INTRODUCTION
When land was colonised, plant morphology significantly diversified (Kenrick and Crane, 1997b; Langdale, 2008). The underlying developmental mechanisms of this diversification during evolution are still poorly understood. Shoot branching is one of the key innovations that led to more complex architecture. Recently, strigolactones were shown to represent a novel class of plant hormones controlling this process (Gomez-Roldan et al., 2008; Umehara et al., 2008). The same molecules act in the rhizosphere as signalling molecules in establishing the symbiotic relationship between arbuscular mycorrhizal fungi and over 80% of extant land plants (Akiyama et al., 2005; Besserer et al., 2006) and in stimulating seed germination of root parasitic weeds (Bouwmeester et al., 2007). Arbuscular mycorrhizal symbiosis evolved for 460 million years before the appearance of roots and is coincident with the appearance of the first bryophyte-like land plants (Remy et al., 1994). In seed plants, strigolactone-deficient mutants in pea (Fig. 1A), Arabidopsis thalianaand rice show an increased branching phenotype (Gomez-Roldan et al., 2008; Umehara et al., 2008). To gain a better understanding of the origin of branching in seed plants and to explore strigolactone functions in early land plants, we investigated whether strigolactones are produced in the moss Physcomitrella patensand what roles they play in a non vascular plant.
Strigolactones are apocarotenoids (Matusova et al., 2005) and the first step in their biosynthesis is catalysed by CAROTENOID CLEAVAGE DIOXYGENASES (CCD). The CCDs are an ancient
family of enzymes with members identified in bacteria, plants and animals (Auldridge et al., 2006). In Arabidopsisand pea, the CCD7 and CCD8 enzymes are encoded by the branching genes MAX3/RMS5 (Booker et al., 2004; Johnson et al., 2006) and MAX4/RMS1 (Sorefan et al., 2003), respectively, and are both involved in strigolactone biosynthesis (Gomez-Roldan et al., 2008; Umehara et al., 2008).
The model moss Physcomitrella patensprovides unique genetic and developmental facilities to undertake evolutionary studies in plant biology (Cove et al., 2006; Lang et al., 2008; Schaefer, 2002). Its life cycle is dominated by the haploid gametophyte that displays a succession of simple and well described developmental transitions that leads from the germinating spore to the colony of differentiated leafy shoots carrying mature diploid sporophytes (Cove et al., 2006).
We generated a mutant in the moss homolog of the strigolactone biosynthesis gene CCD8. Here, we demonstrate that in P. patens, PpCCD8is involved in strigolactone biosynthesis and regulates the branching of filament and colony extension. Furthermore, our data demonstrate that in wild-type (WT) P. patens, secreted strigolactones are directly involved in the regulation of colony extension in response to internal cues or population density. This novel function of strigolactones in plants is reminiscent of quorum-sensing signalling processes that are well described in bacteria (Fuqua and Greenberg, 2002; Ng and Bassler, 2009), and we propose that moss uses strigolactones as quorum-sensing molecules to regulate colonial growth in distinct environmental conditions.
MATERIALS AND METHODS
P. patensgrowth conditions and treatments
The Gransden wild-type strain was used and conditions for cultures were as described (Ashton and Cove, 1977; Ashton et al., 1979; Trouiller et al., 2006). For phenotypic analysis, spores were grown on minimal PP-NO3 medium (Ashton et al., 1979). Sporogenesis was performed in Magenta vessels in which 15-day-old wild-type and mutant colonies were grown for 2 months, then irrigated with sterile water and transferred to growth
Development 138, 1531-1539 (2011) doi:10.1242/dev.058495 © 2011. Published by The Company of Biologists Ltd
1Institut Jean-Pierre Bourgin, UMR1318 INRA-AgroParisTech, Centre de
Versailles-Grignon, 78026 Versailles Cedex, France. 2Weed Science Center, Utsunomiya
University, Utsunomiya 321-8505, Japan. 3Institute of Biology, Laboratory of Cell and
Molecular Biology, University of Neuchatel, CH-2009 Neuchatel, Switzerland. *Author for correspondence ([email protected])
Accepted 28 January 2011 SUMMARY
Strigolactones are a novel class of plant hormones controlling shoot branching in seed plants. They also signal host root proximity during symbiotic and parasitic interactions. To gain a better understanding of the origin of strigolactone functions, we
characterised a moss mutant strongly affected in strigolactone biosynthesis following deletion of the CAROTENOID CLEAVAGE DIOXYGENASE 8 (CCD8) gene. Here, we show that wild-type Physcomitrella patens produces and releases strigolactones into the medium where they control branching of protonemal filaments and colony extension. We further show that Ppccd8mutant colonies fail to sense the proximity of neighbouring colonies, which in wild-type plants causes the arrest of colony extension. The mutant phenotype is rescued when grown in the proximity of wild-type colonies, by exogenous supply of synthetic strigolactones or by ectopic expression of seed plant CCD8. Thus, our data demonstrate for the first time that Bryophytes (P. patens) produce strigolactones that act as signalling factors controlling developmental and potentially ecophysiological processes. We propose that in P. patens, strigolactones are reminiscent of quorum-sensing molecules used by bacteria to communicate with one another.
KEY WORDS: Physcomitrella, Strigolactones, CCD8
Strigolactones regulate protonema branching and act as a
quorum sensing-like signal in the moss
Physcomitrella patens
Hélène Proust1, Beate Hoffmann1, Xiaonan Xie2, Kaori Yoneyama2, Didier G. Schaefer3, Koichi Yoneyama2,
Fabien Nogué1and Catherine Rameau1,*
D
E
V
E
LO
P
M
E
N
putative starvation effect, the experiment was carried out on large Petri dishes (12 cm) and glucose (0.5%) was added in another similar experiment (not shown). Cell measurements were performed with ImageJ software (http://rsb.info.nih.gov/ij/).
Phylogenetic analysis
Evolutionary relationships between CCD proteins were analysed using a selection of proteins from different databases aligned with the program CLUSTALX (Thompson et al., 1997). The evolutionary history was inferred using the Neighbour-Joining method (Saitou and Nei, 1987) and the optimal tree was generated. Phylogenetic analyses were conducted in MEGA4 (Tamura et al., 2007).
Primers
The primers used in this study are described in Table S1 in the supplementary material.
