Copyright1999 by the Genetics Society of America
Molecular Characterization of
tol
, a Mediator of Mating-Type-Associated
Vegetative Incompatibility in
Neurospora crassa
Patrick Ka Tai Shiu and N. Louise Glass
The Biotechnology Laboratory and The Botany Department, The University of British Columbia, Vancouver, British Columbia V6T 1Z3, Canada
Manuscript received July 20, 1998 Accepted for publication October 12, 1998
ABSTRACT
The mating-type locus in the haploid filamentous fungus, Neurospora crassa, controls mating and sexual development. The fusion of reproductive structures of opposite mating type, A and a, is required to initiate sexual reproduction. However, the fusion of hyphae of opposite mating type during vegetative growth results in growth inhibition and cell death, a process that is mediated by the tol locus. Mutations in tol are recessive and suppress mating-type-associated heterokaryon incompatibility. In this study, we describe the cloning and characterization of tol. The tol gene encodes a putative 1011-amino-acid polypeptide with a coiled-coil domain and a leucine-rich repeat. Both regions are required for tol activity. Repeat-induced point mutations in tol result in mutants that are wild type during vegetative growth and sexual reproduction, but that allow opposite mating-type individuals to form a vigorous heterokaryon. Transcript analyses show that tol mRNA is present during vegetative growth but absent during a cross. These data suggest that tol transcription is repressed to allow the coexistence of opposite mating-type nuclei during the sexual reproductive phase. tol is expressed in a mat A, mat a, A/a partial diploid and in a mating-type deletion strain, indicating that MAT A-1 and MAT a-1 are not absolutely required for transcription or repression of tol. These data suggest that TOL may rather interact with MAT A-1 and/or MAT a-1 (or downstream products) to form a death-triggering complex.
E
NTRY into the sexual cycle in the filamentous fun- the sexual cycle, fusion of opposite mating-type hyphae during vegetative growth results in growth inhibition gus, Neurospora crassa, requires the fusion ofrepro-ductive structures of opposite mating types (A and a). and cell death (Beadle and Coonradt 1944; Garn-jobst1953). At least 10 additional genetic determinants Cell fusion results in the presence of opposite
mating-type nuclei within a common cytoplasm. Proliferation called heterokaryon incompatibility (het) loci also re-strict formation of vegetative heterokaryons; genetic dif-of A and a nuclei occurs in the ascogenous hyphae
before formation of the crozier, where opposite mating- ferences at these loci are not required for fertilization nor do they interfere with sexual reproduction (for re-type nuclei are partitioned (Raju1980). Karyogamy
be-tween A and a nuclei occurs in the crozier and is immedi- view seeGlassandKuldau1992;Leslie 1993). The mat A and mat a sequences have been character-ately followed by meiosis and a single mitotic division
to form eight linearly arranged ascospores. Over 200 ized and are composed of dissimilar sequences (Glass et al. 1990; Staben and Yanofsky 1990). The 5301-progeny are typically formed from a single fertilization
event. Mating-type function is required for both cell bp A idiomorph contains three open reading frames (ORFs), mat A-1, mat A-2, and mat A-3. The mat A-1 gene fusion and for postfertilization events, presumably
dur-ing the proliferation and partitiondur-ing of opposite mat- determines mating identity and also confers mating-type-associated heterokaryon incompatibility (Glasset ing-type nuclei in the crozier (GriffithsandDelange
1978;Griffiths1982;GlassandLee 1992;Ferreira al. 1990). Strains that contain null mutations in mat A-1 are both sterile and heterokaryon compatible with a et al. 1998).
In addition to its role during the sexual cycle, the strains (Griffiths 1982; Saupe et al. 1996). MAT A-1 mating-type locus in N. crassa has a function during displays a region of similarity to the mating-type tran-vegetative growth. Although fusion of opposite mating- scriptional regulator, MATa1, of Saccharomyces cerevisiae type reproductive structures is required for entry into (Glass et al. 1990). The mat A-2 and mat A-3 genes are not required for mating identity or heterokaryon incompatibility, but are required for postfertilization This article is dedicated to Dr. Dorothy Newmeyer, who discovered functions (GlassandLee1992;Ferreira et al. 1996). tol.
The a idiomorph contains only one ORF, mat a-1.
Corresponding author: N. Louise Glass, The Biotechnology
Labora-A functional mat a-1 is required for mating identity, tory, Rm. 237 Wesbrook Bldg., The University of British Columbia,
Vancouver, BC, V6T 1Z3 Canada. E-mail: [email protected] heterokaryon incompatibility, and postfertilization
andBarry1977; RLM 04-08, Table 1), RLM 04-08 was crossed functions (Staben and Yanofsky 1990). As with mat
with FGSC (Fungal Genetics Stock Center, Kansas City, KS) A-1, mutations in mat a-1 result in strains that are sterile
strain 2226 (Mauriceville wild-type a). Numerous restriction and that are able to form vigorous heterokaryons with fragment length polymorphisms (RFLPs) have been observed either A or a strains (Griffiths and Delange 1978). between Mauriceville strains (FGSC 2225 and 2226; Table 1) and the Oak Ridge background of standard laboratory strains. An exceptional mat a-1 mutant, am33, is fertile but
hetero-Genomic DNA from ser-3 A progeny were screened for Mau-karyon compatible with A strains (Griffiths and
De-riceville LG IV by hybridization to cosmid G4:A9 (which
con-lange1978). The am33mutant contains a missense
muta-tains trp-4, a LG IV locus closely linked to tol ). A progeny, R5-tion in the carboxyl-terminal region of mat a-1 (Staben 28 (Table 1) had RFLP patterns identical to the FGSC 2226 andYanofsky1990). MAT a-1 has features in common parent when probed with G4:A9.
The strain NE-1 (his-5 tol trp-4 A) was constructed by crossing with transcriptional regulators, specifically an HMG
his-5 A (FGSC 456) with tol trp-4 a (FGSC 2337); a His2and box. The HMG box of MAT a-1 has been shown to bind
Trp2progeny was selected. Crossing of NE-1 to R1-09 (un-3 DNA and to be required for conferring mating identity,
a) gave R5-27 (un-3; his-5 tol trp-4 a). The presence of the tol2 but not for heterokaryon incompatibility (Philleyand allele (N83) in R5-27 was confirmed by crossing with RLM 04-Staben1994). 08 (Table 1); approximately half of the A/a partial diploid progeny displayed normal growth rates (see Figure 2). The A recessive mutation unlinked to the mating-type
lo-strain R5-27 was subsequently crossed with a pan-2-containing cus, tolerant (tol), suppresses mating-type-associated
het-strain (12-21-388) to give het-strains R4-71 and R4-72 (Table 1), erokaryon incompatibility such that tol A and tol a strains which were used for transformation assays.
form a vigorous heterokaryon (Newmeyer1970). Mat- Chromosome walk and physical mapping:A trp-4-containing ing activity of these strains is unaffected and tol strains cosmid was identified from the Orbach/Sachs pMOcosX ge-nomic library (Orbach1994; available from the FGSC) using show normal fertility in crosses with an opposite
mating-a trp-4 clone (Vollmer and Yanofsky1986) as a probe. A type strain. Attempts to identify additional suppressors
chromosome walk was performed in both directions using of mating-type-associated incompatibility resulted only end fragments as probes for each step. Selected cosmids were in the isolation of additional tol alleles (Vellani et al. physically mapped by RFLP analysis (Metzenberget al. 1985).
