ABSTRACT
FETICS, SUSAN KATHLEEN. Structural Analysis of Ras Bound to Raf Kinase and Implications of Ras Allosteric Site. (Under the direction of Dr. Carla Mattos).
The Ras/Raf/MEK/ERK pathway is one of the most prominent signaling cascades studied today. Ras mutations are found in 30% human cancers. Ras is a small GTPase that acts as a molecular switch. Its activation state is dependent on the bound nucleotide. Raf is a kinase that is phosphorylated and binds ATP. When Ras binds Raf, this pathway is active to influence diverse cellular events, such as proliferation, differentiation and transformation. Raf was the first effector of Ras discovered, yet, to date, the structure of Ras bound to a GTP analog in complex with Raf kinase has not been solved. Over the past two decades, the structures of Ras bound to various binding partners have aided in the understanding of the mechanisms of activation for this enzyme. Mutations of Ras, namely valine at residue position 12 (V12) and leucine at residue position 61 (L61), have highly transforming
Structural Analysis of Ras Bound to Raf Kinase and Implications of Ras Allosteric Site
by
Susan Kathleen Fetics
A dissertation submitted to the Graduate Faculty of North Carolina State University
in partial fulfillment of the requirements for the degree of
Doctor of Philosophy
Biochemistry
Raleigh, North Carolina 2012
APPROVED BY:
_______________________________ ______________________________
Dr. Carla Mattos Dr. Michael Goshe
Committee Chair
________________________________ ________________________________
DEDICATION
The six years of hard work spent in North Carolina are dedicated to my proud father George Fetics. Dad passed away February 12, 2012, just months before my graduation. His tangible presence in my life is sadly missed to a depth I've never known.
At 20 years old, in 1954, my dad was drafted into the army and became a nurse. He retired from full time work at the age of 79, after 59 years as a nurse. His impeccable work ethic and purpose in helping others influenced me to find a career that fulfilled me for life. At 30 years old, my dad met my mom. She was hired to be his boss. He was never concerned that she had a higher degree or made more money than he made. He respected her and was always proud of the good work she did as a nurse. Growing up with such an
unconventional father helped me see that no pre-conceived notions can stop me from doing what I love to do.
At 40 years old, my dad became a father for the first time with the birth of my
brother, Barry. As a child, I adored my father. I used to sit on his shoulders in the living room and comb his hair while he watched football. I made him tell me stories from the army or growing up in Pennsylvanian near the coal mines. I cherished lunch dates with my popular dad in the cafeteria of Alexian Brother's Hospital in the town of Elizabeth, New Jersey.
gratitude for the courage and strength you gave me by being your daughter and being at the heart of your existence. Without those gifts, I would not have completed this program. Most importantly, you taught me by example that love always prevails.
BIOGRAPHY
I, Susan Kathleen Fetics, was raised in Roselle, New Jersey. After 13 years of catholic education, I was happy to move to Greenwich Village in New York City and attend New York University as a chemistry major and French minor. While in college, I worked part time at Ambient Labs, Inc, a privately owned analytical lab based in Chelsea. New York City was a wonderful place to live while earning my bachelor's degree. After graduation from NYU, I moved to Washington, DC to pursue graduate studies in chemistry at Georgetown University. Upon graduation with an MS in chemistry, I moved back to New York City and worked as a technician in genetics for 6 years. When most my friends were getting married and having babies, I decided to move to Raleigh, NC, go back to graduate school and committed myself to 6 years of intense work in the Mattos lab. Under the direction of Dr. Carla Mattos, I
ACKNOWLEDGMENTS
I begin by thanking Dr. Carla Mattos. Her enthusiasm ignited my interest in protein interactions on an atomic level and now there is no turning back for me. Greg Buhrman has been the greatest teacher and friend that I could have had during my return to graduate school. Winnell Newman was the best lab manager I've ever worked with. Paul Swartz spent endless hours answering my questions and helping me get a better understanding of my time in graduate school. My committee, consisting of Michael Goshe, Flora Meilleur, Rob Smart, have supported me at each yearly meeting. Kathleen Davis entered the lab as a student I was charged with mentoring, but in the end, she taught me so much with her friendship. Knowing Shya Biswas has inspired me to become a great crystallographer. Thanks to Michelle
DeChene who allowed me to rent her place and create a home where I have a sweet dog waiting for me. Pam, Curt and Nancy in the Biochemistry offices were always available for help. Dr .Dennis Brown did an amazing job as department head of the biochemistry
department at NCSU. Bradley Kearney helped me with countless computer questions and Mychal Smith had good insight with research questions that surfaced. Numerous other undergraduate and graduate students came through the lab during my 6 years that made the lab an exciting place.
TABLE OF CONTENTS
LIST OF TABLES ... xi
LIST OF FIGURES ... xii
CHAPTER 1: Literature Review ... 1
ABSTRACT ... 2
INTRODUCTION ... 2
RasGTPase and signal transduction pathways ... 2
Ras/Raf/MEK/ERK pathway ... 6
Structure of H-Ras ... 11
Ras/Raf Signaling and Cancer ... 16
Structural Analysis of Ras and Cancer Drug Therapies ... 20
Intrinsic Hydrolysis of Ras ... 27
Goals…. ... 29
ACKNOWLEDGEMENTS ... 31
CHAPTER 2: Complex of wtRas-GppNHp bound to Raf kinase shows an active site poised for intrinsic hydrolysis ... 32
ABSTRACT ... 33
INTRODUCTION ... 33
EXPERIMENTAL PROCEDURES ... 37
Raf_RBD_CRD Purification and Ras/Raf Crystallization ... 37
Data Collection and structural refinement ... 39
General features of Ras bound to Ras Binding Domain (RBD) of Raf kinase ... 42
Structural comparison of wtRas-GppNHp / Raf to Raps/Raf ... 50
Omission of CRD model bound to Ras-GppNHp and Raf_RBD ... 58
DISCUSSION ... 60
Concluding Remarks ... 67
ACKNOWLEDGMENTS ... 67
CHAPTER 3: Ras-GppNHp in complex with Raf RBD_CRD: Attempts at ordering the Cysteine Rich Domain ... 68
ABSTRACT ... 69
INTRODUCTION ... 70
EXPERIMENTAL PROCEDURES ... 72
Purification of H-Ras (1-166) and Raf_RBD_CRD (52-184) ... 72
Data Collection ... 73
Structural Refinement ... 73
RESULTS ... 77
Crystal ID 1239 and 1902 ... 79
Crystal 1869 ... 80
Crystal ID 2013, 2014 and 2015 ... 80
Microseeding ... 86
Rigaku Data ... 87
DISCUSSION ... 88
Change reducing agent ... 88
Farnesylation of full-length Ras ... 89
Co-expression of Ras, full length Raf and scaffold protein Shoc2 ... 90
Concluding Remarks ... 91
ACKNOWLEDGMENTS ... 91
CHAPTER 4: Allosteric switch mutant R97L-RasGppNHp stabilizes the ON state and its structure in the GDP-bound form has features seen in the nucleotide-free Ras/Sos complex... 92
ABSTRACT ... 93
INTRODUCTION ... 94
EXPERIMENTAL PROCEDURES ... 96
R97L-Ras-GppNHp mutation, purification and crystallization ... 96
Ras-RafRBD binding assay ... 97
R97L-Ras-GDP purification and crystallization ... 98
RESULTS ... 98
The “on” state of the allosteric switch is stabilized in R97L-RasGppNHp crystals ... 99
R97L-Ras-GDP binds aluminum fluoride in solution unlike wild type Ras-GDP ... 107
R97L-Ras-GDP in the presence of AlF3 is in a conformation similar to that found in the Ras/Sos complex... 110
DISCUSSION ... 118
DTE binding pocket on R97L-Ras-GDP is a targeted cancer drug site ... 128
Concluding Remarks ... 132
ACKNOWLEDGMENTS ... 133
REFERENCES ... 134
APPENDICES ... 146
Appendix A ... 147
Constructs of Raf_RBD_CRD created for expression and purification ... 147