Molecular cloning Ppccd8-KOconstruct
A 2855 bp PCR amplified PpCCD8 genomic fragment (scaffold 14: 2616535-2619389) was cloned into vector pCRII Blunt TOPO (Invitrogen). To disrupt the PpCCD8gene, a 664 bp 3⬘targeting fragment (scaffold 14: 2616135-2616798) was first recovered by enzyme digestion with XmnI and BamHI, and inserted into BamHI- and SmaI-digested pBNRF, which carries a loxP-neoR-loxP selectable marker (Schaefer et al., 2010). The 925 bp 5⬘targeting fragment (scaffold 14: 2618287-2619211), obtained by digestion with AcyI and BseJI, was then inserted between the
ClaI/HpaI sites of vector pBNRF carrying the 3⬘targeting fragment to generate pPpccd8-KO (see Fig. S1 in the supplementary material).
OsACTpro-108-HygR
The CaMVter terminator fragment was excised from pDH51 (Pietrzak et al., 1986) using XbaI/HindIII (HindIII site was blunted) and ligated to pCOR 104 (McElroy et al., 1991) digested by XbaI/SacII (SacII was blunted) to generate pCOR104-CaMVter. A 1800 pb fragment carrying the rice ACTIN-1 gene promoter (GenBank: S44221) and the CaMVter was obtained by digestion of pCOR104-CaMVter vector with XhoI and PvuII enzymes. This fragment was cloned into SwaI and SalI sites of BS-108 vector which contains a genomic P. patensfragment corresponding to locus 108 (Schaefer and Zryd, 1997) giving the OsACTpro-108 vector. The 35S::hygroR cassette (Hajdukiewicz et al., 1994) was excised of pBKS-hygro vector with HindIII and EcoRI enzymes, and integrated in the
HindIII site of p-Act-108 vector to generate OsACTpro-108-HygR(see Fig. S2A in the supplementary material).
OsACTpro:RMS1construct
The pea RMS1 cDNA (GenBank: AY557341) obtained by PCR was subcloned in pTOPO vector and inserted in NcoI/SphI sites of OsACTpro-108-H (see Fig. S2A,B in the supplementary material).
PpCCD8pro:GUS construct
This construct was made in the OsACTpro-108-H vector. A 1616 bp amplified fragment of the PpCCD8promoter (scaffold 14: 2619186-2620801) was cloned behind the GUS:Noster (Fourgoux-Nicol et al., 1999) cassette of the pAF1 vector digested with SacII/HindIII. A fragment containing PpCCD8pro:GUS:Noster was excised using SacII and NcoI, blunted and used to replace the deleted rice actin promoter CaMVter cassette of the OsACTpro-108-H vector with KpnI sites (see Fig. S2E in the supplementary material).
with similar phenotype were obtained and one of them, Ppccd8, was used for subsequent analyses.
OsACTpro:RMS1 and PpCCD8pro:GUS were linearised with SacII before transformation, and transformants were selected on hygromycin (25 g/ml). To confirm the presence of constructs in stable transformants, PCR amplification with specific primers designed on sequences inside and outside the construct was performed (see Fig. S2B-F in the supplementary material).
Quantification of strigolactones released by wild type and
Ppccd8⌬mutant
Approximately 1 g of 7-day-old protonemata from the wild type or
Ppccd8⌬mutant was used to inoculate 150 ml of BCD medium in a 300 ml flask on a rotary shaker. Cultures were grown under standard light conditions for 30 days and the growth medium was replaced with fresh media weekly. Thirty-day-old tissue was transferred to fresh media and grown for an additional 3 days. Plant material was collected by filtration and weighed, while the liquid culture medium was collected and extracted three times with an equal volume of ethyl acetate. The combined ethyl acetate solutions were washed with 0.2 M K2HPO4for removal of acidic compounds, dried over anhydrous MgSO4, filtered and concentrated in vacuo. These crude extracts were stored in sealed glass vials at 4°C until use. Strigolactones released to liquid culture media were determined on LC-MS/MS as described previously (Yoneyama et al., 2008). For quantitative analysis using LC-MS/MS, orobanchol-d1, orobanchyl acetate-d1 and [13C2]strigol were used as internal standards. Levels of other strigolactones were determined as reported previously (Yoneyama et al., 2008) and their recovery rates were estimated based on that of orobanchyl acetate-d1. The experiments were conducted with three replicate samples.
GUS staining of Physcomitrella patens
Colonies developed from ground tissues were incubated for 12 hours at 37°C in 0.8 mM 5-bromo-4-chloro-3-indolyl--D-glucuronide, 2 mM potassium ferrocyanide, 2 mM potassium ferricyanide and 100 mM phosphate buffer (pH 7).
Gene expression analysis
Total RNA was extracted using the RNeasy plant mini kit (Qiagen. Ref 74904). cDNA was synthesised from 2 g of total RNA with RevertAid H Minus M-MuLV Reverse Transcriptase (Fermentas) and dT(18) primer. Quantitative PCR was performed in a Mastercycle ep realplex (Eppendorf) with Realmaster mix SYBR ROX 2.5⫻(5 PRIME). PpAPT, which encodes an ADENOSINE PHOSPHORIBOSYL TRANSFERASE, was used as the constitutive gene (Trouiller et al., 2006). Relative expression of PpCCDwas established with the formula 2–Ct(PpCCD)–Ct(PpAPT). Three biological replicates were carried out with two technical replicates for each sample.
RESULTS
Characterisation of P. patensstrigolactone biosynthesis genes
Unique CCD7-like and CCD8-like sequences were found in the genome of P. patensthat were designated PpCCD7(GenBank
Accession Number HM007803) and PpCCD8 (GenBank
Accession Number HM007802). The predicted moss proteins share 45% and 57% amino acid identity with CCD7 and CCD8 from Arabidopsis, respectively. Phylogenetic analyses demonstrated that they grouped with their respective AtCCD
D
E
V
E
LO
P
M
E
N
clades (see Fig. S3 in the supplementary material). Two other genes involved in strigolactone biosynthesis have been identified by genetic analyses: MAX1 in Arabidopsis, which encodes a member of the CYP450 family; CYP711A1 (Booker et al., 2005); and the rice D27gene, which encodes an iron-containing protein (Lin et al., 2009). No MAX1homologue was identified in the P. patensgenome, whereas several homologues for D27 were found, one (XP001767010) encoding a protein with 47% of identity with D27 from rice.
