台大農藝系 遺傳學 601 20000 Chapter 20 slide 1
CHAPTER 20
Transposable Elements
Peter J. Russell
台大農藝系 遺傳學 601 20000 Chapter 20 slide 2
General Features of Transposable Elements
1. Transposable elements are divided into two classes on the basis of their
mechanism for movement:
a. Some encode proteins that move the DNA directly to a new position or replicate the DNA to produce a new element that integrates elsewhere. This type is found in both prokaryotes and eukaryotes.
b. Others are related to retroviruses, and encode reverse transcriptase for making DNA copies of their RNA transcripts, which then integrate at new sites. This type is
found only in eukaryotes.
2. Transposition is nonhomologous recombination, with insertion into DNA that
has no sequence homology with the transposon.
a. In prokaryotes, transposition can be into the cell’s chromosome, a plasmid or a phage chromosome.
b. In eukaryotes, insertion can be into the same or a different chromosome.
3. Transposable elements can cause genetic changes, and have been involved in
the evolution of both prokaryotic and eukaryotic genomes. Transposons may:
a. Insert into genes.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 3
Transposable Elements in Prokaryotes
1. Prokaryotic examples include:
a. Insertion sequence (IS) elements.
b. Transposons (Tn).
台大農藝系 遺傳學 601 20000 Chapter 20 slide 4
Insertion Sequences
Animation: Insertion Sequences in Prokaryotes
1. IS elements are the simplest transposable elements found in
prokaryotes, encoding only genes for mobilization and insertion of its
DNA. IS elements are commonly found in bacterial chromosomes and
plasmids.
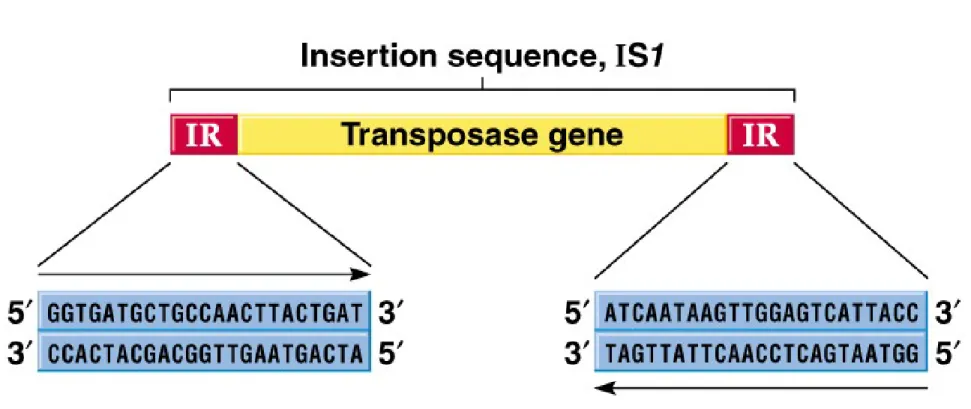
2. IS elements were first identified in
E. coli’s
galactose operon,
wheresome mutations’ were shown to result from insertion of a DNA
sequence now called IS1 (Figure 20.1)
3. Prokaryotic IS elements range in size from 768 bp to over 5 kb.
Known
E. coli
IS elements include:
a. IS1 is 768 bp long, and present in 4–19 copies on the E. coli
chromosome.
b.
IS2 has 0–12 copies on the chromosome, and 1 copy on the F plasmid.
c.
IS10 is found in R plasmids.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 5
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 6
5. Integration of IS elements may:
a. Disrupt coding sequences or regulatory regions.
b. Alter expression of nearby genes by the action of IS element promoters.
c. Cause deletions and inversions in adjacent DNA.
d. Serve as a site for crossing-over between duplicated IS elements.
6. When an IS element transposes:
a. The original copy stays in place, and a new copy inserts randomly into the chromosome.
b. The IS element uses the host cell replication enzymes for precise replication. c. Transposition requires transposase, an enzyme encoded by the IS element. d. Transposase recognizes the IR sequences to initiate transposition.
