JOURNAL OFVIROLOGY,Dec.1994,p.8304-8311 Vol.68, No.12 0022-538X/94/$04.00+0
Copyright © 1994,AmericanSocietyforMicrobiology
Differential Response of
Human
Cells
to
Deletions and
Stop
Codons in the
Yi34.5 Gene of
Herpes
Simplex
Virus
JOANY CHOU, ALICE P. W. POON,JENNIFERJOHNSON,ANDBERNARD ROIZMAN*
The
Marjorie
B. Kovler Viral Oncology Laboratories, The University of Chicago, Chicago Illinois 60637Received28July 1994/Accepted21September 1994
Earlier studies haveshown thatherpessimplexvirus mutantslackingthe
-y,34.5
genearetotallyavirulent onintracerebral inoculationof the virusinto miceand induceprematureshutoff ofprotein synthesisinhuman neuroblastoma (SK-N-SH) cells butnotinVero cells.We report thefollowing. (i) Whereas deletionmutant R3616, lacking 1,000 bp of they,34.5
gene, caused premature shutoff ofproteinsynthesis in both SK-N-SH and humanforeskin fibroblasts(HFF), mutants R4009 and R930(mutantF),carryingstopcodons in allsixframes, 27 and 210 codons from the initiation codon of the-y,34.5
genes, respectively, induced shutoff ofprotein synthesisinSK-N-SHcellsbut not in HFF. The differences in behavior between the R3616 deletion andR4009 stopcodon mutants cannotbe attributedtodifferencesin the rateof inductionof premature shutoffofprotein synthesis and the multiplicity of infection. HFF do not produce detectable truncated'y34.5
protein or truncatedmRNA. (ii) Someclonal lines ofSK-N-SHcellscarrying
a-y34.5
genedrivenbyametallothionein promoter express the-y34.5geneconstitutivelyand do notrequire induction by cadmiumtocomplementthe-y,34.5-
virus. Oneclonal
cell linecomplements
the'y34.5-
virusonly
after inductionby
cadmium. These resultsareconsistent withprevious conclusions that thephenotypeof premature shutoff ofproteinsynthesis isassociated with absence of they,34.5
protein and indicate that theamountsof-y,34.5
protein necessaryto complement the-y34.5-virusesare small. We conclude that human cells differ inthe manner in whichthey respond to the presence of stop codons. Shutoff of protein synthesis in HFF infected with the stop codon mutantscould have been precluded bysmall amountsof-y,34.5
protein producedby splicingoutofanintron containing the stop codon, downstream initiation oftranslation,ortRNAsuppressionof the stopcodon. Several years ago,areportfrom thislaboratorydescribedamost unusual herpes simplex virus 1 (HSV-1) open reading frame(ORF). This ORF, designated
-Yi34.5,
is located in the ab and b'a' inverted repeat sequences flanking the long (L)componentofthe HSV-1 DNA and is therefore present intwo
copies per genome (5). Its TATAA-less promoter was
con-tainedinthe terminalasequence.Thenucleotide sequenceof the HSV-1 strain F [HSV-1(F)]
-y34.5
gene predicted a protein of 263 amino acids consisting of a 159-amino-acidamnino-terminal
domain, 10 repeats of the amino acids Ala, Thr, and Pro, and a 74-amino-acid carboxyl-terminal domain. Because the number oftriplet repeats varied from strain tostrain,its rolewasmostlikely that ofalinker or swivelregion of the protein (6). Agenetically engineered mutant lacking 1,000 bp of the coding domain replicated in Vero cells to the same level as the wild-type parent HSV-1(F) and the recom-binant virus in which the deleted sequences were restored
[HSV-1(F)R]. The deletion mutant did not multiply in the central nervous systems of mice given the virus by the intra-cerebral route, although limited multiplication was seen in miceinoculatedby other routes(4).The phenotypic properties of another mutant, R4009, which carries a stop codon in all six frames inserted between codons 27 and 28 of the
-Yi34.5
gene, were identical to those of the R3616 deletion mutant (4).A remarkable property of the R3616 and R4009 mutants emerged in the course of studies of their replication in human neuroblastoma cell line SK-N-SH. Whereas the phenotype of Verocells infected with these mutants could not be differen-tiated from that of those infected with the wild-type or
*Correspondingauthor. Mailing address:The
Marorie
B.KovlerViralOncology Laboratories,TheUniversityofChicago, 910 E. 58th St.,Chicago,IL60637. Phone:(312) 702-1898. Fax:(312) 702-1631.