To characterise the function of the CCD8gene in P. patens, we generated a Ppccd8 mutant in which genomic sequences encoding part of exon V, exons VI and VII were deleted. RT-PCR analyses confirmed the absence of a full-length PpCCD8transcript in the mutant, showing that the Ppccd8strain is a null allele (see Fig. S1 in the supplementary material). ThePpccd8strain was fertile and we used its selfed progeny to analyse single spore-derived colonies in all the experiments described below. Each colony should, thus, be considered as an individual sibling and not as a clonal replicate.
Ppccd8strains display several developmental phenotypes
Following spore germination, P. patens colonies develop into photosynthetic primary chloronemal filaments that further differentiate into caulonemal filaments implicated in rapid radial
[image:3.612.58.521.61.386.2]extension of the colony (Fig. 1B). Most caulonemal subapical cells further form secondary filaments and some buds (Fig. 1B) that will develop into leafy gametophores. The developmental fate of these caulonemal side branch initials is rather constant in P. patens, with most of them developing into secondary chloronemata and only a few percentage into new caulonemata or buds (Cove et al., 2006). Under our growth conditions, spore germination occurred significantly earlier in Ppccd8spores than in the wild type (see Fig. S4A in the supplementary material) and was associated with a higher frequency of multipolar germination patterns (Fig. 1C,D). Branching of primary chloronemata was also higher in the mutant during this early developmental stage. The percentage of lateral cells was higher in the mutant (37.2%) than in the wild type (10.8%) 93 hours after spore transfer on the medium. This increase in lateral cell number is not due to early germination of the Ppccd8mutant as the percentage was still higher for the mutant at 81 hours (25.6%) (see Fig. S4B in the supplementary material). The first caulonemal cells differentiated ~7 days after spore germination, followed by the formation of the first buds at day 15; timing of chloronemata to caulonemata transition was similar in both wild type and Ppccd8 mutant with a higher number of caulonemata in the mutant very likely to be due to higher branching of chloronemal cells (see Fig. S4C in the supplementary material). Fifteen days after spore germination, branching (i.e. production of secondary filaments or new buds) of caulonemata protruding from Fig. 1. CCD8controls shoot branching in pea and the synthesis of a diffusible signal regulating caulonema branching and colony
extension in P. patens. (A)Wild-type (left) and rms1 (Psccd8) (right) pea plants. (B)Caulonemata (ca), secondary chloronemata (ch) and bud (bud) from a 18-day-old wild-type colony. (C,D)Development of wild-type (C) and Ppccd8⌬(D) colonies, 56 to 106 hours after spore transfer to the medium. (E-H)Wild-type (E,F) and Ppccd8⌬(G,H) colony, periphery of 25-day-old colonies (E,G), 40-day-old colonies (F,H). (I-L)Cross-feeding experiment with a 30-day-old central colony surrounded by four 40-day-old colonies. Scale bars: 200m in B; 100m in C,D; 500m in E,G; 1 cm in F,H-L.
D
E
V
E
LO
P
M
E
N
in the Ppccd8mutant and that the higher branching observed in the mutant was not linked with a general higher cell division rate. A sudden and significant difference appeared 20 days after spore germination: the diameter of wild-type colonies reached a plateau, while mutant colonies continued to extend (Fig. 1F,H; see Fig. S4G in the supplementary material). At this stage, all caulonemal-type filaments, a mixture of caulonemata and rhizoids, protruding from the wild-type colonies were unbranched (Fig. 1E) and had stopped to elongate. By contrast, elongation and branching of Ppccd8 caulonemata continued up to 45 days after spore germination giving rise to much larger colonies (Fig. 1G; see Fig. S4F,G in the supplementary material).
In both genotypes, the first buds appeared ~8 days after caulonema differentiation. Size of gametophores was affected in Ppccd8compared with wild type. Mutant gametophores were in average half the size of wild type, had reduced number of leaves and shorter internodes after 45 days (see Fig. S4H-J in the supplementary material). Yet leaves were normally shaped and displayed normal phyllotaxy, and the general architecture of the leafy shoot was unaffected. Most importantly, we did not observe any increase in gametophore branching, and all Ppccd8 gametophores were formed by a single stem as in wild type (data not shown). Gametophores bearing a capsule were less frequent in the mutant (0.55 sporophyte/gametophore in wild type versus 0.04 in Ppccd8). Nevertheless, capsules from mutant contained viable spores but in reduced numbers compared with wild type (see Fig. S4K,L in the supplementary material).
These results indicated that while cells and organs forming gametophyte and sporophyte were not altered, except in size, control of protonema branching and colony extension was strongly affected in Ppccd8.
A diffusible signal secreted by wild-type colony complements Ppccd8phenotype
In P. patens, hormones such as auxins and cytokinins are secreted in the medium to control the protonemal stage of gametophyte development (Cove et al., 2006). To assess whether the PpCCD8 gene is involved in the biosynthesis of a signal secreted into the medium, we performed a series of cross-feeding assays.
After 2 weeks of co-culture, the Ppccd8colony surrounded by wild-type colonies displayed a wild-type phenotype with a strong decrease in colony diameter (Fig. 1I-L; see Fig. S5 in the supplementary material). These results suggested that wild-type moss produces a signal that diffuses into the medium and complements the phenotype of Ppccd8. This signal inhibits colony extension and filament branching. The mutant does not appear to be producing this inhibitor that accounts for its developmental phenotype.
When a wild-type colony was surrounded by Ppccd8colonies, a slight but significant increase in the diameter of the central wild-type colony was observed (Student’s t-test, P<0.01) (see Fig. S5 in the supplementary material). The central wild-type colony also had
upregulated in strigolactone-deficient mutants suggesting that negative feedback controls strigolactone biosynthesis (Beveridge and Kyozuka, 2010). In rice, application of the synthetic strigolactone GR24 to the strigolactone deficient ccd8(d10) mutant restored CCD8 transcript levels to wild type (Umehara et al., 2008).
Expression of PpCCD8in the wild type was first investigated by quantitative RT-PCR. The expression level of PpCCD8was low but detectable 10 days after spore germination and increased approximately eightfold between days 15 and 21 to reach a plateau (see Fig. S6A in the supplementary material). These results show that PpCCD8is expressed at low levels in the protonemata and is upregulated along with the differentiation of new gametophores where it is more abundantly expressed (see below).
