I wish to heartily thank my advisors, Drs. Louise Ball and Douglas Crawford-Brown, for years of support of many kinds and continuous intellectual stimulation, and to Dr. Johnny L. Carson for his guidance and encouragement as a Reader for this report. The laboratories of Drs. C.-C. Chang and B. V. Madhukar, Dept. of Pediatrics and Human Development, Michigan State University, graciously hosted me in my struggles to use cell culture techniques to explore a bit the relationship which is the subject of this
paper. Their challenging discussions and generosity with space, time and
materials is an example of the best in open and cooperative science. Thanks especially to Dr. Koichiro Namata who patiently continued some of these experiments when time limitations did not permit me to do so.This work is dedicated to the memory of my beloved aunt, Pauline Ross,
ABSTRACT
An apparent linkage betwen gap junctional intercellular communication (GJIC) and promotion of neoplastic growth is frequently mentioned in the literature of carcinogenesis and used as the rationale for several in-vitro assays for tumor promoting compounds. The biological theory behind such an association has not been adequately developed, however, limiting the ability of researchers to optimally design and interpret in-vitro experiments measuring GJIC. In the present work, such theory is first descriptively
developed, and then applied to an evaluation of the utility of currently used assays for assessing tumor promoting ability of test compounds. Results of these experiments and theoretical considerations are used to explicitly state the epistemic basis and strength of claims for a causative relationship between decreased gap junctional commimication and cancer. Next, a type of
experiment is proposed which meets the stated conditions under which a result of diminished GJIC is strongly indicative of increased risk of cancer via promotional effects. Some observations from preliminary attempts to
perform this type of experiment with two heterologous cell systems are
presented. Based on the earlier discussion of promotional theory, and the
TABLE OF CONTENTS
Page Introduction 1
Methods and Detailed Background
I. GJIC as a "Bioeffect" Indicator/Development of In-Vitro Assay 25 n. Links Between In-Vitro Assay Design and Modelling Studies 48 in. Modelling Rat Liver Tumor Promotion by Phenobarbital 51
Results
I. Preliminary Attempts to Perform Heterologous Assays 66
n. Model Calculations of Phenobarbital Promotion in Rat Liver 72
Discussion 78
The multi-stage, multi-pathway development of normal tissue cells into tumours and invasive, metastatic cells is now a widely accepted concept (Weinstein, 1984), having evolved from earlier, more simplistic ideas of "the carcinogen as mutagen" (Ames et al, 1973), discrete initiation and promotion stages (Boutwell, 1974), oncogene control of the process (Bishop, 1985) and hypotheses of cancer as a disease of stem cells (Trosko and Chang, 1988) or a dysftmction of differentiation (Potter, 1978). This acceptance comes from evidence not only of the genotypic and phenotypic diversity of cells within a given tumor, but also that invasive, metastic cells have, unfortunately, an amazing array of options as to how they acquire increasingly tumorigenic phenotypes (Hill, 1990).
The result has been a tangle of complexly linked mechanisms facing those attempting to do quantitative mechanistically-based assessment of
carcinogenic risk from a given occupational or environmental exposure, even in the extremely rare case of short-term exposures to pure substances. In the case of radiation exposure, some success has been achieved in
particles or neutrons act as the principal genotoxic agent (over and above a background mutation rate), as well as a possible promoting agent, via cell killing and removal of contact inhibition of growth. In low radiation exposure cases such as household radon, however, radiation-induced cell killing would be extremely rare and it is more likely that other types of environmental exposures, such as certain components in tobacco smoke, would provide more significant promotional effects.
Earlier asstimptions that chemical carcinogens would all prove to be mutagenic by one or more in-vitro genotoxicity tests have eroded in the past decade (Tennant et al, 1987) as data from a widening array of test compounds has been incorporated into statistical correlations, motivating increased
interest in epigenetic mechanisms of carcinogenesis (Trosko and Chang, 1990; Williams, 1989). Two of the most widely observed effects of xenobiotic
Gap junctions are assemblies of "connexin" proteins inserted into the cellular membrane, lined up with and tightly boimd non-covalently to
connexin assemblies on a neighbouring cell. A single gap junctional channel, or "connexon", consists of two hemi-channels, each a hexagonal arrangement of six connexin molecules contributed by one cell of the contacting pair.
Under normal physiological conditions, and when cells are not proliferating, these junctions are open to form a large pore between the cytoplasms of the two cells (Lowenstein,1981). Several closely related connexin proteins, varying in molecular weight from about 26 to 43 kiloDaltons, have been found in organ tissues, with one type usually predominating in each organ
(Jannsen-Timmen et al, 1986). In culture, however, both established cell lines
and primary cells express primarily the 43kDa connexin (B. V. Madhukar, personal communication). Molecules of up to about 1700 daltons can be passed directly from cell to cell via these junctions, allowing for tissue homeostasis and growth regulation in an organ or cell culture by "contact inhibition" (Loewenstein, 1981). Such physiological changes as lowered intracellular pH or increased intracellular calcium, indicative of impending cell death or lowered metabolic rate, cause the rapidly reversible down-regulation of gap junctional intercellular communication (GJIC), probably due to physical closure of the pore as proposed by Zampighi and Unwin (1979), based on their electron diffraction structural studies.
ͣ
Gap junctions seem to be in a fully open state when the intracellular calcium is below 10-7 M, but dose down in a graded fashion, within minutes, as Ca++ concentration is increased, allowing only smaller and smaller
fluorescent probes to pass, until not even small ions are transmitted at intracellular concentrations above 10-5 M Ca++ (Rose, Simpson and Loewenstein, 1977). This mode and range of closure conditions may help account for threshold effects observed in many communication-dependent cellular processes and for the phenomenon of selective GJIC, to be discussed later. Increased intracellular Ca++ also causes a marked change in the , distribution and packing of connexon particles on the membrane over the same concentration range (Peracchia, 1978), with open junctions rather irregularly arranged, while those which are uncoupled eire arranged in a tightly packed, ordered array.
Phosphorylation of one or more serines on the carboxy-terminus of the connexin molecules, on the cytoplasmic side of the membrane, seems to induce a conformational change which causes a rearrangement of the
example, are known to mimic phosphatidyl inositol (Stagg and Fletcher,
1990), binding to and activating protein kinase C, thus closing junctions. This
effect is reversible within hours after removal of TPA from culture media
(Trosko and Chang, 1990). Some cell lines, such as C3H10T1 /2 mouse
fibroblast cell line, become refractory to TPA's effects after several days of
exposure and resume normal levels of communication (Boreiko et al, 1991).
Activation of protein kinase A by addition of cyclic-AMP apparently results in
phosphorylation at a different site on the connexin molecule, and
up-regulates junctional commimication (Saez et al, 1986; Stagg and Fletcher,
1990). Finally, mutations of genes for the connexiris themselves or the
extracellular matrix proteins (fibronectin, laminin, proteoglycans among
others) and their receptors, which facilitate strong cell-cell contacts, produce
the most irreversible forms of reduced GJIC, often in more highly metastatic
cells (Ren etal, 1990).
