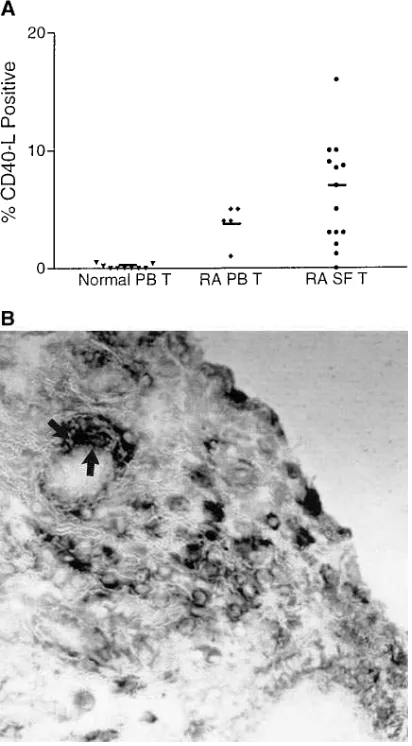
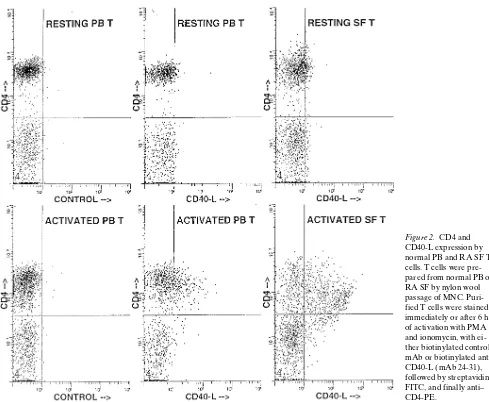
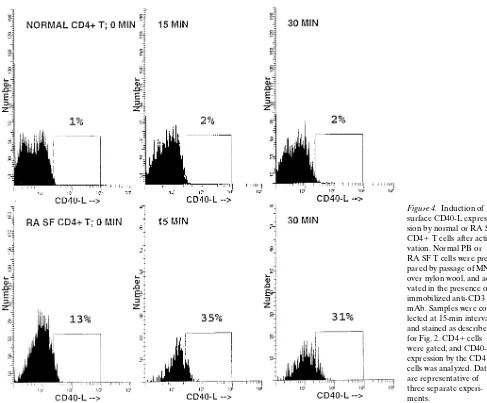
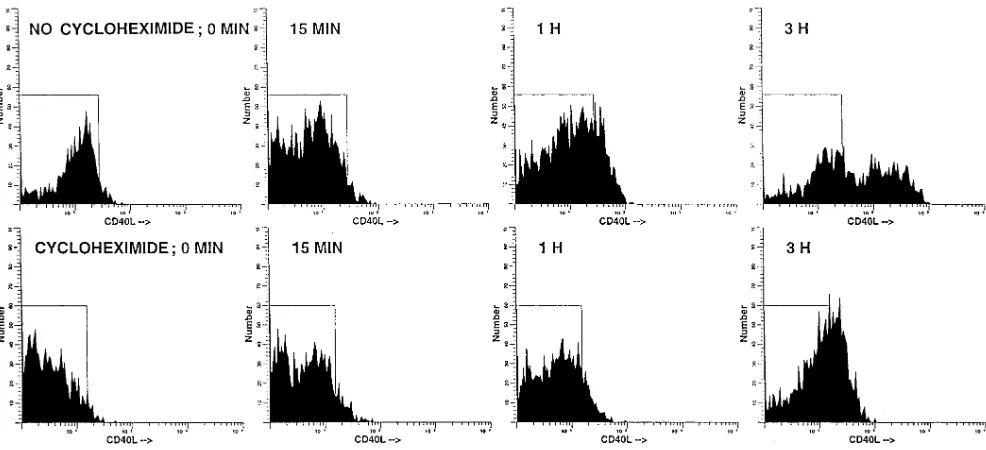
Functional CD40 ligand is expressed by T cells in rheumatoid arthritis
Full text
Figure
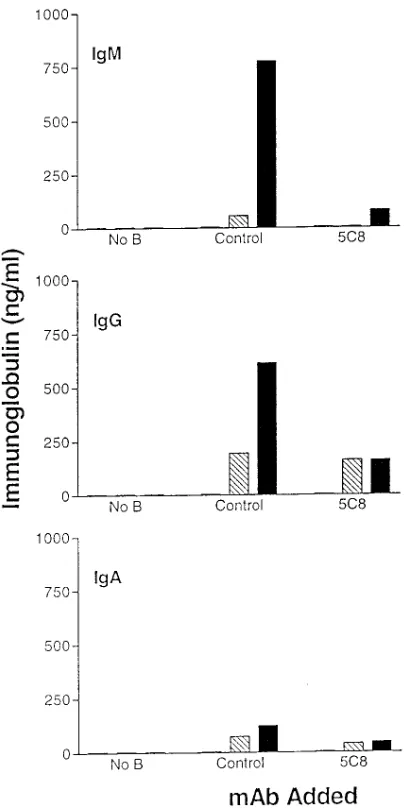
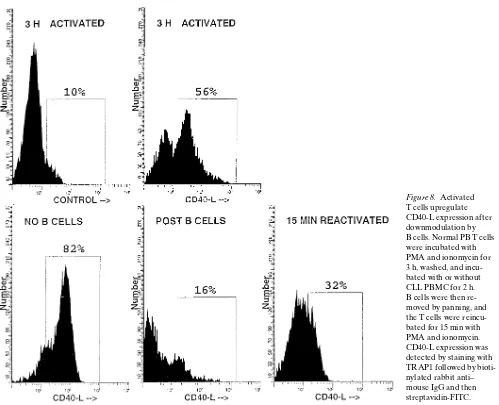
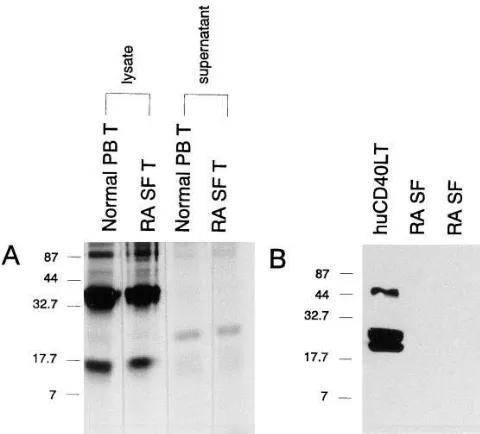
Related documents
Freshly isolated human primary NK cells induce preferential lysis of Oral Squamous Carcinoma Stem Cells (OSCSCs) when compared to differentiated Oral Squamous Carcinoma Cells
Abstract: Squalene has been isolated from rice bran oil fatty acid distillate (RBO FAD) by a combination of biohydrolysis, bioesterification and molecular
The different techniques of image segmentation include thresolding, clustering, region merging, region growing, color segmentation, motion segmentation and automatic
CONCLUSION: This comparative Scintigraphy study suggests that the topical application of quick penetrating solution of diclofenac results in increased penetration of
Based on my observations, four DOFs were identified to perform a needle-based intervention with a robotic manipulator: Two rotations (i.e., pitch and yaw) about the entry port into
(a) TEM imaging; (b) Size distribution; and (c) Day- light image of 3C-SiC QD suspension in aqueous solution.. and size distribution of the QDs are the most important factors on
3) To begin firing disengage the safety, make sure you are ready to fire and the shotgun is pointed in a safe direction to be fired. After you have fired the first round by pulling
In this example, it is possible to show that th~ mean square error can be decreased because the isotonic regression estimates are the nearest points in the restricted parameter space