repaired virus, in SK-N-SH cells infected with the R3616 and R4009 mutants, therewasapremature, total shutoff of protein synthesis (7). The shutoff of proteinsynthesiswastriggered by theonset of viral DNAsynthesis inasmuch as in infected cells treated with phosphonoacetate, a specific inhibitor of viral DNAsynthesis, the premature shutoff of proteinsynthesis did not ensue.Premature shutoff ofproteinsynthesiswastherefore acelltype-specificstressresponseinducedby theonsetof viral DNAsynthesis in the absence of the
yi34.5
geneproduct (7). The predicted amino acid sequence of the-y34.5
gene is partially homologous to two eukaryotic genes, MyD116 andGADD34,whicharemembersofadiversefamilyknownasthe growth arrestand DNAdamage (GADD)genes. Thevarious members ofthisfamily of genesare expressed in cellswhose
growthis arrestedas aconsequenceofdifferentiationorwhose DNAisdamaged.Thefunctionofthese genes appears to be to cause growth arrest and protect cells from programmed cell death(10). Thus, MyD116, the first of the homologs described, is inducedbyinterleukin 6 in murine myeloid leukemia cells. TheMyD116proteinispredicted to consist of 657 amino acids
comprising a long amino-terminal domain, a sequence of 38 amino acids repeated 4.5 times, and a short carboxyl-terminal domain (14). The size and sequence arrangement of the
predictedproduct of GADD34,ahamster gene, are similar to those of theMyD116 protein.Astretchof 64 amino acids of the carboxyl-terminal domain of the
-Yi34.5
protein shows a high degree of homology to the corresponding domains of the MyD116 and GADD34 proteins. Analyses of a series of in-frame deletions and insertions in the-yi34.5
gene have shown that thecarboxyl terminus ofthe-Yi34.5 geneis neces-sary to preclude premature shutoff of protein synthesis and thus suggest a relationship between the functions of the GADD andyi34.5
genes(8).It has also been reported that the strand antisense to the
8304
on November 9, 2019 by guest
http://jvi.asm.org/
DELETIONS AND STOP CODONS IN HSV
-y34.5
GENE 8305-y34.5
gene istranscribed in cells inwhichthemajor
regulatory
protein, infected cell protein 4 is not expressed or is inactive. Work done in this laboratory recently showed that an ORF designated ORF-P that iscontained in the transcribed domain is expressed. The ORF is antisense but overlaps the coding domain of the
-y34.5
gene, and therefore all mutations and deletions introduced into theYi34.5
genewould also appear as deletions in ORF-P (13).This report concerns twoobservations relevant to the func-tion ofthe
-y34.5
gene. First, we report that-y34.5
deletion mutant R3616, but not mutantR4009, carrying the stop codonafter codon27, induces premature shutoff of protein synthesis in humanforeskin fibroblasts (HFF). The discordance between
SK-N-SHand HFFcultures was also noted in another mutant, mutantF(8), carrying a stop codon at codon 210 of the
-Yi34.5
gene. These studies indicate that interaction of cellular ma-chinerywith stop codons may be cell type dependent.
Second, we report that SK-N-SH clonal cell lines carrying the -Yi34.5 gene fused to a mouse metallothionein promoter and expressing the gene after cadmium induction comple-mented the R3616 mutant in precluding premature shutoff of proteinsynthesis. The results of these experiments support the conclusion that premature shutoff of protein synthesis is due to absenceof the -y34.5geneproduct inHSV-infected cells.
MATERUILSANDMETHODS
Cells. The Vero and human neuroblastoma SK-N-SH cell lines used were obtained from the American Type Culture
Collection. The HFF cell strain used was obtained from George Kemble (Aviron Inc., Burlingame, Calif.). The cells were propagated in Dulbecco's modified Eagle's medium
supplemented with 5% (Vero cells) or 10% (SK-N-SH cells andHFF)fetal bovine serum.
Viruses.HSV-1(F)is the prototype HSV-1 strain used in this laboratory (9). The construction and phenotype of all other
-Yi34*5
genemutants havebeen described elsewhere(4, 7,
8).
They were asfollows(Fig. 1). R3616 contains a 1-kbp deletion in the coding domain of the Yi34.5 gene. In recombinant HSV-1(F)R, the -Yi34.5 sequences missing from R3616 had been restored. The phenotype and genotype of this virus are those of the wild-type parent, HSV-1(F). In R4009, a stop
codonwas inserted ataminoacid 28 ofthecodingsequence.
R4003(mutantA) containsaninsertion of17codonsinframe with the first codonof the authentic -Yi34.5 genes. In R4002
(mutant B), 28 codons of the
-yi34.5
genewere deleted andreplaced with the initiation codon of the gene specifying glycoproteinH. In R931 (mutant C),39codonsweredeleted in frame after codon 31. In R908 (mutant D),41 codonswere
deleted in frame after codon 72. In R909
(mutant E),
39 codons weredeleted after codon 107. InR939 (mutant F), a sequenceencodingastopcodon in all sixORFswasinsertedat codon 11 after the last three amino acid repeats. All of themutations
introduced
werepresent inbothcopies
of the-y34.5
gene.
Plasmids. Plasmid pMK, containing the metallothionein
genepromoterfusedtothe HSV-1
thymidine
kinase gene(tk),
was the generous giftof R. Palmitter. TheEcoRI-BglII
frag-ment of pMK, containing only the promoter sequence, was
joinedtothe-Yi34.5leader sequenceattheBamHIsite 100
bp
upstreamof the ATG initiationcodon,nearthe
transcription
initiation site(5). Thechimericfragmentwasthen subcloned
intovectorpSV2-neo between EcoRI andBamHI restriction sites asshown inFig. 1C.
[35S]methionine
labeling of infected-cell extracts.The pro-cedure used forlabelingofinfected-cellproteins
with[35S]me-A
HSU-I
IF)
B R3616
HSU- I
(F)R
R4009R
B
C
a b' Ut e
b ULb a.g. C. us
DI-
o7e
s1Y34.5
0--t- A
stip Tag
n 1'A 1A
D 0 1-A1 11
0 0-ls-A 11I
E - i---AIi s I
F 0
~~StopN
F P Dii
l~~~~~~~ii
c
Rnwar
E KBe 9g/U
Pmetallothionein -T134.5-
pSU2-neo
uectorFIG. 1. (A) Schematic diagram of the HSV-1 genome and the location ofthe
yi34.5
geneand itscodingregion. -y34.5 ispresentintwocopies ofthe viral genome:inthejunction between theLandS components(not shown) andattheterminus of theLcomponent, as showninthediagram (5). ab andb'a' representtheinvertedrepeats flanking the uniquesequencesof theLcomponent(UL),whereas a c' andca representthe invertedrepeatsflankingtheS component(Us) (11, 19, 20). The -y34.5 promoter is contained within the terminal 500-bpa repeat (5). Theverticalrectangles represent 20-bp direct-repeat(DR1)sequencesof thea sequence(16).Thesolidbarandthe arrowindicate thecoding regionand thedirection oftranscription of -y34.5.(B) Schematic representation of the deletions andstopcodons within thecodingsequenceofthe
Yi34.5
gene.The constructionand properties of thesemutantshavebeendescribed previously (4,7,8). The triangle indicates the deletion domain. Stop indicates the six-frame terminationcodons insertedat the BstEII (R4009 virus) andDraIIl
(R939 virus) sites. Tag represents a 17-amino-acid codoninsertionatthemethionine translation initiation codon. (C) Plasmid constructofthepMet-y,34.5gene.The
-Yi34.5
sequencewasfusedto thepromoter ofthemousemetallothioneingeneinapSV2-neovector.TATAindicatestheproximal location of transcription initiation. The solidbarindicatesthecodingsequenceof the-y,34.5gene.N,Be, S,St, E, K,Bg, andBareabbreviationsforthe restrictionenzymecleavage sites NcoI, BstEII, Sacl, StuI, EcoRI,
KpnI,
BglII,
andBamHI,
respectively.thionine has already been described
(7).
