J.micropalaeontol., 3(2): 53-64, September 1984
Structure and encystment in some fossil cavate
dinoflagellate cysts
GEOFFREY L. EATON
BP Research Centre, Sunbury-on-Thames, Middlesex, England
ABSTRACT-A study of some peridinioid and gonyaulacoid cavate cysts which develop a dorsal archaeopyle with a double aperture reveals two significant morphological differences. First, the physical separability of the outer and inner body can only be demonstrated in the peridinioid cysts. Second, although the peridinioid double aperture is covered by two opercula, only the inner aperture is operculate in the corresponding gonyaulacoid archaeopyle. These differences reflect two fundamentally different structural and encystment styles. In the peridinioid cysts, the outer and inner bodies are structurally independent and represent the separate development of outer and inner walls. In the gonyaulacoid cysts, the outer and inner bodies are structurally more intimately related, and represent the differentiation of a single wall into outer and inner layers.
INTRODUCTION
Downie & Sarjeant (1 966) introduced the cavate cyst concept to accommodate fossil taxa such as Deflandrea phosphoritica Eisenack (Pl. 1, fig. 5) in which the cyst
shows a clear separation into an outer and inner body. Conventional interpretation of this structural arrange- ment has the outer body formed by the outer cyst wall (periphragm), and the inner body (capsule) formed by the inner cyst wall (endophragrn). The separating space between the outer and inner bodies is termed the pericoel; the encapsulated space is the endocoel.
In terms of thecal affinity, the majority of cavate cysts have apparently been produced by peridinioid and gonyaulacoid dinoflagellates, and the cysts of both thecal groups often appear to be similarly constructed. This similarity is particularly true of some peridinioid taxa of the Deflandrea and Wetzeliella type, and elongate gonyaulacoid taxa such as the genera Tubo- tuberella Vozzhennikova and Hystrichosphaeropsis Deflandre, and selected species of Gonyaulacysta (Deflandre). However, a study of certain species from these two groups reveals two interesting differences concerning the physical separability of the outer and inner body, and the opercular style of the simple dorsal double-aperture archaeopyle.
The aim of this paper is to describe these differences and discuss their significance in terms of the interpret- ation of structural and encystment style.
The fossil material illustrated in this paper is housed at two localities, where it is available for examination. In the plate and text-figure explanations, slide numbers prefixed
“V”
are located in the palaeontological collec- tion of the British Museum (Natural History), London;numbers prefixed “A” and “S” are located in the palaeontological collection of the Exploration & Pro- duction Division, BP Research Centre, Sunbury-on- Thames, Middlesex, England. The co-ordinates following each slide number refer to a reversed 3” x 1” ‘England Finder’, used with a Leitz Dialux 20 Microscope. Slides were positioned to be read from the right.
STRUCTURE IN CAVATE CYSTS
Separability of the outer and inner cyst body
Peridinioid cysts. In addition to typical examples of
Deflandrea phosphoritica (Pl. 1, fig. 5 ) , Palaeogene cyst assemblages also yield acapsulate outer bodies (Pl. 1, fig. 6; Williams & Downie, 1966, pl. 26, fig. 2), and isolated inner bodies (Manum, 1960, pl. 1, figs. 7, 8; Cookson & Eisenack, 1961, pl. 1, fig. 4). This range of preservational form is comparable with that reported for the cavate cysts of certain Recent peridinioid dino- flagellates (Evitt & Wall, 1968; Norris & McAndrews,
1970).
endophragm in Recent peridinioid cavate cysts: the same interpretation can be made for D . phosphoritica.
Comparable structural relationships can be similarly demonstrated in other fossil taxa. Records of acapsulate examples of fossil peridinioid cavate cysts include Amphidiadema rectangularis (Cookson & Eisenack) in Eisenack (1971, pl. 11, fig. 4); Chatangiellu sp. (Cox, 1971, pl. 7, fig. 2); Zsabelidinium sp. (Evitt, 1961, pl. 1, fig. 1); Wetzeliella spp., this paper (Pl. 1, figs. 1, 3). The specific recognition of isolated inner bodies in fossil cyst assemblages is more difficult (Agelopoulos, 1967, p. 53). Identification is aided by theinner body having a distinc- tive shape as in Kisselovia tenuivirgula (Williams &
Downie) (Pl. 1, fig. 7), or a distinctive surface texture as in D . phosphoritica.
