to approximately 80% of the level of NL4-3 Nef. Polymorphisms N52S and H171A had no effect on Nef function. These data show that the Nef
polymor-FIG 4Structural locations and conservation of amino acids associated with MHC-I downregulation. (A) The 13 codons determined to be under purifying selection are indicated in the predicted three-dimensional structure of the Nef protein (composite crystal structure kindly provided by Art F. Y. Poon). The probability of each amino acid, based on an alignment of all complete, nonrecombinant Nef sequences, including genotypes A to K, submitted to the LANL HIV-1 sequence database through 2010 (n⫽ ⬎2,100 sequences), was calculated for the sites under purifying selection shown inTable 1(B) and sites previously identified as important for Nef MHC-I downregulation (C). term, terminus.
FIG 5Downregulation of MHC-I by mutant forms of Nef identified by CTL immune selection. Eight mutant forms with changes at the seven selected sites with no previously reported role in Nef function were individually introduced into NL4-3 recombinant reporter viruses. Specific amino acid changes were selected on the basis of their presence in primary plasma isolates before selec-tion, except H171A. Their abilities to downregulate HLA-A*0201 were mea-sured by flow cytometry and compared to those of NL4-3 Nef, Delta Nef, and M20A Nef, a mutant form specifically deficient in MHC-I downregulation. (A) Summary of the average HLA-A*0201 downregulation of each mutant form relative to NL4-3 Nef based on at least three separate infections. A single as-terisk indicates a significant difference from NL4-3 with aPvalue of⬍0.05; double asterisks,P⬍0.001. (B) Histogram plots of the levels of HLA-A*0201 on cells infected with a virus that produces either mutant Nef (filled grams, mutation shown in the upper right corner) or NL4-3 Nef (open histo-grams).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.111.470.66.256.2] [image:6.585.300.543.344.584.2]phisms removed from the quasispecies by CTL purifying selec-tion are associated with deficiencies in MHC-I downregula-tion.
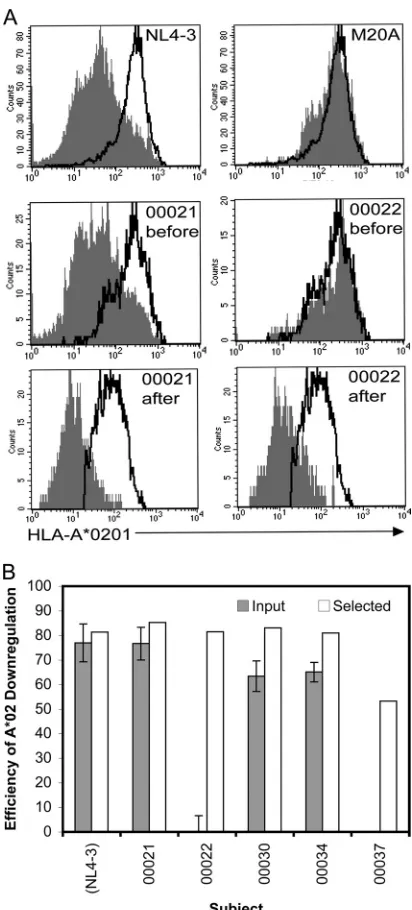
Gag-specific CTLs select for MHC-I downregulatory func-tion within primary Nef quasispecies. Because Nef-mediated downregulation of MHC-I is known to reduce the susceptibility of HIV-1-infected cells to CTLs, the primarynefquasispecies were tested for this function both before and after selection with the Gag-specific CTLs (Fig. 6). Cells infected with recombinant re-porter viruses carrying the nef quasispecies were assessed for MHC-I downregulation in comparison with viruses containing NL4-3 Nef (wild type) and M20A Nef (Fig. 6A). Infection with virus carrying NL4-3 Nef downregulated A*02 by about 80%, and this level of function was unchanged after the passage of this virus in the presence of CTLs. Similarly, virus withnefquasispecies from subject 00021 was functional at the baseline and after selec-tion. However, subjects 00030 and 00034 had Nef quasispecies with partial function at the baseline, which increased to full func-tion after selecfunc-tion. Most strikingly, Nef from subjects 00022 and 00037 (both with late-stage untreated AIDS and minimal CTL responsesin vivo) had no ability to downregulate MHC-I at the baseline, but CTL pressure selected functional populations of Nef
(Fig. 5B). Except for subject 00021, the baseline plasma
