Copyright1998 by the Genetics Society of America
Statistical Analysis of Half-Tetrads
Hongyu Zhao* and Terence P. Speed
†*Department of Epidemiology and Public Health, Yale University School of Medicine, New Haven, Connecticut 06520
and†Department of Statistics, University of California, Berkeley, California 94720
Manuscript received January 20, 1998 Accepted for publication May 18, 1998
ABSTRACT
Half-tetrads, where two meiotic products from a single meiosis are recovered together, arise in different forms in a variety of organisms. Closely related to ordered tetrads, half-tetrads yield information on chromatid interference, chiasma interference, and centromere positions. In this article, for different half-tetrad types and different marker configurations, we derive the relations between multilocus half-half-tetrad probabilities and multilocus ordered tetrad probabilities. These relations are used to obtain equality and inequality constraints among multilocus half-tetrad probabilities that are imposed by the assumption of no chromatid interference. We illustrate how to apply these results to study chiasma interference and to map centromeres using multilocus half-tetrad data.
H
ALF-TETRADS, where two meiotic products from heterozygous at the marker. When there is one crossover between centromere and the marker, there is equal a single meiosis are recovered together, arise indifferent forms in a variety of organisms. The first well- chance of producing homozygous and heterozygous half-tetrads. In Figure 2, the mechanism of meiosis II (MII) studied half-tetrad data were attached-X chromosomes
in Drosophila (Beadle andEmerson1935;Welshons nondisjunctions is shown. Given no crossovers between centromere and a heterozygous marker, MII nondis-1955). Half-tetrads were also constructed using
au-tosomes in Drosophila (BaldminandChovnick1967). junction always results in homozygous half-tetrads, whereas a single crossover always results in heterozygous They have been used in the study of many other
organ-half-tetrads. For half-tetrads from MI nondisjunctions, isms, including maize (Rhoades andDempsey 1966),
the two strands were attached to different centromeres potatoes (Mendiburu and Peloquin 1979), leopard
during meiosis, whereas the two strands in half-tetrads frog (Volpe 1970), rainbow trout (Thorgaard et al.
from MII nondisjunctions were attached to the same 1983;Allendorfet al. 1986), salmonid fish (K. R.
John-centromere during meiosis. MI and MII nondisjunction
sonet al. 1987), catfish (Liuet al. 1992), and zebrafish
are not the only mechanisms that are responsible for (S. L. Johnsonet al. 1995). In mammals, half-tetrads
half-tetrads. For example, attached-X chromosomes in can be studied in the form of autosomal trisomies (
Mor-Drosophila are the result of a different mechanism (
Bea-tonet al. 1990;Shermanet al. 1991) and ovarian
terato-dle and Emerson 1935). In this article, we broadly mas (Ottet al. 1976;EppigandEicher1983;
Chakra-classify half-tetrads into two types: type I half-tetrads, in varti and Slaugenhaupt 1987; Chakravarti et al.
which no crossover between centromere and marker 1989;Dekaet al. 1990). However, the material required
always results in heterozygous half-tetrads, and type II of trisomies and teratomas is rare, and the
recombina-half-tetrads, in which no crossover always results in ho-tion pattern in meiosis that generates trisomies and
mozygous half-tetrads. On the basis of this classification, teratomas can differ from that in normal meiosis (
Sher-tetrads from MI nondisjunctions are type I
half-manet al. 1991, 1994;Lambet al. 1996).Cuiet al. (1992)
tetrads, and those from MII nondisjunctions are type II introduced one technique that uses the polymerase
tetrads. Attached-X chromosomes are type I half-chain reaction to analyze the products of meiosis I in
tetrad. Half-tetrads from fish are mostly type II half-tetrads. individual secondary oocytes. This method has been
Autosomal trisomies and ovarian teratomas can be of since used to map genetic markers in mice (Cuiet al.
either type. In addition, ovarian teratomas can result 1992) and cows (Jarrell et al. 1995).
from mechanisms other than MI or MII nondisjunctions Half-tetrads may arise from different mechanisms. In
(Surtiet al. 1990). Throughout this article, we also make Figure 1, we illustrate how meiosis I (MI)
nondisjunc-the assumptions that nondisjunc-the parental origin of nondisjunc-the half-tions lead to half-tetrads. It is easy to see that when there
tetrads is known and that phases are known in parents. is no crossover between centromere and a heterozygous
These assumptions are usually true for experimental marker, MI nondisjunction results in half-tetrads being
organisms, although human half-tetrad data are more complex and may not satisfy these assumptions. For either type I or type II half-tetrads, a further distinction Corresponding author: Hongyu Zhao, Department of Epidemiology
may be made when two or more markers are studied:
and Public Health, Yale University School of Medicine, 60 College
Street, New Haven, CT 06520. E-mail: [email protected] haplotype information can be either available (attached-X
Figure1.—Diagram illus-trating nondisjunction dur-ing the first meiotic division. No crossover between the centromere and the marker always results in heterozy-gous half-tetrads. One cross-over between the centro-mere and the marker has equal chance of resulting in homozygous half-tetrads and heterozygous half-tetrads.
chromosomes in Drosophila) or unavailable. The two shons 1955); (2) when chromatid interference is ab-sent, chiasma interference can be detected with two loci types of tetrads are called type Ia and type IIa
half-tetrads when haplotype information is available and type and may be detected with just one locus if the locus is sufficiently far from the centromere; and (3) the posi-Ib and type Iposi-Ib half-tetrads when such information is
not available. tion of the centromere can be mapped.
Most studies on half-tetrads (Welshons 1955; Cote As with ordered and unordered tetrads, half-tetrads
are very valuable in studying crossovers during meiosis andEdwards1975;Ottet al. 1976;Chakravartiand Slaugenhaupt1987) used only three loci for the detec-because (1) chromatid interference and chiasma
inter-ference can be distinguished with half-tetrads (Wel- tion of chromatid interference and one locus for the
mapping of centromeres. In the context of chromatid (Griffiths et al. 1996; Zhao and Speed 1998) using half-tetrad data.
interference, Zhaoet al. (1995a) andZhaoandSpeed
(1998) derived a set of linear equality and inequality We use the following notations in this article. Markers are denoted by script letters. For example, we use A constraints on the multilocus probabilities of unordered
and ordered tetrad patterns under the assumption of andBto denote markers. Alleles are denoted by italic letters. For example, A and a denote two alleles of no chromatid interference (NCI). These constraints can
be used to test the assumption of NCI and to order markerA. We use [X, Y, Z, W ] to denote the observed marker configuration for an ordered tetrad, where X markers. Risch and Lange (1983) and Zhao et al.