Briefly, replicatecultureswere either mock infected or
exposed
to the virusesdescribed in Results for 1 h at 37°C. The inoculawerethen
removed, and the cells were incubated at
37°C
in medium199V
consisting of mixture 199(Sigma
Chemical Co., St.Louis, Mo.)
supplemented
with 1% calfserum. At 12 h afterinfection,thecellswereoverlaidwith 1 ml of medium
lacking
methioninebut
supplemented
with 50,uCi
of[35S]methionine
(specific
activity,>1,000
Ci/mmol;
1Ci=37GBq;
Amersham,
ArlingtonHeights, Ill.)for 1 h. Thecellswerethen
harvested,
solubilized,
subjected
toelectrophoresis
indenaturing
poly-acrylamide gels,
transferred to a nitrocellulosesheet,
andsubjected
toautoradiography
aspreviously
described(7).
RNAextractionandNorthern
(RNA) blotting.
RNAswereextractedfrom cells thatwereeither mockinfectedorinfected
withwild-typevirus
HSV-1(F)
ormutant R4009at 13 h after infection byprocedures using
guanidinium thiocyanate
(18).
Poly(A)+
RNAs which representapproximately
5% of totalRNAswere
purified
with akit(PolyATract
mRNA isolationVOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.319.560.75.284.2]8306 CHOU ET AL.
system;Promega, Madison,
Wis.).
Approximately7to8 ,ug of poly(A)+ RNA was then electrophoretically separated on a 1.5%agarose-formaldehyde gel,transferredto anitrocellulose sheet,probed withanantisense-yi34.5
RNAprobesynthesized invitro with anSP6-directed RNAtranscriptionkit(Promega),andsubjected toautoradiography.
Immunoblotting of viral proteins with antibodies. Proteins incell lysateswere electrophoretically separatedin
polyacryl-amidegels cross-linkedwith
N,N'-diallyltartardiamide,
trans-ferredelectrically onto nitrocellulose paper, and reacted with rabbit polyclonal antibody R4 made against the Ala-Thr-Pro-10 repeats of theyi34.5
protein (1).RESULTS
Theability of
-y,34.5
deletionmutantR3616 and stopcodon mutantR4009
toinduce prematureshutoff of protein synthe-sis is cell line specific. Previously we have shown that all proteinsynthesis, both cellular andviral, is prematurelyshut off in human neuroblastoma cells(SK-N-SH)
infected withHSV-1(F)mutantR3616, in which 1 kb of thecodingdomains of bothcopiesof the
-y34.5
genehasbeendeleted,orwith the R4009 mutant, inwhichthe-Yi34.5 proteinhas been truncatedbyastopcodon insertion atamino acid28
(7).
Aremarkable observation whichwereporthere is that in HFFcells,
shutoff ofproteinsynthesiswasobservedafterinfectionwith deletion mutantR3616 butnotafter infection with stop codonmutant R4009. In this series ofexperiments,SK-N-SH, Vero,and HFF cells were infected at equal multiplicities, as described in Materials and Methods, with HSV-1(F), R3616, R4009, andHSV-1(F)R. The infected cells were labeled for 1 h with
[35S]methionineat12 hafter infection and then harvested and processedasdescribed in Materials and Methods. As shown in
Fig. 2, protein synthesis was detected in Vero cells infected withall of the strains tested, in HFFinfected with all of the strains except deletion mutant R3616, and in SK-N-SH cells
infected with thewild-typeandrepairedviruses butnotincells infected witheitherthe R4009orR3616mutantvirus.
The temporal patterns of premature shutoff of protein synthesis by deletion mutant R3616 and stop codon mutant R4009 in neuroblastoma cells are virtually identical. To determine whether the discordance observed in HFF could be a reflection of the temporal pattern of shutoff of protein synthesis, we selected SK-N-SH cells in which shutoff of protein synthesis follows infection with either virus. In this series ofexperiments, we examined the temporal pattern of premature shutoff of protein synthesis in neuroblastoma cells either mock infected or infected with wild-type virus
HSV-1(F), deletionvirusR3616, and stop codon virus R4009 at 2-h intervals from 5 to 20 h after infection. As shown in Fig. 3, premature shutoff of protein synthesis in cells infected with
deletion mutant R3616 or stop codon mutant R4009 was at least partial at 5 h after infection and complete by 11 h after infection.Inthis series of experiments, no significant difference between the phenotypes of the deletion and stop codon mutantswasapparent.