The occurence of isolated inner bodies does not require the complete destruction of the outer body. A rich Middle Eocene cyst assemblage from southern England (Sample W40 in Eaton, 1976) yields several inner bodies of Wetzeliella sp. (Pl. 1, fig. 2) as well as their acapsulate outer bodies (Pl. 1, fig. 1). The irregular tear in the latter specimen seems large enough to allow the inner body to be lost. This also suggests that the occurence of acapsulate outer bodies need not always reflect the failure of the inner body to develop. The second illustrated species of Wetzeliella (Pl. 1, fig. 3) is also represented by an acapsulate example. It shows some damage, and therefore its acapsulate state could be either primary or secondary. These observations only serve to confirm how readily the outer and inner cyst body can be separated.
Gonyaulacoid cysts. Late Jurassic, Oxfordian sediments in the North Sea yield particularly rich well-preserved dinoflagellate cyst assemblages. These assemblages are often dominated by gonyaulacoid cavate taxa such as Scriniodinium crystallinum (Deflandre), Gonyaulacysta jurassica (Deflandre) s. l . , and Endoscrinium galeritum
(Deflandre) to a degree comparable with peridinioid cavate domination in the Late Cretaceous and Palaeo- gene. However, despite this cavate dominance, and my deliberate searching, these assemblages have failed to yield any acapsulate examples of gonyaulacoid cavate cyst taxa, or their isolated inner bodies. Furthermore, there are no such records in the palaeontological litera- ture. One form that does require comment here is “Dinoflagellate type 1” of Fenton, Neves & Pie1 (1980) from the Middle Jurassic, which shows some similarity in overall shape to Tubotuberella Vozzhennikova. I have examined the single recorded specimen and I believe its structural affinities are closer to the Chytroeisphaeridia (Sarjeant) cyst type: a simple, single-walled cyst. It may well have been produced by a Tubotuberella-like dino- flagellate, but I do not consider it to represent an acap- sulate outer body of Tubotuberella.
It is, of course, possible that this absence of gonyaula- coid acapsulate outer bodies and isolated inner bodies is simply due to the vagaries of palaeontological obser- vation and selective reporting; however, I think this is unlikely. I believe that the absence is a genuine one, and that it reflects a different, more intimate structural relationship between the outer and inner body in gonyaulacoid cavate cysts. As a result, the two bodies are not readily separable.
Explanation of Plate 1
The ‘England Finder’ co-ordinates follow the slide number.
Figs. 1-3. Wetzelielfu spp., M. Eocene, southern England: fig. 1, Acapsulate outer body with physical damage (x 400), V.57757, Q57/4; fig. 2, Isolated inner body with endoperculum displaced but retained within the body ( x 400), V.57757, T24/3; fig. 3, Acapsulate outer body with slight physical damage ( x 400), V.57757, J38/0.
Figs. 4, 7. Kisseloviu tenuivirgula (Williams & Downie), M. Eocene, southern England: fig. 4, Complete specimen showing the pentagonal outline of the inner body ( x 400), V.57757, M30/0; fig. 7, Isolated inner body with pentagonal outline ( x 400), V.57757, 053/2.
Figs. 5 , 6. Deflandrea phosphoritica Eisenack, M. Eocene, southern England: fig. 5, Typical specimen showing rounded outline of inner body ( x 500), V.57771, R42/3; fig. 6, Acapsulate outer body with periarchaeopyle formed ( x 500), V.57766, R58/3.