Transformation assay and subcloning of tol: Spheroplast 1994). The tol mutation apparently does not suppress
preparation and transformation were performed as previously incompatibility due to differences at other het loci (
New-described (Schweizer et al. 1981;Vollmer and Yanofsky meyer1970;LeslieandYamashiro1997).
1986) with the exception of selecting for resistance to hygro-This article reports the isolation and molecular char- mycin B (250 units/ml; Calbiochem). Cosmids in the walk acterization of the tol gene of N. crassa. The tol gene were assayed for Tol1activity by cotransforming the cosmids (conferring hygromycin resistance) with a mat A-1-containing encodes a putative 1011-amino-acid (aa) polypeptide
pOKE103 construct (pOKE103 has a pan-2 selectable marker; with a coiled-coil domain and a leucine-rich repeat.
J. GrotelueschenandR. L. Metzenberg,unpublished re-Mutants of tol obtained by repeat-induced point
muta-sults) into strain 71 (his-5 tol trp-4; pan-2 a) and strain R4-tion (RIP;Selker1990) lose their mating-type hetero- 72 (ade-3B; trp-4 cot-1; pan-2 A) spheroplasts. The tol gene karyon incompatibility function but are otherwise phe- was further subcloned into the hygromycin-resistant vector (pCB1004;Carrollet al. 1994). Homokaryotic transformants notypically normal during vegetative growth and sexual
were isolated according toEbboleandSachs(1990). reproduction. Expression studies indicate that tol is not
Nucleic acid isolation, DNA hybridization, and reverse
tran-transcriptionally repressed or activated by MAT A-1 scriptase PCR analysis: Standard molecular biology proce-and/or MAT a-1 during vegetative growth but is appar- dures were used throughout (Sambrooket al. 1989). Genomic ently repressed during sexual development. DNA isolation from Neurospora was adapted fromOakleyet al. (1987). Total RNA was extracted according toLogemann et al. (1987) and enriched for poly(A)1using the OligoTex mRNA kit (QIAGEN, Chatsworth, CA). Gel electrophoresis MATERIALS AND METHODS
and nucleic acids transfer to Nylon filters (Schleicher and
Strains, media, and culturing methods:The N. crassa strains Schuell, Keene, NH) were performed according to manufac-used in the study are listed in Table 1. Culturing and crossing, turer’s specifications. [a-32P]dCTP-labeled probes
(Amer-using Vogel’s (Vogel 1964) and Westergaard’s (Wester- sham, Oakville, ON) were generated from digested DNA using
gaard and Mitchell 1947) media, respectively, were per- the T7 QuickPrime Kit (Pharmacia, Baie d’Urfe, Quebec). formed as previously described, with modification (Davisand For reverse transcriptase (RT)-PCR, cDNAs were synthesized
Deserres1970;Perkins1986). For heterokaryon tests, 2-ml co- by the Not I-d(T)6primer using a procedure from the First
nidial suspensions (107/ml) of two strains containing different
Strand cDNA Synthesis Kit (Pharmacia). PCRs of DNA and auxotrophic markers were coinoculated onto vegetative cDNA were performed using primers tol 13 (59GGGCGGAG growth media (Vogel 1964). A compatible heterokaryon GATAGGAGG 39; bases 641 to 657) and tol 11 (59CCAGCAG forms a vigorously conidiating culture after 3 days of incuba- TGGCTCAGC 39; bases 1144 to 1129). The tol2mutant allele tion with a mean growth rate of 7 cm/day. Mating-type in- (N83) was amplified from strain R5-27 DNA using primers tol compatible heterokaryons are usually aconidial and have a ATG1 (59 CCTGGGCTCACCTATGC 39; base 250 to234) growth rate ofz0.7 cm/day (Vellaniet al. 1994). For perithe- and tol 39-end (59CGGCGGGATCTCTTTCTG 39; base 2894 cial RNA extraction, crosses were performed in petri dishes to 2877). The tol cDNA and tol2 (N83) PCR products were containing Westergaard medium layered with Miracloth mem- cloned into the PCRII vector using a TA cloning kit (In-brane (Calbiochem, La Jolla, CA). Perithecia were scraped vitrogen, San Diego) and subjected to DNA sequencing. The from the Miracloth and RNA was extracted as described below. entire DNA sequence of two different tol2(N83) clones was
Strain construction:To introgress part of Mauriceville link- determined and the mutation point was confirmed from three additional subclones.