quasispe-cies of all of the subjects had amino acid polymorphisms at the sites identified in this analysis that would predict impaired func-tion, and viruses with these polymorphisms were not present after selection (Table 2). Quasispecies sequences were also examined for mutations at other sites previously known to be important for Nef MHC-I downregulation since these would also likely impair baseline function (Table 2). Although we were not able to test selected viruses from all subjects, we previously reported partial impairment of Nef-mediated MHC-I downregulation by the baseline plasma Nef quasispecies of all of the subjects included in this study, with the exception of subject 21 (40). Thus, the pres-ence of these mutations was associated with impaired function of the quasispecies, while reconstitution of function was associated with loss of these polymorphisms from the quasispecies. These data indicate that the sites identified by CTL selection play an important role in Nef-mediated MHC-I downregulation and con-sequent immune evasion and provide a functional context for the sequence evolution ofnefunder CTL selectionin vivo.
DISCUSSION
Human and animal model data suggest that Nef-mediated MHC-I downregulation plays a key role in pathogenesis by promoting viral persistence in the presence of a vigorous CTL response. We previously reported anin vivocorrelation between the breadth of the HIV-1-specific CTL response and the capacity of circulating Nef quasispecies to downregulate MHC-I (40). Furthermore, it has been demonstrated with a laboratory strain of HIV-1 that CTLs exert selective pressure to maintain functional Nef (3,5). The preservation of Nef-mediated MHC-I downregulation in the presence of CTL and its loss in the absence of strong CTL selection is also consistent with the observation of predominately defective Nef in neonates (25,65) and persons with late-stage AIDS and strong pressure to maintain Nef-mediated MHC-I downregula-tion in SIV-infected macaques (11,33,62). Here we demonstrate a selective advantage for primaryin vivoNef quasispecies that can downregulate MHC-I that correlates with the presence of both known and novel amino acid residues important for this function.
Examination of nef quasispecies sequence evolution across subjects due to immune selection by Gag-specific CTLs pin-pointed 13 sites where key amino acid residues are involved in the optimization of Nef-mediated immune evasion. Examination of FIG 6CTL-selected Nef sequences have preserved MHC-I downregulatory function. Levels of A*0201 on the surface of cells infected with reporter viruses carrying input versus CTL-selected Nef quasispecies were measured by flow cytometry. (A) Histogram plots of A*0201 on cells infected with wild-type NL4-3 Nef and M20A Nef, deficient in MHC-I downregulation (top panels) and for subjects 00021 and 00022 before and after immune selection (middle and bottom panels, respectively). Open histograms are cells without Nef; and filled histograms are cells infected with the Nef allele shown in the upper right corner. (B) Summary plots of Nef quasispecies from subjects 00021, 00022, 00030, 00034, and 00037 for input and selected viruses, as well as a wild-type NL4-3 virus control that underwent CTL selection. The error bars indicate the standard deviation for three independent experiments with each input virus group. Note that only one sample of each selected virus was available for testing.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.318.524.64.519.2]more than 2,000 Nef sequences in the LANL HIV-1 database re-vealed⬎90% conservation of the amino acid sequence at 11 of these 13 selected sites, with nearly 100% conservation at 7 sites. This analysis also confirmed several sites that were identified pre-viously through point mutagenesis studies of laboratory-adapted HIV-1nefsequences to be involved in multiple Nef functions. These included residues in the motifs important for dimerization (41), MHC-I downregulation (43,49,67), trafficking via adaptor proteins and-COP binding (59,67), and enhancement of viral replication through cell signaling (20).