Quantitative RT-PCR further revealed that PpCCD7displayed a similar expression profile in both wild-type and Ppccd8colonies. Yet PpCCD7transcript levels were five to 20 times higher in Ppccd8than in wild type (see Fig. S6A,B in the supplementary material). This observation suggests that, in Physcomitrella, the two CCD genes are co-regulated with feedback control of PpCCD7, as previously observed in seed plants (Beveridge and Kyozuka, 2010).
To gain more insight in the regulation of PpCCD8expression, the -glucuronidase (GUS) reporter gene driven by PpCCD8 promoter sequence was introduced into locus 108 in wild-type and Ppccd8strains. Locus 108 is often used as target sequence as it does not contain any coding sequence (Schaefer and Zryd, 1997). GUS expression driven by the PpCCD8 promoter was not detectable in wild type by histochemical staining (data not shown), indicating that the expression level of PpCCD8was very low. By contrast, GUS staining was visible following overnight incubation in the Ppccd8 background, and was observed in every differentiated bud in the protonemata and at the foot of each gametophore (Fig. 2A,B). These data indicate that PpCCD8 expression is also regulated by feedback control, as observed above for the PpCCD7transcript. They further indicate that the biological activity of PpCCD8is developmentally regulated and spatially concentrated at the base of gametophores.
Strigolactone identification and production in wild type andPpccd8
The presence of strigolactones in Physcomitrellawas tested further with the analysis of liquid medium in which differentiated wild-type and Ppccd8tissue were grown for 3 days. As shown for several seed plants, P. patensexuded a mixture of strigolactones (see Fig. S7A,B in the supplementary material). The major strigolactones produced by the wild type were 7-oxoorobanchyl acetate, orobanchyl acetate, 7-hydroxyorobanchyl acetate, fabacyl acetate, strigol and orobanchol (Fig. 3A). The mutant produced only strigol and 7-oxoorobanchyl acetate (Fig. 3A) along with small amounts of 5-deoxystrigol, which could not be quantified in either strains. Wild type produced and released approximately 350
D
E
V
E
LO
P
M
E
N
pg/g fresh weight (FW) of strigolactones in 3 days, while the mutant produced 40 pg/g FW, less than one-eighth that of wild type. These data provide the first characterisation of strigolactones in a moss. They further demonstrate that strigolactone biosynthesis is severely affected in Ppccd8, indicating the involvement of PpCCD8in strigolactone biosynthesis in P. patensand the potential involvement of the strigolactone pool in the developmental phenotype described above.
Ppccd8phenotype is complemented by expression of pea CCD8and by exogenous strigolactone application
To examine whether expression of CCD8genes from seed plants can complement the Ppccd8 phenotype, the pea CCD8gene (RMS1) driven by the rice Actin-1 gene promoter was introduced at locus 108 in the Ppccd8 mutant. Several independent Ppccd8::OsACTpro:RMS1strains were obtained that exhibited the same wild-type developmental pattern: colony extension stopped after 3 weeks and colonies developed leafy shoots of normal size. Furthermore, these strains perfectly restored normal development of Ppccd8colonies in cross-feeding experiments (see Fig. S8A,B in the supplementary material). Consistently, semi-quantitative RT-PCR indicated that the expression of PpCCD7 was reduced to wild-type levels in Ppccd8::OsACTpro:RMS1strains (see Fig. S8C in the supplementary material). These results provide genetic evidence that pea RMS1 is able to restore both normal development and feedback control of PpCCD7 expression in Ppccd8mutant. These data suggest that pea and moss CCD8 have the same biochemical activity that has been conserved during land plant evolution.
To determine whether the synthetic strigolactone GR24 could complement the Ppccd8developmental phenotype, 15-day-old Ppccd8colonies were transferred on media containing 1 M of GR24. After 2 weeks, these colonies showed an intermediate diameter and were surrounded by unbranched caulonema-type filaments (Fig. 3B-G). In a dose-response experiment, the diameters of both wild-type and Ppccd8 colonies were significantly smaller after 23 days on medium supplemented with 100 nM or 1 M GR24 (test of ANOVA, P<0.01, n12; see Fig. S9 in the supplementary material). At the same stage, we observed in both strains that gametophores were larger than in the non-supplemented controls (Fig. 3B-D). To assess whether GR24 was able to restore feedback control on PpCCD7 expression, we analysed its expression in 21-day-old wild-type and Ppccd8 colonies during the 2 days following transfer on media containing
[image:5.612.322.555.56.456.2]1 M GR24. These analyses showed that GR24 downregulated PpCCD7expression in Ppccd8(Fig. 3H). To determine whether the wild-type diffusible signal identified in cross-feeding experiments also regulates PpCCD7 expression, we analysed Ppccd8samples that were transferred on media where wild-type or Ppccd8 colonies had grown previously. The same down regulation of PpCCD7expression was observed in Ppccd8tissue treated with wild-type preconditioned medium (Fig. 3H). Altogether, these data show that GR24 almost completely complements the developmental phenotype of Ppccd8mutant, the Fig. 2. PpCCD8is expressed at the base of each gametophore.
(A)PpCCD8pro:GUS in a 21-day-old Ppccd8⌬colony. (B)Young gametophore (enlargement of A as indicated). Scale bars: 500m in A; 100m in B. Arrowheads indicate bud or gametophore.
Fig. 3. The signal that regulates colony extension in P. patensis a strigolactone. (A)Quantification of strigolactones released by wild type (open bars) and Ppccd8⌬(black bars) in the growth media for 3 days. Data are presented as mean±s.e.m. (n3).
(B-G)Complementation of Ppccd8⌬by 1M GR24 application. Twenty-five-day-old colonies (B-D). Periphery of the same colonies (E-G). ch, chloronema side branches. Scale bars: 1 cm in B-D; 1 mm in E-G. (H)Expression of PpCCD7in wild-type (white) and Ppccd8⌬colonies (21-day-old) 24 and 48 hours after transfer to new media containing 0M (black), 1M GR24 (blue) or to media on which wild-type (light grey) or Ppccd8⌬colonies (dark grey) were grown for 21 days. Error bars indicate s.e.m., n3. Letters indicate means that differ significantly
(P<0.01, ANOVA test).
