J
OURNAL OFV
IROLOGY, Oct. 2010, p. 10812–10819
Vol. 84, No. 20
0022-538X/10/$12.00
doi:10.1128/JVI.00543-10
Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Genetic Analyses of HIV-1
env
Sequences Demonstrate Limited
Compartmentalization in Breast Milk and Suggest Viral
Replication within the Breast That Increases
with Mastitis
䌤
Soren Gantt,
1,5* Jacquelyn Carlsson,
5Laura Heath,
2Marta E. Bull,
5Avinash K. Shetty,
6Junior Mutsvangwa,
7Georgina Musingwini,
8Godfrey Woelk,
8,9Lynn S. Zijenah,
8,10David A. Katzenstein,
11James I. Mullins,
2,3,4and Lisa M. Frenkel
1,3,5Departments of Pediatrics,
1Microbiology,
2Laboratory Medicine,
3and Medicine,
4University of Washington, and
Seattle Children’s Hospital,
5Seattle, Washington; Department of Pediatrics, Wake Forest University Health Sciences,
Winston-Salem, North Carolina
6; Biomedical Research and Training Institute, Harare, Zimbabwe
7;
Zimbabwe AIDS Prevention Project
8and Departments of Community Medicine
9and
Immunology,
10University of Zimbabwe, Harare, Zimbabwe; and Department of
Internal Medicine, Stanford University, Palo Alto, California
11Received 12 March 2010/Accepted 8 July 2010
The concentration of human immunodeficiency virus type 1 (HIV-1) is generally lower in breast milk than
in blood. Mastitis, or inflammation of the breast, is associated with increased levels of milk HIV-1 and risk of
mother-to-child transmission through breastfeeding. We hypothesized that mastitis facilitates the passage of
HIV-1 from blood into milk or stimulates virus production within the breast. HIV-1
env
sequences were
generated from single amplicons obtained from breast milk and blood samples in a cross-sectional study. Viral
compartmentalization was evaluated using several statistical methods, including the Slatkin and Maddison
(SM) test. Mastitis was defined as an elevated milk sodium (Na
ⴙ) concentration. The association between milk
Na
ⴙand the pairwise genetic distance between milk and blood viral sequences was modeled using linear
regression. HIV-1 was compartmentalized within milk by SM testing in 6/17 (35%) specimens obtained from
9 women, but all phylogenetic clades included viral sequences from milk and blood samples. Monotypic
sequences were more prevalent in milk samples than in blood samples (22% versus 13%;
P
ⴝ
0.012), which
accounted for half of the compartmentalization observed. Mastitis was not associated with
compartmental-ization by SM testing (
P
ⴝ
0.621), but Na
ⴙwas correlated with greater genetic distance between milk and blood
HIV-1 populations (
P
ⴝ
0.041). In conclusion, local production of HIV-1 within the breast is suggested by
compartmentalization of virus and a higher prevalence of monotypic viruses in milk specimens. However,
phylogenetic trees demonstrate extensive mixing of viruses between milk and blood specimens. HIV-1
repli-cation in breast milk appears to increase with inflammation, contributing to higher milk viral loads during
mastitis.
Breastfeeding accounts for 30 to 50% of
mother-to-child-transmission (MTCT) of human immunodeficiency virus type 1
(HIV-1) (38). MTCT through breastfeeding occurs primarily
in sub-Saharan Africa, where the use of artificial infant
for-mula is often not feasible because of cost and the associated
infant mortality from infections due to the use of unsafe water
and the lack of the protective effects of breast milk (19, 38, 51).
Numerous strategies to reduce postnatal HIV-1 infection of
infants while preserving the advantages of breastfeeding have
been evaluated, including maternal use of combination
anti-retroviral therapy or infant antianti-retroviral prophylaxis during
the period of breastfeeding (5, 25, 26, 30, 40). Understanding
the biologic events that increase the concentration of HIV-1 in
breast milk is critical to the development and evaluation of
interventions to reduce postnatal MTCT.
The risk of MTCT is strongly associated with the concentration
of HIV-1 in breast milk (28, 46, 47). Although breast milk HIV-1
RNA concentrations correlate with those in plasma, levels in milk
are typically 2 log
10lower (15, 24, 43). This suggests that HIV-1 in
blood and milk may not mix freely, likely because of the closure
of tight junctions between mammary alveolar cells that occurs
once milk production is established and before weaning (16).
Thus, HIV-1 may evolve in the breast without substantial mixing
with blood, i.e., evolving viral variants would become
compart-mentalized—a phenomenon that has been observed in the central
nervous system (50) and in some studies of the genital tract (10,
44, 57). Compartmentalization of HIV-1 variants has been
de-tected in the breast milk of a small number of women (3, 4), but
other data suggest that compartmentalization in breast milk may
be uncommon (22).
Breast inflammation (mastitis) occurs frequently during
lac-tation, most commonly without symptoms. Mastitis is
associ-ated with elevations in HIV-1 RNA levels in milk (15, 31, 47,
55), an increase in the number of inflammatory cells in milk,
and opening of tight junctions in the mammary epithelium that
* Corresponding author. Mailing address: Seattle Children’s
Hospi-tal Research Institute, 1900 Ninth Ave., Seattle, WA. 98101-1304.
Phone: (206) 987-1160. Fax: (206) 884-7311. E-mail: [email protected].
䌤
Published ahead of print on 21 July 2010.
10812
on November 8, 2019 by guest
http://jvi.asm.org/
allows passage of subcellular blood components, of which
so-dium (Na
⫹) serves as a marker (15, 16, 36, 47, 55). Greater
permeability of mammary epithelia may allow the passage of
free virus from the blood into breast milk, which would result
in the mixing of HIV-1 subpopulations from blood and milk.
Alternatively, inflammation in the breast may induce
replica-tion of virus by HIV-1-infected cells within the breast, which
would result in divergence between milk and blood HIV-1
subpopulations. Here we describe detailed genetic analyses of
HIV-1 subpopulations in the blood and breast milk to
deter-mine whether mastitis affects the structure of these
popula-tions and to gain understanding of the processes that may lead
to increased concentrations of HIV-1 in milk.
MATERIALS AND METHODS
Study design.Specimens were selected from participants in a previously de-scribed (15) cross-sectional study of Zimbabwean women who were between 6 and 16 weeks postpartum, HIV-1 infected, and breastfeeding. All participants provided written informed consent, and all procedures were approved by the Institutional Review Board of Seattle Children’s Hospital and the Medical Re-search Council of Zimbabwe. Specimens were selected based on a high HIV-1 concentration in milk and adequate specimen volume for generation of multiple sequences by single-template PCR, and a subset were selected to have mastitis,
defined by a milk Na⫹concentration of⬎12 mM.
Specimen collection and processing.Briefly, breast milk was collected by manual expression, and skim milk was separated from cells using centrifugation
(15). The Na⫹concentration in skim milk was determined using a Roche/Hitachi
902 clinical chemistry analyzer (15). HIV-1 RNA was quantified using the Am-plicor Monitor kit (version 1.5; Roche Diagnostics) in plasma using the standard method (detection limit, 400 copies/ml) and in skim milk using the ultrasensitive method (detection limit, 50 copies/ml) (15).
