DOI: 10.1534/genetics.108.100024
The Functional Transfer of Genes From the Mitochondria
to the Nucleus: The Effects of Selection, Mutation,
Population Size and Rate of Self-Fertilization
Yaniv Brandvain
1and Michael J. Wade
Department of Biology, Indiana University, Bloomington, Indiana 47405 Manuscript received December 22, 2008
Accepted for publication May 6, 2009
ABSTRACT
The transfer of mitochondrial genes to the nucleus is a recurrent and consistent feature of eukaryotic genome evolution. Although many theories have been proposed to explain such transfers, little relevant data exist. The observation that clonal and self-fertilizing plants transfer more mitochondrial genes to their nuclei than do outcrossing plants contradicts predictions of major theories based on nuclear re-combination and leaves a gap in our conceptual understanding how the observed pattern of gene transfer could arise. Here, with a series of deterministic and stochastic simulations, we show how epistatic selection and relative mutation rates of mitochondrial and nuclear genes influence mitochondrial-to-nuclear gene transfer. Specifically, we show that when there is a benefit to having a mitochondrial gene present in the nucleus, but absent in the mitochondria, self-fertilization dramatically increases both the rate and the probability of gene transfer. However, absent such a benefit, when mitochondrial mutation rates exceed those of the nucleus, self-fertilization decreases the rate and probability of transfer. This latter effect, however, is much weaker than the former. Our results are relevant to understanding the probabilities of fixation when loci in different genomes interact.
G
ENOMIC investigations of obligate intracellular endosymbionts (Moranand Wernegreen2000; Akmanet al.2002; Tamaset al.2002; Wernegreenet al. 2002; Klasson and Andersson 2004; Foster et al. 2005) reveal a reduction in genome size and number of protein-coding genes compared to their free-living rela-tives (Charleset al.1999; Gilet al.2002; Wernegreen et al.2002; Moran2003; VanHamet al.2003; Klasson and Andersson2004; Khachaneet al.2007). Similarly, mitochondria—ancient obligate intracellular symbionts of eukaryotes—have retained very few protein-coding genes (Boore 1999; Adams et al.2002) [Reclinomonas americanasis at the extreme of retention of mitochon-drial genes (Lang et al. 1997)]. Understanding the process of gene loss in mitochondria and other endo-symbionts is a major research focus of mitochondrial and endosymbiont genomics (Moran 2003; Timmis et al. 2004; Khachane et al. 2007; Bock and Timmis 2008).The loss of endosymbiont genes can be complete, in which lost genes are absent from the host–endosymbi-ont complex, a substitution, in which a nuclear allele functions in place of the lost symbiont gene, or a func-tional transfer of an endosymbiont gene to the nucleus,
followed by its loss (Adams and Palmer 2003). Such ‘‘functional transfer’’ involves the relocation of a mito-chondrial gene to the nucleus, its acquisition of a promoter, successful targeting to the mitochondria for proper function, and the eventual loss of the gene from the mitochondrial genome altogether. Although this process is probably quite complex and requires numer-ous evolutionary modifications (Murcha et al.2005), there is evidence that some mitochondrial genes are preadapted to functional transfer as they contain signals that target them to the mitochondria before functional transfer to the nucleus (Uedaet al.2008a). The complex evolution of rps16 is an illuminating case of both functional gene transfer and substitution. In some line-ages, the mitochondrial rps16 is functionally expressed in the nucleus but absent from the mitochondria (functional transfer) while in a subset of taxa, the chloroplast copy is also absent and the nuclear gene is also targeted to the chloroplast [substitution (Ueda et al.2008b)].
A number of evolutionary scenarios have been pro-posed to account for the massive loss of genes from endosymbionts. A subset of models argues that endo-symbiont gene loss is a neutral or nearly neutral process. Since endosymbiosis reduces the strength of selection on genes that are unnecessary or redundant in an obligate intracellular environment, these genes may be quickly lost by the neutral fixation of a deletion or other loss-of-function mutations. Moreover, even when
Supporting information is available online athttp://www.genetics.org/ cgi/content/full/genetics.108.100024/DC1.
1Corresponding author:Indiana University, Department of Biology, 1001
E. Third St., Bloomington, IN 47405. E-mail: [email protected]
selection favors the retention of genes in endosym-bionts, such selection may be ineffective because of reduction in effective population size due to recurrent bottlenecking (Rispeand Moran2000). Additionally, frequent input of functional endosymbiont genes into the nucleus makes symbiont genes redundant, exacer-bating gene loss via functional transfer (Berg and Kurland2000).
An alternative class of explanations views the loss of mitochondrial genes (be it complete loss, substitution, or functional transfer) as an adaptive process. The ‘‘mitochondrial competition theory’’ argues that mito-chondrial genomes that either do not contain or do not express a given allele have a replicative advantage over other mitochondria, providing a within-host selective advantage to mitochondrial gene loss (Albert et al. 1996; Selosseet al.2001; Yamauchi2005). The ‘‘bene-fits of sex’’ model posits that the genomic diploid nuclear environment (diploid, sexual) is in some way preferable (e.g., as an escape from Muller’s ratchet or Hill–Robertson interference) to a haploid asexual mitochondrial environment (Blanchard and Lynch 2000). The epistatic model (Wade and Goodnight 2006) does not advance a specific or consistent benefit to transfer, but posits that transfer is explicitly a process of coevolution between mitochondrial and nuclear genomes, where fitness is a function of the gene com-bination rather than of either gene separately.
Because few species are currently undergoing mito-chondrial to nuclear gene transfer, these alternative hypotheses are difficult to distinguish with direct ex-perimentation. However, the distribution of transferred genes across lineages allows for evaluation of the alternative hypotheses. For example, self-pollination reduces the rate of heteroplasmy and consequently the opportunity for competition among genetically distinct mitochondria. Thus, the mitochondrial com-petition theory predicts an excess of transfer events in sexual, outcrossing lineages, with high degrees of ‘‘paternal leakage.’’ Similarly, frequent self-fertilization diminishes the benefits of sex, and thus the benefits of sex hypothesis predicts fewer transfers in selfing and clonally reproducing plants than in outcrossing taxa. The epistatic model makes the opposite prediction. Selfing and clonal reproduction maintain cyto-nuclear gene combinations and increase the response to selec-tion on epistatic combinaselec-tions, potentially encouraging transfer. On the other hand, outcrossing tends to break apart adaptive cyto-nuclear gene combinations, poten-tially decreasing the amount of adaptive transfer in outcrossing lineages.
Plant lineages with high levels of self-fertilization or asexual reproduction transfer more mitochondrial genes to their nuclei than predominantly sexual and outcrossing lineages (Brandvain et al. 2007). This result is consistent with predictions of the epistatic model and is contrary to predictions of the
mitochon-drial competition or benefits of sex models. More specific predictions allowing further empirical tests re-quire more detailed theoretical investigations of the gene transfer process. Here, we investigate the roles of mutation, selection, and random drift in gene transfer using both deterministic models and stochastic simu-lations to refine and extend predictions of patterns of functional mitochondrial to nuclear gene transfer.