Additionally, seven amino acid sites were identified as experi-encing strong purifying selection by CTL pressure and previously had no known role in Nef function. Mutations at six of these sites, reflecting polymorphismsin vivo, resulted in significant impair-ment of Nef-mediated MHC-I downregulation. Two sites in par-ticular, A84 and G140, were both virtually 100% conserved across all genotypes and resulted in complete or nearly complete loss of MHC-I downregulation when mutated. Although H171 was sim-ilarly 100% conserved, mutation at this site to an alanine had no effect on this Nef function. However, H171A was not among the observed polymorphisms at this sitein vivo(i.e., H171N, -P, and -G), and perhaps testing these may yield a different result. The MHC-I downregulation function of Nef with N52A, S169I, and V180E was only modestly affected, suggesting either that these mutations may work cooperatively with other mutations to have a more crippling effect or that they represent tradeoffs to optimize other Nef functions. A recent analysis has shown that site S169 coevolves with N157 (51), perhaps hinting that these sites may contribute to Nef function cooperatively. The exact mechanism whereby these mutations affect Nef function, how they interact with other sites, and whether they affect other Nef functions such as CD4 downregulation are not known but are currently being investigated.
It is also important to note that the 3=portion ofnefoverlaps the U3 region of the 3=long terminal repeat (LTR), and conse-quently, this region is potentially subject to additional LTR-re-lated constraints (36). However, the critical domains, including the NF-B and Sp1 binding sites and the TATAA box, are all downstream of the region ofnefoverlap. Five selected sites with no previously identified Nef function (Y135, G140, S169, H171, and V180) lie within this overlapping LTR region. Although it is pos-sible that these sites are under strong purifying selection due to an
LTR-associated function, the sites we identified were specific for CTL selection (i.e., not identified in both selected and control sequences), and Nef polymorphisms at four of these sites had significant impairment of MHC-I downregulation, making selec-tion due to an LTR funcselec-tion alone unlikely.
The important functional role played by these selected sites is clearly demonstrated by the reconstitution of MHC-I downregu-lation after CTL-mediated purifying selection by removing these mutant proteins from the quasispecies population. Except for subject 00021 Nef, which functioned at wild-type levels at the baseline, plasma quasispecies of all of our study subjects contained amino acid polymorphisms at the sites of purifying selection that would predict impaired function that subsequently were not pres-ent after selection. The most dramatic examples of functional re-constitution were the plasma Nef proteins of subjects 00022 and 00037, who had late-stage AIDS and minimal or undetectable HIV-1-specific CTL responses (40; data not shown), consistent with prior reports of loss of MHC-I downregulationin vivoin the absence of any CTL selective pressure in persons with AIDS (11,
33). Experimental selection by Gag-specific CTLs enriched fornef alleles with the capacity to downregulate MHC-I, suggesting a strong selective advantage for reconstituting this function of Nef in the presence of an active CTL response. The distinct phyloge-netic clustering of CTL-selectednefgenes from subject 00022 in-dicated overgrowth from a small subset of clones from within the baseline quasispecies population, and the convergence of these sequences (as well as those of subject 00034) toward the clade B consensus sequence indicated evolution toward the most gener-ally optimal sequence.
