Behavioural analysis of olfactory conditioning in the moth spodoptera littoralis (Boisd ) (Lepidoptera: noctuidae)
8
0
0
Full text
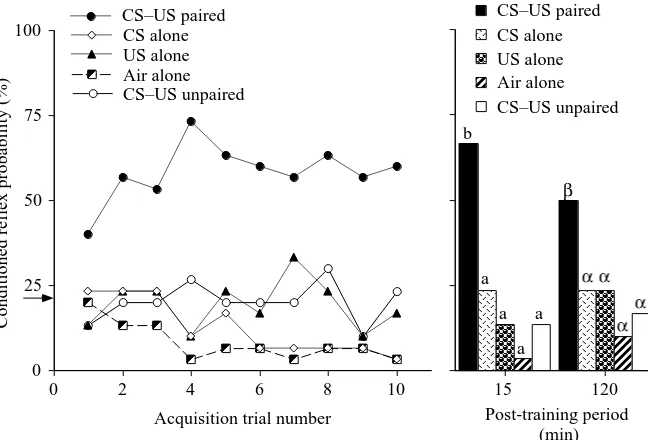
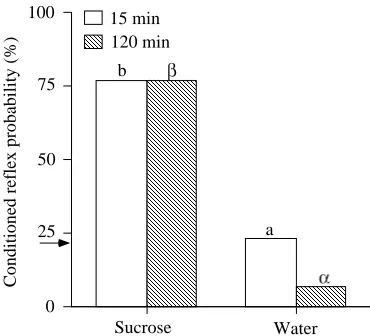
Figure

Related documents