(1995b) fitted chiasma interference models to multilo- and Y are attached to one centromere and Z and W are attached to the other centromere. For example, [AB, cus unordered tetrad data. For half-tetrad data analysis,
Chakravarti et al. (1989) proposed two approaches Ab, aB, ab] represents an ordered tetrad with two strands carrying AB and Ab attached to one centromere and for multilocus analysis. One was to assume that there
are at most three chiasmata across the region under ge- two strands carrying aB and ab attached to the other centromere. For type Ia and IIa half-tetrads, two strands netic study. The other was to treat the proximal marker
as a pseudocentromere relative to the distal marker. are separated by a / . For half-tetrads from MII nondis-junctions, these two strands were attached to the same Because the first approach does not apply to tetrads with
more than three chiasmata and the second approach centromere during meiosis. For half-tetrads from MI nondisjunctions, these two strands were attached to dif-applies only in the absence of chiasma interference,
neither is completely satisfactory. Daet al. (1995) pre- ferent centromeres. For type Ib and IIb half-tetrads, genotypes at each marker are combined and separated sented an approach to analyzing two markers under
the assumptions of NCI and no chiasma interference. by ; in parentheses. For example, aB/Ab represents a half-tetrad with one strand bearing aB and the other Assuming complete chiasma interference, Tavoletti
et al. (1996) proposed a maximum likelihood method. strand bearing Ab, whereas (Aa; Bb) represents a
half-tetrad with genotype Aa at A and genotype Bb at B,
Lamb et al. (1997) followed Weinstein’s (1936)
ap-proach to inferring joint chiasma probabilities at the without knowing whether A and B are on the same strand or A and b are on the same strand.
four-strand stage. There is no assumption on the chi-asma process in this approach except that there are at most two chiasmata in each marker interval.
METHODS
In this article, we assume that when strands attached
to different centromeres during meiosis form a half- No chromatid interference (one marker):With one marker, haplotype information is irrelevant. We need tetrad, each of the two strands attached to the same
centromere has equal chance of being in the half-tetrad, consider only two types: type I and type II half-tetrads.
Type II half-tetrads: For a heterozygous markerAwith
and when strands attached to the same centromere
dur-ing meiosis form a half-tetrad, the two pairs have equal alleles A and a, there are three observed patterns: AA,
Aa, and aa. For MII nondisjunction, patterns AA and
chance of being in the half-tetrad. Under this
assump-tion, four nonsister chromatid pairs have the same aa are derived from ordered tetrads having first division
segregation (FDS) pattern [A, A, a, a]. Pattern Aa is chance of being observed in a half-tetrad from MI
non-disjunctions. For MII nondisjunctions, two sister chro- derived from ordered tetrads having second division segregation (SDS) pattern [A, a, A, a]. Under RRA, matid pairs have the same chance of being recovered.
This assumption is abbreviated as RRA (random recov- tetrads having FDS pattern should give rise to AA and
aa with the same probability. Tetrads having SDS pattern
ering assumption) in the following discussion.
Under RRA and the assumption of NCI, we derive can only give rise to Aa. Therefore, P(AA)5 P(aa) 5
P(FDS)/2 and P(Aa)5 P(SDS).
multilocus half-tetrad probabilities as functions of
multilocus ordered tetrad probabilities. These relations Type I half-tetrads: For MI nondisjunction, patterns AA
and aa can result only from SDS ordered tetrads. Pattern are then used to derive linear equality and inequality
constraints among multilocus half-tetrad probabilities Aa can result from both FDS and SDS tetrads. Under
RRA, SDS gives rise to AA, Aa, and aa with probability 1/4, imposed by NCI. The constraints can be used to test
NCI, order markers, and construct genetic maps under 1/2, and 1/4. Therefore, P(AA)5P(aa) 5P(SDS)/4
and P(Aa) 5 P(SDS)/2 1 P(FDS). This leads to the
a certain chiasma process model. We discuss one-marker
and two-marker cases in detail before presenting the inequality constraint for type I half-tetrads: P(Aa) $
P(AA)1 P(aa) 5 2 P(AA). This inequality constraint
general results for multiple markers. The four half-tetrad
types are discussed in the order of type IIa, IIb, Ia, and is imposed by RRA.
No chromatid interference (two markers, type IIa Ib half-tetrads, respectively.
Since only two of the four strands are recovered in a half-tetrads):For two markersAandB, with the parent undergoing nondisjunction carrying AB on one chro-half-tetrad, the original ordering of the four strands is
Ab/Ab, AB/aB, AB/ab, Ab/aB, Ab/ab, aB/aB, aB/ab, and ab/ h0 5 I 3 p0 and h1 5 H 3 p1, where hi
1 5 (hi10, hi11,
ab. Under RRA, each of the following four pairs should h
i12)9, pi1t5(pi1t0, pi1t1, pi1t2)9,
have the same probability: AB/AB and ab/ab, AB/Ab and
aB/ab, Ab/Ab and aB/aB, and AB/aB and Ab/ab. These
four pairs plus Ab/aB and AB/ab lead to at most six distinct
I5
1 0 0
0 1 0
0 0 1
and H5
1 1⁄4 0
0 1⁄2 0
0 1⁄4 1
. probabilities for half-tetrads with two markers. Two
markersAandBmay be (1) on different chromosomes; (2) on the same chromosome but on different sides of
Therefore, there is a one-to-one correspondence be-the centromere; or (3) on be-the same chromosome and
tween the pi1ti2tand the hi1i2: p05I3h0and p15H 213 on the same side of the centromere. We consider these
h1. Because the pi1ti2tare nonnegative, RRA imposes the
three cases separately.
Two markers on different chromosomes: Let p and q denote constraint that H213h
1 $0.