While MHC-I downregulation is likely to be the main mecha-nism by which Nef promotes HIV-1 survival under selection by CTLs, it is important to note that Nef is a polyfunctional protein with numerous effects on infected cells, including CD4 down-regulation and cellular activation. Our data do not exclude other functions that may be important for viral persistence in the face of CTL pressure. Some functions are likely to be separable due to distinct locations of important functional residues, while others may be interrelated (43). Several of the functionally important areas identified here and in other studies, such as the acidic do-main, PxP motif, and dileucine motif, lie in unstructured flexible loops of the protein (23,38). This may allow Nef to have func-tional flexibility to evolve and optimize different functions or combinations of functions in response to different environmental constraints. It is unexpected to see that L164 of the dileucine motif critical for CD4 downregulation by Nef was identified as a residue under strong purifying selection for viral persistence under CTL pressure. It may be that the function of this motif to bind adaptor proteins is important for both MHC-I and CD4 downregulation but more essential for the latter. It is also possible that other func-tions associated with this amino acid residue, such as enhance-ment of infectivity and replication, played a role in its selection under CTL pressure.
[image:8.585.40.287.87.215.2]Because Nef is an attractive target for pharmacologic or immu-nologic inhibitionin vivo, examining primary isolate Nef proteins for crucial functional sites that could serve as therapeutic targets is important. Mathematical modeling has predicted that blocking MHC-I downregulation by Nef has the potential to decrease viremia in chronically infected individuals by up to 2.4 logs by reducing Nef-mediated evasion of CTLs (66), and thus, inhibition of this function of Nef could be an effective therapeutic approach. TABLE 2Primary plasma Nef mutant genotypesaat selected and known
MHC-I-associated sites
Subject
no. Genotype at selected sites
Genotype at known MHC-I-associated site(s)b
00021 All consensus All consensus
00022 E18D, E62K, G140E R19K, E63V/A, P72A
00026 A84D, G140R, H171G All consensus
00030 E62K, V74A, H171P, D175N R17G, E63D, E65G, P78L
00034 E62G, D123G/N R19K/G, M20I
00035 E18K, E62K, Y135F, L164Y, H171N R19G 00037 N52S, D123N, G140R, D175E E63-65K
00039 E62G, Y135F E63G
00041 S169N/I, V180E All consensus
a
A change in at least one clone in the quasispecies mixture, not necessarily fixed substitutions or consensus sequences.
b
Sites examined: R17, R19, M20, E62-65, P72, P75, and P78.
on November 7, 2019 by guest
http://jvi.asm.org/
Small-molecule inhibitors of Nef have been considered for this purpose (53,55). Alternatively, an appropriately directed vaccine response could achieve this goal by putting immune pressure di-rectly on Nef (3). Of note, two of the six vaccine-induced epitopes that predicted efficacy of vaccination in reducing set point viremia in the HVTN 502 (STEP) trial were the Nef epitopes B*57-re-stricted HW9 (HTQGYFPDW, Nef 116 to 124) and A*02-re-stricted LV10 (LTFGWCFKLV, Nef 137 to 146) (10). These epitopes contain the D123 and G140 sites we identified as being under selective pressure. While D123 is known to be important for Nef dimerization and both CD4 and MHC-I downregulation (41), G140 was not previously known to have an important func-tional role, but now we demonstrate that mutation at this site critically impairs MHC-I downregulation. Thus, examining pri-mary isolate sequences may be important for identifying sites in Nef that are most relevant for its role in immune evasion and for which pharmacologic or immunologic targeting may be most ef-fective due to strict functional constraints.
Prior reports have demonstrated that direct CTL targeting of Nef yields positive selective pressure that leads to loss of function (5,35,64,69), complementing our finding of purifying selection and reconstitution of Nef function in the setting of CTLs targeting Gag and not Nef directly. While the evolution of Nef and other HIV-1 proteinsin vivoappears to be dominated overall by puri-fying selection, reflecting strong functional constraints (39), there is clear positive selective pressure exerted by direct CTL targeting of Nef. This has been demonstrated experimentally;in vitro selec-tion of laboratory-adapted HIV-1 strains with Nef-specific CTL clones resulted in a dramatic pattern of point mutations, dele-tions, and nonsense mutations due to the lack of a fitness cost for Nef deletionin vitro(6,69). Subsequently, these selected labora-tory virus strains deficient in functional Nef were demonstrated to become more susceptible to non-Nef-specific CTLs (6,64). It was further shown that simultaneous addition of Gag-specific CTLs placed a functional constraint on viral escape from Nef-specific CTLs by Nef mutation (5). Our data confirm and extend these findings with more relevant primary isolate Nef alleles and suggest that these principles may apply to therapeutic interventionsin vivo.