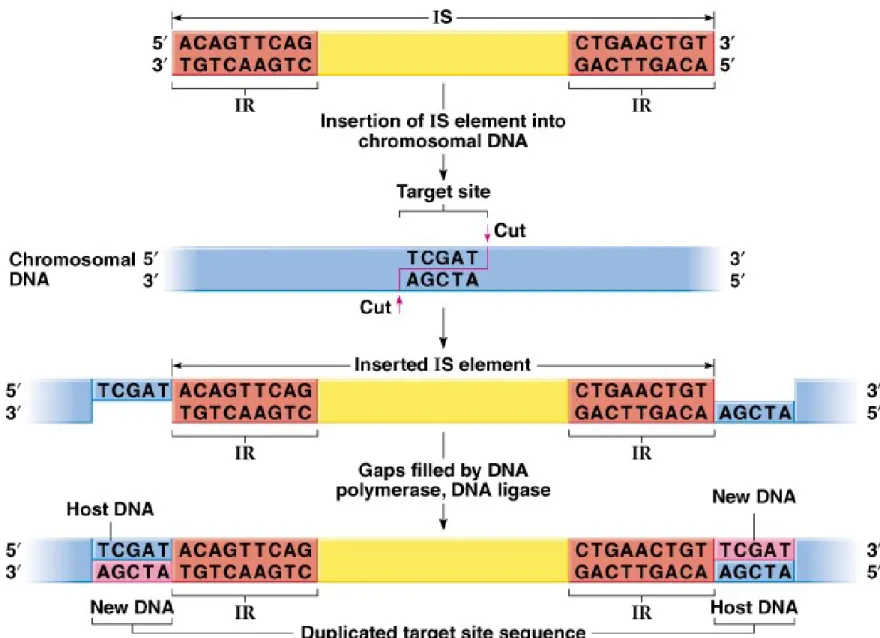
e. IS elements insert into the chromosome without sequence homology (illegitimate recombination) at target sites (Figure 20.2).
i. A staggered cut is made in the target site, and the IS element inserted.
ii. DNA polymerase and ligase fill the gaps, producing small direct repeats of the target site flanking the IS element (target site duplications).
台大農藝系 遺傳學 601 20000 Chapter 20 slide 7
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 8
Transposons
1. Transposons are similar to IS elements, but carry additional genes, and have a
more complex structure. There are two types of prokaryotic transposons:
a. Composite transposons carry genes (e.g., antibiotic resistance) flanked on both sides by IS elements (IS modules).
i. The IS elements are of the same type, and called ISL (left) and ISR (right). ii. ISL and ISR may be in direct or inverted orientation to each other.
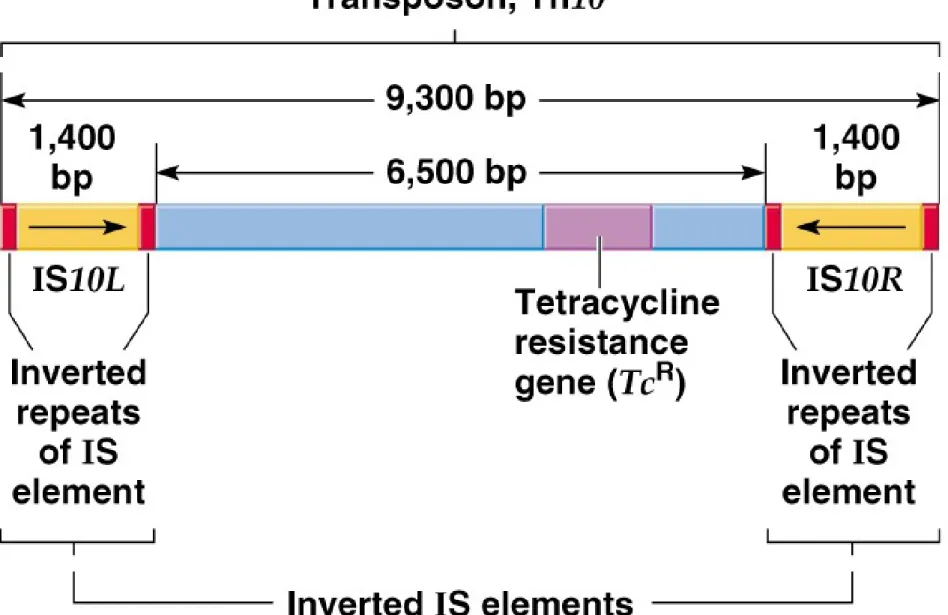
iii. Tn10 is an example of a composite transposon (Figure 20.3). It is 9.3 kb, and contains:
(1) 6.5 kb of central DNA with genes that include tetracycline resistance (a selectable marker).