It was shown in Zhao and Speed (1998) that the the probability of SDS at A and B, respectively. It is
inequality constraints imposed by NCI for ordered tet-easy to show that P(AB/AB)5 P(ab/ab)5P(Ab/Ab)5
rad probabilities are
P(aB/aB)5[(12p)(12q)]/4; P(AB/Ab)5P(aB/ab)5
q(12p)/2; P(AB/aB)5P(Ab/ab)5 p(1 2 q)/2; and
T21
1 p0$0 and T211p1 $0,
P(Ab/aB)5P(AB/ab)5pq/2. These four distinct
proba-bilities are determined by two parameters, p and q. where
Two markers on different sides of the centromere (A–CEN–B):
It was shown inZhaoandSpeed(1998) that there are at most five distinct probabilities for ordered tetrads when
T15
1 0 1⁄4
0 1 1⁄2
0 0 1⁄4
and T21 1 5
1 0 21
0 1 22
0 0 4
. two markers are on different sides of the centromere. The
corresponding five classes can be described as: (1) FDS
at bothAandBand parental ditype betweenAandB; Using these constraints for the p
i1ti2t, it can be shown that,
(2) FDS at bothAandBbut nonparental ditype between
under NCI, the hi1i2satisfy the following inequality
con-AandB; (3) FDS atAand SDS atB; (4) SDS atAand
straints: h00$h02, h01$2h02, h10$h12, 3h11$2h12, and FDS atB; and (5) SDS at both A and B. Denote the
2h12$h11. No equality constraints are imposed by NCI probabilities of these five classes by a, b, g, d, and ε;
among these six half-tetrad probabilities. the constraints imposed by NCI are the proportionality
Using constraints under NCI, we can distinguish dif-constraint P([AB, ab, AB, ab]):P([AB, ab, Ab, aB]):P([Ab,
ferent configurations for two markers: on different
chro-aB, Ab, aB])51:2:1, and the inequality constraintsa $
mosomes, on the same side, or on different sides of the bandg 1 d $2b. Under RRA, P(AB/AB)5P(ab/ab)5
centromere.ZhaoandSpeed(1998) discussed how to a/2, P(Ab/Ab)5 P(aB/aB) 5 b/2, P(AB/Ab)5 P(aB/
apply constraints imposed by NCI to order markers
us-ab)5 g/2, P(AB/aB)5P(Ab/ab)5 d/2, and P(Ab/aB)
ing ordered tetrads.
5 P(AB/ab) 5 ε/2. Therefore, there are five distinct
No chromatid interference (two markers, type IIb probabilities for type IIa half-tetrads when two markers
half-tetrads):For type IIb half-tetrads—because haplo-are on different sides of the centromere. The equality
type information is unavailable—two patterns, AB/ab constraint imposed by NCI is P(Ab/aB) 5 P(AB/ab).
and Ab/aB, which are distinguishable in type IIa half-The constraintsa $ bandg 1 d $2bfor ordered tetrads
tetrads, are no longer distinguishable. This leads to 9, imply similar constraints among half-tetrad probabilities.
instead of 10 distinguishable patterns: (AA; BB), (AA;
Two markers on the same side of the centromere (CEN–A–B,
Bb), (AA; bb), (Aa; BB), (Aa; Bb), (Aa; bb), (aa; BB), (aa;
the case of CEN–B–Acan be discussed similarly): There are Bb), and (aa; bb). Under RRA, the following four pairs
six distinct probabilities for ordered tetrads in this case. should have the same probability: (AA; BB) and (aa; These six types can be distinguished by whetherAshows bb), (AA; Bb) and (aa; Bb), (AA; bb) and (aa; BB), and FDS or SDS pattern and whetherAandBshow parental (Aa; BB) and (Aa; bb).
ditype (P), tetratype (T ), or nonparental ditype (N) . Two markers on different chromosomes: Because AB/ab Denote these types by (it
1it2), where it150 or 1 corre- and Ab/aB have the same probability when markers are sponds to FDS or SDS atA, and it
250, 1, or 2 corre- on different chromosomes, there is no loss of informa-sponds to P, T, or N betweenAandB. The probability tion under NCI compared to type IIa data. The probabil-of (it
1it2) is denoted by pit
1i2t. For half-tetrads, there are six ities are the same as type IIa half-tetrads.
distinct probabilities as well. Each of these six types can Two markers on different sides of the centromere (A–CEN–B):
be denoted by (i1i2), where i15 0 or 1 corresponding As above, AB/ab and Ab/aB have the same probability, toAbeing homozygous or heterozygous, and i250, 1, or and there is no loss of information under NCI compared 2 corresponding 0, 1, or 2 strands showing recombination to type IIa data. We arrive at the same probabilities and betweenA and B. These probabilities are denoted by constraints as those for type IIa half-tetrads. The only exception is that there is no longer the equality
straint P(Ab/aB) 5 P(AB/ab) because these two types g/4 1 d/4 1 ε/8; and P(Ab/aB)5 b 1 g/4 1d/4 1 are not distinguishable for type IIb half-tetrads. ε/8. The equality constraints imposed by NCI are that
Two markers on the same side of the centromere (CEN–A–B, the probabilities of AB/AB and ab/ab are equal to the
the case of CEN–B–A can be discussed similarly): Unlike probabilities of Ab/Ab and aB/aB. Denote the
probabili-the above two cases, AB/ab and Ab/aB have different ties of these five classes by p1, p2, p3, p4, and p5. The probabilities for type IIa half-tetrads whenAandBare probabilitiesa, b,g, d, and εfor ordered tetrads can on the same side of the centromere. Thus, there is some be obtained from the pi, i 51, · · · , 5:
loss of information because AB/ab and Ab/aB cannot be
a 51⁄2(p12 p
22p312p5), distinguished. There are five, instead of six, distinct
probabilities. These five classes can be represented as b 51⁄2(p12 p
22p312p4), (0, i2), which corresponds toAbeing homozygous and
g 52(p3 2p1),
i250, 1, or 2 strands showing recombination between
A andB; and (1, i2), which corresponds to Abeing d 52(p22p1), heterozygous andBbeing homozygous (i250) or het- ε
54p1. erozygous (i25 1). Let ui1i2 denote the probability of
RRA imposes the constraints that the expressions on type (i1i2); we have u05I3p0and u15U3p1, where
the right-hand side of the above equations be nonnega-u05 (u00, u01, u02)9, u15(u10, u11)9,
tive. Because the inequality constraints among ordered tetrad probabilities imposed by NCI are a $ b and U5
0 1⁄2 0
1 1⁄2 1
, g 1 d $ 2b (Zhao and Speed 1998), the inequalityconstraints among the pi imposed by NCI are p5 $ p4 and 3p21 3p3 $5p1 12p4.
and pit
1i2twas defined above as the probability of ordered
Two markers on the same side of the centromere (CEN–A–B,
tetrad pattern (it
1it2).