In summary, these results highlight the close reciprocal rela-tionship between the host CTL immune response and Nef func-tion.nefquasispecies under CTL selection display a pattern of strong purifying selection associated with optimization of MHC-I downregulation. Study of circulating primary isolate Nef alleles revealed novel amino acid residues that are directly important for HIV-1 persistence under immune pressure by the host CTL re-sponse. Better definition of functional sites within circulating plasma Nef quasispecies will be useful for the design of pharma-cologic or immunotherapeutic agents targeting functionally cru-cial regions of Nef capable of disabling its ability to direct immune evasion.
ACKNOWLEDGMENTS
This work was supported by NIH AI068449 (M.J.L.), AI083083 (M.J.L.), and AI051970 (O.O.Y.).
IL-2 was provided by the NIH AIDS Reagent Repository. We thank Mabel Ching Yee Chan for her technical assistance.
REFERENCES
1.Adnan S, et al.2006. Nef interference with HIV-1-specific CTL antiviral activity is epitope specific. Blood108:3414 –3419.
2.Ahmad N, Venkatesan S.1988. Nef protein of HIV-1 is a transcriptional repressor of HIV-1 LTR. Science241:1481–1485.
3.Akari H, et al.2000. Nef-induced major histocompatibility complex class I down-regulation is functionally dissociated from its virion incorpora-tion, enhancement of viral infectivity, and CD4 down-regulation. J. Virol.
74:2907–2912.
4.Ali A, Jamieson BD, Yang OO.2003. Half-genome human immunode-ficiency virus type 1 constructs for rapid production of reporter viruses. J. Virol. Methods110:137–142.
5.Ali A, Ng HL, Dagarag MD, Yang OO.2005. Evasion of cytotoxic T lymphocytes is a functional constraint maintaining HIV-1 Nef expression. Eur. J. Immunol.35:3221–3228.
6.Ali A, et al.2003. Broadly increased sensitivity to cytotoxic T lymphocytes resulting from Nef epitope escape mutations. J. Immunol. 171:3999 – 4005.
7.Arold S, et al.2000. Characterization and molecular basis of the oligo-meric structure of HIV-1 nef protein. Protein Sci.9:1137–1148. 8.Baur AS, et al.1994. HIV-1 Nef leads to inhibition or activation of T cells
depending on its intracellular localization. Immunity1:373–384. 9.Bodéus M, Marie-Cardine A, Bougeret C, Ramos-Morales F, Benarous
R.1995. In vitro binding and phosphorylation of human immunodefi-ciency virus type 1 Nef protein by serine/threonine protein kinase. J. Gen. Virol.76:1337–1344.
10. Brumme ZL, et al.2009. HLA-associated immune escape pathways in HIV-1 subtype B Gag, Pol and Nef proteins. PLoS One4:e6687. doi: 10.1371/journal.pone.0006687.
11. Carl S, et al.2001. Modulation of different human immunodeficiency virus type 1 Nef functions during progression to AIDS. J. Virol.75:3657– 3665.
12. Cohen GB, et al.1999. The selective downregulation of class I major histocompatibility complex proteins by HIV-1 protects HIV-infected cells from NK cells. Immunity10:661– 671.
13. Collins KL, Chen BK, Kalams SA, Walker BD, Baltimore D. 1998. HIV-1 Nef protein protects infected primary cells against killing by cyto-toxic T lymphocytes. Nature391:397– 401.