D
E
V
E
LO
P
M
E
N
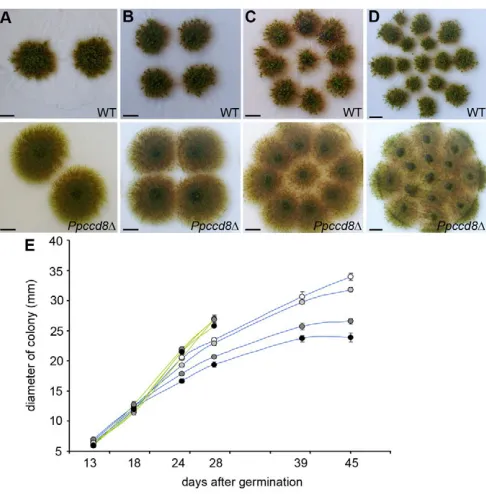
[image:5.612.57.289.62.166.2]mutant colonies. We followed the evolution of wild-type and Ppccd8colonies inoculated at four different densities (Fig. 4A-D). At high densities, the Ppccd8colonies started to merge at 28 days and at 39 days, the surface of the Petri dish was completely covered by mutant colonies (Fig. 4C,D). Consequently, after 28 days we were unable to measure colony diameter for Ppccd8. By contrast, at 39 days, regardless of the initial colony density, wild-type colonies showed a well-delimited round surface (Fig. 4A-D). ANOVA confirmed a significant effect of density on diameter at 28 days in wild type (P<0.001) but not in Ppccd8. The diameter of wild-type colonies was significantly higher at density 2 and 4 colonies per plate, and reached a plateau at 39 days for the two highest densities (Fig. 4E). These data show that in P. patens, strigolactones accumulate in the medium as the number of gametophores increases to a threshold concentration to control colony extension in response to colony density.
DISCUSSION
Conservation of strigolactone biosynthesis among land plants
The first steps in strigolactone biosynthesis involve the cleavage of carotenoids by CCD7 and CCD8 (Beveridge and Kyozuka, 2010; Leyser, 2008). The moss genome contains one orthologue for each corresponding gene and our phylogenetic analysis suggests that the CCD7 and CCD8 clades evolved before the separation of the bryophytes and the vascular plants from a common ancestor 420 million years ago (Kenrick and Crane, 1997b). Mutations in either of these two genes are associated with strigolactone deficiency in pea and rice (Gomez-Roldan et al., 2008; Umehara et al., 2008). Our analysis of strigolactone production in wild type and Ppccd8demonstrates that PpCCD8 is similarly involved in strigolactone biosynthesis in moss. In seed plants, the cytochrome P450 MAX1 is necessary for the synthesis of the shoot branching inhibitor, and grafting experiments indicated that it acts downstream of the two CCDs (Booker et al., 2005). Although there is no MAX1homologue in the moss genome,P. patens produces strigolactones that were previously identified in the exudates of seed plants. This suggests that MAX1 functions in later steps of the active branching inhibiting hormone biosynthesis, i.e. conversion of strigolactones to the shoot branching inhibitor. Hence, the bioactive signal in P. patens and seed plants may be different. Alternatively, moss may have other P450(s) able to replace the function of MAX1 for strigolactone biosynthesis.
The Ppccd8mutant still produced strigolactones, whereas no strigolactones were detected by LC-MS in the ccd8mutants of pea and rice (Gomez-Roldan et al., 2008; Umehara et al., 2008), suggesting that another strigolactone biosynthesis pathway may be present in moss. Hence, seven CCDgenes other than PpCCD8are present in the genome of Physcomitrella (see Fig. S3 in the supplementary material) that may be able to cleave carotenoids to produce small amounts of strigolactones. Alternatively, as root
exudates from the ccd8 mutants of pea and rice did induce germination of root parasitic weed seeds, it could suggest that these mutants may also still produce small amounts of strigolactones (Gomez-Roldan et al., 2008; Umehara et al., 2008).
Strong upregulation of CCD7 and CCD8 transcription is generally observed in strigolactone-deficient mutants (Dun et al., 2009). Our work revealed that transcription levels of both PpCCD7 and PpCCD8genes were also strongly upregulated in Ppccd8, indicating that the transcriptional feedback regulation that controls this biosynthetic pathway is probably conserved among land plants. We demonstrated that expression of pea CCD8fully complements the developmental phenotype of Ppccd8, which provides evidence for functional complementation of CCD8 proteins in these distant species. These results showed that the first steps of strigolactone biosynthesis are conserved among Embryophytes. However, strigolactone biosynthetic pathway is not fully characterised, and even in seed plants the biochemical function of MAX1, for example, is still unknown. A better understanding of the final steps of the strigolactone biosynthesis pathway should highlight difference and similarity between land plants.
Developmental and colonial defects of strigolactone deficient P. patens
[image:6.612.317.560.59.306.2]Detailed analysis of the strigolactone-deficient mutant showed that PpCCD8controls moss development from the germination stage with an earlier spore germination, higher number of primary chloronemata developing from the spore and higher branching of chloronemata in Ppccd8. This indicates that the strigolactone pathway is already involved in the spore development to control specific cellular process. The apparent different phenotypes may Fig. 4. Strigolactones control colony extension in response to colony density. (A-D)Thirty-nine-day-old wild-type (top panels) and
Ppccd8⌬(below panels) colonies grown at different densities. Scale bars: 1 cm. (E)Diameter of wild-type (blue lines) and Ppccd8⌬(green lines) colonies grown in light at different densities, two colonies (white), four colonies (light grey), nine colonies (dark grey) and 15 colonies (black) (n12). Data are mean±s.e.m.
D
E
V
E
LO
P
M
E
N
result from the same cellular mechanism of higher rates of lateral cell differentiation and/or division and would need further investigation.
The subsequent development of colonies was similar in both wild type and Ppccd8until 3 weeks after germination, indicating that strigolactones are not required to establish the normal developmental pattern of colonies. This is consistent with the absence of detectable morphological alteration in the mutant.
Under our growth conditions, and for a density of 15 to 20 colonies per Petri dish, wild-type colonies stop to extent and to differentiate a branched protonema after 3 weeks. By contrast, Ppccd8 colonies maintain their extension with a continuous production of secondary chloronemata and buds on caulonemata. The decrease in gametophore size, in number of gametophore bearing a capsule and in spore number per capsule observed in strigolactone-deficient Ppccd8 should be seen as a trophic response to higher intra-colony competition for resources between gametophores. Similarly, in seed plants, the high branching of strigolactone-deficient ccd8mutants results in an increase in the number of flowers per plant with a concomitant decrease in flower size and seed number per plant (Snowden et al., 2005).