METHODS
To address the effect of mating system on the prob-ability of functional gene transfer, we simulated a mon-oecious Fisher–Wright population with two loci—one nuclear and one mitochondrial. At the mitochondrial locus, which is haploid and strictly matrilineally in-herited,Mrepresents a functional copy of an essential mitochondrial gene, while m represents a nonfunc-tional allele. Initially, the population is fixed for M; however, eachMallele has a probability,mm, of mutating
to the nonfunctional allele,m, every generation whilem
alleles cannot revert to their functional progenitor,M. At the nuclear locus, 1 represents a functioning, transferred copy of the essential mitochondrial gene, while allele 0 represents a nonfunctional (or absent) allele. The nuclear locus is diploid, and biparentally inherited, with Mendelian meiosis. Like the mitochon-drial allele, each 1allele has a probability,mn, of
mu-tating to the nonfunctional allele, 0, and there is no back mutation.
Allele 1 begins at a low frequency (5%); i.e., rare physical transfer has occurred, but the evolutionary resolution of this transfer event is uncertain and may be affected by the nature of selection, the mating system, the effective population size, and the relative mutation rates of the two genomes. Genotypic frequencies in the population in the first generation equal the values expected given the selfing rate (Table 1). We then make new generations by the processes of mutation, repro-duction, and selection.
TABLE 1
Initial frequencies and fitnesses of all two-locus genotypes
Cyto-nuclear
genotype Initial frequency Fitness
M11 f2
1 1Sf1f0 wM11¼1.00
M10 2f1f0ð1SÞ wM10¼1.00
M00 f2
0 1Sf1f0 wM00¼1.00
m11 0 1.00#wm11#wm10
wm10#wm11
m10 0 0.90#wm10
m00 0 wm00¼0.00
Although interesting evolutionary dynamics may occur in between the introduction of a new mutation (at frequency 1/2N) and the initiation of our simulation (5%), much of this initial process will be characterized by neutral evolutionary processes because the nuclear gene is unlikely to have accumulated a substantial amount of loss-of-function mutations and therefore should be unaffected by the parameters investigated below.
Deterministic simulations: We begin at generation zero with a vector of genotypic frequencies (Ft,t ¼0,
Table 1). Genotypic frequencies after selection, Ft9, equal the product of the relative fitness matrix,W˜, in which all values on the main diagonal are the genotypic relative fitnesses, while all values off the main diagonal equal 0 (appendix) and Ft, the vector of initial allele frequencies:
Ft9¼W˜ Ft: ð1Þ
Multiplying the reproduction matrix,R, byF9tyields
F$t—the vector of expected genotypic frequencies after
selection. We complete the transition from genotypes (Ft) at one generation to the next (Ft11; note thatF$t¼
Ft11) by multiplication of the mutation matrix (m) and F$t.
Because loss of the functional mitochondrial allele is approached asymptotically, and because the functional nuclear allele attains different frequencies at mutation– selection balance across fitness functions and selfing rates, we iterate these matrices until the frequency of the functional nuclear allele is common (i.e.,.95%) and report the number of generations needed for the population to reach this frequency.
Parameters: In our deterministic models of infinitely large populations with discrete, nonoverlapping gener-ations, we investigate the effects of mutation by facto-rially combining mitochondrial and nuclear mutation rates (mm and mn, respectively) in increments of an
order of magnitude from 103to 106. We examine the influence of selfing by varying selfing rate from zero to one in 0.001 increments.
We examine a wide array of selection regimes (Table 1). For all selection regimes, all genotypes containing a functional mitochondrial gene have a fitness of one— that is,wM11¼wM10¼wM00¼1.00. Because we assume
that theMallele is essential for survival, we set the fitness of individuals entirely lacking a functional copy of this allele to zero (i.e., wm00 ¼ 0). We vary the fitness of
heterozygotes lacking a functional mitochondrial allele from 0.90 to 1.10 in 0.05 increments (i.e., 0.90#wm10#
1.10) to explore transfer with (wm10,1.00) and without
(wm10$1.00) heterozygote fitness valleys. We vary the
fitness of the genotype homozygous for the functional nuclear allele but with a nonfunctional mitochondrial allele from 1.00 to 1.10 in 0.05 increments (i.e., 1.00#
wm11 # 1.10). Because nuclear overdominance will
maintain the system at an intermediate equilibrium, we excluded cases in whichwm10.wm11.
Stochastic simulations: From initial genotypic fre-quencies (Table 1), we create the next generation one individual at a time until the population consists of N
individuals. Each genotype in the parental generation (t) has anf iwi=wprobability of being the mother of an
individual in the offspring of generation (t11). This mother is also the father with probabilityS, whereSis the selfing rate. With probability 1S, that mother is not the father and a new parent is chosen (with replacement) where each genotype has a probability,
fiwi=w, of being the father. One nuclear allele is chosen
at random from each parent, while the mitochondrial allele is maternally inherited. Each allele is then subject to a risk of mutation according to the rate appropriate for the genome of origin. We iterate this sampling scheme until we createNoffspring. Then, we repeat this process until either the mitochondrial gene is lost (trans-fer) or the nuclear gene is lost (retention) and repeat this algorithm 1000 times for each set of parameters.
Parameters:In our stochastic simulations, generations are discrete and nonoverlapping, and population sizes are stationary. We investigate factorial combinations of three population sizes,N(100, 500, and 1000), and 11 selfing rates, S (0–1 in 0.10 increments). All other parameters (i.e., selective regimes and mutation rates) match those of the deterministic model.
Since nuclear overdominance can maintain the sys-tem at an intermediate equilibrium, we exclude fitness combinations in which wm10 . wm11 and lacks a fully
balanced design. To create a balanced analysis we com-pare the influence of homozygote fitness, keeping het-erozygote fitness constant. Turning our attention to the effect of heterozygote fitness, we compare across hetero-zygote fitnesses, holding homohetero-zygote fitness constant.
RESULTS
Deterministic simulations: The number of genera-tions necessary for a functional nuclear allele to increase from rarity (f0 ¼0.05) to commonness (ft ¼ 0.95) is
determined by an interaction of the selfing rate, relative mutation rates (mmvs.mn), and the selection scenario.
Here we discuss each of the three different classes of mutational parameters: (a)mm>mn, (b)mm¼mn, and
(c)mm?mn(see Figure 1, A, B, and C, respectively).
mn ?mm:When the nuclear mutation rate is greater
than that of the mitochondria, a fitness benefit of nuclear relocation is a necessary but not sufficient criterion for the spread of the nuclear allele. It is not sufficient, since a benefit to transfer (wM11.1.00) does
commonness decreases very quickly with increased selfing rates (Figure 1A). The selfing rate plays a role because the benefit to nuclear transfer is dependent on the mitochondrial genotype, and selfing increases the heritability of mito-nuclear gene combinations (Wade and Goodnight2006). These results are qualitatively consistent across all combinations of mutation rates examined so long asmn?mm(not shown).
mn¼mm:When nuclear and mitochondrial mutation
rates are equal, a fitness benefit is still necessary for deterministic increase in the frequency of a mitochon-drial gene transposed to the nucleus; however, the nuclear allele can increase in frequency over a wider region of parameter space than that described above (wheremn?mm, compare Figure 1A and 1B). Here the
time until commonness decreases as selfing rate in-creases;i.e., the first derivative of the function in Figure 1B is always negative.
Although dy/dx is always negative, the second de-rivative is always positive and generally decreasing, ultimately approaching zero at extreme rates of self-fertilization. This approach is not smooth, and we use critical points representing local minima and maxima ofd2y/dx2(d
minanddmaxcorresponding tod3y/dx3¼0, d4y/dx4.0 andd3y/dx3¼0,d4y/dx4,0, respectively) to describe and compare the shapes of these functions. Hereafter, we designate these pointsd1.05minandd1.10min
andd1.05maxandd1.10max, in which heterozygote fitness, wm10, equals one and the numerical subscript following
drepresents the homozygote fitness,wm11.