14. Couillin I, et al.1994. Impaired cytotoxic T lymphocyte recognition due to genetic variations in the main immunogenic region of the human im-munodeficiency virus 1 NEF protein. J. Exp. Med.180:1129 –1134. 15. Craig HM, Pandori MW, Guatelli JC.1998. Interaction of HIV-1 Nef
with the cellular dileucine-based sorting pathway is required for CD4 down-regulation and optimal viral infectivity. Proc. Natl. Acad. Sci. U. S. A.95:11229 –11234.
16. Crooks GE, Hon G, Chandonia JM, Brenner SE.2004. WebLogo: a sequence logo generator. Genome Res.14:1188 –1190.
17. Daniel MD, Kirchhoff F, Czajak SC, Sehgal PK, Desrosiers RC.1992. Protective effects of a live attenuated SIV vaccine with a deletion in the nef gene. Science258:1938 –1941.
18. Deacon NJ, et al.1995. Genomic structure of an attenuated quasi species of HIV-1 from a blood transfusion donor and recipients. Science270: 988 –991.
19. Dyer WB, et al. 1997. Lymphoproliferative immune function in the Sydney Blood Bank Cohort, infected with natural nef/long terminal repeat mutants, and in other long-term survivors of transfusion-acquired HIV-1 infection. AIDS11:1565–1574.
20. Erdtmann L, et al.2000. Two independent regions of HIV-1 Nef are required for connection with the endocytic pathway through binding to the mu 1 chain of AP1 complex. Traffic1:871– 883.
21. Fackler OT, Baur AS.2002. Live and let die: Nef functions beyond HIV replication. Immunity16:493– 497.
22. Felsenstein J.1989. PHYLIP—phylogeny inference package (version 3.2). Cladistics5:164 –166.
23. Franken P, et al.1997. HIV-1 Nef protein: purification, crystallizations, and preliminary X-ray diffraction studies. Protein Sci.6:2681–2683. 24. Garcia JV, Miller AD.1991. Serine phosphorylation-independent
down-regulation of cell-surface CD4 by nef. Nature350:508 –511.
25. Geffin R, et al.2000. Functional and structural defects in HIV type 1 nef genes derived from pediatric long-term survivors. AIDS Res. Hum. Ret-roviruses16:1855–1868.
26. Geyer M, Fackler OT, Peterlin BM.2001. Structure-function relation-ships in HIV-1 Nef. EMBO Rep.2:580 –585.
27. Geyer M, Munte CE, Schorr J, Kellner R, Kalbitzer HR.1999. Structure of the anchor-domain of myristoylated and non-myristoylated HIV-1 Nef protein. J. Mol. Biol.289:123–138.
on November 7, 2019 by guest
http://jvi.asm.org/
28. Geyer M, et al.2002. Subunit H of the V-ATPase binds to the medium chain of adaptor protein complex 2 and connects Nef to the endocytic machinery. J. Biol. Chem.277:28521–28529.
29. Gibbs JS, Regier DA, Desrosiers RC.1994. Construction and in vitro properties of HIV-1 mutants with deletions in “nonessential” genes. AIDS Res. Hum. Retroviruses10:343–350.
30. Hodge DR, et al.1998. Binding of c-Raf1 kinase to a conserved acidic sequence within the carboxyl-terminal region of the HIV-1 Nef protein. J. Biol. Chem.273:15727–15733.
31. Huang Y, Zhang L, Ho DD.1995. Characterization of nef sequences in long-term survivors of human immunodeficiency virus type 1 infection. J. Virol.69:93–100.
32. Kestler HW, III, et al.1991. Importance of the nef gene for maintenance of high virus loads and for development of AIDS. Cell65:651– 662. 33. Kirchhoff F, et al.1999. Sequence variations in human
immunodefi-ciency virus type 1 Nef are associated with different stages of disease. J. Virol.73:5497–5508.