The first observations that tumor promoting compounds were capable of
reducing gap jvmctional communication were made in 1979 (Yotti et al;
Murray and Fitzgerald). Since that time it has been shown that in many
classes of chemical toxins (tumor promoters, teratogen, neurotoxin,
pesticides) there is a strong correlation between the level of tumor promotion
or other toxicity observed in-vivo and the down-regulation of GJIC as
^^^^^m^s^?^
closely related compoimds, such as PCB's (Tsushimo et ai, 1989) and certain
pesticides (Klatmig, Ruch and Weghorst, 1990), the stronger the in-vivo
promoting activity, the more pronounced the down regulation of cell-cell
communication, causing Trosko, Jone and Chang (1987) to propose reduced
GJIC as a major mechanism of reversible (non-cytotoxic) tumor promotion
and neurotoxicity. A finding of in-vitro down-regulation of intercellular
communication by a suspect chemical may therefore be a useful predictor for
tumor promotion and other in-vivo toxic activities.Promotion in Carcinogenesis
The treatment of arumals with low doses of only initiating (genotoxic)
agents genaerally causes a very small increase in tumor incidence. Over 90%
of the genetic lesions identified in liver of diethylnitrosamine-treated rats by
cellular expression of a placental marker enzyme, are either repaired or
eliminated through cell death within two weeks of treatment (Dragan and
Pitot, 1992). The number of "foci", or clonal lesions arising from single
initiated cells, may decline by another order of magnitude over a period of
seven to eight months unless a promoting stimulus is given (Goldsworthy,
Campbell and Pitot, 1984). Only when conditions in the organ cause cell
proliferation either by cell loss or damage (ie, partial hepatectomy, carbon
them are lethal to the cell; some of them are "promotionally relevant" (Potter, 1983), or affect a gene related to cell survival or proliferation; still others are promotionally irrelevant, having no survival advantages in the presence of promoting agents. In rare cases, multiple lesions occur in the genome of a cell which enable it to escape growth control and progress toward a tumor with no added promotional stimulus. Promotionally relevant
mutations are often associated with inappropriate activation of oncogenes (Chiao et al, 1990) or inactivation of tumor suppressing "anti-oncogenes", such as p53 (Levine, Monandand and Finlay, 1991) or retinoblastoma gene (Weinberg, 1991). Growth-controlling or differentiation-inducing gene products have been termed "chalones" (Bullough, 1962) and normally play tissue-specific roles in homeostasis and suppression of altered cellular phenotypes including rapid proliferation.
A second (or more) irreversible event is almost always required to yield cells capable of promoter-independent progression to a tumor (Land, Parada and Weinberg, 1983). In the example of myeloid leukemia (Sachs et aL 1980), the steps toward malignancy can be viewed as an accumulation of
constitutive or irreversible changes in the expression of regulators of growth and differentiation and their cellular receptors, freeing cells to proliferate in the absence of normal physiological inducers. The exceptions to this
ͣͣͣͣ
"
ͣͣͣ
' ,:- \ ^
occurring in systems where gap jtmctional commimication cannot function. In one such case, single oncogene transfection (SV40) of non-communicating
but growth regulated normal human breast epithelial Type I (stem) cells
(Chang et al, impublished results) created cells which were tumorigenic when
injected in immune-suppressed mice. Transplantation ofoncogene-transfected kerotinocytes to mice, in another example, caused tumor formation when the cells were allowed to grow in plastic barriers isolating
them from direct contact with normal skin cells (Dotto, Weinberg and Ariza,
1988). If one views initiation as causing at least partial constitutive expression
of a growth factor gene (Castagna and Martelle, 1989) or repression of a growth suppressor in a cell which makes a receptor for them, then reducedcommunication with normal cells will prevent transmission of
growth-controlling signals to initiated cells unable to generate them internally. The
most firmly established cellular mechanistic role for gap junctionalintercellular communication (GJIC) is the activation of cyclic-AMP dependent
protein kinases by signals passed from cells with hormone receptors toneighboring cells without them (Stagg and Fletcher, 1990).
this means an effective promoter would increase not only the number of "altered hepatic foci" (AHF) exhibiting altered morphology or enzyme
function, but also the total number of cells in fod. A non-cytotoxic promoter such as phenobarbital aids in the selection of characteristics which enable cells to survive and proliferate in their preneoplastic, partially transformed state
(Moore etal, 1983).
It is dear, however, that the process of promotion requires more than simple mitogenesis. Agents such as nitrate and ethylene dibromide which
cause increased mitotic rates are not sufficient to cause the selection and
growth of altered fod in initiated rat liver (Ledda-Columbano et al, 1989). Only conditions of compensatory growth, following partial hepatectomy or cytotoxic doses of CCL4^ were found by these investigators to support the growth of initiated cells to fod. Under conditions of compensatory growth, gap junctions are reduced by more than 50% in relative surface area of hepatocytes, are smaller and more irregularly distributed (Sugie, Mori and Takahashi, 1987). Phenobarbital causes exactly the same type of
ultrastructural changes in gap junctions, a finding which corresponds well with observations of reduced communication by both normal and initiated hepatocytes upon exposure to phenobarbital in culture (Klaunig, Ruch and Weghorst, 1990; Trosko et al, 1990). These findings point to a role for
10
Based on observations of relative cancer risk in families carrying a single mutation in an anti-oncogen (Li-Fraumeni Syndrome and Retinoblastoma gene), Moolgavkar and Knudson (1981) proposed a model of two rare and critical mutations releasing a cell from normal growth and differentiation controls and thus causing cancer. The role of the promoter here is simply to raise the probability of cells with the critical second mutation by increasing the population of cells carrying the first one, without regard to the detailed biological factors affecting the difference between cell division and death rates (a - P). According to Moolgavkar (1989), raising a while holding a - P
constant will cause a smaller number of large initiated clones, or
preneoplastic foci, while lowering p will increase the mmiber of foci, but decrease their size. Under conditions of high initiation frequency, he predicts that there will be a higher risk of tumors from an a -increasing promoter
than from a P -decreasing one.
Changes in phenotypic expression are increasingly recogiuzed as having major effects on survival of initiated cells and their clonal expansion. Schulte-Hermann and co-workers (1989) proposed an "over-response
phenotypically changing foci are those most dependent on continued presence of the promoter in the early stages of carcinogenesis. Pitot et al (1989) also found the highest rates of cell proliferation in foci which showed altered expression of multiple marker enzymes in response to phenobarbital promotion. Removal of promoter caused a rapid regression of most foci in terms of phenotypic expression and elimination of many cells through increased rates of apoptosis (Schulte-Hermarm et al, 1989). Many initiated cells did survive, however, as altered fod could be rapidly induced again upon readministration of promoter.