Regardless of the mutation rate, the shape of this curve is much the same, withd1.05minandd1.10minand d1.05max and d1.10max occurring at nearly equivalent
selfing rates (d1.05min0.78,d1.10min0.69,d1.05max
0.89,d1.10max0.81). Although the selfing rates at these
inflection points are invariant across mutation rates, the corresponding time until commonness at these points differs. For all four cases ofmm¼mnexamined,dminis
proportional to (1/m), the inverse of the mutation rate. While dmaxis also inversely related with the mutation
rate, the form of this relationship is more complicated (see Table 2).
mn > mm: When the mitochondrial mutation rate
exceeds the nuclear mutation rate, a functional nuclear allele can become common without a direct fitness advantage. However, this mechanism of gene transfer is very slow and varies minimally across selfing rates, until selfing is nearly obligate. By this point (S¼0) the time to commonness is 1.5–2 times greater than it is with no selfing at all (S¼0; Figure 1C9).
When transfer is adaptive (wm11.1.05), the time to
commonness decreases with increased selfing (dy/dx,
0). Again, the second derivative is always positive, but ultimately approaches zero, with a shape similar to that described in the previous section. Table 2 shows that whenevermn>mm, the time until commonness at the
critical points is very sensitive to changes in nuclear mutation rates but not to changes in the mitochondrial mutation rate. As was the case when mutation rates were equal, time to commonness at d1.05min and d1.10min
increases by an order of magnitude as mitochondrial mutation rates decrease by an order of magnitude.
Stochastic simulations:Overall, we investigated 6336 parameter combinations (12 fitness combinations34 nuclear mutation rates 3 4 mitochondrial mutation Figure 1.—Results of deterministic simulation model
across mutation rates and fitnesses. (A)mn¼103 ?m
m¼
104. (B) m
n¼ 104 ¼ mm¼ 104. (C)mn ¼ 105 > mm¼
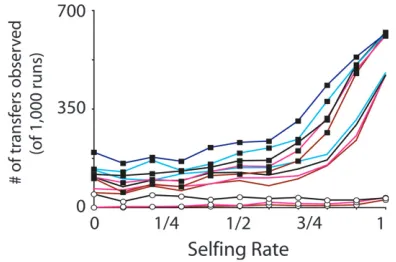
rates33 population sizes311 selfing rates¼6336) and each was replicated 1000 times, totaling .6 million independent binomial results. Below, we describe these results with the aid of the applied logistic regression (Hosmerand Lemeshow1989). The number of trans-fers out of 1000 runs, whenmn¼mm¼13103, andN¼
1000, is presented in Figure 2. Results across all population sizes and mutation rates are presented in
supporting information, File S1. Although we do not correct for multiple comparisons,P-values of significant results are generally very small and supported by greater trends, suggesting that these results are robust to multiple comparisons.
Main effects:For all main effects we report results as odds ratios so that factors that enhance the likelihood of gene transfer give a ratio greater than one, while those that reduce the likelihood have a ratio less than one.
The denominator of our odds ratios is the case where transfer to the nucleus goes through an unfavorable heterozygous stage (wm10¼0.90) with neutral
homozy-gotes (wm11¼1.00), very low mutation rates (mn¼mm¼
106), and small population sizes (N¼100). The focal value of the parameter of interest is the numerator.
Selection: Increasing the fitness of m11 individuals increases the probability of transfer. The odds of achieving functional transfer were twice as high [odds ratio (OR)¼1.97, SE¼0.010,P,0.001] whenwm11¼
1.05 relative to the neutral case (wm11¼1.00) and 2.5
times as high whenwm11¼1.10 (OR¼2.52, SE¼0.012,
P,0.001).
Increasing the fitness of m10 genotypes has similar but much weaker effects (Table 3). Relative to the ‘‘underdominant’’ heterozygote fitness wm10 ¼ 0.90,
wm10¼0.95 has an OR of 1.03 (SE¼0.011,P¼0.003)
and wm10¼1.00 has an OR of 1.09 (SE ¼0.011, P ,
0.001).
Mutation:Increasing the mitochondrial mutation rate increases the probability of transfer, suggesting that
Figure2.—Number of functional transfers of mitochon-drial genes to the nucleus per 1000 stochastic simulations. Here, we report results from large populations (N ¼ 1000) with high mutation rates (mn¼103¼mm¼103). Complete
results, spanning all mutation rates and population sizes ex-amined, are presented inFile S1. Homozygote fitness,wm11, is given by symbols (open circles,wm11¼ 1.00; no symbol, wm11¼1.05; solid boxes,wm11¼1.10). Heterozygote fitness, wm10is given by line color (brown,wm10¼0.90; red,wm10¼ 0.95; black,wm10¼1.00; light blue,wm10¼ 1.05; dark blue, wm10¼1.10).
TABLE 3
Effect of increasingm10heterozygote fitness on the likelihood of functional transfer from the mitochondria
to the nucleus
Wm11 Wm10 OR SE P
1.00 0.95 1.03 1.053102 0.003
1.00 1.00 1.10 1.113102 ,0.001 1.05 0.95 1.04 7.893103 ,0.001 1.05 1.00 1.09 8.193103 ,0.001 1.05 1.05 1.16 8.573103 ,0.001 1.10 0.95 1.05 7.123103 ,0.001 1.10 1.00 1.09 7.343103 ,0.001 1.10 1.05 1.15 7.673103 ,0.001 1.10 1.10 1.23 1.233103 ,0.001
The odds ratio, OR, measures the likelihood of transfer rel-ative to the case wherewm10¼0.90.
TABLE 2
Inflection points describing results from deterministic models across mutation rates
mm mn d1.05min d1.05max d1.10min d1.10max
mm¼mn 103 103 3.43103 7.03102 2.3 3103 4.23102 104 104 3.13104 3.13103 2.0 3104 1.63103 105 105 3.13105 2.33104 2.0 3105 1.03104 106 106 3.03106 2.13105 2.0 3106 9.13105
mm?mn 103 104 1.13103 5.93102 1.1 3103 3.83102
103 105 1.13103 5.83102 1.1 3103 3.73102 103 106 1.03103 5.83102 1.1 3103 3.73102 104 105 8.73103 2.73103 9.5 3103 1.53103 104 106 8.43103 2.73103 9.1 3103 1.53103 105 106 8.23104 2.03104 9.2 3104 9.43103
Homozygote fitness values are indicated by the subscript immediately followingd. dminanddmaxcorrespond
mutation pressure promotes mitochondria-to-nuclear gene transfer. This effect increases nonlinearly with the mitochondrial mutation rate (mm¼105, OR ¼1.04,
SE¼5.903103,P,0.001;m
m¼104, OR¼1.61, SE¼
8.383 103,P , 0.001;m
m ¼103, OR¼4.32, SE ¼
1.993102,P,0.001).
Nuclear mutational pressure has an effect similar in magnitude but opposite in direction. Increasing the nuclear mutation rate from 106to 105has a negligible effect (OR¼1.00, SE¼3.90 3103
, P¼0.666), but increasing it to 104significantly decreases the proba-bility of functional transfer (OR¼0.84, SE ¼3.43 3
103,
P, 0.001). At the highest nuclear mutation rate examined, mn ¼ 103, the probability of transfer is
reduced 60% (OR¼0.41, 2.023103,P,0.001).