Site-directed mutagenesis of human H-Ras for both truncated (1-166) and full-length (1-189) ... 160
Appendix B ... 169
Ras-Raf purification ... 169
Ras Purification ... 169
Raf purification ... 170
Unicorn FPLC method for Ras/Raf_RBD_CRD purification ... 173
Raf Cation Exchange ... 173
Ras/Raf complex purification ... 174
Appendix C ... 176
Rosetta script for finding coordinates for CRD ... 176
Appendix D ... 177
Supporting Information for “Allosteric modulation of Ras positions Q61 for a direct role in catalysis” ... 177
LIST OF TABLES
CHAPTER 1: Literature Review ... 1 Table 1: Occurrence of Ras and B-Raf in Human cancers ... 16 CHAPTER 2: Complex of wtRas-GppNHp bound to Raf kinase shows an active site
poised for intrinsic hydrolysis ... 32 Table 1: Data collection and refinement statistics ... 41 CHAPTER 3: Ras-GppNHp in complex with Raf RBD_CRD: Attempts at
ordering the Cysteine Rich Domain ... 68 Table 1: Data collection and refinement statistics ... 75 CHAPTER 4: Allosteric switch mutant R97L-RasGppNHp stabilizes
the ON state and its structure in the GDP-bound form has
features seen in the nucleotide-free Ras/Sos complex ... 92 Table 1: Data collection and refinement statistics ... 103 Table 2: Missing switch I residues of R97L-Ras-GDP ... 112 Appendix D: Supporting Information for “Allosteric modulation of Ras
LIST OF FIGURES
CHAPTER 1: Literature Review ... 1
Figure 1. Activation of Ras results in signal transduction of many pathways ... 3
Figure 2. Ras is a small monomeric GTPase ... 4
Figure 3. Activation of Ras/Raf/MEK/ERK pathway ... 7
Figure 4. Diagram of full length cRaf ... 8
Figure 5. Structure of H-Ras (PDB ID: 3K8Y), residues 1-166 ... 12
Figure 6. A magnesium ion is hexacoordinated in Ras-GppNHp ... 15
Figure 7a. Amino acid sequence alignment of human K and H-Ras ... 17
Figure 7b. Superposition of K-Ras (raspberry, 3GFT) and H-Ras (yellow, 2RGE) bound to GppNHp ... 19
Figure 7c. Diversity of amino acid sequences within the hypervariable region at carboxy-terminus of H-, N-, and K- Ras isoforms ... 20
Figure 8. Active site of Ras-GDP-AlF3-GAP-344 complex (1WQ1) ... 23
Chapter 2: Complex of wtRas-GppNHp bound to Raf kinase shows an active site poised for intrinsic hydrolysis ... 32
Figure 1a. Initial crystals of wtRasRaf complex ... 39
Figure 1b. Initial crystals of Q61L-RasRaf complex ... 39
Figure 2. Superposition of wtRasRaf and oncogenic Q61L-RasRaf ... 43
Figure 3. wtRasRaf complex is allosterically “on”. ... 44
Figure 4. Calcium ion hexacoordinated near R97 of wtRasRaf ... 46
Figure 6 Hydrophobic pocket of active site in Q61L-Ras-GppNHp-RafRBD structure ... 48
Figure 7. Active site superposition of Q61L-Ras-RafRBD complex with Q61L-Ras-GppNHp in the R32 space group ... 49
Figure 8. Superposition of wtRasRaf complex and Raps-RafRBD ... 51
Figure 9a. Interaction of K84 from RBD and Ras E31/D33... 52
Figure 9b. Electrostatic interaction between R59 and R67 of Raf and E37 of wtRas ... 53
Figure 9c. Raps/Raf structure has calcium ion coordinated near E125 of RafRBD ... 54
Figure 9d. Shift in residues 100-106 in Raf RBD when bound to wtRas-GppNHp ... 56
Figure 9e. Water molecule near Y71 of Ras when bound to Raf kinase ... 57
Figure 10. Electron density map and Ras-RafRBD model in asymmetric unit of wtRasRaf ... 59
Figure 11. Superposition of G12V- Ras-PI3K and G12V-Ras-GppNHp in the presence of calcium acetate ... 62
Figure 12a. Active site superposition of GDP-A85K_Raf-RBD and wtRas-GppNHp_RafRBD ... 64
Figure 12b. Active site superposition of GDP-A85K_Raf-RBD and wtRas-GppNHp_RafRBD ... 65
Figure 12c. Interaction of K84 and A85K from Raf RBD with E31 and D33 of wtRas-GDP ... 66
Chapter 3: Ras-GppNHp in complex with Raf RBD_CRD: Attempts at ordering the Cysteine Rich Domain ... 68
Figure 1b. Many crystals grew from one nucleation site from micorseeded crystals ... 73
Figure 1c. Breaking apart crystals from 1b in order to soak in zinc ... 73
Figure 2a. Truncation of data by UCLA anisotropy server for crystal 1239 ... 76
Figure 2b. Truncation of data by UCLA anisotropy server for crystal 1902 ... 76
Figure 2c. ) Truncation of data by UCLA anisotropy server for merged crystals 2013_14_15 ... 76
Figure 3. SDS-PAGE gel and chromatogram from size-exclusion column purification of Ras/Raf complex ... 78
Figure 4a. CRD model in Ras-Raf structure. ... 82
Figure 4b. CRD model in Ras-Raf structure ... 83
Figure 4c. Electron density and model in asymmetric unit of wtRasRaf ... 84
Figure 4d. Electron density and model in asymmetric unit of wtRasRaf with CRD electron density ... 85
Figure 5. PolyA chain of 12 residues from CRD model ... 86
Chapter 4: Allosteric switch mutant R97L-RasGppNHp stabilizes the ON state and its structure in the GDP-bound form has features seen in the nucleotide-free Ras/Sos complex... 92
Figure 1a. Mutant R97L-Ras-GppNHp is in the allosteric “on” state ... 101
Figure 1b. Active site of R97L-Ras-GppNHp in R32 space group ... 102
Figure 2a. Comparison of wild type and mutant H-Ras in the allosteric “on” and “off” states ... 105
Figure 3a, b, c, d. Chromatograms from size exclusion column of
R97L-Ras-GDP-AlF3 in the presence of Raf-RBD ... 109
Figure 4a. Superposition of 12 molecules of R97L-Ras-GDP ... 111
Figure 4b. R97L-Ras-GDP is allosterically “on” ... 113
Figure 4c. Superposition of R97L-Ras-GDP and Ras/Sos interface ... 114
Figure 4d. DTE molecule with electron density map ... 115
Figure 4e. Conserved residue E62 interacts at the active site of R97L-Ras-GDP ... 116
Figure 4f. In R97L-Ras-GDP, an absent magnesium results in water molecules neutralizing the negative charge of the phosphate groups on the nucleotide ... 117
Figure 5. Superposition of DTE binding pocket from R97L-Ras-GDP and RhoA-GDP ... 124
Figure 6. Conserved switch II regions in R97L-Ras-GDP, RhoA-GDP and Rop-PRONE ... 126
Figure 7. Superposition of DTE binding pocket between R97L-Ras-GDP and Rop-PRONE complex ... 127
Figure 8. Superposition of K-Ras-GDP (G12D, C118S) and H-Ras-GDP (R97L) at switch II... 129
Figure 9. Superposition of full length K-Ras and R97L-H-Ras at switch II ... 130
Figure 10a. Superposition of full length K-Ras, H-Ras and R97L-H-Ras (1-166) at the allosteric site131 Figure 10b. Coordination of acetate molecule at allosteric site of full length K-Ras ... 132
Figure 1. Amino acid sequence alignment of Raf residues 1-186 in pEt28a vector ... 148
Figure 2. Amino acid sequence alignment of Raf residues 1-186 in Gev2 vector ... 149
Figure 3. Amino acid sequence alignment of Raf residues 52-196 in Gev2 vector ... 150
Figure 4. Amino acid sequence alignment of Raf 52-196 in pET28a ... 157
Figure 5. Amino acid sequence alignment of GB1_Raf52-184 in pET21 ... 158
Figure 6. Amino acid sequence alignment of GST_Raf52-184 in pGEX vector ... 159
Figure 7. Amino acid sequence alignment of N-terminal His-tagged GB1_Raf52-184 in Champion vector ... 160
Figure 8. Phosphorylation levels of ERK in NIH-3T3 cells transfected with wild type and R97L-Ras ... 167
Appendix D: Y32F supplemental PNAS ... 177
Figure S1. Switch I and switch II in the well-ordered active site in Ras-GppNHp ... 184
Figure S2. Switch regions in RasY32F-GppNHp ... 185
Figure S3. Chromatogram of RasY32F-GppNHp and RasY32F-GDP in the presence of Raf-RBD-CRD ... 186
ABSTRACT
Ras is an enzyme that has been studied for three decades. Structural analysis of Ras GTPase has helped to understand the mechanisms of activation and deactivation. This review focuses on the influence of Ras within the Ras/Raf/MEK/ERK pathway. Mutant Ras is a prominent protein found in numerous human cancers. We are still deciphering the various functional roles of Ras at the atomic level, using X-ray crystallography as the primary technique to investigate this protein. It is well known that Ras is a molecular switch: active when bound to GTP and inactive when bound to GDP. However, a more recent discovery is the role of the allosteric site on Ras and in this work, we examine the possible impact the allosteric site has on Ras activation.