The Importance of Communication Between Normal and Initiated Cells Constitutive loss of GJIC alone is not sufficient to impart the properties of loss of growth control or contact sensitivity to cells. The undifferentiated stem cells of some organs, where identified, do not communicate through gap junctions with each other or with their differentiated daughter cells (Trosko,
1989) and yet are only stimulated to proliferate imder conditions of tissue loss or damage. The fact that these cells are undifferentiated, however, may mean that they have other characteristics critical to tumorigenesis, including
prevention of normal cellular senescence and death. The
12
It was first thought that all tumor cells had lost the ability to communicate junctionally with all other cells (Yamasaki, 1990), and there are numerous ultrastructiiral studies showing altered arrangements of gap junction particles in neoplastic cells from a variety of tissues (Janssen-Timmen et al, 1986; Ren et al, 1990). Recently, however, it was recognized by Yamasaki (1990) and others (Trosko et al, 1990) that many of these timior cells communicate among themselves, but are unable to carry on GJIC with neighboring differentiated, normal cells. This selective communication often appears even in preneoplastic cells (Mehta, Bertram and Loewenstein, 1986; Trosko et
al, 1990) not yet sufficiently altered to cause tumor formation. In an example
of normal and initiated cells with at least partial heterologouscommunication, Bertram (1990) has shown that clonally-derived initiated C3H10T1/2 cells, which will readily form transformed focal growths when grown by themselves in culture, can be contact inhibited (ie, focal growth is prevented) when grown interspersed with a sufficient number of non-initiated cells of the same parent cell line. A similar contact inhibition of focal growth can be further enhanced in the heterologous (normal and
initiated) BALB/c 3T3 cell system by the addition of cyclic-AMP, known to up-regulate GJIC (Yamasaki and Katoh, 1988).
fibroblasts, and, in-vivo, by growth of tiunors following injection of cells into immunologically compromised mice. In some cases, adding cyclic-AMP to culture medium can up-regulate GJIC sufficiently to bring about contact sensitivity of previously non-communicating tumor cells (Chang,
unpublished results). Injection or transplantation of normal cells along with transformed ones can suppress in-vivo expression of transformed phenotype as well as the growth of tumors which occur when only initiated cells only are used (Terzaghi-Howe, 1987 and 1990; Dotto, Weinberg and Ariza, 1988), indicating that GJIC loss is not an all-or-nothing phenomenon. There is, however, a strong positive relationship between the extent and irreversibility of GJIC loss and the tumorigenic aggressiveness of a particular cell line
(Nicolson, Dulski and Trosko, 1988; Ren et al, 1990; Hill, 1990).
A number of possible mechanisms may be involved in the selective communication between normal and preneoplastic cells. If the initiating mutation has caused alteration of a calcium-responsive protein, the rearrangement of gap junction particles in the membrane in response to varying Ca++ concentration (Peracchia, 1978) may follow a different pattern in normal and initiated cells. Mutation of the genes for the connexin
14
cases (Jongen et al, 1990) and, if altered by an irutiating mutation, may limit the number and type of junctions which are able to form between normal and initiated cells.
Growth Regulation, Cell Cycling and Clonal Selection for Altered Phenotypes All of the above results indicate that the ability to commurucate with
differentiating normal tissue cells should be an important indicator of the ability to control growth of a pre-neoplastic cell or clone. Moreover, for GJIC to perform this role, there must be either: 1) one or more growth regulators produced by the normal cells which are transmissable to other cells via gap jimctions, or 2), a factor or factors produced by the initiated/preneoplastic cells which can counter the effect of growth inhibitors. Prehn (1992) hypothesizes that a "mitosis preventer" is produced by the differentiating cells and affects both the differentiated and stem cells in an organ, causing an appropriate rate of cell proliferation for a particular organ dependent on its rate of cell
preventer" due to a further genetic lesion, cell loss or diminished GJIC, would be needed to bring about a steady increase in cell number. The principal
difficulty wth this hypothesis is that it is known that many stem cells are intrinsically non-communicating cells (Trosko and Chang, 1989) and
therefore would not be subject to regulation by the mitosis preventer, imless by hormonal (extracellular) control.
The simplest interpretation of the observed growth phenomena is that there is a growth regulator or tumor suppressor molecule, or secondary signals from such a suppressor, which can be transferred through gap junctions of normally communicating cells. Differentiated cells which
produced this regulator in normal quantities would continue to proliferate in a controlled way and be resistant to transformation even if GJIC were lost either in a reversible manner through addition of chemicals or other
physiological stresses affecting protein expression, phosphorylation, etc., or, less reversibly, through covalent changes to the genome. However, cells which had even imstable changes in their ability to produce this regulator or suppressor (p53 is the archetypal example), would have a selective growth advantage under reduced GJIC conditions, leading to a cell population which has shifted in its distribution of growth rates and/or production of a
particular gene product, as described by R. P. Hill (1990).
16
importance to the impact of genomic changes which result in even partial reduction of GJIC, as may occur in some initiating events. With irreversibly reduced GJIC and diminished control of proliferation, the cells in the upper end of a distribution of tvmiorigenic phenotypes (related to over- or under-expression of a certain gene product) would have an increased oportunity to acquire other traits (which may accord further survival and growth
advantages) through loss of heterozygosity, oncogene activation, etc. While tumor promoting compounds are not mutagens, they are almost certainly responsible for selective pressure in favor of spontaneous genetic changes which produce short term growth advantages. It should be no surprise that many of the most aggressive cells found in a heterogeneous tumor are also the most resistant to a variety of cytotoxic drugs (Williams, 1989). Other phenotypes which have been used as markers for altered foci in rat liver (and which increase with time of exposure to promoters) include expression of enzymes for xenobiotic metabolism and detoxification (Pitot, 1990), iron or glycogen storage capacity and expression of extracellular matrix proteins
(Williams, 1989). Any of these may be relate'd to the appearance of strongly
phenotypically altered fod with more "distinct borders" and a greatly reduced
rate of normal programmed cell death, or apoptosis, as described bySchulte-Hermann etal (1990).
hybridized a clone of BALB/c 3T3 cells which lost GJIC at confluence and were
highly transformable, with a normally commimicating and transformation
resistant clone, the resulting cells were GJIC negative (producing significantly
reduced quantities of connexin 43 after confluence), yet resistant to
transformation. The most reasonable explanation for this result is provided
by the fact that such hybrid cells have approximately two times the normal
number of chromosomes of each parental cell. Assuming that one or two
copies of a tumor suppressor gene caused expression of enough product to
protect from loss of growth control, even under conditions which would
transform the parental cell line, multiple genetic "hits" would be required on
the larger number of copies of the tvmior suppressor or protooncogene to
result in transformation, or irreversible loss of growth control (BCatoh and
Yamasaki, 1991). Under such conditions of resistance to loss of all autologous
tumor suppressor, the ability to communicate becomes irrelevant to growth
control, ie., contact sensitivity would not be required to prevent cell
overgrowth. If GJIC is indeed critical to normal cell suppression of
transformed growth, this phenomenon of autonomous growth regulation
would not be expected to occur in cells with a normal chromosome number
. JllWJIil^W,)!'!!' ^•^^^v,'iMm^mk^s^W9lllge^^,is!sm
-18
Assays for Gap Junctional Communication
In-vitro assays which have been used to assess the degree of intracellular
communication have evolved to meet the diverse needs of the laboratories
which have worked on the regulation of GJIC as related to promotion and other toxic effects. Among the first assays used was the measurement of electrophysiological conductivity in excitable cells (Furshpan and Potter, 1959; Spray, Harris and Bennett, 1977; Enomoto et al, 1981). Metabolic cooperation assays (Murray and Fitzgerald, 1979; Pitts and Finbow, 1986) are more
complicated in concept and execution, but have been widely used, as they do not depend on toxic effects specific to excitable cells. Two specially engineered strains of a single cell line, designed to be deficient at different points in a metabolic pathway, are intermixed in culture. Cell death results when an added precursor compound is converted successively by the two
communicating cell strains into a cytotoxic product. Addition of subcytotoxic doses of GJIC-inhibiting chemicals thus "saves" some of the cells, as they are less able to transfer the partially metabolized compound.
Perhaps the simplest and most generalizable type of measurement involves the simple transfer of radioactively or fluorescently labelled
compounds between contacting cells (Rose, Simpson and Loewenstein, 1977).