Population size: Large populations are more likely to successfully transfer mitochondrial genes to the nucleus than small populations (relative toN ¼100; N¼500, OR¼1.98, SE¼8.653103,P,0.001;N¼1000, OR¼ 2.87, SE ¼ 1.19 3 103, P , 0.001). Since much of parameter space examined in this simulation involves a selective benefit to transfer, this is expected as the efficacy of selection is mitigated by random drift, which weakens with increases inN.
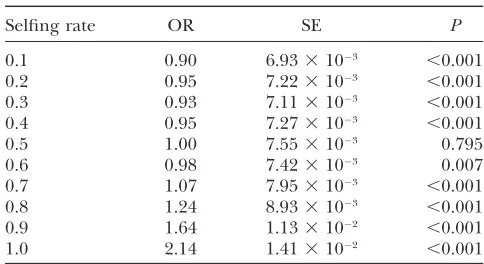
Selfing rate:For most but not all cases, increasing the selfing rate increases the probability of transfer. Com-pared to strict outcrossing, low levels of selfing slightly
decrease the probability of transfer. As selfing rate creases further, the probability of transfer slowly in-creases, until the odds ratio is approximately equal to one at a selfing rate of 0.7. At this point, slight increases in selfing rate dramatically increase the probability of transfer (Table 4). This result reflects the findings of Brandvain et al. (2007) across plant taxa, where strongly selfing groups were much more likely to trans-fer mitochondrial genes to their nuclei than outcrossing species or plants with mixed mating systems.
Interactions among parameters:From the total of 56 interaction terms (15 two-way120 three-way115
four-way1 6 five-way ¼56 total interactions), we discuss a subsample illustrative of the overall trends. InFile S2, we present all pairwise interactions between population size and other parameters. In investigating higher-order interactions, we restrict our analysis to large populations (N¼1000).
Interactions with population size: In large populations, the effect of selection favoring transfer is more pro-nounced than it is in small populations (File S2; Table 1, interaction between N and wm11; Table 2, interaction
betweenNandwm10). Similarly, the influence of selfing
rate on the probability of transfer is more pronounced in large populations (File S2, Table 3). By contrast, the influence of mutation rate on the probability of func-tional transfer is much the same across population sizes (File S2, Tables 4 and 5, respectively), suggesting no interaction between these parameters.
Interactions between selfing rate and homozygote fitness:In large populations (N¼1000), the influence of selfing rate on the probability of transfer depends on homozy-gote fitness. When transfer is neutral (i.e.,wm11¼1.00), odds of transfer steadily decline with increased selfing rates. This pattern is reversed when transfer is adaptive (wm11¼1.05 orwm11¼1.10). In these cases, an initial decrease in odds of transfer with slight selfing rapidly disappears, making way for increased odds of transfer with extreme selfing rates.
The increase in odds of transfer with selfing begins at different threshold selfing values depending on homo-zygote fitness. When homohomo-zygote fitness is 1.05, selfing rates $0.8 have better odds of transfer than obligate outcrossers. When transfer is strongly advantageous (wm11 ¼1.10), the increase in odds of transfer with selfing rate begins at lower selfing rates near 0.6. In either case, the probability of transfer greatly increases with selfing rate once the threshold selfing rate is reached (Figure 3).
Interactions between selfing rate and mutation rates:
Above, we showed that increased selfing decreases the probability of transfer under neutrality. Here we show
TABLE 4
The nonlinear effect of increased selfing on the probability of functional gene transfer
Selfing rate OR SE P
0.1 0.90 6.933103 ,0.001
0.2 0.95 7.223103 ,0.001
0.3 0.93 7.113103 ,0.001
0.4 0.95 7.273103 ,0.001
0.5 1.00 7.553103 0.795
0.6 0.98 7.423103 0.007
0.7 1.07 7.953103 ,0.001
0.8 1.24 8.933103 ,0.001
0.9 1.64 1.133102 ,0.001
1.0 2.14 1.413102 ,0.001
TABLE 5
Effect of relative mutation rate on the probability of neutral (wm11¼1.00) functional transfer of mitochondrial genes to
the nucleus in large populations (N¼1000), across heterozygote fitnesses
Odds ratio as a linear function of selfing rate:
Relative mutation rate
wm10
0.90 0.95 1.00
mn?mm 0.98* 1.09* 0.83*
mn¼mm 0.90* 1.29*** 0.75**
that this result depends on therelative rateof nuclear to mitochondrial mutation. We limit our discussion to large populations (N¼1000) with no benefit to transfer (wm11¼1.00).
When the nuclear mutation rate is less than the mitochondrial mutation rate (i.e.,mn>mm), increasing the selfing rate decreases the odds of transfer. On the other hand, when the nuclear and mitochondrial mu-tation rates are equal, or when nuclear mumu-tation rates are greater than mitochondrial mutation rates, there is no strong association between odds of transfer and selfing rates. This result is consistent across heterozy-gote fitness and is thus unlikely due to dominance effects (Table 5).
DISCUSSION AND CONCLUSIONS
The transfer of mitochondrial genes to the nucleus has been ‘‘a key step in stabilizing the transition from an autonomous endosymbiont to a host-dependent mitochondrion’’ (Lynch 2007, p. 308). Despite the many theories presented to explain this phenomenon, its evolution is not well understood. We explored the interaction of key population-genetic parameters (selection, dominance, mutation rates, and population size) with the rate of self-fertilization to influence the rate and probability of functional transfer of mitochondrial genes to the nu-cleus. Here, we address points that are not addressed by our model, outline our major findings, and relate these findings to the existing literature.
Input bias and the possibility of functional transfer: Our model does not address the input of genes to the nucleus or mechanistic factors that may allow or prevent the functional transfer of mitochondrial genes to the nucleus. Nevertheless, these factors could influence phylogenetic patterns. Sheppard et al. (2008) have shown that chloroplast genes are more likely to join
the nucleus via microgametes than macrogametes. This finding does not influence the input of selfing vs.
outcrossing species, but may severely reduce the input of endosymbiont genes to the nuclei of asexually reproducing plants. Elucidating other molecular mech-anisms, such as the rate of reverse transcription, which could bias the input of mitochondrial genes to the nucleus, will further refine our understanding of this process and distribution of transfer events.
A great deal of research has addressed properties that can influence the functional transfer of specific genes from the mitochondria to the nucleus (Daley and Whelan 2005). For example, the hydrophobic hypothesis proposes that hydrophobic proteins cannot easily cross membrane boundaries and are therefore unlikely to be functionally transferred (Von Heijne 1986), while the redox-control hypothesis argues that certain genes expressed in the electron transport chain need to be expressed within their organelles to maintain redox balance (Allen 1993; Race et al. 1999). Both hypotheses provide important clues as to which genes are likely to be retained in organelles. Our understanding of which genes relocate to the nucleus requires additional information regarding which genes may increase fitness when encoded in the nu-cleus rather than in an organelle.