INTRODUCTION
RasGTPase and signal transduction pathways
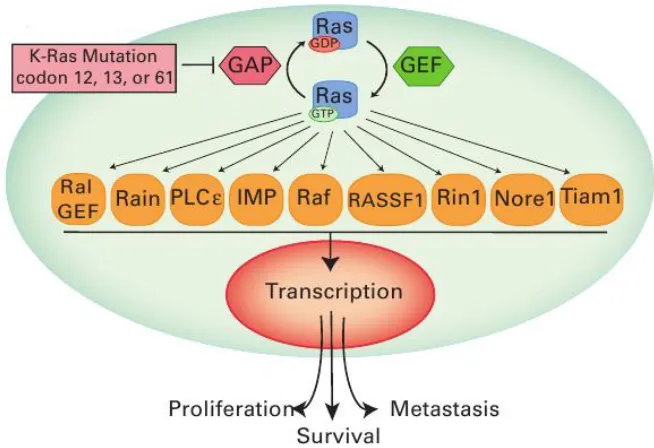
Figure 1: Activation of Ras results in signal transduction of many pathways. Ras has many downstream effectors which activate transcription of genes that induce apoptosis, proliferation, adhesion, and migration. Mutations of Ras at residues 12, 13 and 61 are highly
transforming and discussed in this work. The cytoplasm and nucleus are colored cyan and pink, respectively. Reproduced with permission from Roberts et al, 2010 [4].
GTPase activating proteins (GAPs) bind and inactivate Ras by facilitating the hydrolysis of GTP to GDP, which turns off the phosphorylation cascades regulated by Ras [5]. As explored in this work, Ras also has a low level on intrinsic hydrolysis, converting GTP to GDP without the aid of GAPs [6]. Guanine nucleotide exchange factors (GEFs) bind Ras-GDP and promote the release of GDP. With picomolar affinity for Ras, GTP is present at 10 times higher concentration in the cell; therefore it is likely to replace GDP at the
Figure 2: Ras is a small monomeric GTPase. This figure depicts the activation and deactivation of Ras. GAPs and intrinsic hydrolysis are two mechanisms for hydrolyzing GTP to GDP bound to Ras. GEFs, such as Sos, bind Ras-GDP and promote release of GDP, which
in turn allows for GTP binding and activation of Ras. GTP is present at ten times higher concentration than GDP in the cell while the nucleotide pocket of Ras has picomolar affinity
for guanosine triphosphates. Reproduced with permission from: http://www.munich-interact.org/speakers/alfred_wittinghofer.html
state of the kinase [11].
In eukaryotes, cells communicate using small signaling molecules found in the extracellular matrix. These molecules can be proteins, steroids, amino acids, small peptides, among other molecules [11]. The signals are either secreted from cells by exocytosis or diffusion through the plasma membrane (PM). These molecules can travel either long or short distances to reach their target cell within the multicellular organism [11]. Membrane bound receptors are proteins on the PM of the cell that bind these signaling molecules in order to instruct the cell of a new function. There are three subunits for membrane bound receptors: the amino terminal extracellular domain which contains the ligand-binding cavity; a transmembrane domain inserted into the PM, and a carboxy terminal intracellular domain which experiences a chemical change upon ligand binding [11]. Once the receptor's
cytoplasmic domain is effected, instructions have been received which induce specific functional changes for the cell.
Receptor tyrosine kinases (RTKs) are one type of extracellular receptor [13]. At rest, these proteins are monomers in the PM. RTKs are transmembrane proteins that are kinases which phosphorylate themselves, an event known as autophosphorylation [10]. For these proteins, phosphorylation occurs at tyrosine residues on the cytoplasmic kinase domain. An incoming signaling molecule binds two adjacent RTKs, which induces dimerization and subsequent autophosphorylation of the RTK [14]. Phosphorylated tyrosine residues tend to bind Src Homology region 2 (SH2) domains of other proteins [15].
kinase domain [14]. Growth factors, such as the polypeptide epidermal growth factor (EGF), are the signaling molecules in the extracellular matrix which bind and activate the receptor [15]. Following autophosphorylation, the SH2 domain of the adapter protein Growth receptor binding protein 2 (Grb-2) binds phosphotyrosine residues of the cytoplasmic kinase domain of EGFR [14].
Ras/Raf/MEK/ERK pathway
When Grb-2 binds RTK via its SH2 domain, its Src Homology region 3 (SH3) domain binds the GEF Son of sevenless (Sos) [9]. GEFs act on small monomeric GTPases such as Ras. (figure 3)[8]. Ras has two functional components: an N-terminal GTPase domain and a C-terminal hypervariable region (HPV). A lipid moiety, such as a farnesyl or palmitoyl group, is covalently attached at its carboxy terminus which inserts into the
cytoplasmic side of the PM [16]. Ras is at rest when bound to GDP. When GEF facilitates the dissociation of GDP and binding of GTP, Ras is activated [5]. Ras-GTP associates with classical effectors Rapidly accelerated fibrosarcoma, (Raf) kinase, phosphatidylinositol 3 kinases (PI3K), and Ral guanine nucleotide dissociation stimulator (RalGDS) among other numerous effectors (figure 1) [17]. Many effectors of Ras are kinases which further
Figure 3: Activation of Ras/Raf/MEK/ERK pathway. Growth factors such as EGF bind receptor tyrosine kinases, such as EGFR to promote Ras-GTP activation. Raf is translocated from the cytoplasm to the PM. Upon binding Ras, PAK phosphorylates Raf at
S338 to induce further propagation of the phosphorylation signal to MEK and ERK. Phosphorylated ERK enters the nucleus and induces transcription of genes involved in cell
proliferation and differentiation. The cytoplasm and nucleus are colored cyan and pink, respectively. Reproduced with permission from Roberts et al, 2010 [4].
Figure 4: Diagram of full length cRaf. Three conserved regions of Raf kinase, CR1, CR2, CR3 are seen. CR1 is the amino terminal regulatory region which binds Ras-GTP at residues 51-184. CR2 is a serine/threonine rich region which experiences phosphorylation. CR3 contains the kinase domain which binds ATP and interacts with effectors such as MEK in order to induce phosphorylation of MEK. Full length cRaf has 649 residues. Reproduced
with permission by Bondeva et al, 2002 [22].
The N-terminus is the regulatory region and the C-terminus is the catalytic region of the kinase. CR1 contains the two Ras binding domains: the canonical Ras binding domain (RBD) and the cysteine rich domain (CRD) [23]. The RBD binds the effector region of Ras,
consisting of residues 30-40, which overlaps switch I (residues 26-41). The CRD binds the activator region of Ras, residues 27-29 and 41-45 which flank the effector region on Ras [24]. Both Raf domains must bind Ras to fully activate the Ras/Raf pathway. Once both domains bind Ras, Raf is phosphorylated [25], [26]. This stimulates an association with Map kinase Extracellular-signal Kinase (MEK) [27]. MEK in turn is phosphorylated, which binds and phosphorylates Extracellular signal Regulated Kinase (ERK) [28]. phospho-ERK
migrates into the nucleus and induces transcription of genes that stimulate cell growth, proliferation and division (figure 3) [29], [30]. Aberrant propagation of this phosphorylation cascade leads to tumorigenesis in human cells [31].
Ras-family GTPase Rap1a-GppNHp (PDB ID: 1C1Y) [34]. RBD adopts a ubiquitin-like fold, where two β–sheets, β1 and β2 from RBD form an anti-parallel sheet with β2 and β3 from
Rap1A [34]. When Raf is at rest in the cytoplasm, CRD and CR2 are the locations of the interaction between Raf and 14-3-3 [26], [35],[36]. Once Raf RBD binds activated RasGTP, RafCRD then dissociates from 14-3-3 to bind Ras-GTP. When this occurs, Raf CR2 is dephosphorylated by Protein Phosphatase 2A (PP2A) at serine 259 (S259) [37]. Next, kinase p21rac/cdc42-activated kinase (PAK), phosphorylates Raf at serine 338 (S338) within CR3 [26]. Hence, the activation of Raf along the MEK/ERK pathway is a complex, multi-step process. ATP binding occurs at CR3 of Raf, which contains the catalytic kinase domain. For the mutated isoform of B-Raf, which is known to have increased expression levels in melanomas, the ATP binding pocket has become a target for cancer therapies (discussed in more detail below) [38].
Recent publications suggest the prominent role of scaffold proteins in signal transduction networks [9, 13, 39]. Scaffold proteins bind two or more components of a
a GTP-dependent manner [41]. Research suggests Shoc2 specifically interacts with P34 of Ras [42]. Sieburth et al showed the Ras/RafRBD interaction occurs at E37 and Y40 of Ras effector region. Additionally, with the publication of Ras-GppNHp bound to Raf RBD (discussed in chapter 2 of this dissertation), we see the significance of Ras residues E31, D33, E37, and S39 to interact with RafRBD residues. Levels of phosphorylated ERK and MEK in HeLa cells decrease with the addition of Shoc2-siRNA, Shoc2 indeed influences the activation of this important pathway [43] [41]. The function of Shoc2 was seen to be
enhanced with increased levels of calcium in cells [44]. The various roles of Shoc2 in the cell are still under investigation. For example, one paper concludes that Shoc2 binds Ras-GTP before binding Raf in order to discourage RasGAP’s role to deactivate Ras-GTP [43].