the case of CEN–B–A can be discussed similarly): There
Therefore, we can obtain only p10 1 p12 from the
are six distinct probabilities. Each of these six types is
ui1i2 but not the individual values of p10 and p12. The
denoted by (i1i2), where i1and i2 were defined in the inequality constraints on the pi1ti2t, imposed by NCI are
discussion of type IIa data. Denote the probability of
p00$p02, p01$2p02, p10$p12, and p11$2p12(Zhaoand half-tetrad pattern (i
1i2) by hIi1i2. The relations between
Speed1998). As long as p101p12$0 (i.e., u112u10$0)
the hI
i1i2and the pi1ti2tare h I
051⁄2I3p1and hI15H3p01 the inequalities involving the p1it
2are always satisfied by
1⁄2I3p1, where hI
i1, pi1t, I, and H are similarly defined as
setting p12to 0. Because the relations between the u0i2
for type IIa half-tetrads. RRA imposes the constraints and the p0i2are the same as the relations between the
that the pi1ti2tinferred from the h I
i1i2are nonnegative. The
h0i2and the p0i2, the constraints under NCI are u00$u02,
inequality constraints among the hI
i1i2 imposed by NCI
u01$ 2u02, and u11$ u10.
can be derived from the constraints among the pit 1i2t. It
No chromatid interference (two markers, type Ia
half-can be shown that these constraints are tetrads):For type Ia half-tetrads, the strands in the same
half-tetrad were not attached to the same centromere at the four-strand stage during meiosis. Under RRA, each of the four nonsister chromatid pairs has the same chance of being recovered in a half-tetrad. As for type
21 0 1 1 0 21
0 21 2 0 3 22
0 0 24 0 22 4
2 0 22 0 0 0
0 2 24 0 0 0
0 0 8 0 0 0
hI
00
hI
01
hI
02
hI
10
hI
11
hI
12
$
0 0 0 0 0 0
. IIa half-tetrads, there are 10 distinguishable types and
at most six distinct probabilities.
Two markers on different chromosomes: We also use p and
q to denote the probability of SDS at AandB. It can
No chromatid interference (two markers, type Ib half-be shown that P(AB/AB) 5 P(ab/ab) 5 P(Ab/Ab) 5
tetrads):As type IIb half-tetrads, because haplotype
in-P(aB/aB)5 pq/16, P(AB/Ab)5P(aB/ab)5 p(12q)/
formation is unavailable, patterns AB/ab and Ab/aB
can-41pq/8, P(AB/aB)5P(Ab/ab)5q(12p)/41pq/8,
not be distinguished. There are nine distinguishable and P(Ab/aB)5P(AB/ab)5(12p)(12q)/21p(12
patterns. Under RRA, the following four pairs of
pat-q)/41q(12p)/41pq/8. These four distinct
probabili-terns have the same probability: (AA; BB) and (aa; bb), ties are determined by two parameters, p and q.
(AA; Bb) and (aa; Bb), (AA; bb) and (aa; BB), and (Aa;
Two markers on different sides of the centromere (A–CEN–B):
BB) and (Aa; bb). Therefore, there are at most five
There are five classes, each with a distinct probability.
distinct probabilities for type Ib half-tetrads. Using the same notations as in the discussion of type
Two markers on different chromosomes: Because AB/ab and
IIa half-tetrads to define the probabilities of these five
Ab/aB have the same probability, there is no loss of
infor-classes, we have P(AB/AB) 5 P(ab/ab)5 P(Ab/Ab) 5
mation under NCI compared to type Ia data. The
probabil-P(aB/aB)5 ε/16; P(AB/Ab)5 P(aB/ab)5 d/41ε/8;
Two markers on different sides of the centromere (A–CEN–B): denoted by 0, 1, 2, and 3, respectively, at each marker Ar. The pattern (i1i2. . . in) of each type IIa half-tetrad
Since P(AB/ab) and P(Ab/aB), in general, are different
for type Ia half-tetrads, there is information loss in type is thus defined with ir 5 0, 1, 2, and 3, respectively.
Because two strands in a half-tetrad are not labeled, we Ib half-tetrads compared to type Ia half-tetrads. There
are four distinct probabilities. Because P(AB/ab) 1 can evenly divide the cases by two different labelings of the two strands. The results in the following discussion
P(Ab/aB)5 a 1 b 1 g/21 d/21ε/4, ordered tetrad
probabilitiesa and b cannot be uniquely determined hold under this even division.
There are 233n21distinct probabilities for ordered from type Ib half-tetrad probabilities. Both linear
in-equality constraints, a $ b and g 1 d $ 2b, can be tetrad data under NCI. These different ordered tetrad classes are denoted by (it
1it2. . . itn), where it150 or 1 satisfied by settingbto be 0. The only constraints imposed
by NCI are the equality constraints that the probabilities corresponding to FDS or SDS at A1, and it
r50, 1,
or 2, r5 2, . . . , n, corresponding to parental ditype, of AB/AB and ab/ab are equal to the probabilities of
Ab/Ab and aB/aB. tetratype, and nonparental ditype between Ar21 and
Ar. Let pit
1i2t...intbe the probability of type (i t
1it2 . . . itn). In
Two markers on the same side of the centromere (CEN–A–B,
the case of CEN–B–Acan be discussed similarly): There are the following, we derive the relations between type IIa
five different classes with distinct probabilities. These half-tetrad probabilities, the hi
1i2...in, and ordered tetrad
five classes are denoted by (i1i2), where i1 and i2 were probabilities, the p
it
1i2t...int. For one marker,
defined in the discussion of type IIb data. Let uI
i1i2denote
the probability of pattern (i1i2). It can be shown that uI
05 1⁄2I3 p1and uI1 5 U3 p0 1 1⁄2U3 p1, where uI0, uI1, U and the p
it
1i2tare similarly defined as in the
discus-
h0 h1 h2 h3
5
1⁄2 0
0 1⁄2 0 1⁄2 1⁄2 0
p0 p1 5C1 p0 p1 .
sion of type IIb half-tetrads. From the uI
i1i2, we cannot uniquely determine p00and
p02. Only the sum of p00and p02 can be inferred, and For two markersA
1andA2, we consider four patterns we denote it by p*00. It is easy to show the following: atA
1separately. If the pattern atA1is A1/A1, parental ditype betweenA1 andA2 will result in A2/A2 at A2, tetratype betweenA1andA2will result in A2/a2or a2/
A2 at A2 with the same probability, and nonparental ditype betweenA1andA2will result in a2/a2atA2. If
p *00
p01 p10 p11 p12
5
21 0 21 21 1
0 21 0 2 0
2 0 0 0 0
0 2 0 0 0
0 0 2 0 0
uI 00 uI 01 uI 02 uI 10 uI 11
.the pattern atA1is a1/a1, parental ditype betweenA1 andA2will result in a2/a2atA2, tetratype betweenA1 andA2will result in A2/a2or a2/A2atA2with the same The constraints imposed by RRA are that the pit
1i2tin- probability, and nonparental ditype between A1 and ferred from the ui1i2 being nonnegative. We can also A2 will result in A2/A2 at A2. If the pattern at A1is
A1/a1, under NCI, there is equal chance that the four-derive the inequality constraints imposed by NCI from
strand bundle during meiosis has configuration [A1,a1; the above relations.