Archaeopyle and opercular style
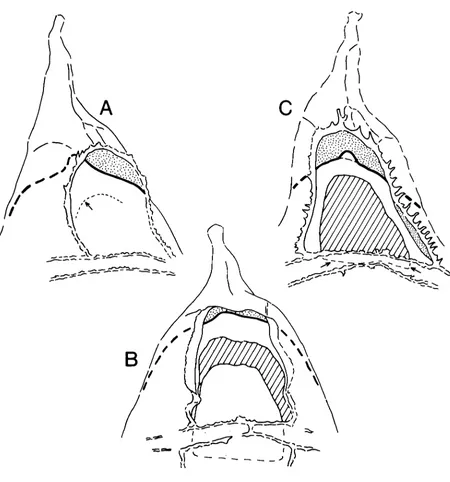
Peridinioid cysts. A characteristic feature of the fossil peridinioid cavate cyst types mentioned in this paper is the development of a simple anterior dorsal archaeopyle involving the displacement of a single apical paraplate, 4' (see Eaton, 1980, p. 675 and fig. 7). An aperture is developed in both the outer and inner body, and both openings (outer, periarchaeopyle; inner, endoarchaeo- pyle; Lentin & Williams, 1976) are operculate. This is well shown by Dracodinium pachyderma (Caro) (Pl. 1, figs. 8-12; Fig. lA, B). The size relationship between the two openings (and their attendant opercula) is extremely variable. When the depth of pericoel develop- ment is limited, and the outer and inner bodies are similar in surface area (as in D . pachyderma), the two operculate openings are similar in size. When the surface area of the outer body is significantly greater than the inner body, then the operculate periarchaeopyle is noticeably larger, as in Chatangiella coronata (McIntyre) in McIntyre (1975, pl. 3, figs. 1-4).
One other significant aspect of this archaeopyle type concerns the planes of the two apertures. In the majority of cases, where the two opercula are separate, the two apertural planes are either parallel, or convergent to beyond the posterior archaeopyle margin. When the two opercula are partially united, as in Deflandrea antarctica Wilson (Wilson, 1967, figs. 23, 24, 26, 27), the two apertural planes converge towards the posterior margin of the archaeopyle.
The development of two operculate openings to form a simple archaeopyle further emphasises the structural independence of the outer and inner cyst walls in these peridiniod cavate cysts.
Gonyaulacoid cysts. In elongate gonyaulacoid genera
such as Gonyaulacysta (Deflandre) and Tubotuberella Vozzhennikova, the archaeopyle is simple, mid-dorsal, and involves the displacement of precingular paraplate
3". Epicystal periocoel development in these genera frequently extends onto the area of 3"; in this case the archaeopyle has a double aperture.
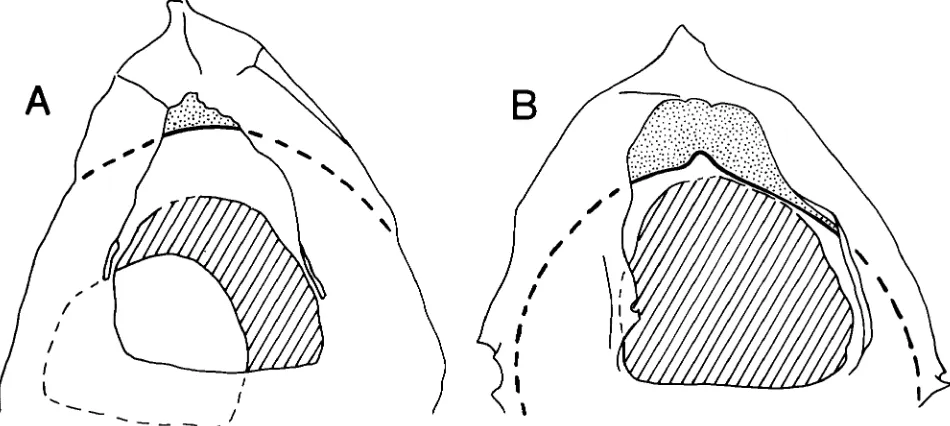
This double aperture is clearly displayed by the specimen of Gonyaulacysta jurmsica in PI. 2, figs. 1-3 and in Fig. 2C. This specimen is exceptional in that the extent of the outer periarchaeopyle can be fully appreci- ated: it is larger than the inner endoarchaeopyle and its limits are defined by the proximal margin of the para- sutural ornament around 3". Also, the two apertural planes are not convergent over the area of 3". This relationship is simply a function of the degree of separ- ation of the outer and inner body in this particular specimen. Separation occurs throughout the area of paraplate 3", and extends posteriorly over and beyond the paracingular zone.