In addition to scaffold proteins influencing phosphorylation pathways, scaffold proteins are themselves subject to regulation. Erbin is a protein with one LRR domain and a carboxy terminal PDZ domain. It functions to bind Shoc2 and inhibit the Ras/Raf interaction [45]. With this role in the cell, Erbin is believed to be a tumor suppressor protein. Erbin also binds unphosphorylated growth factor receptor ERBB2 (also known as HER2), to inhibit activation of the MEK/ERK pathway at the receptor level and prevent tumorigenesis [46]. HER2 is overexpressed in approximately 30% human breast cancers, many of them
Structure of H-Ras
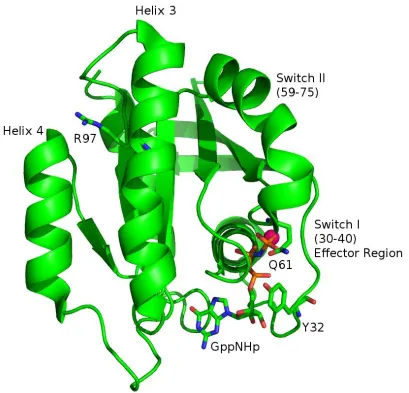
In the Mattos lab, we use X-ray structural analysis to better understand the function of Ras in the cell. Human Ras enzyme has three isoforms: H-, K-, and N-. For our research, we use H-Ras. As is the case for cRaf, H-Ras is the isoform most commonly used in research laboratories. Virtually all available biochemistry and structural biology experiments on Ras have been done using H-Ras due to its high expression in the soluble fractions in E. coli cells and good behavior during purification protocols. Figure 5 shows the structure of the GTPase domain of wild type H-Ras (PDB ID: 3K8Y), consisting of residues 1-166. Ras is a
Figure 5: Structure of H-Ras (PDB ID: 3K8Y), residues 1-166, in the R32 space group. Ras consists of five α-helices, six β-sheets and 10 loops. For this structure, Ras is bound to the slowly hydrolyzing nucleotide GppNHp, to mimic the GTP-bound state. The magnesium ion
is colored magenta. Switch I overlaps the effector region which binds GAPs, GEFs and kinase proteins. Switch II is ordered in this structure. The conformation of the switches is dependent on nucleotide state. The conformation of the switch regions facilitate or hinder binding of various proteins to Ras. Highlighted in this figure are residues Y32 on switch I, Q61 on switch II, and R97 of Helix 3. These residues are discussed in detail in subsequent
chapters.
state of the bound nucleotide [49]. Switch I consists of residues 26-41. This overlaps with the effector region, residues 30-40 [1]. Originally, Ras was crystallized in the P3221 space group, where Y32 adopts an “open” conformation, mimicking the Ras-RasGAP complex (PDB ID: 121P). This conformation of switch I is associated with “state 1” determined by 31P NMR [50], [51]. We see in figure 5 the proximity of switch I to the bound nucleotide. Highlighted in this figure is residue tyrosine 32 (Y32) of switch I. In this structure, Y32 is in the “closed” conformation, observed when Ras is bound to the effector Raf. This is also known as “state 2” determined by 31
P NMR [51]. The Mattos lab was the first to publish a crystal structure of wild type Ras in the R32 space group, where Y32 is in the closed (state 2) conformation [52], where it mimics the Ras-Raf interaction. NMR experiments have shown that when Y32 is in state 1, Ras has low levels of intrinsic hydrolysis compared to state 2. Y32 will be discussed when analyzing the active site of the structures solved in this work in chapters 2 and 4. Figures showing both states 1 and 2 are seen in chapter 4 when discussing the structure of R97L-Ras-GppNHp. Appendix D has X-ray structural details of the Ras-GppNHp mutant Y32F which explores the Role of Y32 and Ras' intrinsic GTP hydrolysis when the hydroxyl group is absent.
state of Ras [55], [56]. Raf is one of the few binding partners of Ras that does not interact with switch II [57], [58], [59, 60]. Switch II contains the catalytic residue glutamine 61 (Q61), highlighted in figure 5. This is another residue that is pertinent to this work and is discussed throughout the chapters.
Overall, Ras has five α-helices, six β-strands and ten loop regions [50], [61]. Helix 3
is adjacent to switch II and contains residue arginine 97 (R97), highlighted in figure 5. Originally, calcium chloride was present when crystals formed in the R32 space group [62]. This resulted in a disordered switch II region. Then, our published Ras structure with PDB ID: 3K8Y was obtained from crystals formed in the presence of calcium acetate. A calcium ion binds between loop 7 and helix 4 and coordinates an acetate molecule bound at R97, at the C-terminus of helix 3. We hypothesize that the acetate mimics a negatively charged group in the lipid bilayer, as this part of the Ras protein has been shown to interact with the
membrane [63], [48], [64]. The bound calcium acetate induces a shift of helix 3 / loop 7 (residues 98-108) away from switch II. We have termed this region the allosteric site. The implications of this allosteric site shift on hydrolysis are discussed in the section called “Ras Intrinsic Hydrolysis” below and experiments in Chapter 4 expand on our knowledge of Ras’
mechanism of intrinsic hydrolysis.
coordinate the magnesium ion. In chapter 4 of this work, a Ras mutant (R97L) bound to GDP has a disordered switch I which includes T35. Interestingly, the magnesium ion is absent from this structure.
Figure 6: A magnesium ion is hexacoordinated in Ras-GppNHp. The positive charge of the magnesium ion (yellow) neutralizes the negative charge of the phosphate groups of the
nucleotide . The Mg2+ is coordinated by one oxygen (red) from the γ- and β- phosphate groups, the hydroxyl oxygen atom (red) from T35 and S17 as well as two conserved water
Ras/Raf Signaling and Cancer
Table 1. Occurrence of Ras and B-Raf in human cancer
explored in the laboratory [66]. In figure 7a, we see the sequence alignment of full length human K- and H-Ras.
Figure 7a. Amino acid sequence alignment of human K and H-Ras. Note the 95% GTPase domain (residues 1-166) sequence identity between the two isoforms. Each line contains 60
residues. MutantK-Ras has been implicated in many human cancers. Mutant H-Ras is less prevalent in tumorigenesis. However, H-Ras is expressed primarily in the lab and results
from X-ray crystallography of H-Ras are used to explain mechanism of K-Ras and malignancy in cells. Web site used for sequence alignment:
http://www.ebi.ac.uk/Tools/msa/clustalw2/
position 106. T/S difference at 126, and K/R difference at 127. The C-terminal hypervariable region (HVR) contains most of the differences among the isoforms, retaining only 15% identity within residues 167-189 [16].
When specifically performing a structural analysis of the G-Domain between the isoforms K- and H-Ras, the backbones are quite similar with a root mean square deviation (RMSD) of 0.42Å (figure 7b) This makes H-Ras an excellent model for a structural analysis of the GTPase domain of oncogenic K-Ras, which contains the switch regions, the guanine nucleotide binding site, the active site and the allosteric site. When including the HVR, the overall sequence identity between the two isoforms is reduced to 86%. However, for
structural analysis, we exclude the HVR (residues 167-189). We see in figure 7c, the effects of different HVR has on lipid bilayer location in the cell. While both isoforms are
carboxymethylated at the C-terminal cysteine following the removal of the last three
Figure 7b. Superposition of K-Ras (raspberry, 3GFT) and H-Ras (yellow, 2RGE) bound to GppNHp. There is good agreement between the backbone conformation of residues and
nucleotide placement from K- and H-Ras. RMSD between these isoforms is 0.42Å, measuring main chain Cα carbons 1-166 while omitting residues 59-75 of switch II which are
known to have high levels of disorder. This makes H-Ras an excellent model in the lab for studying oncogenic K-Ras mechanisms. No publication is associated with the crystal
Figure 7c. Diversity of amino acid sequences within the hypervariable region at carboxy-terminus of H-, N-, and K- Ras isoforms. K-Ras is farnesylated and carboxymethylated (red).
The positively charged poly-K chain is attracted to the polar headgroups of the lipid bilayer of the PM thereby encouraging membrane association. H-Ras experiences farnesylation and double palmitoylation (blue). The dissimilarity of HVR explains the difference in membrane localization of the Ras isoforms within the cell. Reproduced with permission from Hancock
and Parton, 2005 [16].