A1,a1], [A1,a1; a1,A1], [a1,A1; A1,a1], or [a1,A1; a1,A1]. There-No chromatid interference (multiple markers on the
fore, parental ditype betweenA1andA2will result in same side of the centromere): For n markers in the
A2/a2at A2, tetratype between A1 and A2 will have order of CEN–A1–A2– · · · –An, there are 2n21(2n11)
the same probability resulting in A2/A2, A2/a2, a2/A2, or distinguishable patterns (appendix,Proposition 1) for
a2/a2 at A2, and nonparental ditype betweenA1and both type Ia and type IIa half-tetrads. For type Ib and
A2will result in a2/A2atA2. The case that the type at type IIb half-tetrads, there are three possible patterns
A1being a1/A1can be considered similarly. Therefore, at each marker: ArAr, Arar, and arar. Therefore, there
are 3ndistinguishable patterns for type Ib and type IIb
half-tetrads. Under RRA, the number of distinct proba-bilities is at most 4n21 1 2n21 (appendix, Proposition
h00 h01 A h33
5
1⁄2X0 0X0
0X1 1⁄2X1 0X2 1⁄2X2 1⁄2X3 0X3
p00 p01 A p12
, 2) for type Ia and type IIa half-tetrads. Among the 3ndistinguishable patterns for type Ib and type IIb half-tetrads, there are at most (3n11)/2 distinct
probabili-ties (appendix,Proposition 3). where
Type IIa half-tetrads: To simplify the derivation of the
general results for n markers, we proceed differently from the discussion of the one- and two-marker cases.
We first assume that the two strands have already been X05
1 0 0
0 1⁄2 0 0 1⁄2 0
0 0 1
, X15
0 1⁄4 0 1 1⁄4 0 0 1⁄4 1 0 1⁄4 0
, labeled and are thus distinguishable. Then there areshown in Zhao andSpeed (1998) that the inequality constraints for ordered tetrads are
X25
0 1⁄4 0 0 1⁄4 1 1 1⁄4 0 0 1⁄4 0
, X35
0 0 1
0 1⁄2 0 0 1⁄2 0
1 0 0
. T21
n21p0$ 0 and T2n211p1 $0,
where T1 and T21
1 were defined in (1) and T2n211 5 T21^(n21)
1 . The operator^is the standard tensor product For an arbitrary n, the probability hi1i 2...inof the type IIa
(see, e.g.,Bellman1970). Therefore, the inequality con-half-tetrad pattern (i1i2. . . in) can be expressed in terms
straints among the hi1i2...incan be established. A likelihood
of the pi1tit2...itnas
ratio test can be used to test these constraints; seeZhao
hi1i2...in5
o
i1ti2t...itnci1i2...in i1ti2t...intpi
t
1i2t...int. et al. (1995a).
Type IIb half-tetrads: For type IIb half-tetrads, there are
three patterns at each marker. These three patterns are Write the ci1i2...in
i1ti2t...intinto a matrix Cnsuch that the columns
denoted by 0, 1, and 2, corresponding to observing 0, are labeled by it
1it2. . . itn and the rows are labeled by
1, and 2 copies of allele AratAr, r 5 1, · · · , n. The
i1i2. . . in, each in lexicographical order. It can be shown
probability for each type (i1i2. . . in) can be expressed
that (appendix,Theorem 1) the 4r113(233r) matrix
in terms of the ordered tetrad probabilities pit 1i2t...intas
Cr115(c
i1i2...irir11 it
1i2t...irtirt11) can be obtained recursively by
replac-ing each ci1i2...ir it
1i2t...irt
in Cr 5 (c i1i2...ir it
1i2t...irt) by the 4 3 3 matrix ui1i2...in5
o
it1i2t...int ai1i2...in
it 1i2t...intpi
t 1i2t...int. ci1i2...ir
it 1i2t...irtXir.
Because the probabilities hi1...in can be expressed in Write the a i1i2...in it
1i2t...int into a matrix such that the columns
terms of the pit
1...int through the matrix Cn, for two identi- are labeled by it1it2. . . itn and the rows are labeled by
i1i2. . . in, each in lexicographical order. It is easy to see
cal rows in Cn, the corresponding half-tetrad patterns
should have the same probability. Note that some of these equalities are the result of RRA and they can be
readily identified. Equality constraints under NCI can A 15
1⁄2 0
0 1 1⁄2 0
. be established by removing these equality constraints
resulting from RRA.
The inequality constraints can be established as fol- Define lows. Define
D15
1 0 0 1
0 1 1 0
, E05
1 0 0
0 1 0
0 0 1
, E15
0 1⁄ 4 0 1 1⁄
2 1 0 1⁄
4 0
, E25
0 0 1
0 1 0
1 0 0
;
then the matrix Ar11 5 (a
i1i2...irir11 it
1i2t...irtirt11) can be obtained by
Y05
1 0 0 0
0 1 1 0
0 0 0 1
, Y1 5
21⁄2 1 0 21⁄2
2 0 0 2
21⁄2 0 1 21⁄2
,
replacing each ai1i2...ir it
1i2t...irtin Arby the 333 matrix a i1i2...ir it
1i2t...irtEir.
In the discussion of two-marker data, it was noted that
p00and p02cannot be determined from the ui1i2. Similarly
in the n marker case, not all the pit
1i2t...intcan be recovered
Y25
21⁄2 0 1 21⁄2
2 0 0 2
21⁄
2 1 0 21⁄2
, Y35
0 0 0 1
0 1 1 0
1 0 0 0
. from the u
i1i2...in. Equality constraints can be established
as in the discussion of type IIa half-tetrads. To establish inequality constraints, define
The matrix Dr115(ci
t 1it2...itritr11
i1i2...irir11) is defined recursively from
Dr5(d it
1it2...itr
i1i2...ir ) by replacing each d it1it2...itr
i1i2...ir in Drby the 334
B15
1 0 1
0 1 0
,
matrix dit1it2...itr
i1i2...ir Yir. From the facts that
D1C15
1 0 0 1
F05
1 0 0
0 1 0
0 0 1
, F15
21 1 21 2 0 2
0 0 0
, F25
0 0 1
0 1 0
1 0 0
. and YiXi5 I333for i50, 1, 2, and 3, it is easy to show
that
The matrix Br11 5 (b
i1tit2...itri t r11
i1i2...irir11) is defined by replacing
each bi
t 1it2...itr
i1i2...ir in Br 5 (b it1it2...itr
i1i2...ir) by the 3 3 3 matrix
DnCn5
I3n2133n21 0
0 I3n2133n21 .
bit1it2...itr
i1i2...irFir. It can be shown that RRA imposes the
con-So the p0it
2...int and p1i2t...int can be recovered from the straints that p 5 Fnu $ 0, and NCI imposes the
con-straints that T21
n p5 T2n1Fnu$ 0.