Without a direct fitness benefit to gene transfer, selfing does not increase the probability of gene transfer: Brandvain et al. (2007) showed that clonal and selfing plant taxa transferred more mitochondrial genes to their nuclei than outcrossing taxa or those with mixed mating systems. Our model shows that this pattern cannot be due to neutral evolutionary processes. When there is no advantage to transfer (i.e.,w11¼1.00),
the probability of transfer does not increase with selfing rate in any combination of parameters investi-gated. Thus, the positive association between selfing and transfer cannot be explained with strict neutrality, but is consistent with the hypothesis of positive selection. (We did not examine the possibility that there is a mechanistic bias favoring the input of mitochondrial genes to the nucleus in selfing lineages; its existence is an empirical matter, which is not yet supported by any existing data.)
When transfer is neutral, and nuclear mutation rates are smaller than mitochondrial mutation rates, muta-tional pressure drives the transfer mitochondrial genes to the nucleus:Although animal mitochondria contain tRNAs and rRNAs, these mitochondria are notable for the few protein-coding genes (12–13) they contain, relative to the greater number (30–40) found in those of plants. This pattern is particularly extraordinary be-cause the nonstandard genetic code of animal mitochon-dria has precluded the recent transfer of mitochonmitochon-drial genes to the nucleus, while the process of gene transfer is ongoing in plants (Adamsand Palmer2003). We suggest that this disparity can be largely explained by the relative Figure3.—Effects of the interaction between selfing rate
mutation rates of plant and animal genomes. Lynchet al. (2006, Table 1) have shown that in animalsmm/mnranges
from 8.84 in bilaterian invertebrates to 24.68 in reptiles/ amphibians, while in plantsmm/mn0.05, three or four
orders of magnitude smaller. Assuming the value ofmm/
mnof extant animals reflectsmm/mnin the lineage leading
to animals, which suffered extreme losses of mitochon-drial genes, the difference in the relative mutation rates between plants and animals is thus consistent with, and may contribute to explaining, the observed macro-evolutionary pattern of functional gene transfer. Al-though our model is consistent with this pattern, there are numerous plausible alternative explanations. One possible alternative is that the transfer of essential genes from the mitochondrial genome to the nucleus relaxes selection on modifiers of the mitochondrial mutation rate. For neutral transfers, mutational pressure is en-hanced by outcrossing and is diminished by frequent selfing:The stochastic results show a slight but signifi-cant linear decrease in the probability of transfer with increased selfing when transfer is neutral andmm/mn?
1. In contrast the same conditions produce a nonlinear increase in time to commonness with increased selfing in the deterministic model: for selfing rates between 0 and 0.95, the time to commonness is relatively un-affected by selfing rate, but at extreme selfing rates
.0.95, time to commonness becomes larger. The difference between these results suggests that stochastic processes are important to neutral transfer.
Thus, our results show that with mutational pressure and no other benefit to transfer, the transfer of mito-chondrial genes to the nucleus is more likely in pre-dominantly outcrossing species than in highly inbred taxa. Presumably, with outcrossing, functional nuclear alleles can migrate into cytotypes lacking functional mitochondrial alleles and thus rescue these genotypes, providing a selective benefit for gene retention.
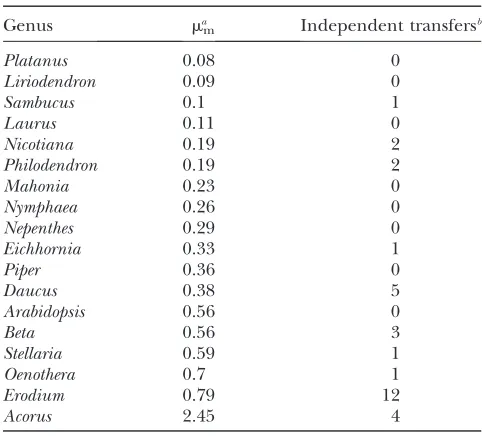
Elevated mitochondrial mutation rates increase the probability of mitochondrial to nuclear gene transfer: Although plant mitochondrial mutation rates are gen-erally much smaller than those of animals, the rates vary widely across taxa (Palmeret al.2000; Choet al.2004; Moweret al.2007). If mitochondrial mutational pres-sure drives the functional transfer of mitochondrial genes to the nucleus, then plants with higher mitochon-drial mutation rates are expected to have transferred more mitochondrial genes to their nuclei than plants with lower mitochondrial mutation rates. Existing data (Table 6) are consistent with this prediction. Although current data are clearly incomplete, there is a significant positive correlation between the mitochondrial muta-tion rate and the frequency of transfers (Spearman’sr¼
0.514,P¼0.029; Kendall’sta¼0.346, Kendall’stb¼
0.397, Kendall’s score¼53,P¼0.037). Although both selfing rate (Brandvainet al.2007) and mitochondrial mutation rate (above) are associated with greater frequencies of functional transfer, these two variables
are uncorrelated (Spearman’sr¼3.79;P¼0.181,N¼
14; data from above and from Brandvainet al. 2007,
File S2), suggesting that each independently influences the probability of functional gene transfer.
When transfer is adaptive, selfing increases the probability of functional gene transfer: Brandvain et al.(2007) found that selfing and clonal plants transfer functional mitochondrial genes to the nucleus more frequently than outcrossing plants. Our model shows that this pattern cannot arise under neutrality. Thus, it is likely that in extant plant lineages, the transfer of mito-chondrial genes to the nucleus is an adaptive process. Although we examined a number of adaptive scenarios that facilitate transfer, we cannot propose a specific fitness advantage to such transfers. This is an empirical question and adaptive explanations must be examined on a case-by-case basis until a general pattern, if one exists, emerges from the data.
The positive relationship between selfing rate and the probability of adaptive transfer sheds light on the roles of linkage and recombination in the fixation of co-adapted gene combinations (Phillips1996; Takahasi and Tajima2005). Coadaptation of mitochondrial and nuclear genes is well documented (Sacktonet al.2003; Randet al.2004, 2006), and thus our results are likely to
TABLE 6
Relationship between mitochondrial mutation rate (mm) and
the number of mitochondrial genes independently transferred to the nucleus
Genus mam Independent transfersb
Platanus 0.08 0
Liriodendron 0.09 0
Sambucus 0.1 1
Laurus 0.11 0
Nicotiana 0.19 2
Philodendron 0.19 2
Mahonia 0.23 0
Nymphaea 0.26 0
Nepenthes 0.29 0
Eichhornia 0.33 1
Piper 0.36 0
Daucus 0.38 5
Arabidopsis 0.56 0
Beta 0.56 3
Stellaria 0.59 1
Oenothera 0.7 1
Erodium 0.79 12
Acorus 2.45 4
Spearman’sr¼0.514,P¼0.029. Kendall’sta¼0.346, Ken-dall’stb¼0.397, Kendall’s score¼53,P¼0.037.
am
mreported in synonymous substitutions per site per
bil-lion years, from Moweret al.(2007). b
be broadly relevant to the evolution of mitochondrial and nuclear genomes. Moreover, although most rele-vant data concerning the transfer of mitochondrial genes to the nucleus in plants have been exhausted in this and a previous article, chloroplast genes also re-locate to the nucleus, much of this transfer is recurrent (Martinet al.1998, 2002), and there is some evidence for genetic interaction between chloroplast and nuclear genes (Grun 1976; Maroof et al. 1992). Thus, the transfer of chloroplast genes to the nucleus provides a unique opportunity for further tests of the epistatic theory of Wadeand Goodnight(2006).