TABLE 1
Neurospora crassastrains
Strain Genotype Origin
— his-5 A FGSC 456
R1-09 un-3 a R. L. Metzenberg
— tol trp-4 a FGSC 2337
— tol a FGSC 4142
NE-1 his-5 tol trp-4 A This study
MV 1c-a Mauriceville WT a FGSC 2226
MV 1 c-A Mauriceville WT A FGSC 2225
RLM 04-08 T(IL→IIR) 39311 ser-3 A R. L. Metzenberg
R5-28 T(IL→IIR) 39311 ser-3 A This study
(Mauriceville background on LG IV)
R5-27 un-3; his-5 tol trp-4 a This study
12-21-388 ad-3B al-2; cot-1; pan-2 A A. J. F. Griffiths
R4-71 his-5 tol trp-4; pan-2 a This study
R4-72 ad-3B; trp-4 cot-1; pan-2 A This study
74-OR23-1VA Oak Ridge WT A FGSC 2489
A2 ad-3A nic-2 cyh-1 a A. J. F. Griffiths
I-20-41 ad-3B arg-1; tol A A. J. F. Griffiths
I-20-26 ad-3B arg-1 A A. J. F. Griffiths
I-10-1 ad-3A nic-2; tol a A. J. F. Griffiths
I-1-51 ad-3A nic-2 a A. J. F. Griffiths
OR8-1a Oak Ridge WT a FGSC 988
RLM 44-02 thi-4; lys-1DmatA FGSC 8292
D25 his-5 tol A/a partial diploid This study
R2-11 nic-2 al-1; pan-1 am33 This study
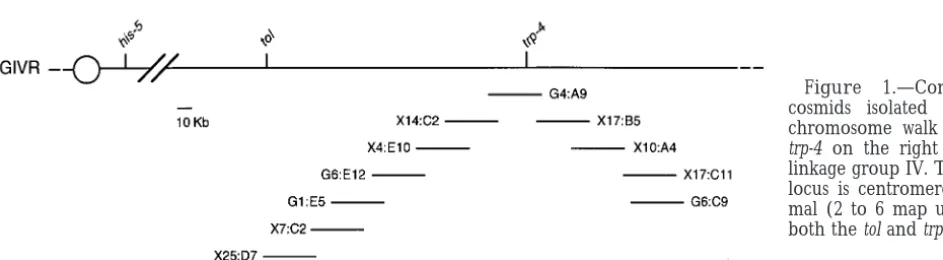
DNA sequence analyses:A 6.9-kb EcoRI tol1-containing con- (2 to 6 map units; Figure 1). A cosmid containing trp-4 struct was subcloned into overlapping fragments suitable for (G4:A9) was identified from a genomic library using the DNA sequencing. DNA sequences were determined for both
trp-4 gene (VollmerandYanofsky1986) as a probe. A strands using the ABI (Mississauga, ON) automated Taq
Dye-chromosome walk was initiated from G4:A9 in both Deoxy Terminator cycle method at the NAPS unit,
Biotechnol-ogy Laboratory, University of British Columbia. Computer directions. Twelve overlapping cosmids spanning a re-sequence analyses for protein and DNA were done using the gion of 350 kb around the trp-4 locus were isolated MacVector/AssemblyLIGN software (International
Biotech-(Figure 1). The physical location of selected cosmids nologies, New Haven, CT) and GCG package available from
was determined using RFLP mapping (Metzenberget the Wisconsin Genetics Computer Group (Devereux et al.
al. 1985) and all mapped to trp-4. 1984).
To determine the orientation of the walk from trp-4, we analyzed progeny that contained crossovers between RESULTS
his-5, tol, and trp-4 from a cross between R5-27 (un-3; his-5 tol trp-4 a; Oak Ridge background) and MV1c-A
Cloning of tol: The tol locus is on the right arm of
LG IV and is flanked by trp-4 (z1 map unit) and his-5 (FGSC 2225; Mauriceville background). Genomic DNA
Figure 2.—The tol locus was bracketed by genetic crossovers and RFLP analy-sis of progeny. (A) Cross-overs between tol and trp-4 were detected in progeny 43-9 and 43-31 (his-51 tol1 trp-4); hybrid RFLP patterns that differed from both pa-rental strains were observed when genomic DNA was probed with X14:C2. (B) Recombination events be-tween his-5 and tol were se-lected for by isolating A/a partial diploid progeny that were his-51, tol, and trp-4. RFLP analysis of genomic DNA from A/a tol trp-4 progeny identified cross-over points in the cross- overlap-ping region of cosmids G13:C8 and X25:D7. When probed with G13:C8, eight progeny (T4, T5, T10, T15, T18, T21, T35, and T37) dis-played the R5-28 (MV LG IV) pattern and one prog-eny (T11) displayed a hy-brid pattern, indicating that the tol locus resided be-tween G13:C8 and X14:C2 in the cosmid walk.
was isolated from His1Trp2progeny and probed with To bracket the tol locus within the cosmid walk, we needed to detect recombination points that were cen-cosmids shown in Figure 1. Recombination points in two
his-51tol1trp-4 progeny (43-9 and 43-31) were identified tromere proximal to tol, i.e., between tol and his-5. A cross was performed between a R5-28 (T(IL → IIR) when cosmid X14:C2 was used as a probe (Figure 2A).
These results indicated that cosmids directed toward 39311 ser-3 A; MV LG IV)3 R5-27 (un-3; his-5 tol trp-4 a) (Figure 2B). The R5-28 strain contains an insertional G13:C8 were oriented toward the tol locus, and that the
in-cludes ser-3, un-3, and mat) is inserted into the right tified at position2413, 85 bp away from the proposed transcription start site (Figure 3). Three pairs of short arm of LG II (PerkinsandBarry1977). Progeny were
selected that displayed wild-type growth in medium lack- perfect repeats (CGCCGCCCA, TTTGTTG, and GAGA AGTTCA) were found 59to the proposed CAAT box. ing serine at 308 (selection for A/a tol partial diploid
progeny; Figure 2B) and that contained tryptophan but Translation of the tol ORF identified a 1011-aa poly-peptide (Figure 3) with a calculated Mrof 113,712. TOL
lacked histidine (selection for progeny with
recombina-tion between his-5 and tol). Genomic DNA from 68 A/a has a predicted isoelectric point of 4.67 and is made up of 40% nonpolar and 26% charged residues. The tol trp-4 progeny were screened for RFLP differences
from the R5-27 parent when probed with the cosmids carboxyl-terminal portion of TOL (aa position 837– 1011) is rather hydrophilic, composed of 30% nonpolar centromere proximal to G4:A9. Fifty-nine progeny
con-tained RFLPs that were identical to the R5-27 parent, and 30% charged residues. BLAST (Altschul et al. 1990) and FASTA algorithm (Pearson and Lipman indicating that in these progeny the recombination
point between his-5 and tol lay centromere proximal to 1988) searches did not reveal significant similarity be-tween TOL and any other protein sequence present in the cosmids identified in the walk. In genomic DNA
from eight progeny, an R5-28 pattern was identified protein databases. However, a heptad repeat structure was identified from aa position 177–211 (Figure 4, A when G13:C8 was used as a probe, but an R5-27 pattern
was observed when probed with X25:D7 (Figure 2B). and B), as predicted by the COILS program (Lupaset al. 1991). A coiled-coil is an a-helical bundle that is These data indicated that the recombination point
be-tween his-5 and tol in these progeny occurred bebe-tween thought to wind into a superhelix, with the hydrophobic residues (a and d) forming the hydrophobic packing G13:C8 and X25:D7, and that tol must be centromere
distal to the crossover point. When genomic DNA from interface (for review see Lupas 1996). Coiled-coil do-mains play structural roles in numerous fibrous proteins progeny T11 was probed with G13:C8, a hybrid RFLP
pattern was identified, consistent with the interpretation (PaulingandCorey 1951; Crick1953) and are also a dimerization motif in the leucine-zipper class of tran-that the tol locus resided between G13:C8 and G4:A9.