In more typical specimens of G . jurmsica (Pl. 2, figs. 7, 8; Fig. 2B) showing more limited epicystal pericoel development, the two apertural planes converge in the anterior portion of the archaeopyle. Here, only the anterior area of the periarchaeopyle is separated and therefore distinguishable. This more typical relationship between the two openings is often difficult to interpret, and probably for this reason this archaeopyle type has received little attention. In the recent review of Gony- aulucysta and its allies (Sarjeant, 1982), for instance, the archaeopyle in Gonyaulacysta (and Tubotuberella) is described simply as having a "reduced operculum".
I
'-'
I
Some fossil cavate dinoflagellate cysts
Turning now to opercular style, it is interesting that G . jurussica does not show evidence of an opercular covering for its periarchaeopyle. This absence has also been reported for Hystrichosphaeropsis quasicribrutu (Wetzel) by Gocht (1976, p. 325) in a paper which also beautifully illustrates the typical archaeopyle style of the elongate gonyaulacoid cyst type (Gocht, 1976, figs. 14a, d). Despite the presence of two archaeopyle apertures, and in contrast to the peridinioid opercular style dis- cussed earlier, there appears to be only a single oper- cular piece in G. jurassica. In the specimen shown in PI. 2, figs. 7, 8 and Fig. 2B, the single opercular piece has been displaced but is retained within the cyst. It clearly only covers the inner opening. If the absence of an outer operculum is genuine, rather than a function of preser- vation, then the specimen of G . jurussica in PI. 2, figs. 4, 5 and Fig. 2A is complete. This is despite the presence of a large opening into the pericoel, anterior to the in-place operculum.
Tubotuberella Vozzhennikova, (Pl. 2, figs. 6,9; Fig. 3) has the same archaeopyle and opercular style as G . jurassica, but displays considerably less parasutural
ornament. In T . upatela (Cookson & Eisenack) (Fig. 3A) there is no parasutural ornament to define the postero- lateral and posterior margins of the periarchaeopyle. In T . rhombiformis Vozzhennikova (Fig. 3B) parasutural crests fully define the lateral margins of the periarchaeo- pyle, but again there is no definition of its posterior margin. In Tubotuberella then, the periarchaeopyle is not a fully defined opening, and therefore it cannot be covered by a definable operculum.
These observations indicate that no periphragm is developed over the area of paraplate 3" in these particu- lar gonyaulacoid cysts.
Summary. The structural relationships around the dorsal archaeopyle in the cyst types discussed above, are summarised in Fig. 4.
In the peridinioid archaeopyle (Fig. 4A), both the peri- and endoarchaeopyle are operculate, with separate or partially united opercula. This emphasises the struc- tural independence of the two cyst walls. The ready separation of the outer and inner body indicates that, in areas away from pericoel development, e.g. posterior to the archaeopyle, the two walls retain their structural independence even where they are in contact with each other.
In Gonyuulacysta jurassica (Fig. 4B), only the endo- archaeopyle is operculate, with the periarchaeopyle developed as a primary, non-operculate opening defined by the proximal margin of the parasutural crests. The lack of evidence for the separability of the outer and inner body in G. jurassica suggests a more complex wall structure than in the peridinioid cyst type. This com- plexity is perhaps best explained by treating periphragm and endophragm in G. jurussica as differentiated layers of a single cyst wall, rather than different walls. Away from the area of pericoel development the periphragm only expresses its presence through the development of parasutural ornament, and the two wall layers remain partially undifferentiated. This would maintain a more intimate structural relationship between the outer and inner body.
Explanation of Plate 2
All photomicrographs are x 600; the 'England Finder' co-ordinates follow the slide number.
Figs. 1-5. Gonyuulucysta jurussicu (Deflandre) subsp. udecta Sarjeant var. longicornis Deflandre, E. Oxfordian, Northern North Sea: fig. 1, Exceptional specimen with pericoel developed over the entire mid-dorsal precingular area, and extending posteriorly beyond the paracingulum. This high focus view shows the discrete posterior margin of the endoarchaeopyle in a different plane to the periarchaeopyle, A.590.4, M48/2; fig. 2, Same specimen, mid-focus, showing the separate anterior margins of the peri- and endoarchaeopyle; fig. 3, Same specimen, mid- low focus, showing the extensive pericoel development; fig. 4, Complete specimen with operculum in place. This low focus view shows the anterior margin of the non-operculate periarchaeopyle, A.1568.3, R49/4; fig. 5, Same specimen, mid-focus, showing the anterior margin of the operculum.