Strigolactone activity in moss are reminiscent of quorum-sensing signal in bacteria
In wild type, the arrest of colony extension is sudden and involves a coordinated response from the whole colony. This response is developmentally associated with a complete arrest of caulonema branching and elongation. By contrast, despite producing a small amount of strigolactones, Ppccd8colonies continue to extend. All these results suggest that wild-type colonies released strigolactones or derived molecules into the medium to control colony extension. These data also indicate that a minimal threshold must be reached for the colony to respond to strigolactones and imply that strigolactones are synthesised and degraded in media as this threshold is never reached by Ppccd8colonies. Strigolactones are characterised by a tricyclic lactone (ABC part) connected, via an enol ether bridge, to a butyrolactone group (the D-ring) (Yoneyama et al., 2009). They are known to be short-lived in the rhizosphere owing to this labile ether bond that spontaneously hydrolyses in water (Akiyama and Hayashi, 2006; Akiyama et al., 2010). This instability creates a steep concentration gradient that has been suggested to be a reliable indicator of the proximity of a host root (Parniske, 2008).
Our analyses further demonstrate that strigolactones act as diffusible signals that regulate colonial extension in response to population densities (Fig. 4A-D). In wild type, the final diameter of 45-day-old colonies is negatively correlated with the number of colonies per plate and this correlation completely disappears for Ppccd8colonies (Fig. 4E). Based on these features, as well as on the fact that strigolactones are low molecular weight molecules that diffuse in the medium, we propose that strigolactones behave as quorum sensing-like molecules. Quorum sensing is a common, if not universal, cell-to-cell communication process in the microbial world. It is mediated by low molecular weight molecules, the most common signals in bacteria being acyl-homoserine-lactones (Ng and Bassler, 2009), that accumulate in the environment depending on the density of the organisms synthesising these molecules (Fuqua and Greenberg, 2002). When a quorum-sensing signal molecule reaches a threshold concentration, i.e. a threshold cell density, the entire population responds, usually through the coordinated expression of specific target genes. By analogy, we
propose that when a ‘quorum’ of strigolactone producing gametophores is reached, the whole colony and the neighbouring colonies respond by halting their extension.
The idea of chemical interactions between mosses regulating spreading of colonies is a long-standing observation for people working with mosses. A diffusible signal called ‘Factor H’, first identified in Funaria hygrometrica, was postulated to account for the maintenance of a clear zone between moss colonies or clones that did not involve competition for nutrients (Bopp, 1959; Klein, 1967). Factor H, which may be a complex of substances, was known to inhibit caulonema growth and lateral spread and to promote bud formation (Bopp and Knoop, 1974; Klein, 1967). Watson (Watson, 1981) proposed that such diffusible substances have developmental as well as ecological implications and may be possible determinant of moss community structure in nature. Strigolactone could be good candidates for the caulonema growth and lateral spread inhibition effect of the factor H. However, strigolactones have no inductive effect on bud formation and rather favour the simultaneous growth of gametophores already differentiated in the population. Thus, more extensive work is needed to asses whether strigolactones could be one component of the factor H.
Possible role of strigolactone in early land plant evolution
The colonisation of land by plants is associated with a large number of developmental, morphological, anatomical and chemical innovations (Kenrick and Crane, 1997b; Langdale, 2008). Arbuscular mycorrhizal symbiosis is an association between one of the oldest group of fungi, the Glomales, and plants. The fungal partner provides nutrients, essentially phosphate, and water to the plant that brings carbohydrates to the fungi (Parniske, 2008). This symbiosis has been found in almost all major lineages of extant land plants and is considered as one of the major innovations that contributed to the colonisation of land by plants (Kenrick and Crane, 1997b; Renzaglia et al., 2000). Mycorrhization is tightly associated with strigolactone biosynthesis (Akiyama et al., 2005; Akiyama et al., 2010; Gomez-Roldan et al., 2008). Despite the fact that P. patens, like most mosses, is not mycorrhizal, other bryophytes, liverworts that are considered the earliest-diverging lineage of land plants, and hornworts are mycorrhizal (Read et al., 2000; Wang and Qiu, 2006; Wang et al., 2010). The evolutionary homology of symbiotic interactions between gametophytes of liverworts and hornworts and mycorrhizal fungi, and those between sporophytes of vascular plants and the fungi has been demonstrated recently (Wang et al., 2010). These data and our demonstration that strigolactones are signalling molecules in the bryophyte P. patens support the hypothesis that strigolactones are very ancient molecules that have played a crucial role in the ecological adaptation of plants to terrestrial environment. Further studies with Marchantia, a mycorrhizal liverwort recently used in biological studies (Bowman et al., 2007; Chiyoda et al., 2008; Ishizaki et al., 2008,) would confirm the role of strigolactones in the establishment of arbuscular mycorrizal symbiosis in bryophytes.
Land plants (embryophytes) evolved from streptophyte green algae, a small group of freshwater algae ranging from unicellular flagellates (Mesostigma) to complex filamentous thalli with branching (Charales) (Becker and Marin, 2009). No CCD7 or CCD8 homologues have been found in streptophyte green algae EST databases, which could mean that strigolactones arose very early in the land plant lineage. However, no complete genome is currently available for streptophyte green algae (Becker and Marin,
D
E
V
E
LO
P
M
E
N
state of this particular organ, the axillary bud. It is quite intriguing that strigolactones control branching of the gametophyte in moss and branching of the sporophyte in seed plants. P. patensgenome analysis and its comparison with the genome of Arabidopsishave suggested that genes controlling gametophyte development have been recruited for sporophyte development (Nishiyama et al., 2003; Rensing et al., 2008). Whether common molecular actors are involved to control gametophyte (protonema) or sporophyte branching in response to strigolactones will be revealed by future studies. However the term ‘branching’ can be misleading and, for example, it is very likely that sporophyte branching due to dichotomy observed in many early taxa, involved different mechanisms from what is currently known in seed plants (Kenrick and Crane, 1997a; Langdale, 2008). Thus, more data will be needed in different taxa in order to decipher branching mechanisms in plants (Boyce, 2010). Nevertheless, this work is a first step in the understanding of the evolution of branching patterns and contributes to the wonderful story of how a small family of molecules has been co-opted during evolution to control chemical interactions between organisms and whole-plant architecture.