Structural Analysis of Ras and Cancer Drug Therapies
The Ras/Raf pathway is implicated in numerous human cancers [30]. Structure based drug design (SBDD) has become a prevalent technique for developing non-classical
and tumor lysis syndrome (TLS), a rapid breakdown of tumor cells which overload the circulation system with purines and uric acid, have resulted in the untimely fatality of some patients [75], [76]. Therefore, more work needs to be done on personalized cancer therapies [77], [78].
The first structure of Ras in complex with another protein was the structure of Ras bound to GAP-334 (1WQ1), a Ras-GAP. In this structure, the nucleotide bound to Ras was GDP. Aluminum fluoride was present in the crystallizing conditions, as it serves as a good transition state mimic of the planar γ-phosphate group of GTP during the hydrolysis reaction
[55]. With the publication of this structure, we saw that GAP interacts with Ras at both switch I and switch II. Y32 is in the open conformation (state 1), which allows room for R789, called the arginine finger, to insert into the active site (figure 8). This structure revealed that the terminal oxygen atom of residue Q61 is positioned to directly interact with the nucleophilic water while the terminal nitrogen atom interacts with a fluoride atom and backbone oxygen atom of R789 from GAP. It was evident that the position of Q61
Figure 8: Active site of Ras-GDP-AlF3-GAP-344 complex (1WQ1). Wild type Ras is colored deep salmon. GAP-334 is colored grey. Highlighted are residues Y32 from Ras, in the open conformation (state 1), which allows for insertion of R789. R789 from GAP-334 is named the “arginine finger”. Dashed lines represent the coordination of AlF3, R789, Q61 and
A year later, the structure of Ras bound to the GEF Sos (PDB ID: 1BKD) was published. From this structure, we see again switch I and switch II are the region of interaction with this binding partner [56]. There is no guanosine nucleoside or magnesium ion present in this complex. This second Ras complex structure revealed a hydrophobic pocket, consisting of residues L56, Y71, M67 and Y64 on switch II that directly interact with H911 and Y912 on helix G of Sos. Two recent independent publications by Genetech and the Fesik group at Vanderbilt University demonstrate that this hydrophobic pocket is still a hot target for cancer drug design if the tumorigenesis is induced by propagation of the Ras signaling pathway [79]. In the Genentech paper, the structure of the small molecule 4,6-dichloro-2-methyl-3-aminoethyl-indole (DCAI) bound to full length K-Ras shows binding at the Ras-Sos hydrophobic interface. Experiments in HEK-293T cells show that the presence of DCAI reduced the recruitment of a construct of Raf containing both the RBD and CRD domains to Ras at the PM. They further showed that when the cells were inundated with EGF in the presence of DCAI, Ras expression levels increased, however there was no increase in Ras signaling in the cell, as measured by the presence of low levels of phosphorylated ERK. In the Fesik publication, they noted the side chain of tyrosine 71 (Y71) within switch II takes a “closed” conformation when Ras has no bound ligand. Upon binding of
N-[2-(1H-indol-3-ylmethyl)-1H-benzimidazol- 5-yl]-L-prolinamide (0QY), Y71 shifts to an “open”
The inhibitors DCAI and 0QY discussed above bind at switch II. GAPs, GEFs and effectors such as PI3K and RalGDS bind at switch I and II [81]. Recall Raf kinase does not bind switch II. It binds Ras at switch I and the activator domain as discussed previously [57]. Therefore these drugs do not directly disrupt the Ras/Raf interaction. The only drug thus far that disrupts the Ras/Raf interaction is methylcyclopropane-1 (MCP-1) and its derivatives MCP-53 and MCP-110 [82]. This inhibitor was originally discovered by yeast two-hybrid screen. More recently, MCP-110 was shown to physically disrupt the Ras/Raf interaction in mammalian NIH-3T3 cells [83]. In this study, phosphorylation levels of ERK decreased in a dose-dependent manner to MCP-110. It was also found that MCP specifically interacts at switch I of Ras, disrupting the RafRBD association, not the Ras-CRD interface, which interacts with Ras' activator region (residues 27-29, 41-45, flanking switch I). With the solved structure of Ras bound to Raf_RBD_CRD, as attempted in this work, SBDD can possibly be utilized to find a small molecule that also disrupts the Ras-Raf_CRD interaction [84]. Interestingly, a recent development in cancer treatment is the combination of multiple small molecules that inhibit protein-protein interactions at different points along the signaling pathway. For example, a study showed that using the drug Lonafarnib in conjunction with Sorafenib considerably reduced the number of melanoma cells [85].Lonafarnib is a farnesyl-transferase inhibitor. Figure 7c shows the farnesylation of Ras which allows for insertion into the PM. Sorafenib, described earlier, binds the ATP pocket of B-Raf and disrupts the
phosphorylation of MEK by Raf. Therefore, with the solved structure of Ras bound to the CRD of Raf kinase, another point of intersection can be targeted for cancer drug design.
not been solved. For full activation of the Ras/Raf/MEK/ERK pathway, two Raf domains must bind Ras for catalytic activity of Raf's kinase domain. [86]. The RBD binds first, with an affinity of 3.5 nM for Ras, and binds switch I in a GTP dependent manner [87]. The CRD binds the activator region of Ras, with lower affinity [57], [84]. The CRD coordinates two zinc atoms [88]. If the CRD does not bind zinc, activation of the Ras/Raf pathway is hindered [24]. In NIH-3T3 Ras-transformed cells, Bondeva et al showed that a green fluorescent protein (GFP)-tagged construct of Raf-RBD alone will not translocate to the PM nor bind Ras when the cells are activated with platelet-derived growth factor (PDGF) [22]. Only the GFP-tagged Raf construct containing both the RBD and CRD is recruited to the PM and binds Ras. Therefore, to perform a complete structural analysis of Ras activation by Raf, both the RBD and CRD regions of Raf must be present in the solved structure. In a separate pull down assay, the RBD alone did associate with Ras in vitro, however more Ras was present using both the RBD and CRD in the construct of Raf [22]. Furthermore, work from the Campbell lab has shown that the association of the CRD is dependent on farnesylation of Ras [89], [90]. The effect of this post-translational modification of Ras on the Raf-CRD
interaction is discussed further in chapter 3.
Thus far, the structure used to analyze the Ras-Raf interaction is the structure of Rap, a Ras-related GTPase with overall 50% sequence identity to Ras, bound to the RBD of Raf kinase (PDB ID: 1GUA) [91]. In this structure, two mutations, (E30D, K31E) in switch I make the effector region of Rap 100% identical to that of Ras. This double mutant of Rap has been termed “Raps” In this Raps-RafRBD structure, we saw that RBD has a ubiquitin
state 2. The hydroxyl group of Y32 is coordinated to an oxygen atom of the γ-phosphate group via a bridging water molecule. Unfortunately, Rap has a T61 in place of Q61, the catalytic residue, on Ras. Despite the fact that T61 interacts with the bridging water
molecule, it is difficult to perform a full structural analysis and obtain information regarding the role of residue 61 in the mechanism of intrinsic hydrolysis for Ras when bound to Raf RBD from this structure.
We also see that the interactions between Raps and RafRBD are primarily
electrostatic. For example, R89 of RafRBD interacts with S39 of Raps. Previous mutational studies confirmed that the R89L mutant of Raf completely abolishes the Ras-Raf interaction [92]. Also, K84 of RafRBD is coordinated to E31 and D33 of Raps. And finally, R59 of RafRBD interacts with E37 of Raps. Note that all the interactions from Raps are contained in the switch I region. These electrostatic interactions will be compared to the solved structure of wild type Ras-GppNHp bound to Raf RBD in chapter 2.
Intrinsic Hydrolysis
GAP-catalyzed hydrolysis and intrinsic hydrolysis are the two modes of deactivation of Ras. In the X-ray structure of Ras bound to GAP-334 (discussed previously), R789 from GAP and Q61 from Ras are involved in GTP hydrolysis (figure 8) [55]. Prior to the
GAP. Wild type Ras has a slow rate of intrinsic hydrolysis, yet oncogenic mutants valine 12 (V12) and leucine 61 (L61) of Ras have decreased levels of hydrolysis compared to wild type [94] [6]. With the GAP-catalyzed mechanism of Ras hydrolysis well studied, much was left unknown about a mechanism of intrinsic hydrolysis and its role in tumorigenesis.