Fig. 6. Tubotuberella rhombiformis Vozzhennikova, probable Volgian, Central North Sea. Mid-focus showing the extent of pericoel development, A.729.2, J37/2.
Figs. 7, 8. Gonyaulucystu jurassica (Deflandre) subsp. jurussica Deflandre, L. Oxfordian, Central North Sea: fig. 7, Specimen with operculum displaced, but retained within the cyst. High focus showing the separate anterior margins of the peri- and endoarchaeopyle, A.859.2, G3 1/2; fig. 8, Same specimen, mid-focus, showing the outline of the operculum.
Some fossil cavate dinoflagellate cysts
h
- - _
-/Fig. 3. Archaeopyle and opercular style in Tubotuberella ( x 1250). A. T . apatela. Specimen with operculum displaced, but retained within the cyst. Note that the posterior and postero-lateral margins of the peri- archaeopyle are undefined, due to the absence of parasutural crests. Reversed, lower surface of specimen; S.9579.4, E22/4. B. T . rhombiformis. Specimen with parasutural crests defining the lateral margins of the periarchaeopyle; the posterior margin remains undefined; A.729.2, 53712.
In Tubotuberella (Fig. 4C) the structural relationship is identical to G. jurassica, but there is n o parasutural crest to define the posterior margin of the periarchaeo- pyle. Consequently, the narrow strip of endophragm immediately posterior to the endoarchaeopyle cannot be distinguished from the combined periphragm and endophragm of the more posterior paracingular zone and beyond.
ENCYSTMENT
On the evidence presented in this paper, particular peridinioid and gonyaulacoid cavate cysts can be divided into a different number of physically separable com- ponent parts. These are illustrated in Fig. 5, which also serves to summarise the structural findings of this paper. This difference in structural style must also reflect a difference in encystment style between particular peridinioid and gonyaulacoid dinoflagellates.
The encystment style which produced the peridinioid cavate cyst of Fig. 5 would have been very similar to that described for Recent Peridinium limbatum by Evitt &
Wall (1968). After initial contraction of the encysting protoplast away from its thecal covering, the outer body is formed by the secretion of the outer wall. Following further contraction of the protoplast, the inner body is formed by the secretion of the inner wall. Failure to develop the inner body apparently does not affect the
outer body, and a non-viable acapsulate cyst is pro- duced. Secondary damage to the outer body of normal cysts can lead to the occurrence of displaced isolated inner bodies.
This encystment style coupled with eventual excyst- ment through a simple dorsal, double aperture, double opercular archaeopyle has a long geological history, ranging through much of the Late Cretaceous and the Palaeogene. It is certainly expressed by fossil peridinioid deflandreacean cysts of the subfamily Wetzelielloideae Vozzhennikova, and the Tribes Deflandreae Bujak &
Davies and Chatangielleae Vozzhennikova of the sub- family Deflandreoideae Bujak & Davies (see Bujak &
Davies, 1983). With different archaeopyle types, this encystment style has an even longer history, extending to the present day in taxa such as Peridinium limbatum.
Some fossil cavate dinoflagellate cysts
areas which do retain contact, (essentially equatorial), remain structurally unstable until secretion of the inner wall layer ultimately defines the inner body. Failure to develop the inner body layer prevents the outer body layer from becoming entirely structurally stable, and it eventually disintegrates. This explains the absence of acapsulate outer bodies of gonyaulacoid cavate cysts in the fossil record. The extent of the area of potential instability must show some variation for a given cyst species, and is presumably genetically controlled. If encystment is successfully completed, the outer and
Fi
inner wall layers co-exist in their areas of contact in a stable intimately related form. The strength of this structural relationship is such that secondary damage does not lead to the occurrence of displaced, isolated inner bodies.