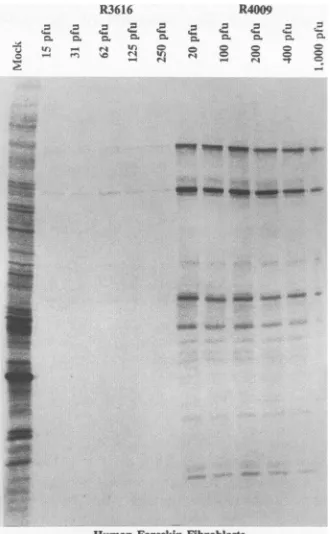
The discordance between the stop codon and the deletion mutants in HFF is not due to differences in multiplicity of infection. In a series of experiments, replicate HFF cultures wereexposed to R3616 at 15 to 250 PFU per cell or to R4009 at 20 to 1,000 PFU per cell. The cultures were processed as described above. Thecells were labeled for 1 h at 12 h after
infection and then harvested and processed as described in Materials and Methods. The results shown in Fig. 4 indicate that the effect of R4009 onviral protein synthesis was
inde-pendentof themultiplicity of infection. We conclude that the
A
j
vn ff X v
- .Da4_4 2
u4
w
0XW c: a: X = lCP
-W 5
_1I
:
-
...-A*
U,-4
W-r ::> 1:04-x^t
0
SK-N-SH
l 25
-i 32
[image:3.612.318.552.72.275.2]HFF Vero
FIG. 2. Autoradiographicimage ofelectrophoretically separated, [35S]methionine-labeledproteins fromlysatesof cells infected withthe
wild-typevirusor mutantviruses. The cellswereinfected with 10PFU percelland labeledat12 h after infection for 1 h. Theprocedures for infection,[35S]methionine labeling, preparation of celllysates, electro-phoresis in N,N'-diallyltartardiamide-polyacrylamide gels, electrical transfer ofproteinstonitrocellulosesheets, andautoradiographyhave been previously described (7). Infected-cell protein (ICP) numbers were assigned in accordance with Honess and Roizman (12) and Morse etal. (17). Mock, mock infection; A, actin.
differences in the premature shutoff ofprotein synthesis seen in the infected HFF cultures described above are intrinsic properties of thecells; i.e., theresultsdescribedabovearedue solely to the fact that SK-N-SH cells are affected by the presence of the stop codon in the -Yi34.5 genes of R4009 whereas HFF are not.
Protein synthesis activities in cells infected with
y,34.5
partial deletion viruses and C terminus stop codon mutant virus F. Elsewhere we reported that the carboxyl terminus domainof the-Yi34.5protein is requiredtopreclude premature shutoff ofprotein synthesis in SK-N-SH cells(8).Inthat study, wetestedaseries ofmutantsgeneratedbyin-frame deletions or insertion of a stop codon 11 codons after the Ala-Thr-Pro-10 repeats(Fig. 1).Inlight of the results described above, itwasofinterest to determine thephenotype-ofthese mutants in HFF cultures. In this series of experiments, replicate SK-N-SH or HFF cultures were infected with 10 PFU ofwild-typeormutantvirus percell. At 12 h after infection, the cells were labeled for 1 h with [35S]methionine and then harvested and processed as described in Materials and Meth-ods. The results (Fig. 5) were as follows. (i) As previously reported, there wasnopremature shutoff of protein synthesis in Vero cells infected with wild-type and mutant viruses.
Similarly, therewasnoprematureshutoff of protein synthesis inSK-N-SHcells infected with the wild-type virus or mutant A, B, C, D, or E. Premature shutoff of protein synthesis was observed in cells infected with mutant R3616 or F.(ii) In HFF
cultures, prematureshutoff of protein synthesis was observed
only following infection of the cells with deletion mutant R3616.
Once again, HFF andSK-N-SH culturesyieldeddiscordant results. Inthis instance again,SK-N-SH cells were affected by the presence of the stop codon in the domain encoding the J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
VOL68,1994
~~~~~~DELETIONS
AND STOP CODONS IN HSV-yi34.5
GENE 83075k VERO 5h S K 7k SK 9k 5K SK 13k SE 15k SK 18k 5K 20k VERO 20k 5K
~~~~~~~
*w
W4
5
11
19 25 32
1-...
O.-iws W ...
IM"
".7.
40M
i;...
itP"
[image:4.612.70.568.76.262.2]'W
FIG. 3.
Temporal
pattern
oflabeling
ofproteinsin Vero andSK-N-SH cells either mock infectedorinfectedwith thewild-type
virusormutant R3616orR4009.The cellswerelabeled with[35S]methionine
for 1hat5,7,9, 11, 13,15, 18,and 20 h afterinfection. Themajor proteinbands formedby lysatesofinfected cells areidentifiedbythe infected-cellproteinnumberson thefight.
carboxyl-terminal
80amino acids of the-yi34.5
gene,whereas HFFwere not.Deletion and
stop
codonmutants donotexpress detectablequantities
of-y3.
proteins
or of truncated mRNA. In thefirst oftwo series of
experiments,
HFF,
SK-N-SH,
andVero cellswere infected withHSV-1(F),
R3616,
orR4009 virus. At 20 h afterinfection,
the cellswere harvested andassayed
forR3616
0.
R4009
0. 0. M.
-'4
-"M* V am"e ".
Human ForeskinFibroblasts
FIG. 4. Autoradiographic image of [35S]methionine-labeled
pro-teins in HFFcells either mock infectedorinfected with virus R3616or
R4009atincreasing multiplicitiesof infection. The number of PFU of virus per cell in each infection is shown.
the presence of
-yi34.5
protein
as described in Materials and Methods. The results shown inFig.
6 indicate that cells infected with R3616orR4009donotmakedetectableamountsof
protein
reactive with the rabbitpolyclonal
antibody
totheAla-Thr-Pro-10 sequencecharacterized in a
preceding
publi-cation. We havepreviously
shownthatthisantibody
reactswitha
variety
of truncated-yi34.5
proteins.
In the second series of
experiments,
RNA was extractedfrom
HFF,
SK-N-SH,
or Vero cells infected with the sameviruses. The RNAwas
electrophoretically
processed
andhy-bridizedtoa
-yi34.5
DNAprobe
asdescribed in Materials and Methods.Figure
7shows that cells infected with the deletion mutantorthestopcodonmutantsaccumulatedtwospecies
of RNA reactive with a-y134.5-specific
probe.
One RNA(A
band),
with apredicted length
of 5kb,
consists of an RNA which contains both-yi34.5
and atO sequences. The second RNA(B band)
is 1.3 kblong
andcontains-yi34.5
sequencesonly.