Generation of single-template sequences.The nucleic acids in 0.2 to 1.0 ml of plasma samples and 0.7 to 2.0 ml of skim milk samples were extracted using Boom silica methods, with slight modifications (6, 8). Purified RNA was reverse transcribed into cDNA using primer BH2JC (TTAGTGGGTGCTACTCCTAA TGG; HXB2 coordinates 7701 to 7723) or ED12 (AGTGCTTCCTGCTGCTC
CCAAGAACCCAAG; HXB2 coordinates 7822 to 7792) forenv(1) and primer
RT1-C (GTTGACTCAGCTTGGATGCAC; HXB2 coordinates 2519 to 2539)
forpol. DNA from 1.2 ml of whole-blood or frozen milk cell pellets derived from
4 to 14 ml of whole breast milk (15) were extracted using the QIAamp minikit
(Qiagen, Hilden, Germany) into 50l of elution buffer, according to the
man-ufacturer’s instructions. The concentration of amplifiable viral templates in ex-tracted nucleic acids was quantified using limiting dilution nested PCR (45), with a multiplexed first round containing the above-mentioned BH2JC and RT1-C forward primers and ED5JC (TGCATGAGGATATAATCAGTTTATGGGA;
HXB2 coordinates 6535 to 6562) to amplifyenvand RT2-C (GTATATCATTG
ACAGTCCAGC; HXB2 coordinates 3321 to 3301) to amplifypol. In a
second-round PCR, the C2-V5 region ofenvwas amplified with primers DR7JC (GTG
GTATCAACTCAACTACTGTTAAATGG; HXB2 coordinates 6984 to 7012) and DR8 (CACTTCTCCAATTGTCCCTCATATCTCCTCC; HXB2 coordi-nates 7638 to 7668). Viral gene sequences were then derived by one of the two following methods. First, up to 100 amplification reactions were performed on
the nucleic acid from each specimen after dilution to⬃0.3 HIV-1 amplifiable
copies/reaction to amplify a single viral template (45, 53). Positive reactions were identified by gel electrophoresis, processed with ExoSAP-IT (USB Corporation, Cleveland, OH), and sequenced directly using fluorescent dye terminators (BigDye Terminator version 3.1 cycle sequencing kit; Applied Biosystems, Foster City, CA) and a Prism 3730XL DNA analyzer (Applied Biosciences). Second, when the average viral input exceeded 0.3 copies/reaction, amplicons were cloned as previously described (8), and one clone per PCR was sequenced to ensure derivation from separate viral templates (32). Each type of specimen (plasma RNA, whole-blood DNA, skim milk RNA, and milk cell pellet DNA) obtained from a participant was evaluated on a different day to minimize the risk of cross-contaminating specimens.
Sequence analysis and phylogenetic tree construction.Sequences were assem-bled and checked for read errors in Sequencher version 3.4 (Gene Codes Cor-poration, Ann Arbor, MI). Alignments were generated in CLUSTALW (52), and hypermutated sequences were excluded and manually adjusted using MacClade version 4.08 (34). An all-inclusive phylogenetic tree was used to verify that each
sequence segregated only with others from the identified participant. Sequences from all participants were carefully examined for contamination by comparisons to known reference sequences and to all other sequences generated in the laboratories, and no matches were found. We also screened each alignment for recombination, since this could confound compartmentalization (56). For all subjects, phylogenetic trees were constructed using the DIVEIN interface (11) (http://indra.mullins.microbiol.washington.edu/DIVEIN/index.html), with a
gen-eral time-reversible model plus invariant plus gamma distribution (GTR⫹I⫹G)
model evolution within PhyML (version 3.0) (18). Four representative subtype C HIV-1 sequences from GenBank (accession numbers AY265933, AY265936, AF268277, and AY265952) were used to root the phylogenies of each partici-pant’s sequences. In order to better evaluate the possibility of dual HIV-1 infection in the phylogenies of those individuals (participants A, D, E, G, and H) with pronounced separation of discrete clades, eight additional subtype C se-quences (GenBank accession numbers AF443091, AY772691, AY162224, AY772699, AF067155, AF286224, AF443112, and AF443106) as well as one each from subtypes A, B, and D (GenBank accession numbers AF286241, U63632, and AY253311) were included. HIV-1 coreceptor usage was predicted for the V3 region amino acid sequences using the subtype C position-specific scoring X4/R5 and syncytium-inducing and non-syncytium-inducing matrices (23) (http://indra .mullins.microbiol.washington.edu/pssm/).
Evaluation of HIV-1 population structure and statistical testing.The topology of each phylogenetic tree was examined, with particular attention to the identi-fication of breast milk- or peripheral blood-specific clades. The compartmental structure of viral sequences in blood and milk specimens obtained from each subject was evaluated by the Slatkin and Maddison (SM) test, which evaluates variation from normalcy in the distribution of sequences over a predicted tree structure using MacClade (8, 49). Each blood and milk specimen was included in SM analysis if at least five sequences were available from that specimen. Com-parisons were performed using each infecting HIV-1 strain separately for those participants that had evidence of dual infection. The SM test was further eval-uated using 1,000 bootstrap replicate phylogenies (7, 44). Genetic differentiation between breast milk and blood specimens was also evaluated using the Bayesian tip-significance (BaTS) testing tool (41) (http://evolve.zoo.ox.ac.uk/Evolve/BaTS .html), which takes into account uncertainty arising from phylogenetic structure error by testing over multiple credible topologies produced by the Bayesian phylogenetics program BEAST (13) (http://beast.bio.ed.ac.uk/Main_Page). BaTS testing evaluates the sequences by running the following two established tests: the association index (AI) (13), which assesses the population structure by weighting the contribution of each internal node based on how deep it is in the tree, and the parsimony score (PS), also known as the Slatkin and Maddison test, described above. BaTS also incorporates a new measure, the maximum single-state clade size (MC), a statistic which quantifies the observation that stronger phylogeny-trait associations should produce larger monophyletic clades whose tips all share the same trait (41). Each participant’s alignment was used to generate a posterior sample of trees (PST) in BEAST, the exponential growth model under the Hasegawa-Kishino-Yano plus gamma invariant model with a
Markov chain Monte Carlo chain length of 5⫻106and parameter logging every
1,000 generations, yielding a PST of 5,000, which was the input data for BaTS testing, minus a burn-in of the first 10%, or 500 trees. BaTS testing was set to
generate 100 null distributions to test the significance of the observed data.P
values of⬍0.05 were considered to be evidence of compartmentalization without
correction for multiple comparisons. The pairwise genetic distances of each breast milk sequence from each blood sequence within a participant were cal-culated using the DIVEIN distance generator under the same model that was estimated for phylogeny calculation (48).
Milk Na⫹medians were compared using the two-sample Wilcoxon rank sum
(Mann-Whitney) test, and the proportions of monotypic virus were compared using Fisher’s exact test (two tailed). Linear regression and the generalized estimating equation (GEE) with robust standard errors were used to evaluate the
association between the milk Na⫹concentration and the mean pairwise genetic
distance between sequences in the breast milk and blood specimens obtained from each participant. These analyses were performed using Stata/SE 9.2 for Macintosh (StataCorp, College Station, TX).
Nucleotide sequence accession numbers.The gene sequences determined in this study were deposited in GenBank under accession numbers HQ172164 through HQ172673.
RESULTS
Participant and specimen characteristics.