(2) 1.4 kb IS elements (IS10L and IS10R) at each end, in an inverted orientation.
iv. Transposition of composite transposons results from the IS elements, which supply transposase and its recognition signals, the IRs.
(1) Tn10’s transposition is rare, because transpose is produced at a rate of ,1 molecule/generation.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 9
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 10
b.Noncomposite transposons also carry genes (e.g.,
drug resistance) but do not terminate with IS
elements.
i. Transposition proteins are encoded in the central
region.
ii. The ends are repeated sequences (but not IS
elements).
iii. Noncomposite transposons cause target site
duplications (like composite transposons).
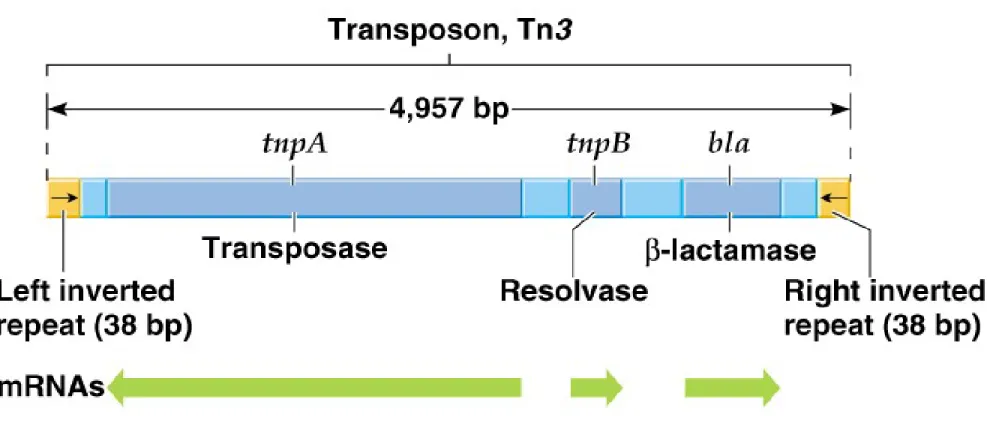
iv. An example is Tn3.
(1) Tn3’s length is about 5 kb, with 38-bp inverted terminal
repeats.
(2) It has three genes in its central region:
(a) bla encodes β-lactamase, which breaks down ampiciliin.
(b) tnpA encodes transposase, needed for insertion into a new site.
(c) tnpB encodes resolvase, involved in recombinational events needed for transposition (not found in all transposons).