Acknowledgements
We thank F. D. Boyer for provided GR24, K. Mori for strigol, T. Asami for [2,3-13C2]strigol, and K. Akiyama for [6’-2H]orobanchol and [6’-2H]orobanchyl acetate. H.P. was supported by a PhD fellowship from the Ministère de l’Education Nationale, de la Recherche et de la Technologie (MENRT). Work from the C.R. group was supported by ANR (Agence Nationale de la Recherche, project ANR.05.BLAN.0262), and work from the K.Y. group was supported by KAKANHI (18208010, 2109111) and Program for Promotion of Basic and Applied Researches for Innovations in Bio-oriented Industry. We thank Y. Dessaux for helpful discussions; H. Höfte, Y. Dessaux, G. Pelletier and S. Bonhomme for their comments on the manuscript; and M. Sawabe for her assistance.
Competing interests statement
The authors declare no competing financial interests.
Supplementary material
Supplementary material for this article is available at
http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.058495/-/DC1
References
Akiyama, K. and Hayashi, H.(2006). Strigolactones: chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann. Bot. (Lond.)97, 925-931.
Akiyama, K., Matsuzaki, K. and Hayashi, H.(2005). Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature435, 824-827.
Akiyama, K., Ogasawara, S., Ito, S. and Hayashi, H.(2010). Structural requirements of strigolactones for hyphal branching in AM fungi. Plant Cell Physiol. 51, 1104-1117.
Ashton, N. W. and Cove, D. J.(1977). The isolation and preliminary characterisation of auxotrophic and analogue resistant mutants of the moss,
Physcomitrella patens. Mol. Gen. Genet.154, 87-95.
Ashton, N. W., Grimsley, N. H. and Cove, D. J.(1979). Analysis of gametophyte development in the moss, Physcomitrella patens, using auxin and cytokinin resistant mutants. Planta144, 427-435.
Auldridge, M. E., McCarty, D. R. and Klee, H. J.(2006). Plant carotenoid cleavage oxygenases and their apocarotenoid products.Curr. Opin. Plant Biol. 9, 315-321.
Becker, B. and Marin, B.(2009). Streptophyte algae and the origin of embryophytes. Ann. Bot.103, 999-1004.
produce a carotenoid-derived branch-inhibiting hormone. Dev. Cell8, 443-449.
Bopp, M.(1959). Versuche zur Analyse von Wachstum and Differenzierung des Laubmoosprotonemas. Planta53, 178.
Bopp, M. and Knoop, B.(1974). Régulation de la différenciation chez le protonéma des Mousses. Bull. Soc. Bot. France121, 145.
Bouwmeester, H. J., Roux, C., Lopez-Raez, J. A. and Becard, G.(2007). Rhizosphere communication of plants, parasitic plants and AM fungi. Trends Plant Sci.12, 224-230.
Bowman, J. L., Floyd, S. K. and Sakakibara, K.(2007). Green genes-comparative genomics of the green branch of life. Cell129, 229-234.
Boyce, C. K.(2010). The evolution of plant development in a paleontological context.Curr. Opin. Plant Biol. 13, 102-107.
Chiyoda, S., Ishizaki, K., Kataoka, H., Yamato, K. T. and Kohchi, T.(2008). Direct transformation of the liverwort Marchantia polymorpha L. by particle bombardment using immature thalli developing from spores. Plant Cell Rep.27, 1467-1473.
Cove, D., Bezanilla, M., Harries, P. and Quatrano, R.(2006). Mosses as model systems for the study of metabolism and development. Annu. Rev. Plant Biol. 57, 497-520.
Dun, E. A., Brewer, P. B. and Beveridge, C. A.(2009). Strigolactones: discovery of the elusive shoot branching hormone. Trends Plant Sci.14, 364-372.
Fourgoux-Nicol, A., Drouaud, J., Haouazine, N., Pelletier, G. and Guerche, P.
(1999). Isolation of rapeseed genes expressed early and specifically during development of the male gametophyte. Plant Mol. Biol. 40, 857-872.
Fuqua, C. and Greenberg, E. P.(2002). Listening in on bacteria: acyl-homoserine lactone signalling.Nat. Rev. Mol. Cell Biol. 3, 685-695.
Gomez-Roldan, V., Fermas, S., Brewer, P. B., Puech-Pages, V., Dun, E. A., Pillot, J. P., Letisse, F., Matusova, R., Danoun, S., Portais, J. C. et al.(2008). Strigolactone inhibition of shoot branching. Nature455, 189-194.
Hajdukiewicz, P., Svab, Z. and Maliga, P.(1994). The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Mol. Biol. 25, 989-994.
Ishizaki, K., Chiyoda, S., Yamato, K. T. and Kohchi, T.(2008). Agrobacterium-mediated transformation of the haploid liverwort Marchantia polymorpha L., an emerging model for plant biology. Plant Cell Physiol.49, 1084-1091.
Johnson, X., Brcich, T., Dun, E. A., Goussot, M., Haurogne, K., Beveridge, C. A. and Rameau, C.(2006). Branching genes are conserved across species. Genes controlling a novel signal in pea are coregulated by other long-distance signals. Plant Physiol.142, 1014-1026.
Kenrick, P. and Crane, P. R.(1997a). The Origin and Early Diversification of land Plants. Washington: Smithsonian Institution Press.
Kenrick, P. and Crane, P. R.(1997b). The origin and early evolution of plants on land. Nature389, 33-39.
Klein, B.(1967). Versuche sur Analyse der Protonemaentwicklung der Laubmoose. IV. Mitteilung. Der endogene Faktor H und seine Rolle bei der Morphogenese von Funaria hygrometrica. Planta73, 12.
Lang, D., Zimmer, A. D., Rensing, S. A. and Reski, R.(2008). Exploring plant biodiversity: the Physcomitrella genome and beyond. Trends Plant Sci.13, 542-549.
Langdale, J. A.(2008). Evolution of developmental mechanisms in plants. Curr. Opin. Genet. Dev. 18, 368-373.
Leyser, O.(2008). Strigolactones and shoot branching: a new trick for a young dog. Dev. Cell15, 337-338.