Our lab has explored the role of intrinsic hydrolysis catalyzed by Ras. We have successfully solved the structure Ras-GppNHp from crystals with symmetry R32 in which the active site of Ras is as observed in the Raps/Raf complex [48]. Based on these structures, two water molecules appear to be involved in the hydrolysis of the GTP γ-phosphate: the nucleophilic water molecule and the bridging water that connects Y32 to the γ-phosphate of
GTP. We propose that a proton is shuttled from the nucleophilic water to the oxygen (O1G) γ–phosphate of GTP and from there to the bridging water molecule which in turn associates with residues Q61 and Y32. This results in a partial positive charge in the vicinity of GTP β-γ
bridging oxygen atom that could stabilize the dissociative-like transition state known to occur during catalysis [95]. We hypothesized that this mechanism is regulated by an allosteric switch in the presence of Raf but not other effectors and have recently shown a directly link between allosteric modulation and the Ras/Raf/MEK/ERK pathway in NIH-3T3 cells [6]. Interestingly, for L61 and V12 Ras, two well established oncogenic Ras mutants, the bridging water is absent [96], [97], [3]. Instead, for L61 a hydrophobic cluster separates the γ–
phosphate from any water molecules. L61 Ras does not detectably hydrolyze GTP in the presence of Raf_RBD_CRD [6]. For V12 Ras, steric hindrance obstructs water from
mutations is the same, therefore hydrolysis of the nucleotide must dictate the dissociation rate [23]. We are presently deciphering the contrast between wtRas and oncogenic L61 and V12 hydrolysis mechanisms to explain the transforming efficiencies of the mutants within cells.
From these findings of the hydrolysis mechanism, we can infer the events for intrinsic hydrolysis when Ras is bound to Raf. Recall Ras-GAP has µM affinity for Ras-GTP, as does PI3K and RalGDS. Raf, on the other hand, has a higher affinity (nM) for Ras-GTP.
Therefore, the specific dissociation of Ras and Raf are unlikely to occur with the
involvement of GAPs. We hypothesize that once Ras binds Raf, the allosteric site is activated by association with a negatively-charged phospholipid. Since Raf does not bind switch II, ordering of switch II is free be effected by the allosteric signal. We see in the structures of Ras in the R32 space group, this renders an ordered active site, where residue 61 is poised to interact with the bridging water, not the nucleophilic water as seen in the Ras-GAP334 interaction. When Ras is bound to Raf, Q61 has a direct role in catalysis, aiding in the intrinsic hydrolysis of GTP to GDP. As a result of the change in nucleotide bound to Ras, the switch regions change conformation. Switch I, which overlaps the effector region, now takes a conformation in which the binding of Raf is no longer favored and dissociation of Raf occurs.
Goals
dissertation. Six new crystal structures are presented throughout chapters 2, 3 and 4. Another structure, Y32F-Ras-GppNHp is discussed in Appendix D.
The questions addressed specifically in this work investigate the Ras/Raf interaction at both the active site and the allosteric site. To this end, I have purified and grown crystals of Ras GTPase in complex with a construct of Raf kinase that contains both the RBD and CRD regions. The objective of this work was to attain the structure of Ras bound to Raf kinase. This work is addressed in chapters 2 and 3. We have attained the structure of wild type Ras and oncogenic Q61L-Ras bound to Raf_RBD with the CRD disordered and unsolvable at this stage. The ultimate goal of this project is to fully solve the structure of the CRD's interaction with Ras. The work for this project will be continued by fellow Mattos Lab members. Upon completion of the structure of Ras bound to Raf-CRD, computational analysis in search of an inhibitory cancer drug can be performed [98].
Another goal of this work is to further elucidate the mechanism of the allosteric switch and intrinsic hydrolysis of Ras. This aspect is discussed in chapters 2 and 4. We see that for the Ras/Raf complex, as expected, the allosteric switch is in what we call the “on” state where Q61 is positioned for a direct role in GTP hydrolysis. We also see that when polar residue R97 is mutated to the hydrophobic residue leucine, the allosteric switch of Ras is shifted to the “on” state. This shift is induced by hydrophobic interactions between helix 3
presence of aluminum fluoride in solution, R97L-Ras-GDP binds Ras_RBD, forming a transition state mimic. This is the first capture of a transition state mimic associated with intrinsic hydrolysis in Ras or any small GTPase.
The appendices contain the extensive mutagenesis work necessary to achieve the results founded in this publication. Cloning and PCR of Ras and Raf are explained in
Appendix A. The purification of the Ras/Raf complex is explained in Appendix B. Appendix C shows the mr_rosetta commands in Phenix used to attempt to fit the CRD to the Ras/Raf data. And finally, Appendix D explains the results from the Y32F-Ras-GppNHp mutant work.
ACKNOWLEDEMENTS
CHAPTER 2:
ABSTRACT
Ras is a GTPase critical in the Ras/Raf/MEK/ERK signaling cascade, a pathway upregulated in many human cancers. When Ras binds Raf, two Raf domains are involved in the interaction: the canonical Ras Binding Domain (RBD) and the Cystein Rich Domain (CRD). Structural analysis of the Ras/Raf interaction has remained elusive over the 20 years since the discovery of this association, with the homologous Raps/Raf-RBD structure providing the best model available. Although computational models of the Ras/Raf-RBD interaction have been developed, there is limited information on the active site of Ras when bound to Raf. In this chapter, we present the structure of wild type Ras-GppNHp and
oncogenic Q61L-Ras-GppNHp bound to Raf kinase. For the wild type Ras complex, we see an ordered active site where the catalytic residue Q61 plays a direct role in intrinsic
hydrolysis of Ras while bound to Raf. For the Q61L-Ras complex, we observe a cluster of hydrophobic residues over the active site. As seen in the previously published crystal structure of uncomplexed Ras-GppNHp with a bound calcium acetate at a remote site, the allosteric switch at helix 3 / loop 7 in the complex promotes ordering of the active site.
INTRODUCTION
effectors [23] [57]. Raf is recruited to the PM by activated Ras and once phosphorylated, Raf in turn phosphorylates Map kinase Extracellular-signal Kinase (MEK), which phosphorylates Extracellular signal Regulated Kinase (ERK) [101]. Phosphorylated ERK translocates into the nucleus and induces transcription of genes that cause cell growth and proliferation [29]. This is the key element that causes malignant cell growth when the Ras/Raf/MEK/ERK pathway is deregulated. Ras binding Raf is a fundamental step along this signaling cascade that plays a critical role in the fate of the cell.
Mutant Raf, Rapidly accelerated fibrosarcoma, is a kinase implicated in the
independent of the RBD [57], [107]. The binding of CRD is not dependent on the nucleotide-bound state of Ras, and its affinity is lower than that of RBD. Mutations in switch I of Ras did not disrupt CRD binding [57]. Binding of the CRD to Ras appears to be dependent on farnesylation of Ras [89, 90]. When the CRD is mutated and does not bind Ras, inhibition of Raf activation is observed, with accompanying decrease in phosphorylation levels of MEK and ERK [89], [108].
Ras signaling is known to be prominent in approximately 30% of human cancers [109]. The most common occurrence of mutational Ras is found in pancreatic cancer, which is also the most lethal form of human cancer [66]. To this end, Ras has become a hot target for drug design as a form of personalized chemotherapy [79]. The solved structures revealing protein-protein interactions lend themselves to computational analysis for discovery of small molecules that inhibit this pathway. Structure based drug design (SBDD) has become a popular method to discover new anti-cancer drugs [70].
slow compared to Ras [111].
For Rap, mutational studies of residue 61 show no direct involvement of T61with hydrolysis [112]. Therefore, even in the presence of computational models of the Ras/Raf-RBD interaction, we are left without a comprehensive understanding of the active site of Ras when bound to its effector Raf [113], [81]. This structure is not ideal for exploring the
mechanism of intrinsic hydrolysis of Ras when bond to Raf.
Recently, our lab has explored the role of intrinsic hydrolysis catalyzed by Ras. We have successfully solved the structure Ras-GppNHp from crystals with symmetry R32 in which the switch I of Ras is as observed in the Raps/Raf complex [62], [48] . Based on these structures, two water molecules appear to be involved in the hydrolysis of the GTP
γ-phosphate: the nucleophilic water molecule and the bridging water that connects Y32 to the γ-phosphate of GTP. We propose that a proton is shuttled from the nucleophilic water to the oxygen (O1G) γ–phosphate of GTP and from there to the bridging water molecule which in
turn associates with residues glutamine 61 (Q61) and tyrosine 32 (Y32). This results in a partial positive charge in the vicinity of GTP β-γ bridging oxygen atom that could stabilize the dissociative-like transition state known to occur during catalysis [95]. We hypothesized that this mechanism is regulated by an allosteric switch in the presence of Raf but not other effectors and have recently shown a direct link between allosteric modulation and the Ras/Raf/MEK/ERK pathway in NIH-3T3 cells [6].
state of the allosteric switch, with helix3 / loop7 shifted toward helix 4 and switch II ordered for catalysis. We see that for wild type Ras bound to Raf kinase, the active site is ordered and Q61 is poised for a role in intrinsic hydrolysis. For the L61 mutant complex, as expected we see a cluster of hydrophobic residues over the active site, which hinders the occurrence of intrinsic hydrolysis. This is expected to prolong the association of Raf and Q61L-Ras, resulting in an enhanced activation of the Ras/Raf/MEK/ERK pathway.