台大農藝系 遺傳學 601 20000 Chapter 20 slide 11
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 12
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 13
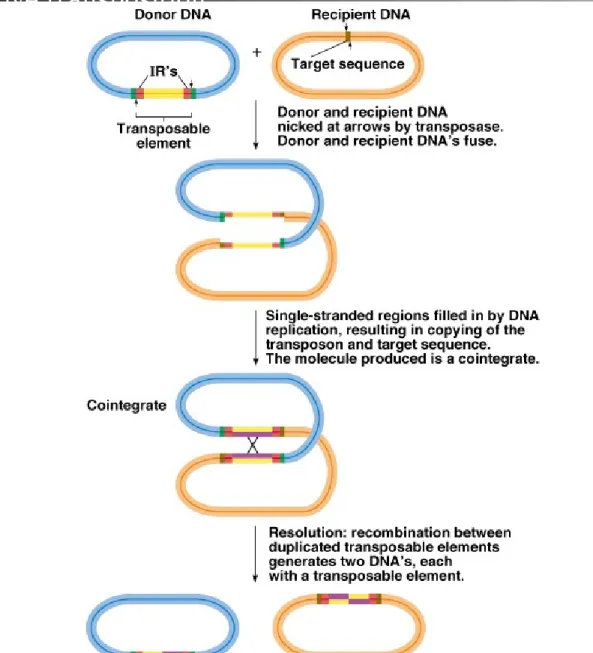
2. Models have been generated for transposition:
a. Cointegration is an example of the replicative transposition that
occurs with Tn3 and its relatives (Figure 20.6).
i. Donor DNA containing the Tn fuses with recipient DNA.
ii. The Tn is duplicated, with one copy at each donor-recipient DNA
junction, producing a cointegrate.
iii. The cointegrate is resolved into two products, each with one copy
of the Tn.
b. Conservative (nonreplicative) transposition is used by Tn10, for
example. The Tn is lost from its original position when it transposes.
3. Transposons cause the same sorts of mutations caused by IS elements:
a. Insertion into a gene disrupts it.
b. Gene expression is changed by adjacent Tn promoters.
c. Deletions and insertions occur.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 14
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 20.6 Cointegration model for transposition of a transposable element by
台大農藝系 遺傳學 601 20000 Chapter 20 slide 15
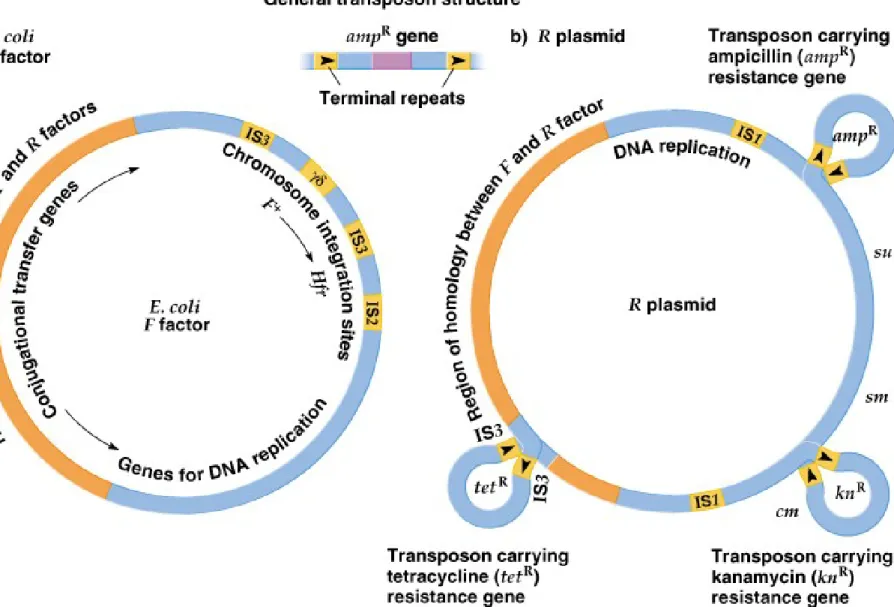
IS Elements and Transposons in Plasmids
1. Bacterial plasmids are extrachromosomal DNA capable of self-replication.
Some are episomes, able to integrate into the bacterial chromosome. The
E.
coli F
plasmid is an example (Figure 20.7):
a. Important genetic elements of the F plasmid are:
i. tra genes for conjugal transfer of DNA from donor to recipient. ii. Genes for plasmid replication.
iii. 4 IS elements: 2 copies of IS3, 1 of IS2, and 1 of γδ (gammadelta). All have homology with IS elements itt the E. coli chromosome.
b. The F factor integrates by homologous recombination between IS elements, mediated by the tra genes.