This encystment style coupled with eventual excyst- ment through a simple dorsal, double aperture, single opercular archaeopyle is well represented in the Late Jurassic and Early Cretaceous and can certainly be recognised in the Late Cretaceous and Tertiary. It is best expressed by genera such as Gonyaulacysta, Tubo-
P E R I D I N I O I D
C A V A T E
G
0
N Y A U L A C
01
D
C A V A T
E
Fig. 5. Component parts of cavate cysts (diagrammatic, not to scale). Peridinioid cavate: left,
Some fossil cavate dinoflagellate cysts
tuberella, Psaligonyaulax and Hystrichosphaeropsis, but may well be applicable to other gonyaulacoid cavate genera.
DISCUSSION
Although it would be unrealistic to apply these inter- pretations of wall structure to all fossil peridinioid and gonyaulacoid cysts, there seems little doubt that they are at least widely applicable. The formal taxonomic groups mentioned in connection with peridinioid encystment style in the previous section of this paper include a large proportion of known fossil peridinioid species. Also, my observations on gonyaulacoid wall structure comple- ment the detailed ultra-thin section studies of Jux (e.g. 1968a, b; 1971) on a range of non-cavate gonyaulacoid cyst types: Jux showed that although two layers can be recognised, it is difficult to separate them optically where they are in contact. Again, there is the possibility of a broad taxonomic distribution of this particular wall structural style.
This being the case, these structural interpretations raise some interesting points concerning dinoflagellate cyst morphology in general. For instance, what is the relationship between the walls of peridinioid cysts and the wall layers of gonyaulacoid cysts? Are the two types of wall simply different expressions of the same “double- phase” structure, or is the relationship more complex? Also, have the two structural styles evolved indepen- dently, or is one derived from the other; if the latter, then in which direction has evolution taken place?
Throughout this paper I have applied the terms peri- phragm and endophragm and their derivatives to both peridinioid and gonyaulacoid cysts, mainly for consist- ency and ease of description. If, however, the peridinioid walls and gonyaulacoid wall layers are not directly
related, then there may be a need for a dual terminology. This would certainly go against the recommendations of Cox (1 971), who discussed at length the problems of cyst body structural nomenclature, and advocated a unified terminology.
Finally, is there a direct relationship between wall structural style and morphological potential in dino- flagellate cysts? There is no doubt that cysts of proven or presumed gonyaulacoid affinity show the greater range of morphological and textural variation. Does the close structural relationship of their wall layers provide the basis for greater morphological expression? Peridinioid cysts tend to be morphologically conservative in com- parison; much of their variation involves subtle differ- ences in wall texture and degree of wall separation, with the occasional ability to produce variable, multi-walled cysts. This last feature in particular appears to emphasise the structural independence of peridinioid cyst walls, which may have an inhibiting effect on overall morpho- logical expression.
Until there is more detailed information available on wall structure in a greater range of peridinioid and gonyaulacoid cyst taxa, these questions must remain unanswered.
ACKNOWLEDGEMENTS
REFERENCES
Agelopoulos, J. 1967. Hystrichosphiiren, Dinoflagellaten und Foraminiferen aus dern eoziinen Kieselton von Heiligenhafen, Holstein. Published doctoral thesis, Eberhard-Karls Univer- sity, Tubingen. 74pp., 14 pls.
Bujak, J. P. & Davies, E. H. 1983. Modern and fossil Peridini- ineae. Am. Ass. stratigr. Palynol., Contr. Ser. 13, 203pp.,
Cookson, I. C. & Eisenack, A. 1961. Tertiary microplankton from the Rottnest Island Bore, Western Australia. J. Proc. r .
SOC. West. Aust., Perth, 44 (2), 39-47, pls. 1-2.
Cox, R. L. 1971. Dinoflagellate cyst structures: walls, cavities, and bodies. Palaeontology, London, 14 ( l ) , 22-33, pl. 7. Downie, C. & Sarjeant, W. A. S. 1966. The morphology, ter-
minology and classification of fossil dinoflagellate cysts. Pp. 10-17. In Davey, R. J., Downie, C., Sarjeant, W. A. S. &
Williams, G. L. Studies on Mesozoic and Cainozoic dino- flagellate cysts. Bull. Br. Mus. nat. Hist., London, (Geol),
Eaton, G. L. 1976. Dinoflagellate cysts from the Bracklesham Beds (Eocene) of the Isle of Wight, southern England. Bull. Br. Mus. nat. Hist., London, (Geol.), 26 ( 6 ) , 225-332, pls.