Classically, this has been done by microinjecting single cells in culture and
then determining the nvimber of cells in a communicating network or the1981). More recently, the "FRAP" (fluorescence recovery after
photobleaching) method (Wade et al, 1986) involves the laser bleaching of an individual cell in a uniformly labelled culture, followed by the photometric determination of the intensity of dye transferred via gap junctions to the bleached cell from other cells with which it is in contact. This test is rapid and quantitative, but the long incubation with dye required can stress some cell types and it is implemented in only a few labs with access to the
instrumentation required.
The "scrape-loading/dye transfer" assay (El-Fouley et al, 1987) is the
simplest assay to date, but as currently used, is at best semi-quantitative. This method of assessing GJIC consists of scoring a thin line through a monolayer of confluent, cultured cells while the medium has been briefly replaced with a solution containing Lucifer Yellow. The dye is able to enter the cells with cut membranes and spreads rapidly from the initially loaded cells to any
20
a test compound can, however, yield an order of magnitude estimate of a threshold for down-regulation of communication.
Finally, in order to asses the role of "heterologous" intercellular
communication, Bertram (1990) has looked at the ability of "normal" (but immortalized) C3H/10T1/2 mouse fibroblasts to suppress the growth of initiated C3H/10T1/2 cells as focal growths in mixed cultures. When a constant proportion of the two tj^es of cells was plated at different starting total densities, plates which were closer to confluence and could quickly establish commimication had fewer focal growths per initiated cell than those which required several generations of proliferation before reaching
confluence. The interpretation of these experiments is complicated,
however, by the different number of total divisions per initiated cell to reach confluence starting with different plating densities, so that it is not possible to distinguish the effects of differences in heterologous GJIC from possible effects of differences in generation number.
With a few exceptions, the correlation between tumor promoting activity for a compound and its ability to down-regulate GJIC in-vitro in established cell lines is high enough that investigators using the above assays often
by a cell and/or the rate constants of reversible or irreversible transitions critical to the process of transformation must be understood sufficiently to model at least a subset of such pathways and to optimally design an assay to emulate the cellular conditions of transformation in an intact organ.
Concluding Remarks
A picture of cellular pathways to tumorigenidty is emerging in which the neat boundaries between the stages of initiation, promotion and progression are perhaps damaged beyond repair. Investigators clearly still use the term "initiation" to refer to irreversible genetic events occuring early in the
process, but, in some cases see little distinction between these and later genetic changes leading to irreversible transformation (Moolgavkar, 1989) and the beginning of the phase of tumor progression. Rapid cell proliferation
(generally considered a promotional phase), for example, may be underway at the time a spontaneous or exogenously induced mutation occurs, helping to rapidly stabilize it and increase the number of daughter cells bearing the
mutation. Further, the nature and extent of genomic changes can vary widely depending on the specific agent and dose, the growth phase and the presence
of hormones or other environmental factors (Williams, 1989).
22
Additionally, it emphasizes the importance of contact with surrounding growth-arrested communicating cells in suppressing the transformed phenotype, and inducing the selective elimination of cells with altered phenotype in reconstructed trachea. If any of the initiating events caused by radiation or chemical genotoxins result in at least partial loss of gap
junctional communication, additional genetic or epigenetic changes causing loss of autologous growth regulation in the same cell would result in an essentially "self-promoting" system, as would these same types of mutational
events in the reverse order.
It has generally been considered that initiating events are those affecting oncogenes related to terminal differentiation (Trosko and Chang, 1990). Just as likely (in the absence of information on relative instability of genes
controlling various phenotypes related to carcinogenesis) is an initial event directly or indirectly affecting levels of GJIC; or, as in the case of inherited Li-Fraumeni syndrome (Levine, Momand and Finlay, 1991), a mutation causing loss of heterozygosity in a gene for a tumor suppressor protein, such as p53; or the structure and function of extracellular matrix (ECM) proteins. These could result in subtly expressed phenotypes generally associated with promotion, progression or metastasis, respectively, but not of biological
further impaired a cell's ability to differentiate and senesce, 4) altered
expression and arrangement of ECM proteins or their receptors, or 5) acquired invasive or migratory properties (also likely to be related to the extracellular matrix or membrane-bound enzymes).
In order to assess the importance of GJIC in carcinogenesis it is necessary to model its effects on cells at any given stage in a pathway and quantitate them as closely as possible. If one accepts the clonal selection theory model of R. P. Hill (1990) and others, it will be important to show whether the kind of attributes being selected for which increase proliferation imder reduced GJIC conditions would also be related to constitutive loss of growth control, the ability to evade terminal differentiation, and increasingly invasive
properties. If so, then the only phenotype which would not be selected for, ie, the only stage at which reversible reduction of GJIC by environmental agents would not increase the probability of neoplastic transformation, would be an irreversibly communication-deficient cell.
24
quantitatively the risk of in-vivo neoplastic transformation due to exposure
to a non-cytotoxic promoter, using dose/response data for phenobarbital in rat
I. Evalulation of Gap Junctional Communication as a "Bioeffect" Indicator for Carcinogenic Hazard Identification and Development of a Proposed In-Vitro Assay
A. Evaluation of GJIC as a Bioeffect Indicator
In order to qualitatively evaluate the importance of down-regulation of GJIC in carcinogenesis, and thus its usefulness as a bioeffect indicative of carcinogenicity, the "criteria for rationality" described by Crawford-Brown et
al (1992) were applied to an extensive sampling of literature in the field.
These criteria—completeness, utility, observed effect and causality—provide
bases for a rating of this bioeffect as "Hi/Med/Lo/No" in its empirical
relationship to a certain stage and mechanism in carcinogenesis.Theory-based inferences concerning the role of diminished GJIC in tumor
promotion was developed in the introductory chapter of this work. With no
evidence that reversible decreases in gap junctional communication have anygenotoxic effects, GJIC-regulating compounds cannot be classified as initiating
agents. Reduced GJIC can, however, by the mechanisms discussed previously,
increase the rate of reversible cell proliferation, thus acting in a promotional
26
In order to draw an etiologic link between down-regulated GJIC and carcinogenesis, a couple of major assumptions must be made: 1) that
hormonal (extracellular) and other means of cell-cell communication,
function with sufficient independence of GJIC to be able to separate them experimentally and conceptually, both in-vitro and in-vivo (In general, all other actions of a putative promoting substance related to proliferation of initiated cells must be controlled for or eliminated.), and 2) that there exists one or more agents, transmissible through gap junctions, which can directly or indirectly modify proliferative behavior of cells. While there may not be a clear dividing line in some cases in-vivo between the processes of promotion to a tumor and progression of a tumor, the work described in this study will focus on GJIC's effects on reversibly increased proliferation of
non-tumorigenic (preneoplastic) cells.
Completeness
(Terzaghi-Howe, 1987,1990; Dotto, Weinberg and Ariza, 1988) and seven
in-vitro studies (Mehta, Bertram and Loewenstein, 1986; Yamasaki et al, 1987,
1988; Bertram, 1990; Terzaghi-Howe, 1990; Trosko et al, 1990; Katoh and
Yamasaki, 1991) of the effect of heterologous communication between normal and initiated/preneoplastic cells on suppression of altered cell growth. In addition, two review articles ( Trosko and Chang, 1988; Yamasaki, 1990) which summarize reports of GJIC regulating effects of over 120 chemicals and
oncogenes, were used to develop a broader perspective of work in the field and major questions to be resolved.