2.
R
plasmids have medical significance, because they carry genes for resistance to
antibiotics, and transfer them between bacteria (Figure 20.7).
a. Genetic features of R plasmids include:
i. The resistance transfer factor region (RTF), needed for conjugal transfer. It includes a DNA region homologous to an F plasmid region, and genes for plasmid-specific DNA replication.
ii. Differing sets of genes, such as those for resistance to antibiotics or heavy metals. The resistance genes are transposons, flanked by IS module-like sequences, and can replicate and insert into the bacterial chromosome.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 16
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 17
Bacteriophage Mu
1. Temperate bacteriophage Mu (mutator) can cause mutations when it transposes.
Its structure includes:
a. A 37 kb linear DNA in the phage particle that has central phage DNA and unequal lengths of host DNA at the ends (Figure 20.8).
b. The DNA’s G segment can invert, and is found in both orientations in viral DNA.
2. Following infection, Mu integrates into the host chromosome by conservative
(non-replicative) transposition.
a. Integration produces prophage DNA flanked by 5 bp target site direct repeats.
b. Flanking DNA from the previous host is lost during integration.
c. The Mu prophage now replicates only when the E. coli chromosome replicates, due to a phage-encocled repressor that prevents most Mu gene expression.
3. Mu prophage stays integrated during the lytic cycle, and replication of Mu’s
genome is by replicative transposition.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 18
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 20.8 Temperate bacteriophage Mu genome shown in (a) as in phage particles and
台大農藝系 遺傳學 601 20000 Chapter 20 slide 19
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 20.9 Production of deletion or inversion by homologous recombination between
台大農藝系 遺傳學 601 20000 Chapter 20 slide 20
Transposable Elements in Eukaryotes
1. Rhoades (1930s) working with sweet corn, observed interactions between two
genes:
a. A gene for purple seed color, the Al locus. Homozygous mutants (a/a) have colorless seeds.
b. A gene on a different chromosome, Dt (dotted) that causes seeds with genotype a/a
Dt/-- to have purple dots.
i. Dt appears to mutate the a allele back to the Al wild-type in regions of the seed, producing a dotted phenotype.
ii. The effect of the Dt allele is dose dependent.
(1) One dose gave an average of 7.2 dots per seed. (2) Two doses gave an average of 22.2 dots/seed. (3) Three doses gave an average of 121.9 dots/seed. c. Rhoades interpreted Dt as a mutator gene.
2. McClintock (1940s-50s), working with corn
(Zea mays)
proposed the existence
of “controlling elements” that regulate other genes and are mobile in the
genome.
3. The genes studied by both Rhoades and McClintock have turned out to be
transposable elements, and many others have been identified in various
eukaryotes.
a. Most studied are transposons of yeast, Drosophila, corn and humans.
b. Their structure is very similar to that of prokaryotic transposable elements.
c. Eukaryotic transposable elements have genes for transposition and integration at a number of sites, as well as a variety of other genes.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 21
Transposons in Plants
Animation: Transposable Elements in Plants
1. Plant transposons also have IR sequences, and generate short direct target site repeats. 2. The result of transposon insertion into a plant chromosome will depend on the properties
of the transposon, with possible effects including:
a. Activation or repression of adjacent genes by disrupting a cellular promoter, or by action of transposon promoters.
b. Chromosome mutations such as duplications, deletions, inversions, translocations or breakage. c. Disruption of genes to produce a null mutation (gene is nonfunctional).