1-2 1.
Eaton, G. L. 1980. Nomenclature and homology in peridinia- lean dinoflagellate plate patterns. Palaeontology, London,
Eisenack, A. 1971. Anomalien bei fossilen Dinoflagellaten. Paliiont. Z., Stuttgart, 45 (1/2), 75-78, pl. 11.
Evitt, W. R. 1961. Observations on the morphology of fossil dinoflagellates. Micropaleontology, New York, 7 (4), Evitt, W. R. & Wall, D. 1968. Dinoflagellate studies IV. Theca and cyst of Recent freshwaterperidinium limbatum (Stokes) Lemmermann. Stanf. Univ. Publs (Geol. Sci.), Palo Alto, 12 Fenton, J. P. G., Neves, R. & Piel, K. M. 1980. Dinoflagellate
cysts and acritarchs from Upper Bajocian to Middle Bathonian strata of central and southern England. Palaeon- tology, London, 23 (l), 151-170, pls. 14-16.
Gocht, H. 1976. Hystrichosphaeropsis quasicribrata (0. Wetzel), ein Dinoflagellat aus dem Maastricht Nordeuropas. Mit einem nomenklatorischen Nachtrag zur Gattung Litho- dinia Eis.Neues Jb. Geol. Paliiont. Mh., Stuttgart, 1976, ( 6 ) ,
321-336, figs. 1-18. 12 pls.
SUPPI. 3, 1-248, PIS. 1-25.
23 (3), 667-688.
385-420, PIS. 1-9.
(2), 1-15, PIS. 1-4.
Jux, U. 1968a. Uber den Feinbau der Wandung bei Cordo- sphaeridium inodes (Klumpp 195 3). Palaeontographica, Stuttgart, B 122 (1-3), 48-54, PIS. 13-14.
Jux, U. 1968b. Uber den Feinbau der Wandung bei Hystricho- sphaera bentori Rossignol 1 96 1. Palaeontographica,
Stuttgart, B 123 (1-6), 147-152, pl. 31.
Jux, U. 1971. Uber den Feinbau einiger tertiarer Dinophyceen- Zysten und Acntarcha. Hystrichosphaeridium, Impleto- sphaeridium, Lingulodinium. Palaeontographica, Stuttgart, Lentin, J. K. & Williams, G. L. 1976. A monograph of fossil peridinioid dinoflagellate cysts. Bedford Inst. Oceanogr. Rept. B1-R-75-16, 1-237.
Manum, S. 1960. Some dinoflagellates and hystrichosphaerids from the Lower Tertiary of Spitsbergen. Nytt Mag. Bot., McIntyre, D. J. 1975. MorphologicchangesinDeflandrea from a Campanian section, District of Mackenzie, N.W.T., Canada. Geoscience and Man, Baton Rouge, 11,61-76, pls.
1-4.
Norris, G. & McAndrews, J. H. 1970. Dinoflagellate cysts from post-Glacial lake muds, Minnesota (U.S.A.). Rev. Palaeobot. Palynol., Amsterdam, 10 (2), 131-156, pls. 1-3.
Sarjeant, W. A. S. 1982. The dinoflagellate cysts of the Gonyaulacysta Group: a morphological and taxonomic restudy. A m . Ass. stratigr. Palynol., Contr. Ser. 9, 81 pp.,
12 PIS.
Williams, G. L. & Downie, C. 1966. Further dinoflagellate cysts from the London Clay. Pp. 215-235. In Davey, R. J., Downie, C., Sarjeant, W. A. S., & Williams, G. L. Studies on Mesozoic and Cainozoic dinoflagellate cysts. Bull. Br. Mu. nat. Hist., London, (Geol.), Suppl. 3, 1-248, pls. 1-25. Wilson, G. J. 1967. Some new species of Lower Tertiary dino-
flagellates from McMurdo Sound, Antarctica, N . Z. Jl Bot., Wellington, 5 (l), 57-83, figs. 1-42.
B 132, 165-174, PIS. 25-28.