34. Kirchhoff F, Greenough TC, Brettler DB, Sullivan JL, Desrosiers RC.
1995. Brief report: absence of intact nef sequences in a long-term survivor with nonprogressive HIV-1 infection. N. Engl. J. Med.332:228 –232. 35. Koenig S, et al.1995. Transfer of HIV-1-specific cytotoxic T lymphocytes
to an AIDS patient leads to selection for mutant HIV variants and subse-quent disease progression. Nat. Med.1:330 –336.
36. Krebs FC, Hogan TH, Quiterio S, Gartner S, Wigdahl B.2001. Lentiviral LTR-directed expression, sequence variation, and disease pathogenesis. Theoretical Biology and Biophysics Group, Los Alamos National Labora-tory, Los Alamos, NM. http://www.hiv.lanl.gov/content/sequence /HIV/COMPENDIUM/2001/partI/Wigdahl.pdf.
37. Lama J, Mangasarian A, Trono D.1999. Cell-surface expression of CD4 reduces HIV-1 infectivity by blocking Env incorporation in a Nef- and Vpu-inhibitable manner. Curr. Biol.9:622– 631.
38. Lee CH, Saksela K, Mirza UA, Chait BT, Kuriyan J. 1996. Crystal structure of the conserved core of HIV-1 Nef complexed with a Src family SH3 domain. Cell85:931–942.
39. Lemey P, Rambaut A, Pybus OG.2006. HIV evolutionary dynamics within and among hosts. AIDS Rev.8:125–140.
40. Lewis MJ, et al.2008. Functional adaptation of Nef to the immune milieu of HIV-1 infection in vivo. J. Immunol.180:4075– 4081.
41. Liu LX, et al.2000. Mutation of a conserved residue (D123) required for oligomerization of human immunodeficiency virus type 1 Nef protein abolishes interaction with human thioesterase and results in impairment of Nef biological functions. J. Virol.74:5310 –5319.
42. Lu X, Yu H, Liu SH, Brodsky FM, Peterlin BM. 1998. Interactions between HIV1 Nef and vacuolar ATPase facilitate the internalization of CD4. Immunity8:647– 656.
43. Mangasarian A, Piguet V, Wang JK, Chen YL, Trono D.1999. Nef-induced CD4 and major histocompatibility complex class I (MHC-I) down-regulation are governed by distinct determinants: N-terminal alpha helix and proline repeat of Nef selectively regulate MHC-I trafficking. J. Virol.73:1964 –1973.
44. Mariani R, et al.1996. High frequency of defectivenefalleles in a long-term survivor with nonprogressive human immunodeficiency virus type 1 infection. J. Virol.70:7752–7764.
45. Niederman TM, Thielan BJ, Ratner L.1989. Human immunodeficiency virus type 1 negative factor is a transcriptional silencer. Proc. Natl. Acad. Sci. U. S. A.86:1128 –1132.
46. Noviello CM, et al.2007. Maintenance of Nef-mediated modulation of major histocompatibility complex class I and CD4 after sexual transmis-sion of human immunodeficiency virus type 1. J. Virol.81:4776 – 4786. 47. Piguet V, Trono D.1999. A structure-function analysis of the nef protein of
primate lentiviruses, p 448 – 459.InKuiken CL, et al (ed), Human retroviruses and AIDS. Los Alamos National Laboratory, Los Alamos, NM.
48. Piguet V, Trono D.1999. The Nef protein of primate lentiviruses. Rev. Med. Virol.9:111–120.
49. Piguet V, et al.2000. HIV-1 Nef protein binds to the cellular protein PACS-1 to downregulate class I major histocompatibility complexes. Nat. Cell Biol.2:163–167.
50. Pond SL, Frost SD, Muse SV.2005. HyPhy: hypothesis testing using phylogenies. Bioinformatics21:676 – 679.
51. Poon AF, et al.2010. Phylogenetic analysis of population-based and deep sequencing data to identify coevolving sites in the nef gene of HIV-1. Mol. Biol. Evol.27:819 – 832.