3. Several families of transposons have been identified in corn, each with characteristic numbers, types and locations.
a. Each family has two forms of transposon. Either can insert into a gene and produce a mutant allele.
i. Autonomous elements, which can transpose by themselves. Alleles produced by an autonomous element are mutable alleles, creating mutations that revert when the transposon is excised from the gene.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 22
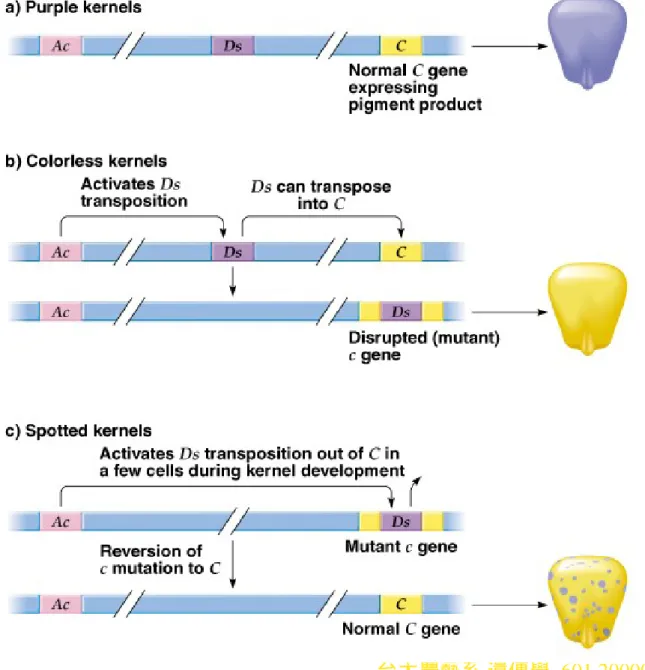
4. Multiple genes control corn color, and classical genetics indicates that a
mutation in any of these genes leads to a colorless kernel. McClintock
studied the unstable mutation that produces spots of purple pigment on
white kernels (Figure 20.10).
a. She concluded that spots do not result from a conventional mutation,
but from a controlling element (now Tn).
b. A corn plant with genotype
c/c
will have white kernels, while
C/--
will
result in purple ones.
i. If a reversion of
c
to
C
occurs in a cell, that cell will produce purple
pigment, and hence a spot.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 23
iii. McClintock concluded that the
c
allele resulted from
insertion of a “mobile controlling element” into the
C
allele.
(1) The element is
Ds
(dissociation), now known to be a
nonautonomous transposon.
(2) Its transposition is controlled by
Ac
(activator), an
autonomous transposon (Figure 20.11).
c. McClintock’s evidence of transposable elements did not fit the
prevailing model of a static genome. More recent studies have
confirmed and characterized the elements involved.
i. The
Ac-Ds
system involves an autonomous element
(Ac)
whose insertions are unstable, and a nonautonomous element
(Ds)
whose insertions are stable if only
Ds
is present.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 24
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 25
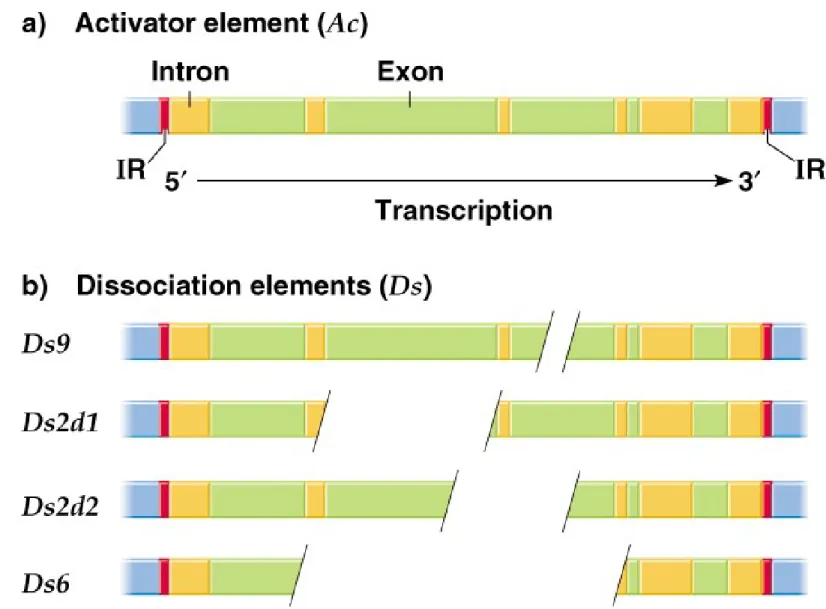
iii.