The eighteen reports of homologous GJIC effects represent a wide variety of test chemicals, both endogenous and xenobiotic, several different test cell types, and numerous variations on the two widely-used methods of assessing junctional communication: metabolic cooperation and dye transfer. With a few exceptions, these studies were undertaken to assess the effects on GJIC of chemicals which are known tumor promoters, anticarcinogens or toxins, and would be predicted on mechanistic grounds to regulate intracellular
communication.
In summary, the studies on which this evaluation of GJIC as a bioeffect indicator are based appear to be representative of the published studies over the past decade. A shortcoming, however, is the investigator bias toward the selection of test compounds expected to have GJIC regulating effects, and a
publication and reviewer bias (Trosko and Chang, 1988) toward reporting of
28
results for compounds which give positive resvilts, ie, down-regulation of
GJIC. For completeness, GJIC studies are rated as "Medium" in quality.
Utility
In-vitro studies. In all of the studies examined in detail, established
cell-culture techniques were applied and details of the history and preparation of
cell lines were reported. Methods for measurement of GJIC varied, but were
closely related to the metabolic cooperation and microinjection/dye transfer
techniques widely used since the late 1970's. Most experiments were
well-designed in terms of dose and treatment time ranges, sample size and
inclusion of negative controls. In all but two of the homologous
communication studies, investigators report their results with standard
deviations or confidence intervals. However, only two of the heterologous
and six of the homologous communication studies present measures of
statistical significance (perhaps a reflection of the general lack of
understanding of such measures in the biomedical research community). In
few of the studies were positive controls—previously studied
compounds-included for comparison. Among the studies using focal growth and
expression of altered phenotype as an endpoint, only Terzaghi-Howe (1990)
used incubation in the presence of conditioned media to exclude the
possibility of effects due to extracellular communication.
Perhaps the greatest shortcoming of the reports of effects of test compounds
established (immortalized) cell lines rather than evaluating GJIC regulating
effects in more biologically relevant normal primary cells and initiated cells.
In only one study (Trosko et al, 1990), is there a comparison of the effects of a
known liver promoter, phenobarbital, on homologous communicationbetween primary hepatocytes and on heterologous GJIC between normal and
initiated cells, with only a single dose tested.
Animal studies. In contrast to the in-vitro studies, animal studies
intrinsically involve heterologous cellular interations, unless specifically
engineered barriers for transplants (Dotto, Weinberg and Ariza, 1988) prevent
them. These experiments necessarily involve the use of endpoints such as
altered cell growth and morphology, or even tumor formation, rather than
direct measurements of intracellular communication, to assess the effects of
GJIC. This has the advantage of a more direct linkage to the actual cellular
changes known to occur in carcinogenesis,thereby increasing the utility of
such studies.The ultrastructural studies relating gap junctional cell surface area to
promoter treatment and cell transformation state, as well as the initiated cell
"transplantation" type experiments used small numbers of animals and
tissue samples. While this leaves open the question of the representativeness
of the measured responses, statistically significant effects were observed in
each study, relative to a variety of relevant controls for exposure conditions
-ͣ'-ͣsyv'Hjja'JIt^
30
The in-vitro homologous GJIC studies, which used only "normal" primary or established cell lines, were of generally mediimi to high quality design for measurements of communication. However, they suffer somewhat from inadequate statistical analysis and, in the absence of studies relating decreased commurucation among normal cells to growth of initiated cells in the same heterologous system, they have questionable relevance to the context of tumor promotion through proliferation of initiated cells. In-vitro and in-vivo studies of preneoplastic growth suppression by normal cells were of high quality, and were more immediately relevant to tumor promotion biology, but often lacked meastu-es of statistical significance. Overall, these studies would rate "Med/Hi" in utility.
Observed Effect
As described under previous criteria, the methods, cells lines and test compounds used in these studies were diverse, preventing pooling of data. Many of the results, though strikingly positive, are reported without
measures of statistical significance, preventing pooling of p-values. However, the results of these studies and the review by Yamasaki (1990), prelinunarily indicate an association between tumor promoting activity in-vivo and down-regulation of GJIC, as well as between "anticarcinogenic" activity or
(suppression of focal growth) and up-regulation of GJIC. One notable
(2,3,7,8-tetrachlorodibenzo-p-dioxin), a powerful rat liver promoter and transformer of immortalized cell
lines (Abemethy et al, 1985; Yang et aL 1992), which has no reported effect on
homologous communication at promoting or transforming concentrations.
The promoting effects of TCDD may be entirely receptor mediated, as are its
other toxic effects, or it is possible that GJIC down-regulation may only be
evident in tests of heterologous commimication.
In the compilation of homologous GJIC assay results for 112 chemicals
prepared by Trosko and Chang (1988), 70 were known promoters, teratogens
or complete carcinogens which were demonstrated to diminish homologous
intracellular communication. Another 28 compounds, not yet tested for
promoting ability, but including many closely related to known promoters,
also showed GJIC regulatory activity. Among the 14 chemicals which were
inactive in regulating GJIC, only one was a weak promoter at subcytotoxic
levels, three were promoters only at cytotoxic concentrations (where
compensatory hyperplasia is known to cause generalized loss of GpC), three
were untested for promotional activity, and seven were known
non-promoters. The authors acknowledge a strong bias in the listed selections for
known promoting and GJIC positive compounds and did not attempt to
survey other toxic effects of the reported chemicals. Thus, though the
apparent correlation between promoting activity and GJIC regulation appears
32
bias in selection of test chemicals made the apparent correlation between
in-vivo carcinogens and Ames-positive tests very high.
The more mechanistically defining heterologous communication studies,
though seldom well-analyzed statistically, report very strong effects for both
GJIC up- and down-regulating chemicals and oncogenes as related to their
growth effects on initiated cells in-vitro. The most contextually relevant
studies of 1) structural effects on gap jimctions following in-vivo promoter
treatment (Sugie, Mori and Takahashi, 1987) and 2) suppression of growth of
irutiated cells to contact with normal tissue cells (Dotto, Weinberg and Ariza,
1988; Terzaghi-Howe, 1987,1990), report strong effects, though with limited
statistical analysis in the latter case.
On the basis of the data available and with the above limitations, a
judgment of "Medium" is given to the strength of association between
down-regulation of GJIC and carcinogenicity.
Causality
Taken by themselves, the in-vitro experiments assessing the effects of
specific compounds on homologous GJIC present no basis for a warrant
stronger than a "common cause" for cancer promotion and diminished
intracellular communication. Even the status of this warrant is dependent
on the confirmation of the current apparently high correlation by further
dose/response data for GJIC down-regulation often bear a strong resemblance
to actual tumor promotion dose/response curves (concentration range of
effect, threshold and maximum effect levels), and the behavior of both phenomena is reversible, temporal correlations are lacking, as in the case ofTPA (Boreiko et aL 1989) or have not been studied.
Heterologous in-vivo experiments, using preneoplastic (or even
neoplastic) growth as the measured endpoint and controlling forconfounding effects by design, allow for a much stronger warrant of causality. The study features which permit this include the use of cell systems more closely resembling the conditions in an in-vivo organ containing some
distribution of initiated cells, and the use of an endpoint (focal growth) which
is considered to be on the direct pathway to tumor formation (Watanabe and
Williams, 1978; Peraino et al, 1984). Though there remain some questions about the generalizability of the observed growth effects in established cell lines (Mehta, Bertram and Loewenstein, 1986; Yamasaki and Katoh, 1988) to other cell systems including normal primary cells, and non-clonal initiated cells, the in-vivo results of Terzaghi-Howe (1987) and Dotto, Weinberg and Ariza (1990) corroborate the overall pattern seen in the in-vitro experiments, making a limited case for a warrant of "operational causality". Theexperimental design to be described in section IB, below, is an attempt to
establish such operational causality as firmly as possible by carefully
34
antecedent conditions for relating GJIC down-regulation to carcinogenesis are
relevant.