Blood and breast
milk specimens obtained from both breasts of nine women
on November 8, 2019 by guest
http://jvi.asm.org/
collected 6 to 16 weeks postpartum were evaluated in a
cross-sectional study (15). All participants had received a single dose
of nevirapine for the prevention of MTCT (17) but were not
otherwise exposed to antiretroviral drugs. The median HIV-1
RNA viral load in plasma samples was 146,224 copies/ml
(range, 25,831 to 986,896 copies/ml) and in milk samples was
4,105 copies/ml (range, 241 to 124,254 copies/ml) (Table 1). A
total of 515 individual HIV-1
env
sequences were generated,
with a median of 54 sequences from each participant (range, 42
to 74) (Table 1 and Fig. 1). Of these sequences, 186 were
generated from HIV-1 RNA, and 329 were generated from
HIV-1 DNA. A total of 171 of these sequences were derived
from blood samples, and 344 were derived from milk samples
(Table 1 and Fig. 1).
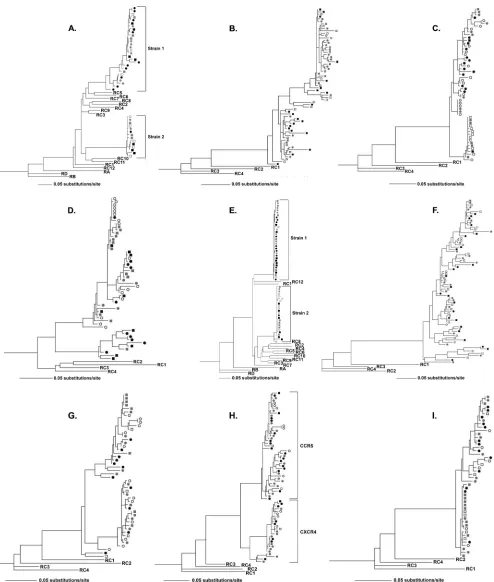
Analysis of HIV-1 population structure.
Pronounced
sepa-ration of discrete clades was apparent in phylogenetic trees
obtained from five of the nine participants (A, D, E, G, and H).
The trees obtained from two of these individuals, participants
A and E, displayed separation of viral clades by reference
sequences without evidence of recombination, indicating
infec-tion with two unrelated strains of subtype C HIV-1 (Fig. 1). A
lack of genetic diversity in cell-associated HIV-1, suggestive of
recent infection, was notable in each infecting virus population
of participant E. Phylogenies obtained from each of the other
participants displayed a monophyletic cluster of sequences
dis-tinct from reference sequences, without evidence of dual
in-fection. The sequences obtained from participant H were
monophyletic with respect to reference sequences yet
dis-played two discrete clades that were found to differ by
geno-typic coreceptor usage prediction (Fig. 1) (phenogeno-typic analysis
for definitive determination of coreceptor usage was not
per-formed [23]). Sequences obtained from participants D and G
formed distinct clades that were not explained by dual
infec-tion or coreceptor use.
Across all the participants’ phylogenies, HIV-1 sequences
from breast milk specimens were interspersed with those from
blood specimens, without clades comprised exclusively of
blood or milk viruses. RNA and DNA sequences were largely
interspersed across the individual’s phylogenies. Because
cell-free and cell-associated HIV-1 sequences in untreated
individ-uals largely reflect actively replicating virus (33, 35) and
be-cause of the small numbers of sequences of each type available
for analysis, RNA and DNA sequences were combined in the
evaluations of compartmentalization.
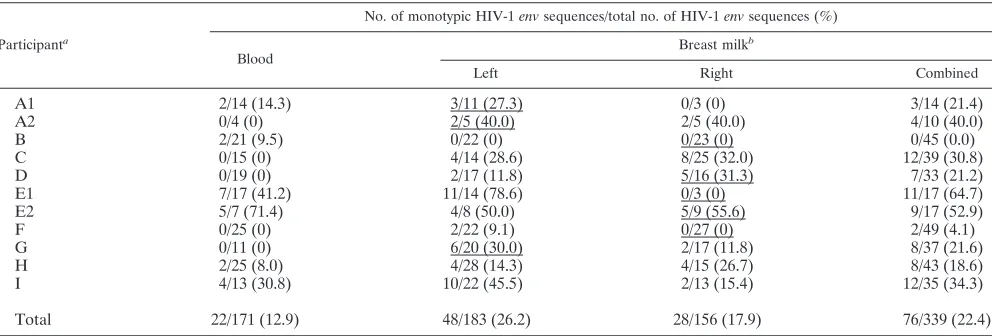
The frequency of finding two or more monotypic sequences
(those with zero nucleotide differences) was similar across
breast milk and blood specimens (15/18 versus 5/9,
respec-tively;
P
⫽
0.175). However, monotypic variants comprised a
greater proportion of all sequences in milk specimens than in
blood specimens (22.4% versus 12.9%;
P
⫽
0.012) (Table 2).
Despite the interspersion of sequences on phylogenetic trees
noted above, statistical evaluation of the population structure
of viral sequences using SM testing based on maximum
likeli-hood phylogenies detected compartmentalization of virus
be-tween breast milk and blood in 6 of 17 (35.3%) specimens
obtained from 4 of 9 individuals (participants B, C, D, and E)
(Table 3). Results were similar when statistical estimation of
compartmentalization was performed using parsimony scores,
association indices, and maximum monophyletic clade sizes
based on Bayesian phylogenies (12, 41) (Table 3). In addition,
performing all analyses using balanced numbers of sequences
from each participant’s samples did not substantially change
estimates of compartmentalization (data not shown). No
evi-dence for compartmentalization between right and left breast
milk specimens was observed (data not shown). After
mono-typic sequences, which can inflate statistical estimates of
pop-ulation structure (7, 8, 21), were collapsed into a single
se-quence, 3 of 6 (50%) specimens no longer showed evidence of
compartmentalization by SM testing (Table 3), reducing the
overall rate to 3 of 17 (17.6%) specimens.
Effect of mastitis on HIV-1 population structure.
The
me-dian concentration of breast milk Na
⫹across all specimens
analyzed was 8 mM (range, 2 to 86 mM), and 6 participants
had mastitis in one breast, as defined by a milk Na
⫹concen-tration of
⬎
12 mM (Table 1) (15, 47). Milk specimens with
mastitis had a median HIV-1 viral load of 45,228 copies/ml
(range, 1,553 to 124,254 copies/ml) compared with that of
2,660 copies/ml (range, 400 to 55,227 copies/ml) in specimens
without mastitis (
P
⫽
0.043).
[image:3.585.42.543.88.244.2]The proportion of monotypic viruses in milk specimens was
not associated with mastitis (18.4% versus 24.4% in samples
with and without mastitis, respectively;
P
⫽
0.218) (Table 2).