EXPERIMENTAL PROCEDURES
Wild type and mutant Q61L-Ras were expressed as truncated versions, residues 1-166 as previously published [62]. Purification and loading of GppNHp have also been published [62].
Raf_RBD_CRD purification and Ras/Raf crystallization: Human cRaf kinase residues 52-196 with N-terminal B1 immunoglobulin-binding domain of streptococcal protein G (GB1 domain) in Gev vector was a gift from Sharon Campbell, UNC Chapel Hill. A new Raf construct that consisted of residues 52 to 184 was cloned from that construct for the experiments performed in this work as follows: GB1_Raf_RBD_CRD was cloned into the EcoRI and BamHI restriction sites on Champion pET302/NT-His vector (Invitrogen) by Genewiz (South Plainfield, NJ, USA). More details of this construct is found in Appendix A. GB1 is a fusion protein known to increase solubility and expression levels of difficult
induction. At 4 °C cell paste was resuspended in Buffer A (20 mM HEPES, pH 7.0-7.1, 20 mM NaCl, 20 μM ZnCl2 , 5 mM MgCl2 5% glycerol 10mM DTT) containing protease
inhibitor cocktail. Nucleic acids were precipitated with benzonase nucleases (EM Industries). Elution Buffer B was identical to Buffer A with 1.0 M NaCl. Protein was run over HiPrep SP_FF 16/10 column (GE Healthcare) and eluted with gradient of 0-60% B. GB1_Raf-RBD-CRD purity was verified with SDS-PAGE gel. Fractions were immediately pooled. Promptly, 0.9-1.1 molar equivalent of H-Ras-GppNHp was added to Raf solution. Ras/Raf complex was then concentrated to contain 2mg/mL of Raf to ensure proceeding proteolysis step. A thrombin-cleavable linker is present connecting the C-terminus of GB1 and N-terminus of Raf. Concentrated complex was dyalized against 20 mM Tris (pH 8.1), 100 mM NaCl, 25 mM CaCl2, 5 mM MgCl2, 20 μM ZnCl2, 1 mM DTE for 4 hours at 4 °C. Complex was then dialyzed into fresh thrombin media at 18 °C. Thrombin (1U / μL) was added at 20 Units thrombin per mg of total Raf. Reaction went for 18 h, 18 °C. Proteolysis was arrested by dropping the temperature to 4 °C for 4h. Protein was concentrated to 1 mL for size exclusion column purification (50 mM HEPES, pH 7.2, 50 mM NaCl, 10 mM MgCl2 , 5% Glycerol, 1 mM DTE, 10 μM ZnCl2) Ras-Raf fractions were checked for purity with SDS-PAGE gel.
Fractions were pooled and concentrated for final concentration of 10-16 mg/mL of complex. Initial crystals were grown at 18 °C using the vapor diffusion, sitting drop method by mixing 1 μL Ras/Raf and 1 μL reservoir, and with 90 μL reservoir solution. Reservoir consisted of
Hampton Research Crystal Screen Hit 46 (200 mM calcium acetate, 100 mM sodium cacodylate pH 6.5, 18% PEG 8000.) Subsequently, crystals were grown using vapor
and with 400 uL reservoir solution. Crystals of wtRasRaf have a hexagonal morphology with one axis longer than the other two, measuring approximately 0.05 x 0.01 x 0.01mm (figure 1a). Q61L-Ras/Raf crystals are also hexagonal; however their dimensions are approximately 0.02 x 0.01 x 0.01 mm (figure 1b). Both complexes crystallized in space group P321 with the twin law -h, -k, l. Hundreds of crystals were screened.
Figure 1a (left). Initial crystals of wtRasRaf complex. Figure 1b (right). Initial crystals of Q61L-RasRaf complex. Crystals for both wild type and mutant complex crystallized in the
same condition (200 mM calcium acetate, 100 mM sodium cacodylate, 18% PEG 8000), 18C°. Crystals took approximately 2 to 3 weeks to form. Initial wild type complex crystals
were grown by sitting drop. Initial Q61L complex crystals grew by hanging drop vapor diffusion.
Data Collection and structural refinement. For initial wild type Ras-Raf crystals, X-ray
Our previously published model with PDB ID: 3K8Y was used for molecular replacement of Ras and the RafRBD domain was phased with the structure of Raf-RBD taken from the Raps/Raf-RBD complex (PDB ID: 1GUA). Unsuccessful attempts were made using PDB ID: 1FAR to fit the CRD with PHENIX AutoMR (advanced interface) where the Ras and RBD were held fixed and CRD was the searchable component. When CRD coordinates were obtained, MR_Rosetta was used for further fitting into the weak electron density map [118], [119]. At this stage, no solution for the CRD has been found with these crystals. Attempts to fit the CRD are discussed in chapter 3. For Q61L-Ras-Raf, initial X-ray diffraction data sets were collected at APS, SER-CAT beamline ID-22. Analysis of this data set by the UCLA diffraction anisotropy server indicate that the diffraction along the c* axis was superior than the other directions.
(http://services.mbi.ucla.edu/anisoscale/) [120]. On the basis of an F/σ(F) cutoff of 3 along each reciprocal space axis, reflections were subjected to an anisotropic truncation with resolution limits of 3.3, 3.3 and 3.2 Å along a*, b* and c* axis, respectively, before use in refinement. We report this structure to an overall resolution of 3.3 Å. Coordinates from wild type Ras-Raf
Table 1. Data collection and refinement statistics
Crystal Code 993 1642
Structure wtRas-Raf-RBD Q61L-Ras-Raf-RBD Data Collection
Space group P321 P321
Temperature 100K 100K
Cell dimensions
a, b, c (Å) 90.44 90.44 92.703 91.4 91.4 93.11 α, β, ɣ (°) 90 90 120 90 90 120 Resolution (Å) 50-2.45 (2.48-2.44) 50-3.3 (3.31-3.25) Rsym or Rmerge 0.146 (.649) .179 (.477)
I/σ 2.0 (25) 1.5 (26)
Completeness (%) 97.6 (74.1) 87.4 (55.6)
Redundancy 8.9 (3.5) 4.6 (3.4)
Refinement
Resolution 32.368-2.446 32.613-3.249
No. reflections 16218 6462
Rwork / Rfree 19.75% / 24.80% 24.61% / 30.54% No. atoms
Protein 1861 1861
GppNHp 32 32
waters 100 25
Mg 2 2
Ca 1 0
B-factors
protein 53.55 53.14
nucleotide 33.32 37.57
Water 49.66 27.97
Mg 29.46 22.53
Ca 58.42 ---
R.m.s. deviations
Bond lengths (Å) 0.009 0.008
Bond angles (°) 1.6 1.4
RESULTS
Complexes of wtRas-GppNHp and Q61L-Ras-GppNHp bound to cRaf_RBD_CRD (hereafter called Raf) crystallized in the space group P321 with unit cell dimensions 90.440 x 90.440 x 92.703 Å and 91.400 x 91.400 x 93.110 Å respectively. Each asymmetric unit contained a single Ras/Raf complex. The electron density maps for both complexes reveal well-resolved features for Ras, nucleotide and Raf RBD. Electron density for CRD however is not present. The resolution of wtRas-Raf is 2.45Å and Q61L-Ras-Raf is 3.3Å. An exhaustive attempt to crystallize G12V-Ras-Raf complex was also screened, however crystals never formed of this mutant Ras complex.
The critical impediment of structural analysis of Ras bound to a construct of Raf containing both the RBD and CRD has been the outstanding insolubility of the CRD. We overcame this challenge firstly by using a construct of Raf with the fusion protein GB1 attached at the N-terminus. GB1 is a 56 amino acid fusion protein of the B1 immunoglobulin-binding domain of streptococcal protein G from the GEV vector system [114]. Due to GB1 small size and stable fold, it does not interfere with the interaction of Ras-Raf as previously reported in binding studies from our lab [62]. Secondly, once Raf was purified using cation exchange, we
immediately added 0.9 – 1.1 molar equivalent Ras-GppNHp to the Raf solution. With no Ras present or a less than equimolar amount of Ras in solution with Raf, Raf consistently precipitated out of solution (verified with SDS-PAGE gel, unpublished results).
General features of Ras bound to Ras Binding Domain (RBD) of Raf kinase
Figure 2. Superposition of wtRasRaf and oncogenic Q61L-RasRaf. Wild type Ras complex (dark red) has an overall good agreement with Q61L-Ras complex (pink). Magnesium ions are see in
yellow and magenta for wild type and Q61L-Ras complexes respectively. For both structures, Ras is allosterically on. Raf RBD has a ubiquitin superfold, as seen in the Raps/Raf structure. Raf
interacts at the switch I region of Ras.
additional electron density representing CRD was identified.