The role of gene interactions in the evolutionary process is an historically contentious issue (Coyneet al. 1997, 2000; Wadeand Goodnight1998; Lopez-Fanjul et al. 1999; Goodnight and Wade 2000). Wright’s ‘‘shifting balance theory’’ emphasizes the fixation of adaptive gene combinations in some subpopulations and the exportation of these adaptive combinations across the larger metapopulation by differential migra-tion (Wadeand Goodnight1991). Goodnight(1987, 1988) has shown that fixing one background in an epistatic system ‘‘converts’’ the segregating variation at another, interacting locus into additive variation, which is subsequently available for a response to selection.
In selfing or clonal organisms, each lineage acts as a subpopulation in which the genetic background is fixed, so that epistatic effects of nuclear genes appear additive and selectable when random drift changes cytoplasmic allele frequencies (Wadeand Goodnight 2006). Our results and the observed pattern of a positive association between selfing/clonal reproduction and number of gene transfers (Brandvain et al. 2007) together suggest that these forces influence the func-tional transfer of mitochondrial genes to the nucleus. Moreover, the variation in mating systems among extant plant taxa and the frequency of cyto-nuclear epistasis for fitness (e.g., Pollak 1991; Galloway and Fenster 1999; Sambattiet al.2008) provide fertile grounds for further tests of the role of selection on epistasis in subdivided populations in evolutionary change.
This article was inspired by an insightful discussion with M. W. Hahn and has benefitted from discussions and comments from S. Dickinson, M. S. Barker, L. C. Moyle, A. O. Richardson, D. van Dyken, T. Cruickshank, P. Nista, P. Zee, D. Drury, D. M. Rand, and two anonymous reviewers. This work was made possible by support from the National Institutes of Health (NIH) training grant, Common Themes in Reproductive Diversity, by a National Science Foundation Predoctoral Fellowship to Y.B., and by NIH grant R01-GM084238 to M.J.W.
LITERATURE CITED
Adams, K. L., and J. D. Palmer, 2003 Evolution of mitochondrial
gene content: gene loss and transfer to the nucleus. Mol. Phylo-genet. Evol.29:380–395.
Adams, K. L., Y. L. Qiu, M. Stoutemyer and J. D. Palmer,
2002 Punctuated evolution of mitochondrial gene content: high and variable rates of mitochondrial gene loss and transfer
to the nucleus during angiosperm evolution. Proc. Natl. Acad. Sci. USA99:9905–9912.
Akman, L., A. Yamashita, H. Watanabe, K. Oshima, T. Shibaet al.,
2002 Genome sequence of the endocellular obligate symbiont of tsetse flies,Wigglesworthia glossinidia.Nat. Genet.32:402–407.
Albert, B., B. Godelle, A. Atlan, R. DePaepeand P. H. Gouyon,
1996 Dynamics of plant mitochondrial genome: model of a three-level selection process. Genetics144:369–382.
Allen, J. F., 1993 Control of gene expression by redox potential and
the requirement for chloroplast and mitochondrial genomes. J. Theor. Biol.165:609–631.
Berg, O. G., and C. G. Kurland, 2000 Why mitochondrial genes are
most often found in nuclei. Mol. Biol. Evol.17:951–961.
Blanchard, J. L., and M. Lynch, 2000 Organellar genes—Why do
they end up in the nucleus? Trends Genet.16:315–320. Bock, R., and J. N. Timmis, 2008 Reconstructing evolution: gene
transfer from plastids to the nucleus. BioEssays30:556–566. Boore, J. L., 1999 Animal mitochondrial genomes. Nucleic Acids
Res.27:1767–1780.
Brandvain, Y., M. S. Barker and M. J. Wade, 2007 Gene
co-inheritance and gene transfer. Science315:1685–1685.
Charles, H., D. Mouchiroud, J. Lobry, I. Goncalvesand Y. Rahbe,
1999 Gene size reduction in the bacterial aphid endosymbiont,
Buchnera. Mol. Biol. Evol.16:1820–1822.
Cho, Y., J. P. Mower, Y. L. Qiu and J. D. Palmer, 2004
Mito-chondrial substitution rates are extraordinarily elevated and vari-able in a genus of flowering plants. Proc. Natl. Acad. Sci. USA
101:17741–17746.
Coyne, J. A., N. H. Bartonand M. Turelli, 1997 Perspective: a
cri-tique of Sewall Wright’s shifting balance theory of evolution. Evo-lution51:643–671.
Coyne, J. A., N. H. Bartonand M. Turelli, 2000 Is Wright’s
shift-ing balance process important in evolution? Evolution54:306– 317.
Daley, D. O., and J. Whelan, 2005 Why genes persist in organelle
genomes. Genome Biol.6:110–115.
Foster, J., M. Ganatra, I. Kamal, J. Ware, K. Makarovaet al.,
2005 The Wolbachia genome ofBrugia malayi: endosymbiont evolution within a human pathogenic nematode. PLoS Biol.3:
599–614.
Galloway, L. F., and C. B. Fenster, 1999 The effect of nuclear and
cytoplasmic genes on fitness and local adaptation in an annual legume,Chamaecrista fasciculate. Evolution53:1734–1743. Gil, R., B. Sabater-Munoz, A. Latorre, F. J. Silvaand A. Moya,
2002 Extreme genome reduction inBuchnera spp.: toward the minimal genome needed for symbiotic life. Proc. Natl. Acad. Sci. USA99:4454–4458.
Goodnight, C. J., 1987 On the effect of founder events on epistatic
genetic variance. Evolution41:80–91.
Goodnight, C. J., 1988 Epistasis and the effect of founder events on
the additive genetic variance. Evolution42:441–454.
Goodnight, C. J., and M. J. Wade, 2000 The ongoing synthesis: a
reply to Coyne, Barton, and Turelli. Evolution54:317–324. Grun, P., 1976 Cytoplasmic Genetics and Evolution.Columbia
Univer-sity Press, New York.
Hosmer, D. W., and S. Lemeshow, 1989 Applied Logistic Regression.
Wiley, New York.
Khachane, A. N., K. N. Timmisand V.dosSantos, 2007 Dynamics
of reductive genome evolution in mitochondria and obligate in-tracellular microbes. Mol. Biol. Evol.24:449–456.
Klasson, L., and S. G. E. Andersson, 2004 Evolution of
minimal-gene-sets in host-dependent bacteria. Trends Microbiol. 12:
37–43.
Lang, B. F., G. Burger, C. J. Okelly, R. Cedergren, G. B. Golding
et al., 1997 An ancestral mitochondrial DNA resembling a eu-bacterial genome in miniature. Nature387:493–497.
Lopez-Fanjul, C., A. Fernandezand M. A. Toro, 1999 The role of
epistasis in the increase in the additive genetic variance after pop-ulation bottlenecks. Genet. Res.73:45–59.
Lynch, M., 2007 The Origins of Genome Architecture.Sinauer
Associ-ates, Sunderland, MA.
Lynch, M., B. Koskellaand S. Schaack, 2006 Mutation pressure
and the evolution of organelle genomic architecture. Science
Maroof, M. A. S., Q. Zhang, D. B. Neale and R. W. Allard,
1992 Associations between nuclear loci and chloroplast DNA genotype in wild barley. Genetics131:225–231.
Martin, W., B. Stoebe, V. Goremykin, S. Hansmann, M. Hasegawa
et al., 1998 Gene transfer to the nucleus and the evolution of chloroplasts. Nature393:162–165.