Both RNAs have beenpreviously
described(5).
The results of these
experiments
revealed no detectablequantities
of either a truncated RNA that could express atruncated form of
-yi34.5
protein
orthe presence ofatruncated-yi34.5
protein
that can contribute to thepartial
protein
synthesis
in R4009stop
codonmutantvirus-infected HFFcells. The SK-N-SH cellline,
which iscapable
ofexpressing
the"y,34.5
protein, complements
the'y,34.5S
phenotype
of the R3616 deletion mutant. In the first of two series ofexperi-ments, SK-N-SH cells were transfected with
pRB321O
andplated
in the presence of G418(400 ~xg/ml
ofmedium).
Theresulting
G418-resistant cloneswere subcloned andtested for the presence of-yi34.5
protein
(Fig. 8)
and theability
ofthe-yi34.5
gene,by
cadmiumsulfate(50 pLM)
induction,
to restoreprotein synthesis
incellsinfected with the R3616 virus(Fig. 9).
Three clonal cell lines
carrying
the-yi34.5
gene and a linecarrying
only
the G418 resistance marker were selected forthese studies. Asshown in
Fig.
8,clonal linesPmet--y134.5-11
and
Pmet--y134.5-1
expressed
the-yi34.5
protein
atrelatively
high
levels after cadmium induction and atlower levels in the absence of cadmium. ThePmet--y134.5-2
cell lineexpressed
barely
detectable levels of the-y,34.5
protein
inthepresence ofcadmium,
andexpressed
no detectableyi34.5
protein
inun-induced cells. VOL.
68,
1994-:4
k
.uo;.W,#
11"ko:
a ."Wft
z
;,6,,
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.105.270.418.685.2]8308 CHOU ET AL.
oC
V
O
_~~
A B C D E F ICP
A'z.*
z *ii *5i5
~~~~~ 4~~~~~~~~11
i 9U1: i 25
I ~~~~~~~~~~~32
dA
B C D E F
X4
Q ;> Z
>S m P A B C D E F
ICP
s- 11
19
35
32
A
[image:5.612.60.546.75.299.2]SK-N-SH HFF
Vero
FIG. 5. Autoradiographic imageof[35S]methionine-labeled proteinsinSK-N-SHcells, HFF,and Vero cellseithermockinfectedorinfected
with wild-typevirusHSV-1(F) ordeletionmutantR3616,A, B, C, D, E,orF. ICP,infected-cellprotein; A,actin.
In thesecondseries ofexperiments,parentand clonal lines
wereinfected with the wildtypeand the deletionmutantinthe
presence orabsenceof cadmium. Thekey findings, shown in
Fig. 9,were that proteinsynthesis wasshut offby the R3616
virus in the parent cell line and in cells carrying the G418 resistancemarker. Proteinsynthesiswaspresent atnearlythe wild-type level when the R3616 virus infected the
Pmet-yi34.5-11
andPmet--y,34.5-1
clonal cell lines in the absence of cadmium andwasenhanced in both cell lineswhencadmiumwasaddedtothemedium. Thesignificant findingisthat in the Pmet-y134.5-2 clonal cellline,which didnotexpresstheYi34.5 proteinat adetectable level in the absence of cadmium (Fig.
8), protein synthesis was shut off by the R3616 virus to the
same levelas in control cell lines. This premature shutoffof protein synthesisin thePmet-y134.5-2 clonal cell line infected
LO)
crt
ClIL
tD
cnto C
0 0°qetCV C) 0
E E c
with R3616 was precluded by addition of cadmium to the medium.
We conclude the following. (i) Induction of the activity whichprecludedtheshutoff ofprotein synthesiswastiedtothe inductionofthe metallothionein promoterbycadmium. Con-sistent with these results, the Yi34.5 protein in expressor cell lines complemented the deletion in the -y34.5 gene of the R3616virus ininfection toprecludethe premature shutoff of protein synthesis. (ii) The amounts of the -y34.5 protein
B
A
B
SK-N-SH Vero HFF
FIG. 6. Photographic image of -Yi34.5 proteins in infected-cell
extracts,visualizedby stainingwith polyclonal antibodyR4.SK-N-SH cells, HFF,and Verocellswereeithermockinfectedorinfected with
R3616, R4009,orwild-type virusHSV-1(F) for 24 h. The procedures
used forharvesting ofextracts,electrophoresis, transfer, and immuno-blottingaredescribed inMaterials and Methods.
HFF SK-N-SH Vero
FIG. 7.
-y,34.5-specific
RNAtranscripts in cells infected with the wild-type virus or the R4009 mutant virus. Poly(A)+ RNAs wereextracted from HFF, SK-N-SH cells, and Vero cells either mock infectedorinfectedwith theHSV-1(F)orR4009 virus and probed with anantisense RNAprobe specificforYi34.5.A,5-kb transcriptinitiated
fromtheasequencepromoterof-Yi34.5 and 3' coterminate with that
ofaOtranscripts; B, 1.3-kb transcriptof the -y34.5 codingsequence
only.Bothtranscriptshave beenreported previously (5).
ICP
5
11
19
25
32 J. VIROL.
5 a*VW,
i,; ...