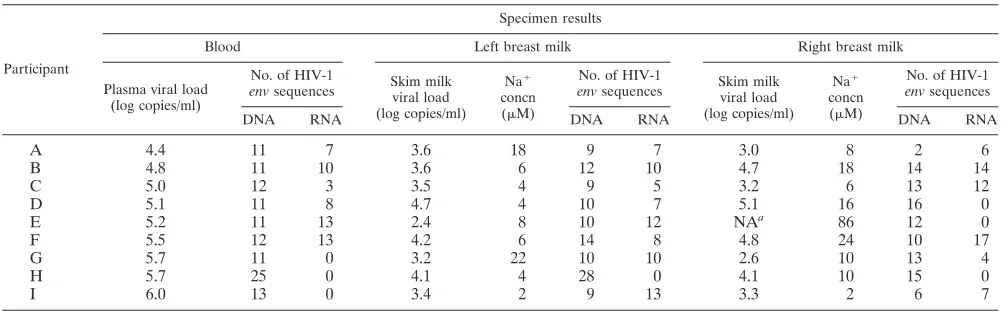
TABLE 1. Plasma and breast milk HIV-1 viral loads, breast milk Na
⫹concentrations, and numbers of
single-template
env
DNA and RNA sequences analyzed
Participant
Specimen results
Blood Left breast milk Right breast milk
Plasma viral load (log copies/ml)
No. of HIV-1
envsequences Skim milkviral load
(log copies/ml)
Na⫹
concn
(M)
No. of HIV-1
envsequences Skim milkviral load
(log copies/ml)
Na⫹
concn
(M)
No. of HIV-1
envsequences
DNA RNA DNA RNA DNA RNA
A
4.4
11
7
3.6
18
9
7
3.0
8
2
6
B
4.8
11
10
3.6
6
12
10
4.7
18
14
14
C
5.0
12
3
3.5
4
9
5
3.2
6
13
12
D
5.1
11
8
4.7
4
10
7
5.1
16
16
0
E
5.2
11
13
2.4
8
10
12
NA
a86
12
0
F
5.5
12
13
4.2
6
14
8
4.8
24
10
17
G
5.7
11
0
3.2
22
10
10
2.6
10
13
4
H
5.7
25
0
4.1
4
28
0
4.1
10
15
0
I
6.0
13
0
3.4
2
9
13
3.3
2
6
7
a
NA⫽sample not available.
10814
GANTT ET AL.
J. V
IROL.
on November 8, 2019 by guest
http://jvi.asm.org/
FIG. 1. HIV-1
env
sequences from both breast milk and blood specimens are present in all phylogenetic clades, suggesting a lack of viral
compartmentalization. Maximum likelihood phylogenetic analysis of the HIV-1
env
region C2-V5 derived from single-amplicon-derived sequences
from RNA (squares) and DNA (circles) from blood specimens (black filled symbols) and breast milk specimens (left breast, gray filled symbols;
right breast, open symbols). Dual HIV-1 infection is evident in participants A and E, with each infecting strain delineated by a bracket. The
phylogeny for participant H shows distinct clades, shown with brackets, which differ by predicted coreceptor usage (CCR5 or CXCR4). Mastitis,
as defined by a breast milk Na
⫹concentration of
⬎
12 mM, was present in samples from subject A (left breast), B (right breast), D to F (right
breast), and G (left breast). Sequences obtained from all participants are rooted by four reference sequences of subtype C (labeled RC1, etc.).
Eight additional subtype C sequences and one each from subtypes A, B, and D were used to root phylogenies for participants A and E in order
to better characterize their dual infections. The number of substitutions per site is indicated by the horizontal line below each phylogeny.
10815
on November 8, 2019 by guest
Similarly, compartmentalization of virus was not associated
with mastitis, with compartmentalization observed in 1 of 6
(16.7%) breast milk specimens with mastitis compared to 5 of
11 (45.5%) of those without mastitis (
P
⫽
0.334) (Table 3) by
SM testing. Furthermore, the median Na
⫹concentration did not
differ significantly between milk specimens with
compartmental-ization detected by SM testing and those without (
P
⫽
0.155).
The relationship between milk Na
⫹concentration as a
marker of inflammation within the breast and the genetic
dis-tance between HIV-1 populations in breast milk and blood
samples was evaluated using the GEE. Breast milk Na
⫹was
positively correlated with the genetic distance between HIV-1
in milk and blood specimens, indicating that with increasing
inflammation HIV-1 populations in milk specimens
progres-sively diverged from those in blood specimens (

coefficient
⫽
3.8
⫻
10
⫺5; 95% confidence interval, 1.6
⫻
10
⫺6to 7.4
⫻
10
⫺5;
P
⫽
0.041).
DISCUSSION
Three observations are noteworthy from our study. First, we
found only limited compartmentalization of HIV-1 in breast
milk specimens, suggesting substantial interchange of viruses
between the milk and blood specimens obtained from the
participants we studied. Second, monotypic sequences were
overrepresented in the HIV-1 populations from milk
speci-mens compared to those from blood specispeci-mens, suggesting
local HIV-1 production within the breast, possibly from the
proliferation of infected cells. Third, the mean genetic distance
between milk and blood viruses correlated positively with
breast milk Na
⫹concentrations, suggesting full cycles of viral
replication within the breast and that the increased breast milk
HIV-1 concentrations observed with mastitis are not simply
due to the passage of virus from blood into breast milk.
Viral compartmentalization was detected in a minority of
breast milk specimens using a panel of statistical methods
commonly used to evaluate population structure. However,
among the participants evaluated, no tissue-specific clades
were observed in phylogenetic trees. Rather, viral sequences
from milk and blood specimens were intermingled, suggesting
that viruses mix readily between these fluids. Similarly, studies
of HIV-1 populations in the genital tract and lung have found
compartmentalization by statistical testing when segregation
by fluid/tissue is not apparent in phylogenies (7, 21). The
im-portance of such compartmentalization is unclear but is
con-sistent with replication or proliferation within a small virus
population (i.e., limited effective population size) (7, 21, 56).
[image:5.585.45.541.80.248.2]Detection and quantification of minority sequence variants
are dependent on the method of sampling and the number of
sequences obtained. We sequenced virus derived from single
PCR amplicons, which prevents bias due to resampling of
individual variants (9, 32). Resampling of viral sequences may
explain the apparent compartmentalization of HIV-1 in breast
milk specimens previously reported for some women (3, 4). In
support of our findings, a study using a heteroduplex tracking
assay was unable to distinguish differences between HIV-1
populations from breast milk and blood specimens obtained
from 8 women (22), as did a recent study that cloned viruses
but employed methods to avoid resampling of viral templates
(20). The time and expense required for the generation of
single-amplicon sequences limits the sample size feasible for
phylogenetic analyses and therefore affects the precision of
estimates of population differences. We analyzed a large
num-ber of sequences, but these sequences were derived from a
small number of nonrandomly selected participants,
poten-tially limiting the generalizability of our findings. In addition, it
was not possible to perform analyses on cell-free virus
popu-lations separately, as few sequences were generated from
breast milk specimens with low concentrations of HIV-1 RNA.
Finally, because the sequences were derived at a single time
point early in lactation, we cannot rule out the possibility that
HIV-1 compartmentalization in an individual’s breast milk
changes significantly over time. Relevant to this last point, a
comparison of the compartmentalization of blood and milk
when these specimens were collected on the same or different
dates gave discrepant results (20). Viruses in specimens
col-lected on the same dates were rarely compartmentalized,
TABLE 2. Monotypic HIV-1
env
sequences in breast milk and plasma specimens
Participanta
No. of monotypic HIV-1envsequences/total no. of HIV-1envsequences (%)
Blood Breast milk
b
Left Right Combined
A1
2/14 (14.3)
3/11 (27.3)
0/3 (0)
3/14 (21.4)
A2
0/4 (0)
2/5 (40.0)
2/5 (40.0)
4/10 (40.0)
B
2/21 (9.5)
0/22 (0)
0/23 (0)
0/45 (0.0)
C
0/15 (0)
4/14 (28.6)
8/25 (32.0)
12/39 (30.8)
D
0/19 (0)
2/17 (11.8)
5/16 (31.3)
7/33 (21.2)
E1
7/17 (41.2)
11/14 (78.6)
0/3 (0)
11/17 (64.7)
E2
5/7 (71.4)
4/8 (50.0)
5/9 (55.6)
9/17 (52.9)
F
0/25 (0)
2/22 (9.1)
0/27 (0)
2/49 (4.1)
G
0/11 (0)
6/20 (30.0)
2/17 (11.8)
8/37 (21.6)
H
2/25 (8.0)
4/28 (14.3)
4/15 (26.7)
8/43 (18.6)
I
4/13 (30.8)
10/22 (45.5)
2/13 (15.4)
12/35 (34.3)
Total
22/171 (12.9)
48/183 (26.2)
28/156 (17.9)
76/339 (22.4)
a
Analyses for those participants (A and E) who had dual HIV-1 infections were performed separately on each of the infecting HIV-1 strains (1, upper clade in phylogenies; 2, lower clade in phylogenies) (Fig. 1).
b
Breast milk specimens with mastitis (Na⫹concentrations of⬎12 mM) are underlined.