Martin, W., T. Rujan, E. Richly, A. Hansen, S.Cornelsenet al.,
2002 Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proc. Natl. Acad. Sci. USA99:1246–1251.
Moran, N. A., 2003 Tracing the evolution of gene loss in obligate
bacterial symbionts. Curr. Opin. Microbiol.6:512–518. Moran, N. A., and J. J. Wernegreen, 2000 Lifestyle evolution in
symbiotic bacteria: insights from genomics. Trends Ecol. Evol.
15:321–326.
Mower, J. P., P. Touzet, J. S. Gummow, L. F. Delphand J. D. Palmer,
2007 Extensive variation in synonymous substitution rates in mitochondrial genes of seed plants. BMC Evol. Biol.7:135.
Murcha, M. W., C. Rudhe, D.Elhafez, K. L. Adams, D. O. Daley
et al., 2005 Adaptations required for mitochondrial import fol-lowing mitochondrial to nucleus gene transfer of ribosomal pro-tein S10. Plant Physiol.138:2134–2144.
Palmer, J. D., K. L. Adams, Y. R. Cho, C. L. Parkinson, Y. L. Qiuet al.,
2000 Dynamic evolution of plant mitochondrial genomes: mo-bile genes and introns and highly variable mutation rates. Proc. Natl. Acad. Sci. USA97:6960–6966.
Phillips, P. C., 1996 Waiting for a compensatory mutation: phase
zero of the shifting-balance process. Genet. Res.67:271–283.
Pollak, P. E., 1991 Cytoplasmic effects on components of fitness in
tobacco cybrids. Evolution45:785–791.
Race, H. L., R. G. Herrmannand W. Martin, 1999 Why have
or-ganelles retained genomes. Trends Genet.15:364–370. Rand, D. M., R. A. Haneyand A. J. Fry, 2004 Cytonuclear
coevolu-tion: the genomics of cooperation. Trends Ecol. Evol.19:645–653. Rand, D. M., A. Fryand L. Sheldahl, 2006 Nuclear-mitochondrial
epistasis and Drosophila aging: introgression ofDrosophila simu-lansmtDNA modifies longevity inD. melanogasternuclear back-grounds. Genetics172:329–341.
Rispe, C., and N. A. Moran, 2000 Accumulation of deleterious
mu-tations in endosymbionts: Muller’s ratchet with two levels of se-lection. Am. Nat.156:425–441.
Sackton, T. B., R. A. Haneyand D. M. Rand, 2003 Cytonuclear
co-adaptation inDrosophila: disruption of cytochrome c oxidase ac-tivity in backcross genotypes. Evolution57:2315–2325.
Sambatti, J. B. M., D. Ortiz-Barrientos, E. J. Baack and L. H.
Rieseberg, 2008 Ecological selection maintains cytonuclear
in-compatibilities in hybridizing sunflowers. Ecol. Lett.11:1082– 1091.
Selosse, M. A., B. R. Albertand B. Godelle, 2001 Reducing the
genome size of organelles favours gene transfer to the nucleus. Trends Ecol. Evol.16:135–141.
Sheppard, A. E., M. A. Ayliffe, L. Blatch, A. Day, S. K. Delaney
et al., 2008 Transfer of plastid DNA to the nucleus is elevated during male gametogenesis in tobacco. Plant Physiol. 148:
328–336.
Takahasi, K. R., and F. Tajima, 2005 Evolution of coadaptation in a
two-locus epistatic system. Evolution59:2324–2332.
Tamas, I., L. Klasson, B. Canback, A. K. Naslund, A. S. Eriksson
et al., 2002 50 million years of genomic stasis in endosymbiotic bacteria. Science296:2376–2379.
Timmis, J. N., M. A. Ayliffe, C. Y Huang and W. Martin,
2004 Organelle genomes forge eukaryotic chromosomes. Nat. Rev. Genet.5:123–135.
Ueda, M., M. Fujimoto, S. Arimura, N. Tsutsumiand K.Kadowaki,
2008a Presence of latent mitochondrial signal in gene on mito-chondrial genome. Mol. Biol. Evol.25:1791–1793.
Ueda, M., T. Nishikawa, M. Fujimoto, H.Takanshi, S. Arimura
et al., 2008b Substitution of the gene for chloroplast RPS16 was assisted by generation of a dual targeting signal. Mol. Biol. Evol.25:1566–1575.
vanHam, R., J. Kamerbeek, C. Palacios, C. Rausell, F. Abascal
et al., 2003 Reductive genome evolution inBuchnera aphidicola.
Proc. Natl. Acad. Sci. USA100:581–586.
vonHeijne, G., 1986 Why mitochondria needs a genome. FEBS
Lett.198:1–4.
Wade, M. J., and C. J. Goodnight, 1991 Wright shifting balance
theory—an experimental-study. Science253:1015–1018. Wade, M. J., and C. J. Goodnight, 1998 Perspective: the theories of
Fisher and Wright in the context of metapopulations: when na-ture does many small experiments. Evolution52:1537–1553. Wade, M. J., and C. J. Goodnight, 2006 Cyto-nuclear epistasis:
two-locus random genetic drift in hermaphroditic and dioecious spe-cies. Evolution60:643–659.
Wernegreen, J. J., A. B. Lazarusand P. H. Degnan, 2002 Small
ge-nome ofCandidatus blochmannia, the bacterial endosymbiont of
Camponotus, implies irreversible specialization to an intracellular lifestyle. Microbiology148:2551–2556.
Yamauchi, A., 2005 Rate of gene transfer from mitochondria to
nu-cleus: effects of cytoplasmic inheritance system and intensity of intracellular competition. Genetics171:1387–1396.