:40 "WA
-ii ::: .".. .mzi
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.347.515.463.652.2] [image:5.612.60.293.518.670.2]DELETIONS AND STOP CODONS IN HSV y,34.5 GENE 8309
Vector G418 Prnet-y34.5-11 Pmet-y34.5-2 Pmet-y34.5-1
Cd++ + - + - + - +
-Cd + ^ c c _ KIe Y
° 'O, 0.0
.-R4anti-serum
FIG. 8. Photographic image of
y,34.5
proteins expressed by clonal lines ofSK-N-SH cells. Vector G418 is the designation of a control cell line selected in thepresence of G418.Pmet--y,34.5-11,
Pmet--y,34.5-2,
andPmet-y134.5-1 were selected fromSK-N-SH cultures transfected with the plasmids shown in Fig. 1C and plated in medium containing G418.The clonal cell lineswereuntreated (-) orexposed to 50 ,uM cadmium sulfate (+) during infection. The procedures used for preparation of cell lysates, electrophoresis in N,N'-diallyltartardia-mide-polyacrylamide gels, electrical transfer of proteins to nitrocellu-losesheets, and reaction with antibody to the-y34.5protein with rabbit polyclonal antibody R4 were as previously described. 0, position ofyi34-5 protein.
required to preclude shutoff of protein synthesis were low. For
example, the amounts of the
-y34.5
protein expressed in the uninducedPmet--y,34.5-11
andPmet--y,34.5-1
clonal cell lines appear tobesufficient to preclude premature shutoff of proteinsynthesiswhereas the amounts expressed in the
Pmet--y,34.5-2
clonal line were notsufficient.Inthe presence of cadmium, the smallamountmade in thePmet-y134.5-2 cell line was sufficient to complementthe-y34.5-
virus.DISCUSSION
Thesalient features of this report are as follows. (i) Cultures of two human cell lines, SK-N-SH neuroblastoma cells and HFF, responded identically to infection with
-Yi34.5
deletion mutantR3616butin very different ways to infection with stop codonmutants R4009 and F. Specifically, SK-N-SH cells and HFFinfectedwith-y,34.5
deletion mutant R3616 exhibited the samephenotype, i.e.,premature, total shutoff ofproteinsyn-thesis. Whereas SK-N-SH cells infected with stop codon mutants R4009 and R939 (mutant F) exhibited the same
phenotypeas thatobserved afterinfection with deletion mu-tantR3616, HFFcells infected with the stop codon mutants exhibitedawild-type phenotype; i.e.,protein synthesiswas not
prematurely shut off. (ii) The -y34.5 protein expressed in
SK-N-SH cellscomplementedthe R3616 deletionmutantand
precludedprematureshutoff ofproteinsynthesis.Theamounts of
-y,34.5
protein required for this function appeared to be small.Relevant to the results presented in this report are the
following findings.
(i)
In our studies, R3616 and R4009be-havedidenticallyinbothVero andSK-N-SH cellsin culture.
Furthermore,asshown inthisreport,the
temporal
patternsof prematureshutoffofproteinsynthesisinSK-N-SH cellsbythe twoviruseswerevirtuallyidentical. Both R3616 and R4009arehighly attenuatedin mice infectedby the intracerebralroute
(4, 21).
Specifically,
the[R4009
PFU/LD50]/[HSV-1(F)
PFU/
LD5O]
ratio,whereLD50is the50% lethaldose,was>105
(4).
Incontrast to ourresults,Bolovan etal.
(3) reported
thatan HSV-1 strain 17 mutant, 1771, in which a stop codon was insertedatthesamepositionasin theR4009 mutant,i.e.,
after codon 27 of the-Yi34.5
gene, exhibited a(1771
PFU/LD50)/
(HSV-1strain 17
PFU/LD5O)
ratioof 25 with thesame routeofs
--r ti E:
^lw ...., t _ * w
.w.
-'5Z
* -S ,jge
w , w .*. *a
F .! |
2
t.ss;
jP
::
+ §
.4: w
.. ],
,. s:W
*:.X
r:S
311F'
*^F
ICP
19
_
25
;m:: 32
Fw 9A
w'.
I'arentCcll Vector(;418 Pme,- 4.4-1l P Y34.5-2 P.,,y34.n-i
Line CellLine Cell Line CellLine Cell Line
FIG. 9. Autoradiographic images of electrophoresis-separated,
[35S]methionine-labeledproteinsinSK-N-SHclonal cells infected with viruses. Thevarious SK-N-SH clonal cell lines shown in Fig. 8 were infected with virusHSV-1(F) or R3616 and were either untreated (-) ortreated(+) with 50 ,uM cadmium sulfate during infection prior to labelingwith [35S]methionine. Mock, mock infection. ICP, infected-cellprotein. A, actin.
inoculation,i.e.,several orders of magnitude different from the results reported in our studies. Their data are difficult to interpret, particularly in light of the studies by McKie et al. (15), who inserted a stop codon after codon 9 of both copies of the
-yi34.5
gene(mutant termA)andreported that the(termAPFU/LD50)/(HSV-1
strain 17PFU/LD50)
ratio was>105,
i.e.,similartothatreported byChouetal. (4).Preliminary studies indicate that R939(mutantF)is alsohighly attenuated follow-ing intracerebral inoculation (12a).
(ii)Theexperimentsreported here indicate that the
-Yi34.5
geneproductisessential for full viral geneexpressionin HFF. This conclusion is based on the observation that the
-Yi34.5
gene deletion mutant caused premature shutoff of protein synthesis.Itfollows that the failure of the stop codonmutants toinduce premature shutoff ofproteinsynthesisindicates that therequirement for the-y34.5 proteinwas satisfied in some fashion. The disparity between the replicative cycles of the R3616 and R4009 viruses is not due to some gene dosage effect, inasmuchas we have shown that the phenotypeof the mutants in HFF cells is independent of the multiplicity of infection.
Theformalpossibilitieswhichcould accountfor the results
observed are the following.
Hypothesis
1 is thatprotein
synthesis is initiated downstream from the stop codons.
For-mally,we cannotexclude this
hypothesis.
However,theyi34.5
genecontainsasinglemethionine codon.
Although
alternative(non-methionine)
initiations abound, toexplain
theobserva-tion thatrecombinant R939
(mutant
F),
withthe stop codon downstream from thetripletrepeatatcodon205,
also failsto induce prematureshutoff ofprotein
synthesis
in HFFcells,
it is necessarytopostulate eitherafamily
oftruncatedRNAsandcorresponding shorter -y34.5
proteins
or that the initiator codon is 3'tothe stop codoninR939(mutant
F),
which hasa codingcapacityofonly
60amino acids. As shown inFig.