Lin, H., Wang, R., Qian, Q., Yan, M., Meng, X., Fu, Z., Yan, C., Jiang, B., Su, Z., Li, J. et al.(2009). DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth. Plant Cell21, 1512-1525.
Matusova, R., Rani, K., Verstappen, F. W., Franssen, M. C., Beale, M. H. and Bouwmeester, H. J.(2005). The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche spp. are derived from the carotenoid pathway. Plant Physiol.139, 920-934.
McElroy, D., Blowers, A. D., Jenes, B. and Wu, R.(1991). Construction of expression vectors based on the rice actin 1 (Act1) 5⬘region for use in monocot transformation. Mol. Gen. Genet.231, 150-160.
Ng, W. L. and Bassler, B. L.(2009). Bacterial quorum-sensing network architectures. Annu. Rev. Genet.43, 197-222.
Nishiyama, T., Fujita, T., Shin, I. T., Seki, M., Nishide, H., Uchiyama, I.,
Kamiya, A., Carninci, P., Hayashizaki, Y., Shinozaki, K. et al.(2003).
D
E
V
E
LO
P
M
E
N
Comparative genomics of Physcomitrella patens gametophytic transcriptome and Arabidopsis thaliana: implication for land plant evolution. Proc. Natl. Acad. Sci. USA100, 8007-8012.
Parniske, M.(2008). Arbuscular mycorrhiza: the mother of plant root endosymbioses.Nat. Rev. Microbiol.6, 763-775.
Pietrzak, M., Shillito, R. D., Hohn, T. and Potrykus, I.(1986). Expression in plants of two bacterial antibiotic resistance genes after protoplast transformation with a new plant expression vector. Nucleic Acids Res. 14, 5857-5868.
Read, D. J., Ducket, J. G., Francis, R., Ligron, R. and Russell, A.(2000). Symbiotic fungal associations in ‘lower’ land plants. Philos. Trans. R. Soc. Lond. B Biol. Sci.355, 815-831.
Remy, W., Taylor, T. N., Hass, H. and Kerp, H.(1994). Four hundred-million-year-old vesicular arbuscular mycorrhizae. Proc. Natl. Acad. Sci. USA91, 11841-11843.
Rensing, S. A., Lang, D., Zimmer, A. D., Terry, A., Salamov, A., Shapiro, H., Nishiyama, T., Perroud, P. F., Lindquist, E. A., Kamisugi, Y. et al.(2008). The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science319, 64-69.
Renzaglia, K. S., Duff, R. J., Nickrent, D. L. and Garbary, D. J.(2000). Vegetative and reproductive innovations of early land plants: implications for a unified phylogeny. Philos. Trans. R. Soc. Lond. B Biol. Sci.355, 769-793.
Saitou, N. and Nei, M.(1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol.4, 406-425.
Schaefer, D. G.(2002). A new moss genetics: targeted mutagenesis in Physcomitrella patens. Annu. Rev. Plant Biol. 53, 477-501.
Schaefer, D. G. and Zryd, J. P.(1997). Efficient gene targeting in the moss Physcomitrella patens. Plant J.11, 1195-1206.
Schaefer, D. G., Delacote, F., Charlot, F., Vrielynck, N., Guyon-Debast, A., Le Guin, S., Neuhaus, J. M., Doutriaux, M. P. and Nogue, F.(2010). RAD51 loss of function abolishes gene targeting and de-represses illegitimate integration in the moss Physcomitrella patens. DNA Repair (Amst.)9, 526-533.
Snowden, K. C., Simkin, A. J., Janssen, B. J., Templeton, K. R., Loucas, H. M., Simons, J. L., Karunairetnam, S., Gleave, A. P., Clark, D. G. and Klee, H. J.
(2005). The decreased apical dominance1/Petunia hybrida CAROTENOID CLEAVAGE DIOXYGENASE8 gene affects branch production and plays a role in leaf senescence, root growth, and flower development. Plant Cell17, 746-759.
Sorefan, K., Booker, J., Haurogne, K., Goussot, M., Bainbridge, K., Foo, E., Chatfield, S., Ward, S., Beveridge, C., Rameau, C. et al.(2003). MAX4 and RMS1 are orthologous dioxygenase-like genes that regulate shoot branching in Arabidopsis and pea. Genes Dev.17, 1469-1474.
Tamura, K., Dudley, J., Nei, M. and Kumar, S.(2007). MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol.24, 1596-1599.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F. and Higgins, D. G.(1997). The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25, 4876-4882.
Timme, R. E. and Delwiche, C. F.(2010). Uncovering the evolutionary origin of plant molecular processes: comparison of Coleochaete (Coleochaetales) and Spirogyra (Zygnematales) transcriptomes. BMC Plant Biol. 10, 96.
Trouiller, B., Schaefer, D. G., Charlot, F. and Nogue, F.(2006). MSH2 is essential for the preservation of genome integrity and prevents homeologous
recombination in the moss Physcomitrella patens. Nucleic Acids Res. 34, 232-242.
Umehara, M., Hanada, A., Yoshida, S., Akiyama, K., Arite, T., Takeda-Kamiya, N., Magome, H., Takeda-Kamiya, Y., Shirasu, K., Yoneyama, K. et al.
(2008). Inhibition of shoot branching by new terpenoid plant hormones. Nature 455, 195-200.
Wang, B. and Qiu, Y. L.(2006). Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza16, 299-363.
Wang, B., Yeun, L. H., Xue, J. Y., Liu, Y., Ane, J. M. and Qiu, Y. L.(2010). Presence of three mycorrhizal genes in the common ancestor of land plants suggests a key role of mycorrhizas in the colonization of land by plants. New Phytol.186, 514-525.
Watson, M. A.(1981). Chemically mediated interactions among juvenile mosses as possible determinants of their community structure. J. Chem.7, 367-376.
Yoneyama, K., Xie, X., Sekimoto, H., Takeuchi, Y., Ogasawara, S., Akiyama, K. and Hayashi, H.(2008). Strigolactones, host recognition signals for root parasitic plants and arbuscular mycorrhizal fungi, from Fabaceae plants. New Phytol.179, 484-494.
Yoneyama, K., Xie, X. and Takeuchi, Y.(2009). Strigolactones: structures and biological activities. Pest Manag. Sci.65, 467-470.