10816
GANTT ET AL.
J. V
IROL.
on November 8, 2019 by guest
http://jvi.asm.org/
whereas viruses in those collected more than 100 days apart
were consistently compartmentalized across statistical
anal-yses, suggesting that significant evolution occurred in this time
frame.
Even using methods that preclude resampling, identical
se-quences were found in the majority of specimens. Indeed, we
may have slightly underestimated the number of identical
se-quences, as approximately half of the sequences were
gener-ated by cloning, which we estimate could introduce an average
of
⬍
1- to 3-base-pair changes per sequence (14). These
mono-typic sequences likely reflect very recent viral replication or, in
the case of DNA sequences, the clonal expansion of
HIV-1-infected cells (2, 7, 8, 21, 53). Thus, the disproportionate
prev-alence of monotypic sequences in breast milk specimens
com-pared to those in blood specimens may represent a relatively
greater production of HIV-1 in milk. Alternatively, the
likeli-hood of sampling identical sequences from blood specimens
compared to that from breast milk specimens may differ due to
the large total circulating volume and higher HIV-1
concen-trations in blood than in milk. It is also possible that the fact
that slightly more blood sequences than milk sequences were
obtained by cloning and may include additional errors
intro-duced during PCR resulted in an underestimate of monotypic
viruses in blood specimens. In either case, the presence of
identical sequences can bias statistical estimates of population
structure (2, 7, 8, 21). Of note, monotypic sequences accounted
for half of the instances of compartmentalization detected.
Our findings are remarkably similar to those from studies of
rhesus macaques (42) that had large numbers of monotypic
SIV
env
sequences detected in the breast milk specimens but
not in blood specimens. Furthermore, although breast milk
and blood simian immunodeficiency virus (SIV) sequences
were interspersed in phylogenetic trees, SM testing detected
compartmentalization that was again diminished by exclusion
of monotypic sequences. We contend that
compartmentaliza-tion based on monotypic or nearly identical sequences may not
reflect a separate viral population but rather might be a
snap-shot of recent events, namely, a burst of viral replication and/or
proliferation of HIV-1-infected cells.
Mastitis is associated with increased breast milk HIV-1
loads, presumably by either influx of virions from the blood
and/or local viral production within the breast (15, 31, 47, 55).
We observed a statistically significant positive correlation
be-tween breast milk Na
⫹concentrations and the genetic distance
between milk and blood HIV-1 subpopulations. This is
consis-tent with increased replication of HIV-1 within the breast due
to mastitis, resulting in divergence of breast milk HIV-1
sub-populations from those in blood. The plausibility of mastitis
stimulating local HIV-1 replication in the breast is supported
by observations of the effects of infection and inflammation in
other settings. For example, increased production of HIV-1 in
lungs (29, 37) and lymph nodes (39) has been associated with
opportunistic infections, and viral replication can be stimulated
in T cells by various inflammatory cytokines (27, 54).
When inflammation renders mammary epithelial tight
junc-tions permeable, HIV-1 from the blood may more easily enter
breast milk, as occurs with other blood components (15, 16, 36,
47, 55). Although the passage of HIV-1 from blood into breast
milk may contribute to the increase in milk HIV-1
concentra-tions due to mastitis, our observaconcentra-tions suggest that this is not
TABLE
3.
Evaluation
of
breast
milk
HIV-1
compartmentalization
using
dif
ferent
statistical
methods
Participant a P values for breast milk vs. blood using indicated methods b Left breast Right breast SM test PS AI MC, milk MC, blood SM test PS AI MC, milk MC, bloodA1
0.201
(0.404)
0.090
(0.40)
0.260
(0.520)
0.120
(0.450)
0.930
(0.880)
—
—
———
A
2
—
—
—
—
————
—
—
B
0.011
(0.030)
<0.001
(<0.001)
<0.001
(0.010)
0.010
(0.010)
0.340
(0.360)
0.392
(0.505)
0.080
(0.180)
0.020
(0.070)
0.010
(0.020)
0.430
(0.350)
C
0.003
(0.010)
<0.001
(<0.001)
<0.001
(<0.001)
0.060
(0.050)
<0.001
(0.090)
0.032
(0.124)
0.050
(0.140)
0.100
(0.150)
0.180
(0.510)
0.710
(0.800)
D
0.001
(0.002)
<0.001
(<0.001)
<0.001
(0.010)
0.250
(0.150)
0.050
(0.050)
0.018
(0.153)
<0.001
(0.999)
0.020
(0.160)
0.280
(0.999)
0.010
(0.010)
E1
0.023
(—)
0.010
(—)
0.120
(—)
0.010
(—)
0.430
(—)
—
—
—
—
—
E2
0.008
(—)
0.070
(—)
0.280
(0.280)
0.720
(0.430)
0.310
(0.650)
0.854
(—)
0.780
(—)
0.710
(—)
0.910
(—)
0.410
(—)
F
0.484
(0.573)
0.470
(0.620)
0.140
(0.500)
0.410
(0.430)
0.590
(0.650)
0.741
(0.741)
0.580
(0.580)
0.230
(0.230)
0.510
(0.510)
0.999
(0.999)
G
0.540
(0.701)
0.380
(0.590)
0.480
(0.500)
0.080
(0.170)
0.590
(0.110)
0.125
(0.174)
0.540
(0.620)
0.240
(0.480)
0.970
(0.999)
0.060
(0.150)
H
0.613
(0.613)
0.550
(0.390)
0.770
(0.740)
0.999
(0.999)
0.590
(0.410)
0.097
(0.314)
0.020
(0.210)
0.030
(0.170)
0.110
(0.710)
0.820
(0.800)
I
0.059
(0.443)
0.220
(0.930)
0.280
(0.800)
0.190
(0.970)
0.650
(0.490)
0.340
(0.651)
0.210
(0.480)
0.440
(0.890)
0.870
(0.810)
0.890
(0.710)
a Analyses for those participants (A and E) who had dual HIV-1 infections were performed separately on each of the infecting HIV-1 strains (1, upper clad e in phylogenies; 2, lower clade in phylogenies) (Fig. 1). b P values shown are derived from comparing all sequences, excluding monotypic sequences, from the breast milk and blood specimens obtained from each su bject using various methods, as follows: Slatkin and Maddison (SM) test, parsimony score (PS), association index (AI), and maximum single-state clade size (MC). In parentheses are the P values for the same comparison after elimination of identical sequences within a specimen. P values of ⬍ 0.05 are shown in boldface. For the MC test, a P value ⬍ 0.05 for either the milk or blood sample is considered evidence of compartmentalization. Specimens yielding ⬍ 5 sequences were not analyzed for compartmentalization by SM (indicated by “—”). Breast milk specimens with mastitis (Na ⫹ concentrations of ⬎ 12 mM) are underlined.on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.60.248.66.726.2]the sole mechanism, because an influx of blood viruses would
decrease the genetic differences between these subpopulations.