The Tol1 activity of contiguous cosmids between scription factors (Landschulzet al. 1988). The putative coiled-coil domain in TOL had five heptad repeats and G13:C8 and G4:A9 was assayed by cotransforming each
cosmid with pOKEmat A-1 into R4-71 (his-5 tol trp-4; contained mostly hydrophobic residues in positions a and d with some charged residues in positions b, c, e, pan-2 a) and R4-72 (ad-3B; trp-4 cot-1; pan-2 A)
sphero-plasts. Transformants were selected for both hygro- f, and g. Occasional polar residues in the core (a and d, such as in the case of TOL) favor dimerization over mycin resistance (cosmid marker) and for growth in the
absence of panthothenic acid (pOKEmat A-1 marker). trimer- or tetramerization, because they can still be partly solvated (Harbury et al. 1993; Lumb and Kim Transformants fail to regenerate following the
induc-tion of mating-type-associated incompatibility (Glasset 1995).
A single putative leucine-rich repeat (LRR) was found al. 1990) and therefore a cosmid with Tol1 activity
should exhibit significantly lower transformation fre- in the carboxyl-terminal portion of TOL (position 804– 823; Figure 4A). LRRs, which have been found in quencies when introduced with mat A-1 into tol a
sphero-plasts as compared to A spherosphero-plasts. Only cosmid numerous proteins, are thought to mediate protein-protein interactions (for reviews, seeKobeand Deisen-X25:D7 (plus mat A-1) exhibited a significant reduction
in transformation frequency in a tol a background. Fur- hofer1994;BuchananandGay1996). Each LRR con-tributes an exposed b-sheet that could participate in ther subcloning identified a 6.9-kb EcoRI fragment from
X25:D7 that caused a 20- to 30-fold reduction in trans- strong interactions with other proteins. The LRR in TOL is predicted to form ab-sheet by both the Chou-formation frequencies when introduced into tol a
sphe-roplasts as compared to A sphesphe-roplasts. Fasman (Chou and Fasman 1978) and the Robson-Garnier algorithms (Garnier et al. 1978). Although
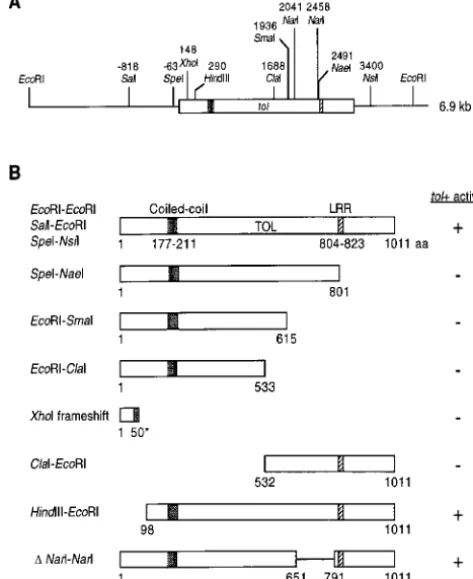
Molecular characterization oftol:DNA sequence
de-termination of a 4.2-kb Sal I-Nsi I construct within the LRRs are usually present in tandem arrays, at least three proteins with a single LRR have been reported: Rev 6.9-kb EcoRI fragment revealed an ORF of 3127 bp
inter-rupted by a putative intron of 94 bp. The 59 and 39 (HIV-1 nuclear regulatory protein;Malimet al. 1989), GPIbb (human platelet glycoprotein Ib b subunit; and internal sites of the tol intron fit intron splicing
consensus sequences (Bruchez et al. 1993; Figure 3). Lopezet al. 1988), and GPIX (human platelet glycopro-tein IX; Hickey et al. 1989). These single LRRs are The sequence of a tol cDNA spanning the intron site
was characterized and confirmed the presence of the thought to be functionally important (Malimet al. 1989; Noris et al. 1997).
intron in the genomic tol sequence. DNA sequences
surrounding the tol ATG start codon were in good agree- Functional analysis oftolconstructs:Several tol dele-tion and frameshift constructs were obtained to deter-ment with the consensus for N. crassa (Edelman and
Staben1994). A sequence matching the transcription mine what portions of TOL were important for function (Figure 5). A 11 frameshift mutation at the XhoI site initiation consensus for N. crassa was found at position
iden-Figure 3.—Nucleotide and amino acid sequence of the tol gene of N. crassa. The nucleotide sequence (4.2-kb Sal I-Nsi I fragment) and the predicted TOL protein sequence are shown. The in-tron sequence is italicized and the 59, 39, and inter-nal splicing consensus se-quences are underlined. Se-quences matching the con-sensus for a transcriptional start site and CAAT box are underlined in bold. Short repetitive sequences (three pairs) in the promoter re-gion are underlined. The putative coiled-coil domain and the leucine-rich repeat (LRR) are bracketed with {} and [], respectively. The original tol mutation, E858 (GAA) → Stop (TAA) is underlined in bold and itali-cized. The GenBank acces-sion number for this se-quence is AF085183.
portion also abolished Tol1activity. However, an inter- otic pan-21 tol a (pOKE103, SE5.1, or pOKE-EE6.9) transformants were isolated and subjected to nal deletion of 139 aa in the region before the LRR did
not affect TOL function. N-terminal deletion constructs heterokaryon tests with tol1 A (I-20-26) and tol A (I-20-41) testers. As expected, homokaryotic pan-21 tol indicated that a region between aa position 98 and 531,
which includes the coiled-coil domain, was also essen- a transformants containing only the vector (pOKE103) formed a vigorous heterokaryon with the tol A (I-20-41) tial.
Complementation of the tol mutant and DNA se- strain, but formed an incompatible heterokaryon with
the tol1 A tester (I-20-26). In contrast, the pan-21tol a quence analysis of thetol2allele:The original tol
muta-tion was identified in an A/a partial diploid strain that (pOKE-SE5.1 or pOKE-EE6.9) transformants exhibited typical mating-type-associated incompatibility with both had escaped from inhibited to wild-type growth rates
(Newmeyer 1970). The mutation was subsequently tol A and tol1A testers. These data indicated the original tol mutant phenotype was reverted to wild type (Tol1) mapped to LG IV, near trp-4. To confirm that we cloned
tol, we performed complementation experiments with by the introduction of either of the pOKE-SE5.1 or pOKE-EE6.9 clones. To confirm further that we had the tol2mutant and determined the DNA sequence of
the tol2allele (N83). cloned tol, we determined the DNA sequence of the tol2 mutant allele. The tol2 allele (N83) contained a Two pOKE103-tol1 constructs, Sal I-EcoRI (5.1 kb;
pOKE-SE5.1) and EcoRI-EcoRI (6.9 kb; pOKE-EE6.9; Fig- transversion mutation at position 2666 (G → T) that resulted in the change of a glutamine codon (GAA) ure 5), plus a vector control (pOKE103), were
Figure4.—Structural features identified in TOL. (A) The amino acid sequence of the coiled-coil domain (position 177– 211) in TOL is shown with a schematic heptad position a-g underneath. The leucine-rich repeat (LRR; aa position 804– 823) is shown and the consensus for Arabidopsis thaliana RPS2 LRR (Mindrinoset al. 1994) is depicted underneath. For the LLR, an “X” stands for an arbitrary amino acid while an “a” stands for an aliphatic amino acid. (B) A helical wheel repre-sentation of the coiled-coil domain in TOL. The hydrophobic residues of the heptad repeats are in bold while charged
resi-Figure5.—Functional analysis of tol deletion and frameshift dues are underlined.
constructs. (A) Schematic representation of the 6.9-kb tol DNA sequence including relevant restriction sites. DNA position is given according to Figure 3. (B) Schematic diagram of various
Isolation of tolmutants by RIP mutation:The muta- tol constructs used to examine functional domains of TOL.