6and7,wehavenot detectedasmaller,truncated
protein
inlysates
I I.- I I
'l-M
4'-.s
VOL.68, 1994
I I* I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.325.563.76.292.2] [image:6.612.63.302.76.180.2]8310 CHOU ET AL.
ofR4009-infected cells or shorter truncated RNAspeciesthat could giverise to a smallerproteintofavor thishypothesis.We have, on occasion, observedfaster-migrating proteins reactive with the anti-y134.5 antibody in cells infected with the wild-type virus but not in cells infected with R4009. The faster-migrating bands reacting with the antibody to the
-Yi34.5
protein shown in Fig. 6 were not reproducibly found in all of theimmunoblots, and those shown in Fig. 6reflect proteolytic digestion rather than polypeptides obtained by alternative codon usage for translation of the protein.
Hypothesis 2 is that the stop codons are contained within introns. The stop codon in the R4009 virus is within a sequence
surrounded by potential splice donor and acceptor sites. To explain the differential growth of the R939 mutant in SK-N-SH and HFF cells, we would have to postulate that both stop codons are within introns and are spliced out and that,
furthermore, the splicing of RNA is human cell type (fibro-blasts versus neuroblastoma cells) specific. We cannot formally exclude this hypothesis, but testing of this hypothesis is ren-deredtremendously more complex by our inability to verify the existence of truncated gene products in HFF cells.
Hypothesis 3 is that the stop codons are suppressed at a low level. Stop codon suppression by a tRNA suppressor is a
well-documented event, and suppression can vary from 1 to 20% in some cells. This hypothesis differs from the preceding two in that it does not require the existence of truncated RNA orproteins, and furthermore, such events may well be tissue specific. Consonant with this hypothesis are two observations. First, stop codon mutant R4009 and deletion mutant R3616 were equally avirulent in mice inoculated intracerebrally and little or no virus could be detected in central nervous system tissues in the days following inoculation. However, both of these viruses were detected in vaginal tissues and the stop codon mutant was present at a titer greater than that of the deletion mutant after vaginal inoculation (21). These observa-tions suggest that both mutants have a reduced capacity to multiply in the central nervous system, a conclusion that is consistent with the results of studies on the human neuroblas-toma cell line. However, both viruses have a residual capacity tomultiply in tissues peripheral to the central nervous system, andmoreover, the stop codon mutant is better able to multiply; these findings are also consistent with the behavior of the two viruses in HFF. The second relevant observation stems from results presented in Fig. 8 and 9 showing that cadmium induction of the
-y34.5
gene in SK-N-SH cells constitutively expressing small amounts of theyi34.5
protein was not neces-sary to complement the-Yi34.5-
mutant. The cell line which required induction to complement the-Yi34.5-
mutant pro-duced no detectableYi34.5
prior to induction and barely detectable amounts afterward. These observations suggest that traceamounts of-Yi34.5
proteins are sufficient to preclude the shutoff of protein synthesis induced by a deletion virus in the absence of theYi34.5
protein.Lastly, two series of experiments indicate that both the in vivoattenuated phenotypeand the premature shutoff of pro-tein synthesis in vitro are associated with mutations in the
yi34.5
gene and bothreflect defects introduced into that gene and not into ORF-P, which is located exactly antisense to the-Yi34.5
gene.First,
McKieetal.(15)
tooknoteof the existenceof the transcript reported by Bohenzky et al. (2) and by Yeh and Schaffer (22) and constructed its
Yi34.5
gene stop codon mutant in such a manner as to preclude mutagenesis of ORF-P. Theobservation that the mutant produced by McKie etal.(15) is nearly identical to R3616 and R4009 in the murine model indicates that the properties associated with neuroviru-lence are indeed due to the-yi34.5
gene. The studies presentedhere indicate that induction of the metallothionein promoter-driven
Yi34.5
gene by cadmium resulted in full or enhanced complementation of the-Yi34.5-
virus and support the argu-ment that premature shutoff of protein synthesis in cells is indeed due to absence of the-Yi34.5
gene.The scenario that bestfits theavailable data is that initiation of viral DNAsynthesis triggers in infected cellsahost response to infection similar to those triggered in murine myelogeous leukemia cells induced to differentiate byinterleukin 6 orcells subjected to DNA damage. According tothishypothesis, HSV or its ancestor co-opted a domain of a cellular GADD gene and tacked it on at the carboxyl terminus of a forbear of the
-Yi34.5
gene topreclude
cells fromshutting
off allprotein
synthesis
and thus interfere with viralreplication.
ACKNOWLEDGMENTS
We thank George Kemble for bringing to our attention the fact that R3616 does not multiply in human foreskin fibroblasts.
These studies were aided by grants from the National Cancer Institute (CA47451), the National Institute of Allergy and Infectious Diseases (AI124009), and the U.S. Public Health Service and by an unrestricted grant from the Bristol-Myers Squibb Program in Infec-tious Diseases.
REFERENCES
1. Ackermann, M., J. Chou, M. Sarmiento, R. A. Lerner, and B. Roizman. 1986. Identification by antibody to a synthetic peptide of a protein specified by a diploid gene located in the terminal repeats of the L component of herpes simplex virus genome. J. Virol.58:843-850.
2. Bohenzky, R. A., A. G. Papavassiliou,
I.
H. Gelman, and S. Silverstein. 1993. Identification of a promoter mapping within the reiterated sequences that flank the herpes simplex virus type 1 UL region. J. Virol. 67:632-642.3. Bolovan, C. A., N. M. Sawtell, and R. L. Thompson. 1994.