Type Ia half-tetrads: For type Ia half-tetrads, let hI
i1i2...in have been labeled. Any type IIa half-tetrad pattern can
be represented by ij5(i1i2. . . in; j1j2. . . jm), where each
and pit
1i2t...itn denote the half-tetrad and ordered tetrad
ik(k5 1, · · · , n) or jl(l 51, · · · , m) is 0, 1, 2, or 3.
probabilities. The relations between the hI
i1i2...inand the
The probability of this half-tetrad pattern is denoted by
pit
1i2t...intcan be expressed as h
(i1i2...in;j1j2...jm). If the centromere were observable, ordered
tetrad pattern could be represented by itjt 5 (it
1it2. . .
hI
i1i2...in5
o
it1i2t...int
ci1i2...in it
1i2t...intpi t 1i2t...int.
it
n;jt1jt2. . . jtm), where each itk( jtl) is 0, 1, or 2,
correspond-ing to parental ditype, tetratype, or nonparental ditype Write theci1i2...in
it
1i2t...intinto a matrixCnsuch that the columns
between Ak21 andAk(Bl21andBl). Both A0andB0 are labeled by it
1it2. . . itn and the rows are labeled by
correspond to CEN. The hijcan be expressed in terms i1i2 . . . in, each in lexicographical order. Using
argu-of the pitjtas
ments similar to those used in the proof of type IIa half-tetrads in theappendix,it can be shown that
hij5
o
gijitjtpitjt.It is shown in theappendix(Theorem 2) that
gijitjt5
o
it1,jt1
vi1j1 it
1j1tq i itqjjt,
C15
0 1⁄4 1⁄2 1⁄4
1⁄2 1⁄4
0 1⁄4
where vi1j1it
1j1t is the element in the (i1j1)th row and the
and that the matrix Cr11 5 (c
i1i2...irir11 it
1i2t...irtirt11) is obtained by (i t
1jt1)th column of the following matrix: replacing each ci1i2...ir
it
1i2t...irt in Cr by the 4 3 3 matrix
ci1i2...ir it
1i2t...irtXir, where X1, X2, X3, and X4 were defined in the
discussion of type IIa half-tetrads. Linear equality and inequality constraints imposed by RRA and NCI can be similarly established.
Type Ib half-tetrads: For type Ib half-tetrad data, the
relations between the uI
i1i2...inand the pi1ti2t...int can be
ex-pressed as
uI
i1i2...in5
o
i1ti2t...intφi1i2...in it
1i2t...intpi t 1i2t...int.
(00) (01) (02) (10) (11) (12) (20) (21) (22)
(00) 1⁄2 0 0 0 0 0 0 0 1⁄2
(01) 0 1⁄4 0 0 0 0 0 1⁄4 0 (02) 0 1⁄4 0 0 0 0 0 1⁄4 0 (03) 0 0 1⁄2 0 0 0 1⁄2 0 0 (10) 0 0 0 1⁄4 0 1⁄4 0 0 0
(11) 0 0 0 0 1⁄4 0 0 0 0
(12) 0 0 0 0 1⁄4 0 0 0 0
(13) 0 0 0 1⁄4 0 1⁄4 0 0 0 (20) 0 0 0 1⁄4 0 1⁄4 0 0 0
(21) 0 0 0 0 1⁄4 0 0 0 0
(22) 0 0 0 0 1⁄4 0 0 0 0
(23) 0 0 0 1⁄4 0 1⁄4 0 0 0 (30) 0 0 1⁄2 0 0 0 1⁄2 0 0 (31) 0 1⁄4 0 0 0 0 0 1⁄4 0 (32) 0 1⁄4 0 0 0 0 0 1⁄4 0
(33) 1⁄2 0 0 0 0 0 0 0 1⁄2
Write theφi1i2...in it
1i2t...intinto a matrix such that the columns are
labeled by it
1it2. . . itnand the rows are labeled by i1i2. . . in,
each in lexicographical order. It can be shown that
F15
0 1⁄ 4 1 1⁄
2 0 1⁄
4
and that the matrix Fr11 5 (φ
i1i2...irir11
i1ti2t...irtirt11) is obtained by
replacing eachφi1i2...ir it
1i2t...irtinFrby the 33 3 matrixφ i1i2...ir it
1i2t...irtEir Write the coefficients q i it5 q
i1i2...in
it
1i2t...int into a matrix Qnsuch
where E0, E1, and E2 were defined in the discussion of that the columns are labeled by it
1it2...itn and the rows
type IIb half-tetrads. The linear equality and inequality are labeled by i1i2...in, each in lexicographical order. As
constraints under RRA and NCI can be established simi- in the derivation of Theorem 1 (appendix), it can be larly to those for type IIb half-tetrads. shown that
No chromatid interference (multiple markers on different sides of the centromere): Consider markers on different sides of the centromere in the order of
Bm– · · · –B1–CEN–A1– · · · –An. Here we show only Q15
1 0 0
0 1⁄2 0
0 1⁄2 0
0 0 1
the relations between type IIa and type IIb half-tetrad probabilities and ordered tetrad probabilities. Con-straints imposed by RRA and NCI can be derived using
and that the matrix Qr115 (q
i1i2...irir11 it
1i2t...irtirt11) is obtained by
re-these relations following the approach described above.
placing each qi1i2...ir it
1i2t...irtby the the 433 matrix q i1i2...ir it
1i2t...irtXir, where
Derivations for type Ia and type Ib half-tetrads are
simi-X0, X1, X2, and X3were defined in the discussion of type lar, and we omit the details here.
centromere. The coefficients qjjt5q j1j2...jm jt
1j2t...jmtare defined the between this marker and the centromere can be easily
derived. same as qi
it5qi1i2...in it
1i2t...int.