Providing Support for Claims of Carcinogenicity
In the sequence of "tasks" described by Crawford-Brown and Brown (1992) required for concluding that an observed bioeffect, such as reduced gap
junctional communication, is related to carcinogenesis, the completion of several "working tables" is recommended. In Working Table 2 (Fig. 1), the characteristics of GJIC data observed in the general context of in-vitro tissue culture studies is summarized, using the results of the evaluations for completeness, utility, observed effect and causality. The completion of Working Table 3 (Fig. 2) for GJIC uses the results of the etiological theory developed, and empirical correlations from the data evaluated in Table 2, to classify an observation of decreased GJIC according to the taxonomic scheme for claims of carcinogenicity shown in Figure 3 and to give an overall rating
to the evidence. Substances which caused reduced commimication would be
classified as "partial carcinogens" in the "helper role" of modifying the conditions (through non-genotoxic mechanisms) under which a another agent with genotoxic activity exerts its carcinogenic effect. Strength of
evidence for this classification is rated "Med". Finally, Working Table 5 (Fig. 4) is used to summarize the premises and conditions under which
WORKING TABLE 2. DATA CHARACTERISTICS FOR OBSERVATIONAL CONTEXTS
CONTEXT NO.
(from Crawford-Brown and Brown, 1992) Data Category/Item Completeness
(Hi/Me/Lo/No) Utility* (Hi/Me/Lo/No) Observed Effect^ (Hi/Me/Lo/No) 1 Causality' (AA/CC/EC/OC) Exposure Context-
Organism-Effect Specific Specific Measurement Measurement |
Tumor Response TRl TR2
ͣ
Biophysical Effects ^
BEicejjc)
BE2
M^cL
lli/M<d.
Mi^d,
Pharmacodynamics
PDI PD2
•
Host Charactoistks
n HCl
FIGURE 2
WORKING TABLE 3. INTRA-CONTEXT SUPPORT FOR CLAIMS OF CARCINOGENICITY*
CONTEXT NO.
Claims of Carcinogenicity^
Reference Strategy I.O.» Increases Incidence of Cancer
Classification(s) Stage(s) Mechanism(s)
Complete Partial Mixer Helper Neo. Conv. Neo. Devel. Geno-toxic Non-genotoxlc Direct Empirical (D.E.)
1 Semi-Empirical
Extrapolation (S.E.E.) Empirical Correlation (E.C.) • Theory-based Inference (T.B.I.) Existentiali Insight (E.I.)
1 Column Summary
1 Overall Summary
Top half of each entry is completed using dose-response data observed in the context. Bottom half of each entry includes
"floater-data" as well (see text).
Choices for cell entries are Hi/Me/Lo/No. See text of Section 5.1. This assignment is made independently of the assignment
of intellectual obligation.
"Intellectual Obligation" (Hi/Me/Lo/No). See text of Section 5.1. This assignment is made independently of the assignment in the cells of the table (footnote 2).
Classification
Staoe(s)
M6chanlsm(s)
of Carcinogenicity ,^ ^
(from Crawford-Brown and Brown, 1992)
Substance increases tlie incidence of cancer.
Substance Is a direct carcinogen
Complete | | Partial I
Carcinogen i | Carcinogen '
I
Substance inducesI Neoplastic
! Conversion
Neoplastic
Development
Substance induces
transitions through
Genotoxic i j Non-genotoxic
Mechanisms i ! Mechanisms
!__________________________________JSubstance is an indirect carcinogen
Mixing
Agent
[helping
Agent
I____________^F._____________I
Substance modifies
Neoplastic |
Conversion I
Neoplastic
Development
Substance modifies
I Genotoxic j j Non-genotoxic
FIGURE 4
WORKING TABLE 5. SUPPORT FOR INTER-CONTEXT EXTRAPOLATION PREMISES
CONTEXT NO. TO CONTEXT NO.
1 Reference
1 Strategy .
Exposure to BSDR Conversion
BSDR to Effect Conversion
Host Factors Environmental
Conditions
Intra(Inter)-Subject Variability
Direct Empirical (D.E.)
Semi-Empirical Extrapolation
(S.E.E.)
Empirical
Correlation (E.C.) Theory-based Inference (T.B.I.) Existential Insight
(E.I.)
1
Overall
Assessment
the target context of in-vivo carcinogenicity are valid. For in-vitro
nneasurement of GJIC, the broad category of "environmental conditions" will be discussed, in the form of the following list.
Assumptions and Antecedent Conditions for Inter-Context Extrapolation from Observation of GTIC Down-Regulation to In-Vivo Carcinogenicity
1. Heterologous Cell System. As it is the growth of iiutiated cells which is etiologically related to the production of tumors, experiments should
evaluate communication between "normal" cells and initiated or
preneoplastic cells of the same tissue or a biologically relevant, contiguous
tissue.
2. Cell Types. Established cell lines, such as C3H/10T1/2 and BALB/c 3T3 cells have well known growth characteristics and are easily grown to large cell numbers for these experiments. However, attention must be paid to the passage number of immortalized cells used to ensure inter-experiment comparability, as phenotypic or genotypic changes may occur.
Terzaghi-Howe (1987) found that, unlike primary epithelial cells,
36
that established cell lines would be more sensitive to down-regulation by environmental substances being tested (ie, exhibit a lower threshold of GJIC regulating effects), thus providing a more conservative risk characterization. However, this possible advantage would seem to be outweighed by the fact that immortalization often produces more readily transformable cells which are not well characterized in their growth regulating properties. Strongly communicating, primary epithelial cells are more characteristic of normal in-vivo conditions surrounding initiated cells. Therefore, experiments using differentiated primary cells and initiated or preneoplastic cells from the same tissue will provide the most useful information about growth and
communication in in-vivo simulating conditions. In the absence of test chemicals, there should be sufficient heterologous communication present to suppress focal growth by initiated or preneoplastic cells.
3. Cellular metabolism and culture conditions. In evaluating a test
GJIC, it is necessary to include direct measurements of both homologous and heterologous GJIC (via dye or radio-label transfer or metabolic cooperation)
and correlated measurements, under the same culture conditions, of an
endpoint related to proliferation of the initiated cells (such as focal growth in co-cultures or growth of initiated cells on a normal "cell mat"). A complete characterization of altered cell growth in the presence of varying proportions of normal cells (such as the type of experiment proposed in section IB)
permits a choice of heterologous culture conditions for maximum
responsiveness to GJIC effectors. All measurements of GJIC should be made after cells reach confluence, when junctional communication is maximally established (Boreiko et aL 1986). Cell growth experiments will require longer culture periods of two or more weeks for development of effects. Tests
should also be made of reversibility of observed effects upon removal of test chemical from the culture medium, with time dependence information where possible to allow for temporal correlations.
5. Dose / Response. For any chosen endpoints in a given heterologous system, and for a given test compound, as complete dose/response data as possible should be collected, from concentrations about one order of
. 38 .