Figure 3. wtRasRaf complex is allosterically “on”. Superposition of wild type Ras-GppNHp (PDB ID 3K8Y, green) in the R32 space group with calcium acetate bound at helix 3 /
loop 7 and complex of wild type Ras-GppNHp bound to RafRBD (dark red). Note the shift of helix 3 / loop 7 of the Ras complex away from switch II. This shift induces an ordering of the active site and places catalytic residue Q61 for a direct role in hydrolysis when Ras is bound to Raf. The wild type Ras/Raf complex has a calcium ion coordinated at R97. RMSD for residues 98-108 for these structures is 0.6Å. RMSD calculated for 3K8Y and Q61L-Ras/Raf complex for
residues 98-108 is 0.74Å.
Both wild type and oncogenic complexes are allosterically “on”, with helix 3 / loop 7
(2RGE) of the allosteric switch. As we have previously shown, helix 3/loop 7 needs to be shifted away from switch II in order to avoid clashes between side chains of Val 103 on helix 3 / loop 7 and Met 67 on switch II.
Figure 4. A calcium ion is hexacoordinated near R97 of wtRasRaf. The calcium ion found in the Ras/Raf complex is coordinated by four water molecules (red), the oxygen atom from the main chain of D107 and the oxygen atom from the main chain of Y137. Despite the presence of calcium acetate in the crystallizing conditions, electron density for an acetate ion is not
seen. R97 is within hydrogen bonding distance to one water molecule coordinating the calcium ion. The calcium ion, however, is not directly interacting with R97. Electron density
Figure 5. Active site of wtRasRaf. Superposition of wild type Ras/Raf (dark red) and wtRas bound to calcium acetate (green, 3K8Y). Both bridging (189) and catalytic (175) waters are present in the complex structure. Side chain oxygen atom of Q61 is 2.8Å from bridging water molecule in the new Ras/Raf structure. This agrees with the 2.8Å distance between the side chain
oxygen of Q61 and the bridging water molecule of 3K8Y, placing it in the pre-catalytic conformation for hydrolysis. Dashed lines denote atoms are within hydrogen bonding distances.
Figure 6. Hydrophobic pocket of active site in Q61L-Ras-GppNHp-RafRBD structure. A hydrophobic pocket is seen in the active site of the oncogenic mutant Q61L-Ras bound to RafRBD. This promotes a reduction of intrinsic hydrolyisis, thereby increasing the time of the Ras/Raf association. The prolonged propagation of the Ras/Raf/MEK/ERK pathway is known to
cause tumorigenesis. Electron density contoured at 1σ.
Q61L-Ras bound to Raf could be a limitation of the resolution of this complex structure (3.3Å). The conformation of L61 in the Ras-Raf complex agrees with the placement of L61 in the higher resolution structure of Q61L-Ras in the R32 space group.
Figure 7. Active site superposition of RafRBD complex with Ras-GppNHp in the R32 space group. The catalytic water molecule (175) is seen in the Q61L-Ras structure (3OIU, cyan) with a calcium acetate bound at the allosteric site. This structure
With this direct comparison of active sites for the structure of Q61 and L61 Ras bound to Raf RBD, a deeper understanding of the mechanism of intrinsic hydrolysis and its influence on the activation of the MEK/ERK pathway is coming into focus. We see from the structure of Ras bound to Raf RBD, Q61 interacts with the bridging water molecule poised to participate in intrinsic hydrolysis catalyzed by Ras while bound to Raf. We also see with the structure of Q61L-Ras-GppNHp bound to Raf kinase, a hydrophobic cluster excludes water from the active site, which in turn severely impairs intrinsic hydrolysis of Q61L-Ras bound to Raf. This can possibly explain the high transforming capability of cells found with Q61L-Ras present.
Structural comparison of wtRas-GppNHp/Raf to Raps/Raf:
Previously, the structure used to model Ras bound to Raf has been the structure of Rap, a member of the Ras superfamily that has 57% overall sequence identity in the G-domain [109]. To create 100% sequence identity at the effector region, mutations E30D and K31E make up the “Raps” effector region. This was solved in the space group P21 21 21 and to date, this has served
Figure 8. Superposition of wtRasRaf complex and Raps-RafRBD. As expected, good backbone agreement is seen with the overall fold. wtRasRafRBD is dark red and Raps-RafRBD is magenta (1GUA). Electrostatic interactions from switch I of Raps to RafRBD were retained in
wtRasRaf. As explained in the text, there were some differences between these structures.
For example, the interaction of K84 from RafRBD and E31 and D33 from switch I of wtRas is seen (figure 9a). Recall K31E was one of the mutations in switch I created in Raps to render the effector region 100% identical to Ras. Another association in good agreement is the interaction of guanidinium group of R59 from the RafRBD in hydrogen bonding distance with the carboxyl group of E37 from switch I of Ras (figure 9b). However for wtRas/Raf structure, Ras E37 also associates with Raf R67. This is not seen in the Raps/Raf structure. There is a shift in the main chains of residues 35 to 38 which might explain the lack of interaction in the Raps complex.
Figure 9a. Interaction of K84 from RBD and Ras E31/D33. wtRas-RafRBD complex (dark red) superimposed with Raps/RafRBD (magenta). Electrostatic interactions are seen between
the switch I region of Ras and RafRBD. To create 100% sequence identify in the effector region, E30D and K31E were mutated to make Raps. This mutation is critical to promote an
Figure 9b. Electrostatic interaction between R59 and R67 of Raf and E37 of wtRas.
Previously, with the Raps/Raf structure publication, E37 of Ras and R59 of Raf was seen as an important interaction. Interestingly, for wtRas/Raf structure, we also see an interaction between Ras E37 and Raf R67, an association not seen in the Raps/Raf structure. A shift in residues of 35 to 38 in switch I of the GTPases is seen on the bottom left of the figure, may be influencing the lack of E37-R67 interaction in the Raps-Raf structure.
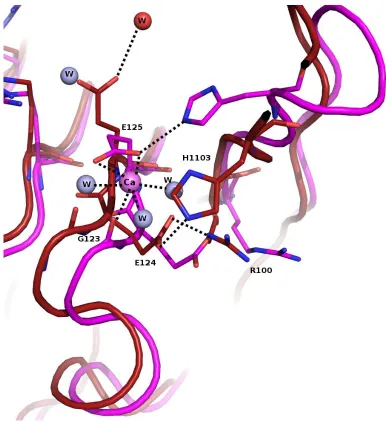
Figure 9c. Raps/Raf structure has calcium ion coordinated near E125 of RafRBD. Raps/Raf (magenta, 1GUA) and wtRas/Raf (dark red) show different conformations at E125 and H103.
In fact, a shift occurs at the backbone of residues 119-125 between the RBDs. For Raps/Raf, we see H103 hydrogen bonding with a terminal oxygen atom of E125. In wtRas/Raf, we see
H103 interacting with a terminal oxygen atom of E124. A water molecule in the Raps complex is placed where E125 is located in the wtRas complex. Possibly the bound calcium
molecules, both side chain oxygen atoms of E125 and the back bone carboxyl oxygen of G123. wtRasRaf crystals were also grown in the presence of calcium acetate, but at a much higher concentration (200 mM). This calcium ion near His103 of the RBD is missing from the wtRasRaf structure.
We also see in figure 9c, H103 interacts with a side chain oxygen atom of E125 in the Raps/Raf structure. For the wtRas complex, H103 interacts with a terminal oxygen atom of E124. A water molecule is found in the Raps complex where E125 is seen in Ras/Raf. In the Ras/Raf structure, the new conformation of E125 allows for an interaction between the terminal oxygen atom and a water molecule (red). Lastly, a shift in R100 of RBD is observed. For the Ras complex, a hydrogen bonding interaction is present between the ε-nitrogen atom and a side chain oxygen atom of E124. This interaction is not observed in the Raps complex. Figure 9d shows the electron density map for residues 100-106 of RafRBD bound to wtRas. This loop region is found between β-sheets 3 and 4 of RBD. Electron
Y71 of Ras is another residue with a different conformation that that in Raps. It appears a water molecule is hydrogen bonding with the hydroxyl group in the wtRas/Raf complex. This water molecule is missing from the Raps/Raf structure (figure 9e). The conformation of Y71 is similar to the conformation seen in our structure of wtRas-GppNHp in the allosteric on state (3K8Y).
Figure 9e. Water molecule near Y71 of Ras when bound to Raf kinase. Y71 of the switch II region in Ras (dark red) has a different conformation than Y71 in Raps (magenta). Y71 for the wtRasRaf structure agrees with the conformation seen in 3k8y. Note residues 63 and 64 are missing from switch II in wtRas/Raf complex. It appears for the space group P321, the allosteric site orders the active site, including Q61, but not all of switch II, as seen in the R32