A crossover process model is needed for multilocus
Type IIb half-tetrads: As for type IIa half-tetrads, the
analysis. Different models have been proposed in the liter-probability uijfor type IIb half-tetrad pattern ij5 (i1i2
ature to model the crossover process (McPeekandSpeed . . . in;j1j2. . . jm), where each ikor jlis 0, 1, or 2, can be
1995). Among them, the chi-square model was found to expressed in terms of the pitjtas
provide better fit to data from different organisms (Zhao
et al. 1995b).
uij5
o
fij itjtpitjt.
The chi-square model, which was first introduced by It can be shown that Fisheret al. (1947), was suggested as a plausible biological model byFosset al. (1993). The model, which is
repre-fijitjt5
o
it1,jt1
wi1j1 it
1j1ts i
itsjjt,
sented as Cx(Co)m, assumes that the crossover
intermedi-ates, C events, are randomly distributed along the four-where wi1j1
it
1j1t is the element in the (i1j1)th row and the strand bundle, and every intermediate resolves either as
(it
1jt1)th column of the following matrix:
a crossover (Cx) or not (Co). When an intermediate re-solves as a Cx, the next m intermediates must resolve as a
Co, and after m Co’s the next intermediate must resolve
as a Cx. The process is made stationary by letting the leftmost crossover intermediate have the same chance of being one of Cx(Co)m. The chi-square model has recently
been generalized to a more general class, the Poisson-skip (00) (01) (02) (10) (11) (12) (20) (21) (22)
(00) 1⁄2 0 0 0 0 0 0 0 1⁄2
(01) 0 1⁄2 0 0 0 0 0 1⁄2 0
(02) 0 0 1⁄2 0 0 0 1⁄2 0 0
(10) 0 0 0 1⁄2 0 1⁄2 0 0 0
(11) 0 0 0 0 1 0 0 0 0
(12) 0 0 0 1⁄2 0 1⁄2 0 0 0
(20) 0 0 1⁄2 0 0 0 1⁄2 0 0
(21) 0 1⁄2 0 0 0 0 0 1⁄2 0
(22) 1⁄2 0 0 0 0 0 0 0 1⁄2
model (Lange et al. 1997). Both the chi-square model and the Poisson-skip model lead to closed-form expression for the probability of any ordered tetrad pattern. This gives a rather flexible and tractable class of models for genetic linkage analysis. Note that the Poisson model is a special case of the chi-square model.
For an arbitrary number of markers on the same side Write the si1i2...in
it
1i2t...intinto a matrix Sn, then or different sides of the centromere,Zhao andSpeed (1998) derived general closed-form expressions for or-dered tetrad probabilities under the Cx(Co)m model.
Using these results and the relations we derived between S1 5
1 0 0
0 1 0
0 0 1
,
half-tetrad probabilities and ordered tetrad probabili-ties, we can evaluate any half-tetrad probability. There-fore, maximum likelihood estimates of the interference and Sr11is obtained by replacing each s
i1i2...ir
i1ti2t...irtin Srby the
parameter m and the genetic distances among the mark-3 3 3 matrix si1i2...ir
i1ti2t...irtEir, where E0, E1, E2, and E3 were ers and the centromere are tractable under this class
defined in the discussion of type IIb half-tetrads for
of models. markers on the same side of the centromere. The
coef-ficients sjjt5 s j1j2...jm
j1tj2t...jmt are defined the same as s i
it5si1i2...in i1ti2t...itn.
RESULTS
Multilocus genetic mapping:In the studies of ordered tetrads,ZhaoandSpeed(1998) compared various map
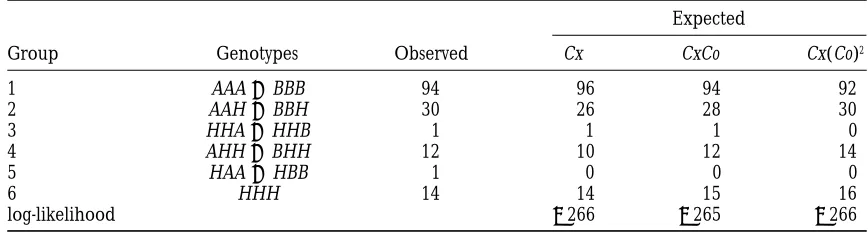
TABLE 1
Observed and expected counts of six different half-tetrad types of alfalfa progeny
Expected
Group Genotypes Observed Cx CxCo Cx(Co)2
1 AAA1BBB 94 96 94 92
2 AAH1BBH 30 26 28 30
3 HHA1HHB 1 1 1 0
4 AHH1BHH 12 10 12 14
5 HAA1HBB 1 0 0 0
6 HHH 14 14 15 16
log-likelihood 2266 2265 2266
Three markers in the order of CEN–UWg 119–MTSc 9–UWg65 were typed. The observed genotypes are denoted by G1G2G3, where Gi5A or B corresponding to homozygous alleles at the ith marker and Gi5H corresponding to heterozygous alleles at the ith marker.
et al. 1996) and rainbow trout (Thorgaardet al. 1983) three intervals. The CxCo model, which imposes
moder-ate amount of chiasma interference, gave almost perfect via the method of maximum likelihood. Haplotype
in-formation is unavailable in both data sets. Because there fit to the observed data under the assumption of no meiosis I nondisjunctions. Recall that complete chiasma is little consistent evidence of chromatid interference
(Zhao et al. 1995a) and both data sets yield little evi- interference was assumed in deriving the estimate of
the meiosis I nondisjunction proportion in alfalfa by dence of chromatid interference, NCI is assumed in the
following analyses. For both data sets, we assume known Tavolettiet al. (1996). Therefore, it is difficult to dis-tinguish the model studied by Tavolettiet al. (1996) marker order and use the chi-square model for the
chiasma process. and the chi-square model studied here using this data set. Note that in some cases, the meiosis I nondisjunc-Alfalfa:By assuming complete chiasma interference,
Tavolettiet al. (1996) introduced a maximum likeli- tion proportion, pMI, and the map distance between the centromere and the most proximal marker, dCEN–A,
hood approach to analyzing half-tetrads from alfalfa.