Amino acid position is according to Figure 3. A tol construct tion in the original tol2allele (N83) occurred near the
is considered to have TOL activity if it results in a reduction carboxyl-terminus of TOL, and therefore it is possible
in transformation frequency (at least 20-fold) when cotrans-that TOL retains partial function. We therefore at- formed with mat A-1 into tol a spheroplasts, as compared to tempted to obtain additional tol mutants by RIP muta- frequencies when cotransformed into A spheroplasts. *XhoI frameshift contains 50 functional amino acids and terminates tion (Selker1990). RIP is a mechanism in Neurospora
at position 68. that causes duplicated sequences to undergo multiple
G-C to A-T transition mutations during the sexual cycle.
A 2.7-kb EcoRV fragment (cloned into pCB1004) that as RIP strains (to1-43, 83, 95, 106, and tol-contained an internal portion of tol (position 3–2735; 135).
Figure 3) was transformed into strain A2 (ad-3A nic-2 To confirm that the suppressor phenotype in the tol-cyh-1 a; Table 1). A hygromycin-resistant homokaryotic RIP mutants segregated with the tol locus, tol-43 a (as a transformant (4-9) was obtained and crossed with FGSC male) and tol-83 a (as a female) were crossed separately 456 (his-5 A). Forty-nine his-51; ade-3A nic-2 a progeny to his-5 A. Of the 30 His1progeny tested from the two
were analyzed for Tol1 activity in heterokaryon tests. crosses, all but one formed vigorous heterokaryons with Five his-51; ad-3A nic-2 a progeny formed vigorous het- both tol a (I-10-1) and tol A (I-20-41) strains (3.3% recom-erokaryons with the tol A tester (I-20-41), indicating that bination), indicating that the new mutations in tol-43 the tol allele in these strains was not functioning to and tol-83 were closely linked to the his-5 (and hence confer mating-type-associated incompatibility (Figure tol) locus. By Southern blot and RFLP analysis, it was 6A). Heterokaryons between these same five his-51; ad- determined that the tol sequence in the tol-RIP strains 3A nic-2 a progeny and the tol1 A tester (I-20-26) dis- showed restriction site changes (Figure 6B), consistent played typical heterokaryon incompatibility (Figure with the presence of transition mutations that are char-6A), indicating that the tol mutation in these strains was acteristic of sequences that have undergone RIP ( Sel-recessive. The tol-RIP strains were phenotypically normal ker1990).
yon (A1am33); the am33strain forms compatible
hetero-karyons with A strains, but sexual function is not affected (GriffithsandDelange1978).
Following fertilization, opposite mating-type nuclei coexist and divide within a common cytoplasm. There-fore, mating-type-associated incompatibility mediated by tol must be suppressed during sexual reproduction. To determine if tol is transcriptionally repressed dur-ing a cross, thus allowdur-ing the coexistence of opposite mating-type nuclei, we analyzed the presence of the tol cDNA by RT-PCR in mRNA preparations from peri-thecia at 3, 5, 6, and 9 days postfertilization. Although a cDNA for mat A-2 could easily be detected by RT-PCR from perithecial RNA preparations for all time points, a tol cDNA could not be detected at any time point postfertilization (cross; Figure 7 and data not shown).
DISCUSSION
This study describes the cloning, characterization, and expression analysis of tol, a mediator of mating-type-associated incompatibility in N. crassa. The tol locus is the only molecularly characterized mediator of allelic incompatibility from fungi. Nonallelic suppressors of incompatibility have been characterized in Podospora an-serina and encode proteins thought to be involved in developmental processes and signal transduction ( Lou-bradou et al. 1997;Barreauet al. 1998). In N. crassa, it is not clear how tol, mat a-1, and mat A-1 function to Figure6.—Phenotypic and genotypic analysis of tol-RIP
mu-trigger incompatibility during vegetative growth. One tants. (A) The top three plates are controls showing growth
characteristics of compatible (tol A1tol a) and incompatible hypothesis is that, in an A 1 a heterokaryon or A/a (tol1A1tol a and tol1A1tol1a) heterokaryons. The bottom partial diploid, MAT a-1 and MAT A-1 form a hetero-plates show the phenotype of heterokaryon with tol-43. The
dimer and this complex acts as a transcription regulator tol-43 strain was compatible with a tol A strain, but incompatible
to activate or repress the expression of tol (Glassand with a tol1 A strain. Strains I-10-41 (tol A), I-20-26 (tol1A),
Staben1990), which would then trigger incompatibil-I-10-1 (tol a), I-1-51 (tol1a), and R5-52 (tol-43 a) (Table 1)
were used in heterokaryon tests. (B) RFLP analysis of tol se- ity. Our studies show that the expression of tol occurs quences in the tol-RIP strains. Genomic DNA from parental in an A, a, and even in aDmat strain. These data indicate strains (his-5 and transformant 4-9) plus the transformation
that MAT A-1 and MAT a-1 are not required for either recipient strain (A2; Table 1) and each tol-RIP strain (tol-43,
repression or activation of tol, although we cannot rule tol-83, tol-95, tol-106, and tol-135) was digested with MboI (M)
out the possibility that the mating-type polypeptides may and Sau3AI (S) and probed with a 3.5-kb fragment containing
tol. The presence of altered restriction patterns in the RIP modulate tol expression levels. The expression of tol was progeny as compared to their parental controls is indicative also detected in an A/a tol partial diploid and an A1 of GC→AT transition mutations that occur during the RIP
a incompatible heterokaryon, indicating that if a MAT process (Selker1990).
A-1/MAT a-1 heterodimer is formed, it does not materi-ally affect the expression of tol.