Communicating editor: D. M. Rand
APPENDIX 1
Matrices for the deterministic models:
Vector of allele frequencies: F ¼
fm11
fm10
fm00
fM 11
fM 10
fM 00
:
The fitness matrix:
˜
W¼
˜
wm11 0 0 0 0 0
0 w˜m10 0 0 0 0
0 0 w˜m00 0 0 0
0 0 0 w˜M 11 0 0
0 0 0 0 w˜M 10 0
0 0 0 0 0 w˜M 00
The recombination matrix R:The rows and columns of the transition matrices (below) are labeled with the relevant genotypes. In the reproduction matrix (R), values in rowi, columnjrepresent the frequency with which an individual of genotypejparents an individual of genotypei:
m11 m10 m00 M 11 M 10 M 00
m11 S1ð1SÞf11fm1
2 ð1SÞ
f01fm0
2 0 ð1SÞ
fM 1
2 ð1SÞ
fM 0
2 0
m10 S
41ð1SÞ
f11fm1
4 S21ð1SÞ 11fm
4 S41ð1SÞ
f01fm0
4 ð1SÞ
fM 1
4 ð1SÞ
fM
4 ð1SÞ
fM 0
4
R¼m00 0 ð1SÞf11fm1
2 S1ð1SÞ
f01fm0
2 0 ð1SÞ
fM 1
2 ð1SÞ
fM 0
2
M 11 ð1SÞfm1
2 ð1SÞ
fm0
2 0 S1ð1SÞ
f11fM 1
2 ð1SÞ
f01fM 0
2 0
M 10 ð1SÞfm1
4 ð1SÞ
fm
4 ð1SÞ
fm0
4 S41ð1SÞ
f11fM 1
4 S21ð1SÞ 11fM
4 S41ð1SÞ
f01fM 0
4
M 00 0 ð1SÞfm1
2 ð1SÞ
fm0
2 0 ð1SÞ
f11fM 1
2 S1ð1SÞ
f01fM 0
2
:
The mutation matrix m: In the mutation matrix (m), values inijrepresent the frequency with which a zygote of genotypejmutates to genotypei:
m11 m10 m00 M 11 M 10 M 00
m11 ð1mnÞ2 2m
Nucð1mnÞ m2n 0 0 0
m10 0 1mn mn 0 0 0
m¼m00 0 0 1 0 0 0
M 11 mmð1mnÞ2 m
m2mnð1mnÞ mmm2n ð1mmÞð1mnÞ2 ð1mmÞ2mnð1mnÞ m2Nucð1mmÞ
M 10 0 mmð1mnÞ mnmn 0 ð1mmÞð1mnÞ mnð1mmÞ
M 00 0 0 mm 0 0 ð1mmÞ
Supporting Information
http://www.genetics.org/cgi/content/full/genetics.108.100024/DC1
The Functional Transfer of Genes From the Mitochondria to the Nucleus:
The Effects of Selection, Mutation, Population Size
and Rate of Self-Fertilization
Yaniv Brandvain and Michael J. Wade
Y. Brandvain and M. J. Wade 2 SI
FILE S1
Numbers of transfers observed out of 1,000 simulations or each set of parameters. Populations of 1,000 individuals
are shown in A and B. Populations of 500 individuals are shown in C and D. Populations of 100 individuals are shown
in E and F. Results from nuclear mutation rates of 10-3, and 10-4 are shown in A, C, and E. Results from nuclear
Y. Brandvain and M. J. Wade 3 SI
3ELFING2ATE
3ELFING2ATE
OFTR
ANSF
ERSOBSER
VEDOFRUNS
-IT
O
CHONDR
IAL-UTA
TION2
AT
E
. .UCLEAR-UTATION2ATE
!
Y. Brandvain and M. J. Wade 4 SI
3ELFING2ATE
3ELFING2ATE
OFTR
ANSF
ERSOBSER
VEDOFRUNS
-IT
O
CHONDR
IAL-UTA
TION2
AT
E
. .UCLEAR-UTATION2ATE
"
Y. Brandvain and M. J. Wade 5 SI
3ELFING2ATE
3ELFING2ATE
OFTR
ANSF
ERSOBSER
VEDOFRUNS
-IT
O
CHONDR
IAL-UTA
TION2
AT
E
. .UCLEAR-UTATION2ATE
#
Y. Brandvain and M. J. Wade 6 SI
3ELFING2ATE
3ELFING2ATE
OFTR
ANSF
ERSOBSER
VEDOFRUNS
-IT
O
CHONDR
IAL-UTA
TION2
AT
E
. .UCLEAR-UTATION2ATE
$
Y. Brandvain and M. J. Wade 7 SI
3ELFING2ATE
3ELFING2ATE
OFTR
ANSF
ERSOBSER
VEDOFRUNS
-IT
O
CHONDR
IAL-UTA
TION2
AT
E
. .UCLEAR-UTATION2ATE
%
Y. Brandvain and M. J. Wade 8 SI
3ELFING2ATE
3ELFING2ATE
OFTR
ANSF
ERSOBSER
VEDOFRUNS
-IT
O
CHONDR
IAL-UTA
TION2
AT
E
. .UCLEAR-UTATION2ATE
&
Y. Brandvain and M. J. Wade 9 SI
FILE S2
Interactions with population size
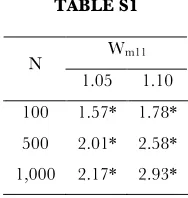
TABLE S1
Wm11
N
1.05 1.10 100 1.57* 1.78* 500 2.01* 2.58* 1,000 2.17* 2.93*
Combined influence of homozygote and population size on the probability of functional transfer, represented by the Odds Ratio (relative to neutral homozygotes). To enforce balance, comparisons were limited to neutral or deleterious heterzygote fitness (wm10≤ 1.00). Note that increases in homozygote fitness have a greater influence on the odds ratio
Y. Brandvain and M. J. Wade 10 SI
TABLE S2
Wm10
N
0.95 1.00 100 1.00 1.02- 500 1.04* 1.09* 1,000 1.06* 1.12*
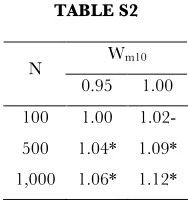
Combined influence of heterozygote and population size on the probability of functional transfer, represented by the Odds Ratio (relative to strongly deleterious heterozygote, i.e. wm10 = 0.90). To enforce balance, comparisons were
limited to neutral or deleterious heterozygote fitness. Note that increases in heterozygote fitness have a greater influence on the odds ratio in large than small populations, suggesting an interaction among theses parameters. * signifies p < 0.05. Above, all results significant at the 0.05 level are also significant at the 0.001 level.
Y. Brandvain and M. J. Wade 11 SI
TABLE S3
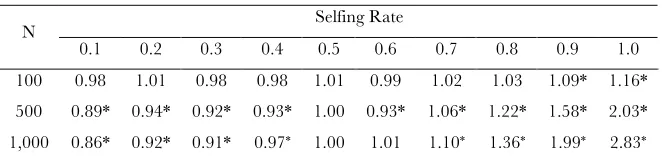
Selfing Rate N
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 100 0.98 1.01 0.98 0.98 1.01 0.99 1.02 1.03 1.09* 1.16* 500 0.89* 0.94* 0.92* 0.93* 1.00 0.93* 1.06* 1.22* 1.58* 2.03* 1,000 0.86* 0.92* 0.91* 0.97* 1.00 1.01 1.10* 1.36* 1.99* 2.83*
Combined influence of selfing rate and population size on the probability of functional transfer, represented by the Odds Ratio (relative to obligate outcrossing in a population of size N). Note that large selfing rates have a greater influence on the odds ratio in large than small populations, suggesting an interaction among theses parameters. * signifies p < 0.05. Above, all results significant at the 0.05 level are also significant at the 0.001 level.
Y. Brandvain and M. J. Wade 12 SI
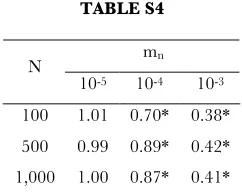
TABLE S4
mn
N
10-5 10-4 10-3
100 1.01 0.70* 0.38* 500 0.99 0.89* 0.42* 1,000 1.00 0.87* 0.41*
Combined influence of nuclear mutation rate and population size on the probability of functional transfer, represented by the Odds Ratio (relative to low nuclear mutation rates, mn = 10-6). Note that the influence of nuclear
mutation rate on the odds ratio is relatively constant across population sizes, suggesting no interaction among these parameters. * signifies p < 0.05. Above, all results significant at the 0.05 level are also significant at the 0.001 level.
Y. Brandvain and M. J. Wade 13 SI
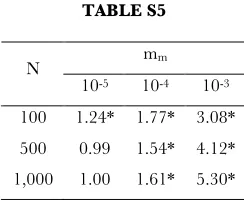
TABLE S5
mm
N
10-5 10-4 10-3
100 1.24* 1.77* 3.08* 500 0.99 1.54* 4.12* 1,000 1.00 1.61* 5.30*
Combined influence of mitochondrial mutation rate and population size on the probability of functional transfer, represented by the Odds Ratio (relative to low nuclear mutation rates, mm = 10-6). Note that the influence of