While the absolute divergence of breast milk HIV-1 we
de-tected in mastitic milk was small, it is important to note that
the duration of increased viral replication would at most be 6
to 16 weeks in duration due to the timing of sample collection
after the onset of lactation. In addition, the magnitude of the
genetic differences between milk and blood HIV-1 may have
been diminished by the countervailing contribution of blood
virus leaking into milk. Of note, although there was a
signifi-cant association between milk Na
⫹concentrations and the
genetic distance between HIV populations in blood and milk
specimens, we did not detect an association between mastitis
and the prevalence of monotypic sequences in milk specimens.
This discrepancy suggests that mastitis may stimulate viral
rep-lication across a larger population of virus in the infected
breast compared to that in the healthy breast, perhaps due to
an influx of infected inflammatory cells. Furthermore,
com-partmentalization of virus in breast milk specimens was not
associated with mastitis, suggesting either that a relatively
di-verse group of viruses replicated in milk specimens or that the
net effect of inflammation on replication within the breast was
not sufficient to affect HIV-1 population structure by these
measures.
In summary, phylogenetic analyses of HIV-1 genomes from
breast milk and blood specimens obtained from lactating
women found limited viral compartmentalization, indicating
relatively free mixing of viruses between milk and blood
spec-imens. The greater prevalence of monotypic HIV-1 in breast
milk specimens suggests viral replication and/or proliferation
of cells containing proviruses within the breast. Statistical
mod-eling of the effect of inflammation on genetic distance between
HIV-1 subpopulations in milk and blood specimens found
ev-idence for increased HIV-1 replication in breasts with mastitis.
ACKNOWLEDGMENTS
We are grateful to the participants. We appreciate the contributions
of Mary Mucheche and Lynda Stranix-Chibanda in participant
enroll-ment; Rhona Jack, Ingrid Beck, Patrick Abe, and Kuda Matasa for
technical assistance; and the Zimbabwe Ministry of Health, the
Chi-tungwiza Health Department, and the Department of Pediatrics and
Child Health, University of Zimbabwe College of Health Sciences.
This work was supported by the National Institutes of Health grants
R21 AI065288 (to L.M.F.), KL2 RR025015 (to S.G.), and T32
HD07233 (to S.G.) and the University of Washington Center for AIDS
Research Computational Biology Core (grant P30 AI27757 to J.I.M.).
We all made substantial contributions to the project and agree to
our positions in the byline.
We have no conflicts of interest to declare.
REFERENCES
1.Altfeld, M., E. S. Rosenberg, R. Shankarappa, J. S. Mukherjee, F. M. Hecht, R. L. Eldridge, M. M. Addo, S. H. Poon, M. N. Phillips, G. K. Robbins, P. E. Sax, S. Boswell, J. O. Kahn, C. Brander, P. J. Goulder, J. A. Levy, J. I. Mullins, and B. D. Walker. 2001. Cellular immune responses and viral diversity in individuals treated during acute and early HIV-1 infection. J.
Exp. Med.193:169–180.
2.Bailey, J. R., A. R. Sedaghat, T. Kieffer, T. Brennan, P. K. Lee, M. Wind-Rotolo, C. M. Haggerty, A. R. Kamireddi, Y. Liu, J. Lee, D. Persaud, J. E. Gallant, J. Cofrancesco, Jr., T. C. Quinn, C. O. Wilke, S. C. Ray, J. D. Siliciano, R. E. Nettles, and R. F. Siliciano.2006. Residual human immu-nodeficiency virus type 1 viremia in some patients on antiretroviral therapy is dominated by a small number of invariant clones rarely found in
circulat-ing CD4⫹T cells. J. Virol.80:6441–6457.
3.Becquart, P., N. Chomont, P. Roques, A. Ayouba, M. D. Kazatchkine, L. Belec, and H. Hocini.2002. Compartmentalization of HIV-1 between breast
milk and blood of HIV-infected mothers. Virology300:109–117.
4.Becquart, P., V. Courgnaud, J. Willumsen, and P. Van de Perre.2007. Diversity of HIV-1 RNA and DNA in breast milk from HIV-1-infected
mothers. Virology363:256–260.
5.Bedri, A., B. Gudetta, A. Isehak, S. Kumbi, S. Lulseged, Y. Mengistu, A. V. Bhore, R. Bhosale, V. Varadhrajan, N. Gupte, J. Sastry, N. Suryavanshi, S. Tripathy, F. Mmiro, M. Mubiru, C. Onyango, A. Taylor, P. Musoke, C. Nakabiito, A. Abashawl, R. Adamu, G. Antelman, R. C. Bollinger, P. Bright, M. A. Chaudhary, J. Coberly, L. Guay, M. G. Fowler, A. Gupta, E. Hassen, J. B. Jackson, L. H. Moulton, U. Nayak, S. B. Omer, L. Propper, M. Ram, V. Rexroad, A. J. Ruff, A. Shankar, and S. Zwerski.2008. Extended-dose ne-virapine to 6 weeks of age for infants to prevent HIV transmission via breastfeeding in Ethiopia, India, and Uganda: an analysis of three
random-ised controlled trials. Lancet372:300–313.
6.Boom, R., C. Sol, M. Salimans, C. Jansen, P. W.-V. Dillen, and J. V. d. Noordaa.1990. Rapid and simple method for purification of nucleic acids.
J. Clin. Microbiol.28:495–503.
7.Bull, M., G. Learn, I. Genowati, J. McKernan, J. Hitti, D. Lockhart, K. Tapia, S. Holte, J. Dragavon, R. Coombs, J. Mullins, and L. Frenkel.2009. Compartmentalization of HIV-1 within the female genital tract is due to monotypic and low-diversity variants not distinct viral populations. PLoS One4:e7122.
8.Bull, M. E., G. H. Learn, S. McElhone, J. Hitti, D. Lockhart, S. Holte, J. Dragavon, R. W. Coombs, J. I. Mullins, and L. M. Frenkel.2009. Monotypic human immunodeficiency virus type 1 genotypes across the uterine cervix
and in blood suggest proliferation of cells with provirus. J. Virol.83:6020–
6028.
9.Bull, M. E., G. H. Learn, J. I. Mullins, and L. M. Frenkel.2007. Methods used to examine compartmentalization of viral populations between the
genital tract and peripheral blood. J. Infect. Dis.196:493–494. (Author’s
reply,196:494–495.)
10.Delwart, E. L., J. I. Mullins, P. Gupta, G. H. Learn, Jr., M. Holodniy, D. Katzenstein, B. D. Walker, and M. K. Singh.1998. Human
immunodefi-ciency virus type 1 populations in blood and semen. J. Virol.72:617–623.
11.Deng, W., B. S. Maust, D. C. Nickle, G. H. Learn, Y. Liu, L. Heath, S. L. K. Pond, and J. I. Mullins.2010. DIVEIN: a web server to analyze phylogenies,
sequence divergence, diversity, and informative sites. Biotechniques48:405–
408.
12.Drummond, A. J., G. K. Nicholls, A. G. Rodrigo, and W. Solomon.2002. Estimating mutation parameters, population history and genealogy
simulta-neously from temporally spaced sequence data. Genetics161:1307–1320.
13.Drummond, A. J., and A. Rambaut.2007. BEAST: Bayesian evolutionary
analysis by sampling trees. BMC Evol. Biol.7:214.