On the basis of the data presented in this article, a used to amplify tol mRNA from vegetatively growing
cultures. Figure 7 shows that a tol cDNA could be de- second more plausible possibility for the mechanism of mating-type-associated incompatibility can be formu-tected from mRNA from A, a, and a mating-type deletion
strain grown in vegetative and crossing media (V and lated. In our current hypothesis, TOL physically inter-acts with the MAT A-1/MAT a-1 complex or their down-C; Figure 7). These data indicated that neither
mating-type constitution nor growth conditions materially af- stream products to trigger incompatibility. The 1011-aa TOL protein possesses a putative coiled-coil domain and fected the expression of tol. A tol cDNA could also be
detected in an A/a tol partial diploid strain, indicating a LRR, both of which are thought to mediate protein-protein interactions. Putative LRRs are found in both that the presence of both mat A and mat a in the same
nucleus did not significantly alter activation or repres- MAT A-1 and MAT a-1 (P. K. T. Shiu, unpublished results), and alteration of these motifs affects the capac-sion of tol. Exprescapac-sion of tol was also detectable in both
Figure7.—Expression of tol as analyzed by RT-PCR. Primers spanning the in-tron (tol 13 and tol 11) were used to amplify tol cDNA from different mRNA prep-arations. The size of the ge-nomic tol product is 504 bp with these primers, while the tol cDNA product is 410 bp. Strains used are as fol-low: 74-OR23-1VA (A), OR8-1a (a), RLM44-02 (DmatA), D25 (A/a tol), I-20-261 I-1-51 (A1a), and I-20-261 R2-11 (A1am33). mRNA was
iso-lated from the above cul-tures grown either in vegeta-tive medium (Vogel 1964; V) or crossing medium (Westergaardand Mitch-ell1947; C) or from perithe-cia (cross). A tol cDNA could be detected in mRNA from all strains, with the exception of peritheperithe-cial mRNA. The quality of the perithecial mRNA samples was checked by the amplification of a mat A-2 mRNA (using primers rI.1 and 2423-2406;Ferreira et al. 1996); mat A-2 is expressed at low levels throughout the sexual cycle (data not shown). DNA size markers (1-kb ladder) are given on the left.
A 4-aa deletion in the carboxyl terminus of MAT a-1 a vigorous heterokaryon. However, when mat A and mat a from N. sitophila were introgressed into N. crassa, abolished incompatibility but not DNA binding or
mat-ing activity (Philley and Staben 1994). These data mating-type-associated incompatibility was observed (Perkins 1977). In the pseudohomothallic species N. suggest that for MAT a-1, mating (which probably
in-volves transcriptional activation) and vegetative incom- tetrasperma, opposite-mating-type nuclei normally re-side in a common cytoplasm. Similar to the results ob-patibility are mediated by biochemically distinct
mecha-nisms. served with N. sitophila, mat A and mat a from N.
tetra-sperma displayed mating-type incompatibility when in-Some het genes have cellular functions other than to
limit heterokaryosis (for review, see Be´gueret et al. trogressed into N. crassa (Metzenberg andAhlgren 1973). The introgression of tol1from N. crassa into N. 1994). The original tol mutant and our tol -RIP strains
exhibit normal vegetative growth and sexual reproduc- tetrasperma induced mating-type-associated incompati-bility and disrupted the pseudohomothallic nature of tion. However, the tol mutation has been shown to
sup-press the phenotype of an fmf-1 mutant (Johnson the species (Jacobson 1992). All of these data argue that tol is the major mediator of mating-type-associated 1979), which is sterile as both a male and a female in
crosses to a wild-type partner. Fertility is restored when incompatibility. The evolutionary history and selection for or against mating-type-associated incompatibility in an fmf-1 tol strain is crossed to an fmf-11tol strain. Our
expression analysis of tol is consistent with the hypothesis the different species within the genus Neurospora re-main an enigma. The molecular characterization of tol that tol activity is suppressed during sexual development
to allow the coexistence of opposite mating-type nuclei from N. crassa and related species will provide the neces-sary tools to address these questions.
in the ascogenous hyphae. It is possible that FMF-1
regu-lates tol either directly or indirectly, and thus may play Both allelic and nonallelic incompatibility systems have been described for a number of filamentous fungi, a role in the repression of tol transcription.
Presum-ably, the misexpression of tol during the sexual cycle and it is believed that heterokaryon (or vegetative, so-matic, or heterogenic) incompatibility is a universal would result in mating-type-associated incompatibility
and block further sexual development. The presence phenomenon among filamentous ascomycetes and basi-diomycetes (Glass and Kuldau 1992; Leslie 1993; of short repetitive sequences in the promoter of tol may
provide putative binding sites for such regulators. Be´gueretet al. 1994;EsserandBlaich1994;Worrall 1997). It has been proposed that vegetative incompati-Some species within the genus Neurospora do not
exhibit mating-type-associated incompatibility. This fact bility may protect an individual from the transfer of deleterious cytoplasmic factors (Caten1972). Cytoplas-is presumably due to the presence of tol mutations within
these species that suppress mating-type-associated in- mic transfer of hypovirulence-associated dsRNA virus in Cryphonectria parasitica and detrimental KalDNA plas-compatibility. In the heterothallic species N. sitophila,
Beadle, G. W.,andV. L. Coonradt,1944 Heterokaryosis in Neuro-(Anagnostakis1982;Debetset al. 1994). In black
As-spora crassa. Genetics 29: 291–308.
pergilli, vegetative incompatibility completely blocks Be´gueret, J., B. TurcqandC. Clave´,1994 Vegetative incompatibil-ity in filamentous fungi: het genes begin to talk. Trends Genet. transfer of mycoviruses (Van Diepeningenet al. 1997).
10:441–446. Vegetative incompatibility mediated by mating type
Bruchez, J. J. P., J. EberleandV. E. A. Russo,1993 Regulatory could be selected for by two mechanisms. The first sequences in the transcription of Neurospora crassa genes: CAAT box, TATA box, introns, poly(A) tail formation sequences. Fungal mechanism is that incompatibility mediated by mating
Genet. Newsl. 40: 89–96. type could promote outbreeding by eliminating the
pos-Buchanan, S. G., andN. J. Gay,1996 Structural and functional sibility of heterokaryon formation with opposite mating- diversity in the leucine-rich repeat family of proteins. Prog.
Bio-phys. Mol. Biol. 65: 1–44. type siblings. However, in an outbreeding population,
Bucher, P.,1990 Weight matrix descriptions of four eukaryotic RNA the formation of vegetative heterokaryons is also
ex-polymerase II promoter elements derived from 502 unrelated cluded by differences at any of 10 additional het loci in promoter sequences. J. Mol. Biol. 212: 563–578.
Carroll, A. M., J. A. SweigardandB. Valent, 1994 Improved N. crassa; N. crassa populations are highly polymorphic
vectors for selecting resistance to hygromycin. Fungal Genet. for het loci (Mylyk1976). A second mechanism is that
Newsl. 41: 22.
mat A-1 and mat a-1 evolved solely for sexual reproduc- Caten, C. E.,1972 Vegetative incompatibility and cytoplasmic infec-tion in fungi. J. Gen. Microbiol. 72: 221–229.
tion, but that molecular divergence of tol resulted in
Chen, H., and J. A. Kinsey, 1995 Purification of a heteromeric mating-type-associated incompatibility during
vegeta-CCAAT binding protein from Neurospora crassa. Mol. Gen. Genet. tive growth. Because the mating-type locus is always poly- 249:301–308.