5. Test Substances. Each test compound should be dissolved in as small a volume as possible of DMSO, ethanol or other non-cytotoxic, water miscible solvent to be added to culture medium at no more than 0.5% by volume. In some cases, such as the exposure of lung epithelial cells to tobacco smoke, it may be desirable to provide the test compotmd or mixture in particulate form. At the end of the exposure period, the cell layer should be washed with fresh medium to remove as much residual compound as possible. All
compounds must also be tested for activity in one or more genotoxidty assays to distinguish promoting effects from growth regulatory changes caused by secondary genetic lesions.
6. Controls. Positive: Indude at least an abbreviated dose-response for a known promoter, such as TPA, in both communication and initiated cell growth assays.
Negative: Solvent controls should be included for the same exposure times as test compounds. Cytotoxidty/Plating Efficiency: The survival and colony formation rate in sparse plating trials of each cell type should be used to modifiy mixing ratios for heterologous experiments and determine upper limits for test chemical exposure.
through gap junctions.
Genotoxicity: Putative tumor promoting compounds to be evaluated for effects on GJIC and growth of initiated cells must have been shown to be non-genotoxic by one or more short term tests.
B. Development of Proposed In-Vitro Assay For Gap Jimctional Function Using the Endpoints of Focal Growth of Initiated Cells and Dye Transfer
Initial Design Rationale for Heterologous (Normal/Initiated) Communication Experiments
The work of Mehta, Bertram etal (1986), Yamasaki (1987,1988) in-vitro and
the in-vivo studies of Terzaghi-Howe (1987) and Dotto, Weinberg and Ariza (1988) have shown that normal, communicating cells can suppress the growth of preneoplastic fod (piled-up, morphologically altered cells,
descended from cells with an initial genetic lesion). Further, we know that
there is a threshold communication level below which there is no
suppression of focal growth (Mehta et al, 1986), and, if this suppression is a function only of GJIC, there will be a maximum growth suppression level corresponding to maximal heterologous communication.
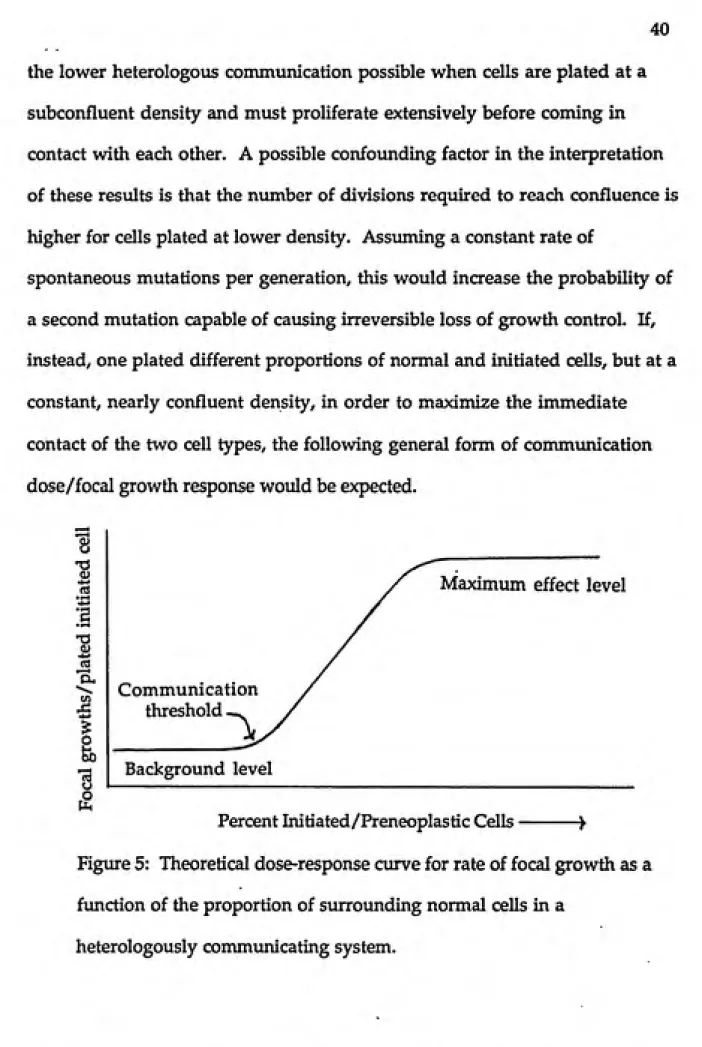
40
the lower heterologous communication possible when cells are plated at a subconfluent density and must proliferate extensively before coming in contact with each other. A possible confounding factor in the interpretation of these resvdts is that the number of divisions required to reach confluence is higher for cells plated at lower density. Assuming a constant rate of
spontaneous mutations per generation, this would increase the probability of a second mutation capable of causing irreversible loss of growth control. If, instead, one plated different proportions of normal and initiated cells, but at a constant, nearly confluent density, in order to maximize the immediate contact of the two cell types, the following general form of communication dose/focal growth response would be expected.
fS
•2
Si
«j
'a,
^.^
(A
Maximum effect level
Communication
threshold -^
Background level
Percent Initiated/Preneoplastic Cells ->
Figure 5: Theoretical dose-response curve for rate of focal growth as a function of the proportion of surrounding normal cells in a
For any given heterologous cell system, in any specified culture conditions, this cvirve would have a characteristic background rate of focal growth, slope and location of the transition region of the curve, and maximum focal
growth rate. These characteristics would depend on the type of genetic lesion(s) present in the initiated cells, the intrinsic communication of the two cell types, the distribution of contacts achieved by neighboring cells in the test culture and any exogenous factors which may up- or down-regulate GJIC. If a compound is added to the culture medium known to inhibit
communication—and there are some indications that some established cell
lines (Fransson-Steen and Warngard, 1992) and initiated cells (Trosko et al, 1990) are more sensitive to down-regulation than primary cells—the above curve would shift to the left, as there would be less suppression of focal growth at each proportion of co-plated cells. Similarly, the introduction of additional genetic lesions in the initiated cells (by spontaneous mutations or treatment with genotoxin) which altered either the communication
competence or the ability to respond to heterologous regulatory signals, would shift this curve to the left and perhaps change the background and maximum focal growth rates. Addition of cyclic-AMP or other compounds known to up-regulate GJIC in some cell systems, would be expected to shift the transition to the right. Such experiments, especially when coupled with independent measurements of GJIC, could establish a definitive role for
42
endpoint of focal growth, and provide the basis for a relatively simple short-term test for suspect tumor promoters.
Heterologous Cell Systems Chosen
1. C3H/10T1/2 and H-23 cells. The mouse fibroblast cell line C3H/10T1/2
(Reznikoff et al, 1973), shown in phase contrast and fluorescent images in Figure 6, and various initiated clones of this line have been extensively used for both GJIC and focal growth experiments (Mehta, Bertram and
Loewenstein, 1986; Boreiko, 1986; Hossain et al, 1989; Boreiko et al, 1989;
Bertram, 1990). These lines are considered relatively easy to culture and, because of their well-characterized behavior in these types of experiments (Boreiko, 1986), they were chosen for these preliminary studies. A further advantage of a clonally-derived initiated cell line, such as the H-23 cells (3-methylcholanthrene initiated) used here, is the uniformity of its genome. In the absence of suppression by communication with normal cells, changes in phenotypic expression of initiated cells are observable in the form of an increased rate of focal growth. Focal cells, while often referred to as
"transformed", are not irreversibly committed to tumor growth, however; when dispersed and cultured with non-initiated cells under commvmicating conditions, these cells will often grow normally, with altered phenotype suppressed.