14.Eckert, R. A., and T. A. Kunkel.1991. DNA polymerase fidelity and the
polymerase chain reaction. PCR Methods Appl.1:17–24.
15.Gantt, S., A. K. Shetty, K. D. Seidel, K. Matasa, G. Musingwini, G. Woelk, L. S. Zijenah, D. A. Katzenstein, and L. M. Frenkel. 2007. Laboratory indicators of mastitis are not associated with elevated HIV-1 DNA loads or
predictive of HIV-1 RNA loads in breast milk. J. Infect. Dis.196:570–576.
16.Georgeson, J. C., and S. M. Filteau.2000. Physiology, immunology, and
disease transmission in human breast milk. AIDS Patient Care STDS.14:
533–539.
17.Guay, L. A., P. Musoke, T. Fleming, D. Bagenda, M. Allen, C. Nakabiito, J. Sherman, P. Bakaki, C. Ducar, M. Deseyve, L. Emel, M. Mirochnick, M. G. Fowler, L. Mofenson, P. Miotti, K. Dransfield, D. Bray, F. Mmiro, and J. B. Jackson.1999. Intrapartum and neonatal single-dose nevirapine compared with zidovudine for prevention of mother-to-child transmission of HIV-1 in
Kampala, Uganda: HIVNET 012 randomised trial. Lancet354:795–802.
18.Guindon, S., and O. Gascuel.2003. A simple, fast, and accurate algorithm to
estimate large phylogenies by maximum likelihood. Syst. Biol.52:696–704.
19.Harris, J. R., S. K. Greene, T. K. Thomas, R. Ndivo, J. Okanda, R. Masaba, I. Nyangau, M. C. Thigpen, R. M. Hoekstra, and R. E. Quick.2009. Effect of a point-of-use water treatment and safe water storage intervention on
diar-rhea in infants of HIV-infected mothers. J. Infect. Dis.200:1186–1193.
20.Heath, L., S. Conway, L. Jones, K. Semrau, K. Nakamura, J. Walter, W. D. Decker, J. Hong, T. Chen, M. Heil, M. Sinkala, C. Kankasa, D. M. Thea, L. Kuhn, J. I. Mullins, and G. M. Aldrovandi.2010. Restriction of HIV-1 genotypes in breast milk does not account for the population transmission
genetic bottleneck that occurs following transmission. PLoS One5:e10213.
21.Heath, L., A. Fox, J. McClure, K. Diem, A. B. van ’t Wout, H. Zhao, D. R. Park, J. T. Schouten, H. L. Twigg, L. Corey, J. I. Mullins, and J. E. Mittler. 2009. Evidence for limited genetic compartmentalization of HIV-1 between
lung and blood. PLoS One4:e6949.
22.Henderson, G. J., N. G. Hoffman, L. H. Ping, S. A. Fiscus, I. F. Hoffman, K. M. Kitrinos, T. Banda, F. E. Martinson, P. N. Kazembe, D. A. Chilongozi, M. S. Cohen, and R. Swanstrom.2004. HIV-1 populations in blood and
breast milk are similar. Virology330:295–303.
23.Jensen, M. A., G. S. Gottlieb, A. B. van ’t Wout, F. S. Li, D. C. Nickle, K. Wong, H. X. He, S. McLaughlin, R. Shankarappa, J. B. Margolick, and J. I. Mullins.2003. A bioinformatic predictor of coreceptor usage corre-lates with markers of disease progression and supports the gradual evo-lution of X4 virus via R5X4 intermediates, poster 498. Abstr. 10th Conf. Retroviruses Opportunistic Infect., Boston, MA.
10818
GANTT ET AL.
J. V
IROL.
on November 8, 2019 by guest
http://jvi.asm.org/
24.John, G. C., R. W. Nduati, D. A. Mbori-Ngacha, B. A. Richardson, D. Panteleeff, A. Mwatha, J. Overbaugh, J. Bwayo, J. O. Ndinya-Achola, and J. K. Kreiss.2001. Correlates of mother-to-child human immunodeficiency virus type 1 (HIV-1) transmission: association with maternal plasma HIV-1 RNA load, genital HIV-1 DNA shedding, and breast infections. J. Infect. Dis.183:206–212.
25.Kilewo, C., K. Karlsson, A. Massawe, E. Lyamuya, A. Swai, F. Mhalu, and G. Biberfeld. 2008. Prevention of mother-to-child transmission of HIV-1 through breast-feeding by treating infants prophylactically with lamivudine in Dar es Salaam, Tanzania: the Mitra Study. J. Acquir. Immune Defic.
Syndr.48:315–323.
26.Kilewo, C., K. Karlsson, M. Ngarina, A. Massawe, E. Lyamuya, A. Swai, R. Lipyoga, F. Mhalu, and G. Biberfeld.2009. Prevention of mother-to-child transmission of HIV-1 through breastfeeding by treating mothers with triple antiretroviral therapy in Dar es Salaam, Tanzania: the Mitra Plus Study. J.
Acquir. Immune Defic. Syndr.52:406–416.
27.Kinter, A., A. Moorthy, R. Jackson, and A. S. Fauci.2003. Productive HIV
infection of resting CD4⫹T cells: role of lymphoid tissue microenvironment
and effect of immunomodulating agents. AIDS Res. Hum. Retroviruses 19:847–856.
28.Koulinska, I. N., E. Villamor, B. Chaplin, G. Msamanga, W. Fawzi, B. Renjifo, and M. Essex.2006. Transmission of cell-free and cell-associated
HIV-1 through breast-feeding. J. Acquir. Immune Defic. Syndr.41:93–99.
29.Koziel, H., S. Kim, C. Reardon, X. Li, R. Garland, P. Pinkston, and H. Kornfeld.1999. Enhanced in vivo human immunodeficiency virus-1 replica-tion in the lungs of human immunodeficiency virus-infected persons with
Pneumocystis carinii pneumonia. Am. J. Respir. Crit. Care Med.160:2048–
2055.
30.Kumwenda, N. I., D. R. Hoover, L. M. Mofenson, M. C. Thigpen, G. Kafu-lafula, Q. Li, L. Mipando, K. Nkanaunena, T. Mebrahtu, M. Bulterys, M. G. Fowler, and T. E. Taha.2008. Extended antiretroviral prophylaxis to reduce
breast-milk HIV-1 transmission. N. Engl. J. Med.359:119–129.
31.Lee, E. J., R. Kantor, L. Zijenah, W. Sheldon, L. Emel, P. Mateta, E. Johnston, J. Wells, A. K. Shetty, H. Coovadia, Y. Maldonado, S. A. Jones, L. M. Mofenson, C. H. Contag, M. Bassett, and D. A. Katzenstein.2005. Breast-milk shedding of drug-resistant HIV-1 subtype C in women exposed
to single-dose nevirapine. J. Infect. Dis.192:1260–1264.
32.Liu, S.-L., A. G. Rodrigo, R. Shankarappa, G. H. Learn, L. Hsu, O. Davidov, L. P. Zhao, and J. I. Mullins.1996. HIV quasispecies and resampling.
Science273:415–416.
33.Liu, S. L., T. Schacker, L. Musey, D. Shriner, M. J. McElrath, L. Corey, and J. I. Mullins.1997. Divergent patterns of progression to AIDS after infection from the same source: human immunodeficiency virus type 1 evolution and
antiviral responses. J. Virol.71:4284–4295.