Ac
is 4,563 bp, with 1 1-bp imperfect terminal IRs and 1 transcription
unit producing a 3.5 kb mRNA encoding an 807 amino acid transposase.
Insertion generates an 8-bp target site duplication (Figure 20.12).
iv.
Ac
activates
Ds
to transpose or break the chromosome where it is
inserted.
v.
Ds
elements vary in length and sequence, but all have the same terminal
IRs as
Ac,
and many are deleted or rearranged versions of
Ac.
vi. Unique to corn transposons, timing and frequency of transposition and
gene rearrangements are developmentally regulated.
vii.
Ac
transposes only during chromosome replication, and does not leave a
copy behind. There are two possible results of
Ac
transposition,
depending on whether the target DNA has replicated or not (Figure
20.13).
-
(1) If
Ac
transposes during replication into a replicated target site, its
chromatid’s donor site will be empty since that copy of
Ac
has
inserted elsewhere. In the homologous donor site on the other
chromatid, a copy will remain. There is no net increase in copies of
Ac.
(2) Transposition to an unreplicated chromosome site also leaves one
donor site empty (and the other with a copy of
Ac).
The DNA into
which
Ac
inserts will then be replicated, resulting in a net gain of one
copy of
Ac.
viii. Replication of Ds is the same, except that the transposition protein is
台大農藝系 遺傳學 601 20000 Chapter 20 slide 26
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 20.12 The structure of the Ac autonomous transposable element of corn and of
台大農藝系 遺傳學 601 20000 Chapter 20 slide 27
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 28
5. In Mendel’s wild-type (SS) peas the starch grains are
large and simple, while in wrinlded peas
(ss)
they are
small and fissured.
a. SS seeds contain more starch and less sucrose than
ss
seeds.
b. The sucrose difference makes
ss
seeds larger, with higher water
content, so that when dried they are wrinided.
c. One type of starch-branching enzyme (SBEI) is missing in
ss
plants, reducing their starch content.
d. The SBEI gene corresponding to the
s
allele has a 0.8 kb
台大農藝系 遺傳學 601 20000 Chapter 20 slide 29
Ty
Elements in Yeast
1.
Ty
elements share characteristics with bacterial transposons:
a. Terminal repeated sequences.
b. Integration at non-homologous sites.
c. Generation of a target site duplication (5 bp).
2.
Ty
element is diagrammed in Figure 20.14:
a. It is 5.9 kb including 2 terminal direct repeats of 334 bp, the long terminal repeats (LTR) or deltas (δ).
b. Each delta contains a promoter and transposase recognition sequences.
c. Ty elements encode one 5.7 kb mRNA beginning at the delta 5’ promoter (Figure 20.14). d. There are two ORFs in the mRNA, designated TyA and TyB, encoding two different
proteins.
e. Ty copy number varies between yeast strains, with an average of about 35.
3.
Ty
elements also share similarities with retroviruses, ssRNA viruses that replicate
via dsDNA intermediates.
a. Ty elements transpose by making an RNA copy of the integrated DNA sequence, them making DNA using reverse transcriptase. This DNA can integrate at a new
chromosomal site. Evidence for this includes:
i. An experimentally introduced intron in the Ty element (which normally lacks
introns) was monitored through transposition. The intron was removed, indicating an RNA intermediate.
ii. Ty elements encode a reverse transcriptase.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 30
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 31
Drosophila
transposons
1. It is estimated that 15% of the
Drosophila
genome is mobile! These
transposons fall into different classes:
a. The
copia
retrotransposons include several families, each highly
conserved and present in 5-100 widely scattered copies per genome
(Figure 20.15).
i. All
copia
elements in
Drosophila
can transpose, and there are
differences in number and distribution between fly strains.
ii. Structurally,
copia
elements are similar to yeast
Ty
elements:
(1) Direct LTRs of 276 bp flank a 5 kb DNA segment.