Chou, P. Y.,andG. D. Fasman,1978 Prediction of the secondary morphic in populations in a heterothallic species like
structure of proteins from their amino acids sequence. Adv. Enzy-N. crassa, mating-type-associated incompatibility would
mol. Relat. Areas Mol. Biol. 47: 45–148.
be an efficient way to restrict heterokaryon formation Crick, F. H. C.,1953 The packing ofahelixes: simple coiled-coils. Acta. Crystallogr. 6: 689–697.
with 50% of the population, even in the absence of
Davis, R. H.,andF. J. Deserres,1970 Genetic and microbiological polymorphisms at other het loci.
research techniques for Neurospora crassa. Methods Enzymol. 27A: Mating-type-associated incompatibility has been re- 79–143.
Debets, F., X. YangandA. J. F. Griffiths,1994 Vegetative incom-ported in other fungal species, such as in Ascobolus
stercor-patibility in Neurospora: its effect on horizontal transfer of mito-arius (G. N. Bistis,personal communication),
Aspergil-chondrial plasmids and senescence in natural populations. Curr. lus heterothallicus (KwonandRaper1967), and Sordaria Genet. 16: 113–119.
Devereux, J., P. HaeberliandO. Smithies,1984 A comprehensive brevicollis (J. Bond,personal communication). It is
possi-set of sequence analysis programs for the VAX. Nucleic Acids ble that the phenomenon of mating-type-associated
in-Res. 12: 387–395.
compatibility is even more widespread in filamentous Ebbole, D.,andM. Sachs, 1990 A rapid and simple method for isolation of Neurospora crassa homokaryons using microconidia. ascomycetes but cannot be assessed because of the lack
Fungal Genet. Newslett. 37: 17–18. of isogenic strains that differ only at the mating-type
Edelman, S.,andC. Staben,1994 A statistical analysis of sequence locus. The characterization of the three major compo- features within genes from Neurospora crassa. Exp. Mycol. 18:
70–81. nents of mating-type-associated incompatibility, mat
Esser, K.,andR. Blaich,1994 Heterogenic incompatibility in fungi, A–1, mat a-1, and tol, will provide tools to analyze the
pp. 211–232 in The Mycota I: Growth, Differentiation and Sexuality, molecular mechanism of mating-type-associated incom- edited byJ. D. H. WesselsandF. Meinhardt.Springer-Verlag,
Berlin. patibility in filamentous fungi and will facilitate the
Ferreira, A. D. V., S. SaupeandN. L. Glass,1996 Transcriptional challenge of delineating the molecular mechanism of
analysis of the mt A idiomorph of Neurospora crassa identifies two growth inhibition and cell death that is a characteristic genes in addition to mt A-1. Mol. Gen. Genet. 250: 767–774.
Ferreira, A. D. V., Z. An, R. L. MetzenbergandN. L. Glass,1998 feature of this phenomenon.
Characterization of mat A-2, mat A-3 andDmatA mating-type
mu-We thank Drs. X. Wu and S. J. Saupe and Ms. T. S. Vellani for tants of Neurospora crassa. Genetics 148: 1069–1079.
isolating some of the cosmids in the walk and performing some hybrid- Garnier, J., D. J. OsguthorpeandB. Robson,1978 Analysis of ization experiments. We are indebted to V. Kyritsis for the tol allele the accuracy and implications of simple methods for predicting the secondary structure of globular proteins. J. Mol. Biol. 120: isolation. We thank Drs. A. J. F. Griffiths, D. J. Jacobson, and M. L.
97–120. Smith for critical reading of the manuscript. P. K. T. Shiu is a recipient
Garnjobst, L.,1953 Genetic control of heterokaryosis in Neurospora of the Natural Sciences and Engineering Research Council of Canada
crassa. Am. J. Bot. 40: 607–614.
(NSERC) postgraduate fellowship. This work was supported by an
Glass, N. L.,andG. A. Kuldau,1992 Mating type and vegetative NSERC grant to N.L.G.
incompatibility in filamentous ascomycetes. Annu. Rev. Phytopa-thol. 30: 201–224.
Glass, N. L.,andL. Lee,1992 Isolation of Neurospora crassa A mating-type mutants by repeat-induced point (RIP) mutation. Genetics LITERATURE CITED
132:125–133.
Glass, N. L.,andC. Staben,1990 Genetic control of mating in
Altschul, S. F., W. Gish, W. Miller, E. W. MyersandD. J. Lipman,
1990 Basic logical alignment search tool. J. Mol. Biol. 215: 403– Neurospora crassa. Semin. Dev. Biol. 1: 177–184.
Glass, N. L., J. GrotelueschenandR. L. Metzenberg,1990
Neu-410.
Anagnostakis, S. L., 1982 Biological control of chestnut blight. rospora crassa A mating-type region. Proc. Natl. Acad. Sci. USA
87:4912–4916. Science 215: 466–471.
Barreau, C., M. Iskander, G. Loubradou, V. Levallois and J. Griffiths, A. J. F.,1982 Null mutants of the A and a mating-type alleles of Neurospora crassa. Can. J. Genet. Cytol. 24: 167–176.
Be´gueret,1998 The mod-A suppressor of nonallelic
heterokar-yon incompatibility in Podospora anserina encodes a proline-rich Griffiths, A. J. F.,andA. M. Delange,1978 Mutations of the a mating-type gene in Neurospora crassa. Genetics 88: 239–254. polypeptide involved in female organ formation. Genetics 149:
between two-, three-, and four-stranded coiled coils in GCN4 Newmeyer, D.,1970 A suppressor of the heterokaryon-incompatibil-ity associated with mating type in Neurospora crassa. Can. J. Genet. leucine zipper mutants. Science 262: 1401–1407.
Hickey, M. J., S. A. WilliamsandG. J. Roth,1989 Human platelet Cytol. 12: 914–926.
Noris, P., S. Simsek, J. StibbleandA. E. Von Dem Borne,1997 A glycoprotein IX: an adhesive prototype of leucine-rich
glycopro-teins with flank-center-flank structures. Proc. Natl. Acad. Sci. USA phenylalanine-55 to serine amino-acid substitution in the human glycoprotein IX leucine-rich repeat is associated with Bernard-86:6773–6777.
Jacobson, D. J.,1992 Control of mating type heterokaryon incom- Soulier syndrome. Br. J. Haematol. 97: 312–320.
Oakley, C. E., C. F. Weil, P. L. KretzandB. R. Oakley,1987 Clon-patibility by the tol gene in Neurospora crassa and N. tetrasperma.