34.Maddison, D. R., and W. P. Maddison.2000. MacClade 4: analysis of phy-logeny and character evolution. Sinauer Associates, Inc., Sunderland, MA. 35.Mohri, H., A. S. Perelson, K. Tung, R. M. Ribeiro, B. Ramratnam, M. Markowitz, R. Kost, A. Hurley, L. Weinberger, D. Cesar, M. K. Hellerstein, and D. D. Ho.2001. Increased turnover of T lymphocytes in HIV-1 infection
and its reduction by antiretroviral therapy. J. Exp. Med.194:1277–1287.
36.Morton, J. A.1994. The clinical usefulness of breast milk sodium in the
assessment of lactogenesis. Pediatrics93:802–806.
37.Nakata, K., W. N. Rom, Y. Honda, R. Condos, S. Kanegasaki, Y. Cao, and M. Weiden.1997. Mycobacterium tuberculosis enhances human
immunodefi-ciency virus-1 replication in the lung. Am. J. Respir. Crit. Care Med.155:
996–1003.
38.Nduati, R., G. John, D. Mbori-Ngacha, B. Richardson, J. Overbaugh, A. Mwatha, J. Ndinya-Achola, J. Bwayo, F. E. Onyango, J. Hughes, and J. Kreiss.2000. Effect of breastfeeding and formula feeding on transmission of
HIV-1: a randomized clinical trial. JAMA283:1167–1174.
39.Orenstein, J. M., C. Fox, and S. M. Wahl.1997. Macrophages as a source of
HIV during opportunistic infections. Science276:1857–1861.
40.Palombi, L., M. C. Marazzi, A. Voetberg, and N. A. Magid.2007. Treatment acceleration program and the experience of the DREAM program in
pre-vention of mother-to-child transmission of HIV. AIDS21(Suppl. 4):S65–
S71.
41.Parker, J., A. Rambaut, and O. G. Pybus.2008. Correlating viral phenotypes with phylogeny: accounting for phylogenetic uncertainty. Infect. Genet. Evol. 8:239–246.
42.Permar, S. R., H. H. Kang, A. B. Wilks, L. V. Mach, A. Carville, K. G. Mansfield, G. H. Learn, B. H. Hahn, and N. L. Letvin.2010. Local replica-tion of simian immunodeficiency virus in the breast milk compartment of
chronically infected, lactating rhesus monkeys. Retrovirology7:7.
43.Pillay, K., A. Coutsoudis, D. York, L. Kuhn, and H. M. Coovadia.2000. Cell-free virus in breast milk of HIV-1-seropositive women. J. Acquir.
Im-mune Defic. Syndr.24:330–336.
44.Poss, M., A. G. Rodrigo, J. J. Gosink, G. H. Learn, D. de Vange Panteleeff, H. L. Martin, Jr., J. Bwayo, J. K. Kreiss, and J. Overbaugh.1998. Evolution of envelope sequences from the genital tract and peripheral blood of women
infected with clade A human immunodeficiency virus type 1. J. Virol.72:
8240–8251.
45.Rodrigo, A. G., P. C. Goracke, K. Rowhanian, and J. I. Mullins.1997. Quantitation of target molecules from polymerase chain reaction-based
lim-iting dilution assays. AIDS Res. Hum. Retroviruses13:737–742.
46.Rousseau, C. M., R. W. Nduati, B. A. Richardson, M. S. Steele, G. C. John-Stewart, D. A. Mbori-Ngacha, J. K. Kreiss, and J. Overbaugh.2003. Longitudinal analysis of human immunodeficiency virus type 1 RNA in breast milk and of its relationship to infant infection and maternal disease.
J. Infect. Dis.187:741–747.
47.Semba, R. D., N. Kumwenda, D. R. Hoover, T. E. Taha, T. C. Quinn, L. Mtimavalye, R. J. Biggar, R. Broadhead, P. G. Miotti, L. J. Sokoll, L. van der Hoeven, and J. D. Chiphangwi.1999. Human immunodeficiency virus load in breast milk, mastitis, and mother-to-child transmission of human
immuno-deficiency virus type 1. J. Infect. Dis.180:93–98.
48.Shankarappa, R., J. B. Margolick, S. J. Gange, A. G. Rodrigo, D. Upchurch, H. Farzadegan, P. Gupta, C. R. Rinaldo, G. H. Learn, X. He, X. L. Huang, and J. I. Mullins.1999. Consistent viral evolutionary changes associated with the progression of human immunodeficiency virus type 1 infection. J. Virol. 73:10489–10502.
49.Slatkin, M., and W. P. Maddison.1989. A cladistic measure of gene flow
inferred from the phylogenies of alleles. Genetics123:603–613.
50.Strain, M. C., S. Letendre, S. K. Pillai, T. Russell, C. C. Ignacio, H. F. Gunthard, B. Good, D. M. Smith, S. M. Wolinsky, M. Furtado, J. Marquie-Beck, J. Durelle, I. Grant, D. D. Richman, T. Marcotte, J. A. McCutchan, R. J. Ellis, and J. K. Wong.2005. Genetic composition of human immuno-deficiency virus type 1 in cerebrospinal fluid and blood without treatment
and during failing antiretroviral therapy. J. Virol.79:1772–1788.
51.Thior, I., S. Lockman, L. M. Smeaton, R. L. Shapiro, C. Wester, S. J. Heymann, P. B. Gilbert, L. Stevens, T. Peter, S. Kim, E. van Widenfelt, C. Moffat, P. Ndase, P. Arimi, P. Kebaabetswe, P. Mazonde, J. Makhema, K. McIntosh, V. Novitsky, T. H. Lee, R. Marlink, S. Lagakos, and M. Essex. 2006. Breastfeeding plus infant zidovudine prophylaxis for 6 months versus formula feeding plus infant zidovudine for 1 month to reduce mother-to-child HIV transmission in Botswana. A randomized trial: the Mashi Study. JAMA296:794–805.
52.Thompson, J. D., D. G. Higgins, and T. J. Gibson.1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice.
Nucleic Acids Res.22:4673–4680.
53.Tobin, N. H., G. H. Learn, S. E. Holte, Y. Wang, A. J. Melvin, J. L. McK-ernan, D. Pawluk, K. M. Mohan, P. F. Lewis, J. I. Mullins, and L. M. Frenkel.2005. Evidence that low-level viremias during effective highly active antiretroviral therapy result from two processes: expression of archival virus
and replication of virus. J. Virol.79:9625–9634.
54.Unutmaz, D., V. N. KewalRamani, S. Marmon, and D. R. Littman.1999. Cytokine signals are sufficient for HIV-1 infection of resting human T
lym-phocytes. J. Exp. Med.189:1735–1746.
55.Willumsen, J. F., S. M. Filteau, A. Coutsoudis, K. E. Uebel, M. L. Newell, and A. M. Tomkins.2000. Subclinical mastitis as a risk factor for
mother-infant HIV transmission. Adv. Exp. Med. Biol.478:211–223.
56.Zarate, S., S. L. Pond, P. Shapshak, and S. D. Frost.2007. Comparative study of methods for detecting sequence compartmentalization in human
immunodeficiency virus type 1. J. Virol.81:6643–6651.
57.Zhu, T., N. Wang, A. Carr, D. S. Nam, R. Moor-Jankowski, D. A. Cooper, and D. D. Ho.1996. Genetic characterization of human immunodeficiency virus type 1 in blood and genital secretions: evidence for viral
compartmen-talization and selection during sexual transmission. J. Virol.70:3098–3107.
on November 8, 2019 by guest
http://jvi.asm.org/