(2) The end of each LTR has 17 bp inverted repeats.
(3) An
RNA
intermediate and reverse transcriptase are used for
transposition.
(4) Virus-like particles (VLPs) occur with
copia.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 32
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 20.15 Structure of the transposable element copia, a retrotransposon found in
台大農藝系 遺傳學 601 20000 Chapter 20 slide 33
b. P elements cause hybrid dysgenesis, a series of defects
(mutations, chromosomal aberrations and sterility) that
result from crossing certain
Drosophila
strains (Figure
20.16).
i. A mutant lab strain female (M) crossed with a
wild-type male (P) will result in hybrid dysgenesis.
ii. A mutant lab strain male (M) crossed with a
wild-type (P) female (reciprocal cross) will have normal
offspring.
iii. Thus, hybrid dysgenesis results when
台大農藝系 遺傳學 601 20000 Chapter 20 slide 34
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 35
iv. The model is based on the observation that the M strain has no
P
elements, while the haploid genome of the P male has about 40
copies.
(1)
P
elements vary from full-length autonomous elements
through shorter versions resulting from a variety of internal
deletions.
(2)
P
element transposition is activated only in the germ line.
(3) The F1 of an M female crossed with a P male have
P
elements inserted at new sites, flanked by target site repeats.
(4)
P
elements are thought to encode a repressor protein that
prevents transposase gene expression, preventing
transposition.
(5) Cytoplasm in an M oocyte lacks the repressor, and so when
fertilized with P-bearing chromosomes, transposition occurs
into the maternal chromosomes, leading to hybrid dysgenesis.
v.
P
elements are used experimentally to transfer genes into the germ
line of
Drosophila
embryos. For example (Figure 20.18):
(1) The wild-type
rosy (ry)
gene was inserted into a P element,
cloned in a plasmid and microinjected into a mutant
ry/ry
strain.
(2) Insertion of the recombinant P element into the recipient
chromosome introduced the
ry
allele, and produced wild-type
台大農藝系 遺傳學 601 20000 Chapter 20 slide 36
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 20.17 Structure of the autonomous P transposable element found in
Drosophila
台大農藝系 遺傳學 601 20000 Chapter 20 slide 37
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 20.18 Illustration of the use of P elements to introduce genes into the
Drosophila
台大農藝系 遺傳學 601 20000 Chapter 20 slide 38
Human Retrotansposons
1. Retrotransposons also appear to be present in mammals. For example, a very
abundant human SINE repeat (short interspersed sequence) is the Mu
family, named for the
AluI
restriction site in its sequence.
a. Mu sequences are about 300 bp, repeated 300,000-500,000 times in the human
genome (up to 3% of total human DNA).
b. Sequences are divergent, related but not identical.
c. Each Mu sequence is flanked by 7-20 bp direct repeats.
d. At least a few Mu sequences can be transcribed, and the model is that
transcriptionally active Mu sequences are retrotransposons that move via an
RNA intermediate.
e. A human case of a genetic disease, neurofibromatosis, provides some evidence.
i. Neurofibromas (tumorlike growths on the body) result from an autosomal
dominant mutation.
ii. In a patient’s DNA, an unusual Mu sequence was detected in one of the
introns of the neurofibromatosis gene.
iii. The resulting longer transcript is incorrectly proessed, removing an exon
from the mRNA and producing a nonfunctional protein.
iv. Neither parent had this Mu sequence in the neurofibromatosis gene.
v. Divergent Mu sequences made it possible to track this particular version to
an insertion event in the germ line of the patient’s father.
台大農藝系 遺傳學 601 20000 Chapter 20 slide 39