HGURE 6. Mouse fibroblast C3H/10T1/2 cells, phase contrast and fluorescent
43
HME cells are primary cells isolated from normal human breast tissue
(Chang) and include both imdifferentiated putative stem cells (Type I), which
are not communication-competent, and differentiated Type n cells, which are
highly commimicating (Fig. 7), as well as a few fibroblastic cells. These cell
types have differing kinetics of attachment to tissue culture plastic
(fibroblasts>Typen>TypeI) and a fairly pure culture of Type n cells can be
obtained by selective plating.
HS578T tumor cells were first isolated from a human breast tumor and
characterized by Hackett et al (1977). They are highly communication
competent among themselves (Fig. 8), but, unlike most non-communicating
tumor cells and some homologously communicating tumor cells, such as
Wilm's Tumor (Trosko et al, 1990), HS578T cells are unable to cause tumors
when injected into immune-suppressed mice (Hackett et al, 1977), leading to
the hypothesis that there is some at least minimal level of heterologous
communication which allows the tumor phenotype and uncontrolled cell
growth to be suppressed. When cultured alone, they display many
characteristics of malignant cells, including altered morphology, imstable
chromosome number (aneuploidy) and high density, multilayered growth
(Hackett, 1977), perhaps indicating more than one accumulated genetic lesion,
but without irreversible commitment to tumor formation.
m
e
hp.
'A
m
u
*^%
- ^--—.^ifc-ͣ*i^:^«ͣ:. «ͣ>.&ͣͣ*.*»ͣ-..: .- .: .m^---^-l 't^^ftTf^ra.
m
$.
type was first grown up to the required cell numbers separately, in optimal media, then they were plated together in desired proportions and grown in a "compromise" medium which supported slower proliferation for both cell types (see experimental design for details). Scrape-loading experiments with preliminary heterologous cultures were at least suggestive of partial
communication between the cobbled appearing HME Type n cells and the larger, more spindle-shaped HS578T cells (Fig. 9).
Heterologous Culture Experiments
The purpose of these experiments was to define the characteristic focal growth response curve as a function of communication between normal and
initiated cells. At the same time and under the same culture conditions,
independent, semi-quantitative assessments of gap junctional
communication were made by observation of Lucifer Yellow transfer between normal and initiated cells (heterologous), as well as between cells of the same tj^e. If the primary mechanism controlling rate of focal growth is regulation of GJIC, culture in the presence of known tumor promoters would be
i^^^^r
^
m
s
>?^
i^
s
^
^
n
¥
FIGURE 9. Heterologous oolture of HME (cobbled appearance) and HS578T
Cell Culture Conditions
The above-described cell types were rapidly thawed from liquid nitrogen frozen stocks and grown imder 5% CO2 at 37° in the following media.
C3H/10T1/2 and C3H/10T1/2 (clone H-23) were grown in D medium (Gibco)
with 10% FBS (Gibco). Cells were subcultured at about 80% confluence imtil
desired cell numbers were acquired. Retinal acetate (1 [iM) was added to H-23 cultures to suppress spontaneous focal growth (Roimdhaug et aJ, 1987), until co-plating with normal cells. Human Mammary Epithelial cells were
suspended in 1:1 D medium/MCDB (Gibco) plus 5% FBS and plated on 90 mm culture dishes to allow attachment of fibroblasts. After two hours, supernatant was transferred to a T-75 flask overnight to attach Type n cells. Supernatant, containing most of the Type I cells, was removed and the medium changed to 1:1 D/MCDB with no FBS. These primary cells could only be subcultured two or three times, at moderate density, before terminal
differentiation and sloughing would occur. HS578T cells were suspended and
grown for several days in D mediim\ plus 5% FBS. Cells recovered andproliferated most rapidly when subcultured at about 80% confluence, at which time the medium was changed to 1:1 D/MCDB.
Heterologous Gap Junctional Communication Assays
,. 46
to improve visualization of communication between cell types. The evening before initiating the heterologous cultures, l|i.m diameter fluorescent beads
(Polysdences) were added at a density of about 108/cm2 to cultures of H-23 and
HS578T cells, which phagocytized several beads per cell. In the morning, these cultures were washed five times with phosphate buffered saline (PBS), trypsinized, and resuspended in medium containing 5% PBS. At least 1 X 105 cells of the appropriate initiated type were added to the plates containing small colonies of C3H/10T1/2 (D mediimi plus 10% FBS) or HME cells (1:1 D/MCDB). Plates were grown about three days to confluence before addition of promoters (TPA at 10 and 100 ng/ml or TCDD at 0.1 to 1 ng/ml) to media. Media were changed every three days and communication levels were assessed at 0, 24, 72 and 96 hours after addition of test compounds.
Scrape-Loading/Dye Transfer Assay (after El-Fouley jgt al, 1986) 1. Medium poured off and cell layer washed three times with PBS.
2. 2 ml of 0.5 mg/ml Lucifer Yellow solution in PBS added to cover cells. Cell
layer repeatedly scored at scattered locations with a scalpel blade, using a
rocking motion to avoid tearing. Cells are allowed to take up dye for three
minutes. I
3. The cell layer is washed five times with PBS . Cells are covered with
medium and observed immediately on a Nikon fluorescent microscope with
permit the researcher to readily differentiate initiated/preneoplastic cells
from normal ones.
Heterologous Culture Focal Growth Experiments
For these experiments, all cell types were grown, and H-23 and HS578T
cells were bead-labelled, as described above. However, in order to maximize
mixing of cells and thus contact between normal and initiated/preneoplastic
cells, both cell types of each heterologous pair were trypsinized, counted and
plated at the same time with extensive physical mixing. For each
heterologous experiment, ten 35 mm plates were set up with each of the
following proportions of initiated/preneoplastic cells: (A), 0%; (B), 20%; (C),
40%; (D), 60%; (E), 80%; (F), 100%. Approximately 1.25 X 105 total cells were
plated per dish. C3H/10T1/2 and H-23 cultures were grown in D medium
plus 10% FBS; HME/HS578T cultures in 1:1 D/MCDB. After reaching
confluence, duplicate plates at each relative concentration of cells were treated
with: 10 nl EtOH (control), 100 ng/ml TPA for 24 or 72 hovirs, or 1 ng/ml
TCDD for 24 or 72 hours. At 3.5 weeks, all dishes were washed with PBS,
stained with crystal violet to visualize focal growths, washed, air-dried and
48
n. Conceptual Links Between the Development of an In-Vitro Assay and
Modelling of Rat Liver Tumor Promotion by Phenobarbital
The work done in the preparation of this thesis is of two rather different
types: first, the in-vitro methods development of an assay to increase the
empirical data base for a major bioeffect of suspected tumor promotors and
secondly, the presentation and testing of a simple theory for tumor
promotion based on phenotypic change which is at least partly induced by
decreased gap junctional communication. It is therefore important to
integrate the conceptual bases for the assay development and the proposed
promotional theory as tested on in-vivo data.
Both the in-vitro data that would be generated by the proposed
heterologous cell assay and the literature data (mostly from in-vivo systems)
used to evaluate our simplified model involve the clonal growth of cells of
recognizably altered phenotype into foci observable by differential staining.
The purpose of both approaches, then, is to quantify the expression of
chemically-induced, reversibly altered phenotypes which lead to more rapid
cell proliferation and to directly measure gap junctional commimication
under the same exposure conditions.