Genome 35: 347–353. ing of the riboB locus of Aspergillus nidulans. Gene 53: 293–298.
Orbach, M. J.,1994 A cosmid with a HyR marker for fungal library
Johnson, T. E.,1979 A Neurospora mutation that arrests perithecial
development as either male or female parent. Genetics 92: 1107– construction and screening. Gene 150: 159–162.
Pauling, L.,andR. B. Corey,1951 The structure of synthetic poly-1120.
Kobe, B.,andJ. Deisenhofer,1994 The leucine-rich repeat: a versa- peptides. Proc. Natl. Acad. Sci. USA 37: 241–250.
Pearson, W. R.,andD. J. Lipman,1988 Improved tools for biologi-tile binding motif. Trends Biochem. Sci. 19: 415–421.
Kwon, K.,andK. B. Raper,1967 Heterokaryon formation and ge- cal sequence comparison. Proc. Natl. Acad. Sci. USA 85: 2444– 2448.
netic analyses of color mutants in Aspergillus heterothallicus. Am.
J. Bot. 54: 49–60. Perkins, D. D.,1977 Behavior of Neurospora sitophila mating-type alleles in heterozygous duplications after introgression into
Neu-Landschulz, W. H., P. F. JohnsonandS. L. McKnight,1988 The
leucine zipper: a hypothetical structure common to a new class rospora crassa. Exp. Mycol. 1: 166–172.
Perkins, D. D.,1986 Hints and precautions for the care, feeding of DNA binding proteins. Science 240: 1759–1764.
Leslie, J. F., 1993 Fungal vegetative incompatibility. Annu. Rev. and breedings of Neurospora. Fungal Genet. Newsl. 33: 35–41.
Perkins, D. D.,andE. G. Barry,1977 The cytogenetics of Neuro-Phytopathol. 31: 127–150.
Leslie, J. F.,andC. T. Yamashiro,1997 Effects of the tol mutation spora. Adv. Genet. 19: 133–285.
Philley, M. L.,andC. Staben,1994 Functional analyses of Neuro-on allelic interactiNeuro-ons at het loci in Neurospora crassa. Genome 40:
834–840. spora crassa MT a-1 mating type polypeptide. Genetics 137: 715–
722.
Logemann, J., J. Schell and L. Willmitzer, 1987 Improved
Raju, N. B.,1980 Meiosis and ascospore genesis in Neurospora. Eur. method for the isolation of RNA from plant tissues. Anal.
Bio-J. Cell. Biol. 23: 208–223. chem. 163: 16–20.
Sambrook, J., E. F. FritschandT. Maniatis,1989 Molecular
Clon-Lopez, J. A., D. W. Chung, K. Fujikawa, F. S. Hagen, E. W. Davie
ing: A Laboratory Manual, Ed. 2. Cold Spring Harbor Laboratory et al., 1988 Theaandbchains of human platelet glycoprotein
Press, Cold Spring Harbor, NY. Ib are both transmembrane proteins containing a leucine-rich
Saupe, S., L. Stenberg, K. T. Shiu, A. J. G. Griffiths andN. L.
amino acid sequence. Proc. Natl. Acad. Sci. USA 85: 2135–2139.
Glass,1996 The molecular nature of mutations in the mt A-1
Loubradou, G., J. Be´gueretandB. Turcq,1997 A mutation in
gene of Neurospora crassa A idiomorph and their relation to mat-an HSP90 gene affects the sexual cycle mat-and suppresses vegetative
ing-type function. Mol. Gen. Genet. 250: 115–122. incompatibility in the fungus Podospora anserina. Genetics 147:
Schweizer, M., M. E. Case, C. C. Dykstra, N. H. GilesandS. R.
581–588.
Kushner,1981 Identification and characterization of
recombi-Lumb, K. J.,andP. S. Kim,1995 A buried polar interaction imparts
nant plasmids carrying the complete qa gene cluster from Neuro-structural uniqueness in a designed heterodimeric coiled coil.
spora crassa including the qa-11regulatory gene. Proc. Natl. Acad. Biochemistry 34: 8642–8648.
Sci. USA 78: 5086–5090.
Lupas, A.,1996 Coiled coils: new structures and new functions.
Selker, E. U.,1990 Premeiotic instability of repeated sequences in Trends Biochem. Sci. 21: 375–382.
Neurospora crassa. Annu. Rev. Genet. 24: 579–613.
Lupas, A., M. Van DykeandJ. Stock,1991 Predicting coiled coils
Staben, C.,andC. Yanofsky,1990 Neurospora crassa a mating-type
from protein sequences. Science 252: 1162–1164.
region. Proc. Natl. Acad. Sci. USA 87: 4917–4921.
Malim, M., S. Bo¨ hnlein, J. HauberandB. Cullen,1989 Functional
Van Diepeningen, A. D., A. J. M. DebetsandR. F. Hoekstra,1997 dissection of the HIV-1 Rev activator—derivation of a
trans-Heterokaryon incompatibility blocks virus transfer among natural dominant repressor of Rev function. Cell 58: 205–214.
isolates of black Aspergilli. Curr. Genet. 32: 209–217.
Metzenberg, R. L.,andS. K. Ahlgren,1973 Behaviour of
Neuro-Vellani, T. S., A. J. F. GriffithsandN. L. Glass,1994 New
muta-spora tetrasperma mating-type genes introgressed into N. crassa. tions that suppress mating-type associated incompatibility in
Neu-Can. J. Genet. Cytol. 15: 571–576. rospora crassa. Genome 37: 249–255.
Metzenberg, R. L., J. N. Stevens, E. U. SelkerandE. Morzycka- Vogel, H. J., 1964 Distribution of lysine pathways among fungi: Wroblewska,1985 Identification and chromosomal distribu- evolutionary implications. Am. Nat. 98: 435–446.
tion of 5S RNA genes in Neurospora crassa. Proc. Natl. Acad. Sci. Vollmer, S. J.,andC. Yanofsky,1986 Efficient cloning of genes USA 82: 2067–2071. from Neurospora crassa. Proc. Natl. Acad. Sci. USA 83: 4869–4873.
Mindrinos, M., F. Katagiri, G. L. YuandF. M. Ausubel,1994 The Westergaard, M.,andH. K. Mitchell,1947 Neurospora V. A
syn-A. thaliana disease resistance gene RPS2 encodes a protein con- thetic medium favoring sexual reproduction. Am. J. Bot. 344:
taining a nucleotide-binding site and leucine-rich repeats. Cell 573–577.
78:1089–1099. Worrall, J. J.,1997 Somatic incompatibility in Basidiomycetes.
My-Mylyk, O. M.,1976 Heteromorphism for heterokaryon incompati- cologia 89: 24–36. bility genes in natural populations of Neurospora crassa. Genetics