52. Posada D, Crandall KA.1998. MODELTEST: testing the model of DNA substitution. Bioinformatics14:817– 818.
53. Richter SN, Frasson I, Palu G.2009. Strategies for inhibiting function of HIV-1 accessory proteins: a necessary route to AIDS therapy? Curr. Med. Chem.16:267–286.
54. Riggs NL, Craig HM, Pandori MW, Guatelli JC.1999. The dileucine-based sorting motif in HIV-1 Nef is not required for down-regulation of class I MHC. Virology258:203–207.
55. Saksela K.2004. Therapeutic targeting of interactions between Nef and host cell proteins. Curr. Drug Targets Immune Endocr. Metabol. Disord.
4:315–319.
56. Saksela K, Cheng G, Baltimore D.1995. Proline-rich (PxxP) motifs in HIV-1 Nef bind to SH3 domains of a subset of Src kinases and are required for the enhanced growth of Nef⫹viruses but not for down-regulation of CD4. EMBO J.14:484 – 491.
57. Salter RD, Howell DN, Cresswell P.1985. Genes regulating HLA class I antigen expression in T-B lymphoblast hybrids. Immunogenetics21:235– 246.
58. Sawai ET, et al.1994. Human immunodeficiency virus type 1 Nef asso-ciates with a cellular serine kinase in T lymphocytes. Proc. Natl. Acad. Sci. U. S. A.91:1539 –1543.
59. Schaefer MR, Wonderlich ER, Roeth JF, Leonard JA, Collins KL.2008. HIV-1 Nef targets MHC-I and CD4 for degradation via a final common beta-COP-dependent pathway in T cells. PLoS Pathog.4:e1000131. doi: 10.1371/journal.ppat.1000131.
60. Schwartz O, Marechal V, Le Gall S, Lemonnier F, Heard JM.1996. Endocytosis of major histocompatibility complex class I molecules is in-duced by the HIV-1 Nef protein. Nat. Med.2:338 –342.
61. Smith BL, Krushelnycky BW, Mochly-Rosen D, Berg P.1996. The HIV nef protein associates with protein kinase C theta. J. Biol. Chem.271: 16753–16757.
62. Swigut T, et al.2004. Impact of Nef-mediated downregulation of major histocompatibility complex class I on immune response to simian immu-nodeficiency virus. J. Virol.78:13335–13344.
63. Tomiyama H, Akari H, Adachi A, Takiguchi M.2002. Different effects of Nef-mediated HLA class I down-regulation on human immunodefi-ciency virus type 1-specific CD8(⫹) T-cell cytolytic activity and cytokine production. J. Virol.76:7535–7543.
64. Ueno T, et al.2008. CTL-mediated selective pressure influences dynamic evolution and pathogenic functions of HIV-1 Nef. J. Immunol.180:1107– 1116.
65. Walker PR, et al.2007. Polymorphisms in Nef associated with different clinical outcomes in HIV type 1 subtype C-infected children. AIDS Res. Hum. Retroviruses23:204 –215.
66. Wick WD, Gilbert PB, Yang OO.2009. Predicting the impact of blocking human immunodeficiency virus type 1 Nef in vivo. J. Virol.83:2349 – 2356.
67. Williams M, Roeth JF, Kasper MR, Filzen TM, Collins KL. 2005. Human immunodeficiency virus type 1 Nef domains required for disrup-tion of major histocompatibility complex class I trafficking are also nec-essary for coprecipitation of Nef with HLA-A2. J. Virol.79:632– 636. 68. Yang OO, et al.2002. Nef-mediated resistance of human immunodeficiency
virus type 1 to antiviral cytotoxic T lymphocytes. J. Virol.76:1626 –1631. 69. Yang OO, et al.2003. Determinant of HIV-1 mutational escape from
cytotoxic T lymphocytes. J. Exp. Med.197:1365–1375.