We analyze a subset of three markers in their study. cannot be simultaneously identified. The Poisson model is the simplest such model, under which dCEN–A varies
These three markers are in the order of CEN–UWg 119–
MTSc9–UWg 65 and were genotyped in 152 progeny. according to pMI. In general, the estimate of dCEN–A
in-creases as the estimate of pMIdecreases. This is because Tavolettiet al. (1996) found that a very small
percent-age, approximately 6%, of half-tetrads in this organism as pMIdecreases, more crossover events are needed be-tween CEN andAto explain the observed heterozygous were the results of meiosis I nondisjunctions. To study
whether the observed data can be explained by a moder- half-tetrads at A, thus increasing the estimated map distance between them. As an example, for the alfalfa ate chiasma interference and no meiosis I
nondisjunc-tions, we fitted the chi-square model to the data set, data, the estimated map distance between CEN and
UWg 119 is 5 cM when no meiosis I nondisjunctions are
and the results are presented in Table 1. In our analysis,
all half-tetrads were assumed to be type IIb half-tetrads assumed, whereas the estimated map distance is 3 cM when the meiosis I nondisjunction rate is estimated at (i.e., they all resulted from meiosis II nondisjunctions).
The CxCo model gave the best fit among the chi-square around 6%.
Rainbow trout: Two markers in the order of CEN– models. The estimated map distances under the CxCo
model were 5, 4, and 11 cM in the three intervals CEN– Idh2–Est1 in rainbow trout were studied byThorgaard
et al. (1983). A total of 138 progeny were genotyped. UWg119, UWg 119–MTSc 9, and MTSc 9–UWg65,
respec-tively. The standard errors were estimated using the The number of progeny for each of the six observed half-tetrad types are given in Table 2. When the Cx(Co)m
parametric bootstrap method by (1) simulating data
sets with the same sample size under the CxCo model models, where m 50, · · · , 6, were fitted to the data, the model with the greatest degree of chiasma inter-assuming the estimated parameter values; (2) estimating
model parameters for each simulated data set; and (3) ference, the Cx(Co)6 model, gave the best fit to this data set. This is consistent with the conclusion of Thor-approximating the standard errors of the parameter
estimates using the standard errors of the estimated gaard et al. (1983) that there is high interference in rainbow trout. The estimated map distances under the parameter values from these simulated data sets. Using
this method, the standard errors were estimated to be Cx(Co)6model were 36 and 11 cM in the two intervals
CEN–Idh2 and Idh2–Est1. The corresponding standard
1, 1, and 2 cM, respectively. The above estimated genetic
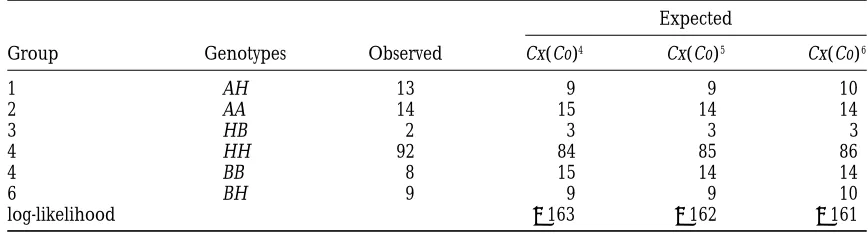
TABLE 2
Observed and expected counts of six different half-tetrad types of rainbow trout progeny
Expected
Group Genotypes Observed Cx(Co)4 Cx(Co)5 Cx(Co)6
1 AH 13 9 9 10
2 AA 14 15 14 14
3 HB 2 3 3 3
4 HH 92 84 85 86
4 BB 8 15 14 14
6 BH 9 9 9 10
log-likelihood 2163 2162 2161
Two markers in the order of CEN–Idh2–Est1 were typed. The observed genotypes are denoted by G1G2, where
Gi5A or B corresponding to homozygous alleles at the ith marker and Gi5H corresponding to heterozygous alleles at the ith marker.
genetic distances in these two intervals are 35 and 9 cM, address chiasma interference issue in their half-tetrad analysis. But their methods either were only applicable respectively. Our estimates agree fairly well with their
esti-mates. to one-marker data or made different assumptions
about chiasma interference in the same analysis for mul-In both examples, we have assumed no chromatid
interference. If chromatid interference indeed exists, tiple markers. In contrast, our proposed approach in this article applies to any crossover process model that map distances can be either over- or underestimated
depending on the specific pattern of chromatid interfer- incorporates chiasma interference. Zhao and Speed (1998) noted that most map functions proposed in the ence. In addition, chiasma interference can be
incor-rectly “detected” even if it is absent. A detailed study of context of centromere mapping can be well approxi-mated by the map function under the Cx(Co)2 model. chromatid interference is reported inH. ZhaoandT. P.
Speed (unpublished results). Therefore, multilocus gene-centromere mapping, as de-scribed above using the chi-square model as the cross-over process model, may provide a tractable and flexible
DISCUSSION
approach to analyzing multilocus half-tetrad data. The chi-square model has recently been extended to allow In this article, four types of half-tetrad data, type Ia,
Ib, IIa, and IIb, were studied. Half-tetrads, just like or- a more general class of interarrival distributions, yet the tractability of the model is still preserved (Langeet al. dered and unordered tetrads, provide information on
both chromatid interference and chiasma interference. 1997). This class of generalized chi-square models can also be used to analyze multilocus half-tetrad data. In addition, they can be used to map centromeres.
Under the assumptions of no chromatid interference H. ZhaoandT. P. Speed(unpublished results) stud-ied a Markov model for chromatid interference. They and random recovering of half-tetrads, the relations
between ordered tetrad probabilities and half-tetrad showed how this chromatid interference model can be applied with the chi-square model to study both chroma-probabilities were established. Using these relations, we
derived constraints among half-tetrad probabilities im- tid and chiasma interference. Their approach can be easily adopted here to study both types of genetic inter-posed by RRA and NCI. These constraints can be used
to test for NCI. When the order of the markers and ference using half-tetrads.
Throughout this article, we have assumed that the their relations to the centromere are not known, the
best order for these markers can be established through type of half-tetrads observed is known (type Ia, Ib, IIa, or IIb) and the phases in parents are known. Although examining the constraints imposed by NCI.
The relations between tetrad probabilities and half- these assumptions cover many data sets from experimen-tal organisms, they are often violated in human data. For tetrad probabilities can be used to construct genetic maps
and to locate centromeres under a given chiasma process example, autosomal trisomies could be the result of MI or MII nondisjunction events. If the probability of each model. This provides an approach to incorporating
chi-asma interference in genetic analysis. Because of the half-tetrad type is known, the maximum likelihood method can still be applied for multilocus gene-centro-presence of chiasma interference in most organisms, the
approach ofDaet al. (1995), which assumes no chiasma mere mapping. Alternatively, we can introduce parame-ters to account for the uncertainties in determining interference, is not consistent with biological evidence.
Several articles in the literature on half-tetrads (Ottet half-tetrad types (H. Zhao,unpublished results). Another assumption we have made is that the
ob-al. 1976;ChakravartiandSlaugenhaupt1987;