ICP34.5
mutants of herpes simplex virus type 1 strain17syn+
are attenu-ated for neurovirulence in mice and for replication in confluent primary mouse embryo cell cultures. J. Virol. 68:48-55. 4. Chou, J., E. R. Kern, R. J. Whitley, and B. Roizman. 1990.Mapping of herpes simplex virus-1 neurovirulence to -y34.5, a gene nonessential for growth in culture. Science 250:1262-1266. 5. Chou, J., and B. Roizman. 1986. The terminal a sequence of the
herpes simplex virus genome contains the promoter of a gene located in the repeat sequences of the L component. J. Virol. 57: 629-637.
6. Chou, J., and B. Roizman. 1990. The herpes simplex virus 1 gene for ICP34.5, which maps in inverted repeats, is conserved in several limited-passage isolates but not in strain17syn+. J. Virol. 64:1014-1020.
7. Chou, J., and B. Roizman. 1992. The
-Yi34.5
gene of herpes simplex virus 1 precludes neuroblastoma cells from triggering total shutoff of protein synthesis characteristic of programed cell death in neuronal cells. Proc. Natl. Acad. Sci. USA 89:3266-3270. 8. Chou, J., and B. Roizman. 1994. The herpes simplex virus 1-y34.5
gene function which blocks the host response to infection maps in the homologous domain of the genes expressed during growth arrest and DNA damage. Proc.Natl. Acad. Sci. USA 91:5247-5251. 9. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1967. Characteriza-tion of herpes simplex virus strains differing in their effects on social behavior of infected cells. J. Gen. Virol. 2:357-364. 10. Fornace, A. J., J. Jackman, M. C. Hollander, B.
Hoffman-Liebermann, and D. A. Liebermann. 1992. Genotoxic-stress-re-sponse genes and growth-arrest genes. Ann. N. Y. Acad. Sci. 663: 139-153.
11. Hayward, G. S., R. J. Jacob, S. C. Wadsworth, and B. Roizman. 1975. Anatomy of herpes simplex virus DNA: evidence for four populations of molecules that differ in the relative orientations of their long and short segments. Proc. Natl. Acad. Sci. USA 72: 4243-4247.
12. Honess,R W., and B. Roizman. 1975. Regulation of herpesvirus macromolecular synthesis: sequential transition of polypeptide J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
DELETIONS AND STOP CODONS IN HSV -y,34.5 GENE 8311
synthesis requires functional viral polypeptides. Proc. Natl. Acad. Sci. USA 72:1276-1280.
12a.Kern, E., R. J. Whitley, J. Chou, and B. Roizman. Unpublished data.
13. Lagunoff, M., and B. Roizman. 1994. Expression of a herpes
simplex virus 1open reading frame antisensetotheyi34.5 gene
and transcribed by an RNA 3' coterminal with the unspliced latency-associated transcript. J. Virol. 68:6021-6028.
14. Lord, K. A., B. Hoffman-Liebermann, and D. A. Liebermann. 1990. Sequence ofMyD116 cDNA:anovelmyeloiddifferentiation
primary response gene induced by IL6. Nucleic Acids Res. 18: 2823.
15. McKie, E. A., R. G. Hope, S. M. Brown, andA. R.MacLean. 1994. Characterization of the herpes simplex virus type 1 strain 17+ neurovirulencegeneRL1and itsexpression inabacterialsystem.
J. Gen.Virol.75:733-741.
16. Mocarski, E. S., and B. Roizman. 1982. Thestructureand role of the herpessimplex virus DNA termini in inversion, circularization andgeneration of virionDNA. Cell 31:89-97.
17. Morse, L. S., L. Pereira, B. Roizman, and P. A.Schaffer. 1978.
Anatomyof herpes simplex virus (HSV) DNA. X. Mapping of
viralgenesby analysis ofpolypeptides and functions specified by
HSV-1 x HSV-2 recombinants. J. Virol. 26:389-410.
18. Puissant, C., and L.M.Houdebine.1990. An improvement of the single-step method of RNA isolation by acidguanidinium thiocy-anate-phenol-chloroform extraction. BioTechniques 8:148-149. 19. Sheldrick, P., and N. Berthelot. 1975. Invertedrepetitionsinthe
chromosome of herpes simplex virus. Cold SpringHarborSymp. Quant. Biol.39:667-678.
20. Wadsworth, S., R. J. Jacob,and B.Roizman. 1975. Anatomy of herpes simplex virus DNA. II. Size, composition, and
arrange-mentsofinverted terminal repetitions. J. Virol. 15:1487-1497. 21. Whitley, R. J., E. R. Kern, S. Chatterjee, J. Chou, and B.Roizman.
1993. Replication, establishment of latency, and induced
reactiva-tion ofherpes simplex virus -Yi34.5 deletion mutants in rodent
models.J. Clin. Invest.91:2837-2843.
22. Yeh, L., and P. A. Schaffer. 1993. A novel class of transcripts expressed with late kinetics in the absence of ICP4 spans the
junction between the long and short segments of the herpes simplex virustype1genome.J. Virol.67:7373-7382.
VOL.68, 1994
![FIG. 2.wild-typewerebeenphoresisperinfection,transferMorse[35S]methionine-labeled Autoradiographic image of electrophoretically separated, proteins from lysates of cells infected with the virus or mutant viruses](https://thumb-us.123doks.com/thumbv2/123dok_us/1289850.81834/3.612.318.552.72.275/typewerebeenphoresisperinfection-transfermorse-methionine-autoradiographic-electrophoretically-separated-proteins-infected.webp)

![FIG. 9.viruses.orlabelingcellinfected[35S]methionine-labeled treated Autoradiographicimagesofelectrophoresis-separated, proteins in SK-N-SH clonal cells infected with The various SK-N-SH clonal cell lines shown in Fig](https://thumb-us.123doks.com/thumbv2/123dok_us/1289850.81834/6.612.325.563.76.292/viruses-orlabelingcellinfected-methionine-autoradiographicimagesofelectrophoresis-separated-proteins-infected-various.webp)
