1535-9778/05/$08.00⫹0 doi:10.1128/EC.4.4.787–798.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
A Point Mutation in the Cargo-Binding Domain of Myosin V Affects
Its Interaction with Multiple Cargoes
Natasha Pashkova,† Natalie L. Catlett,†‡ Jennifer L. Novak, and Lois S. Weisman*
Department of Biochemistry, University of Iowa, Iowa City, IowaReceived 20 December 2004/Accepted 26 January 2005
Class V myosins move diverse intracellular cargoes, which attach via interaction of cargo-specific proteins to the myosin V globular tail. The globular tail of the yeast myosin V, Myo2p, contains two structural and functional subdomains. Subdomain I binds to the vacuole-specific protein, Vac17p, while subdomain II likely binds to an as yet unidentified secretory vesicle-specific protein. All functions of Myo2p require the tight association of subdomains I and II, which suggests that binding of a cargo to one subdomain may inhibit cargo-binding to a second subdomain. Thus, two types of mutations are predicted to specifically affect a subset of Myo2p cargoes: first are mutations within a cargo-specific binding region; second are mutations that mimic the inhibited conformation of one of the subdomains. Here we analyze a point mutation in subdomain I,
myo2-2(G1248D), which is likely to be this latter type of mutation.myo2-2 has no effect on secretory vesicle
movement. The secretory vesicle binding site is in subdomain II. However, myo2-2 is impaired in several Myo2p-related functions. While subdomains I and II of myo2-2p tightly associate, there are measurable differences in the conformation of its globular tail. Based solely on the ability to restore vacuole inheritance, a set of intragenic suppressors ofmyo2-2 were identified. All suppressor mutations reside in subdomain I. Moreover, subdomain I and II interactions occurred in all suppressors, demonstrating the importance of subdomain I and II association for Myo2p function. Furthermore, 3 of the 10 suppressors globally restored all tested defects inmyo2-2. This large proportion of global suppressors strongly suggests thatmyo2-2(G1248) causes a conformational change in subdomain I that simultaneously affects multiple cargoes.
Class V myosins are actin-based molecular motors that func-tion in relatively long-range movements of many intracellular cargoes including organelles, membrane vesicles, and mRNA. These motors are ubiquitously found in all eukaryotes, and the corresponding genes or cDNAs have been cloned from multi-ple species (3, 28). The in vivo functions of several myosin V motors have been extensively studied in both vertebrate ani-mals and yeast.
Mammals have three myosin V motors: myosin Va, myosin Vb, and myosin Vc (28). Myosin Va is the best characterized mammalian myosin V. In mouse, myosin Va is encoded by the dilute locus. The dilute mutants have a lightened coat color, and null mutants die of a neurological disorder within a few weeks of birth (32). Similarly, mutations in the human ortholog of dilute cause Griscelli’s syndrome, a rare recessive disease characterized by neurological and pigmentation defects (31, 36). Several studies have demonstrated that myosin Va is im-portant for melanosome transport in melanocytes (34, 37, 39, 49, 51) and transport of smooth endoplasmic reticulum in Purkinje cells (46). In addition, myosin Va moves chromaffin vesicles (41) and membranous vesicles in nerve cells (5, 12). While these myosin Va studies report on the movement of a single cargo within a selected cell type, it is likely that myosin Va moves multiple cargoes within a single type of cell (9).
In the budding yeastSaccharomyces cerevisiae,MYO2is an essential myosin V. Defects inMYO2abolish polarized growth by disrupting the delivery of secretory vesicles (14, 24, 44). In
addition to secretory vesicle transport, Myo2p moves many yeast organelles including the vacuole (8, 16, 50), the late Golgi (42), peroxisomes (17), and mitochondria (4, 21, 22). Myo2p is also required for mitotic spindle orientation (1, 53).MYO4, a nonessential yeast myosin V, moves the peripheral endoplasmic reticulum (11) and in addition transports several mRNAs (45). Myosin V moves via attachment of its amino terminal head (motor) domain to actin cables; its carboxyl terminal tail do-main anchors it to cargoes via attachments to organelle-specific receptors. Individual myosin V motors move multiple cargoes, where the cargoes move to distinct locations at different times. Both the spatial and temporal regulation of movement of an individual cargo occurs in part via organelle-specific receptors. To date, a few organelle-specific myosin V receptors have been identified. The melanosome-specific myosin Va receptor is composed of melanophilin, which binds directly to myosin Va and simultaneously to Rab27a (52). Similarly, the myosin Vb globular tail interacts with Rab11a and the Rab11 family in-teracting protein 2 (15). The vacuole-specific myosin V re-ceptor is composed of Vac17p, which binds directly to yeast Myo2p and simultaneously to the vacuole membrane protein Vac8p (6, 20, 47). In a separate functional complex, the Myo2p globular tail binds directly to Kar9p, which in turn binds to the microtubule end binding protein, Bim1p/EB1 (1, 53).
Several studies have demonstrated that the myosin V glob-ular tail can bind to cargoes in the absence of a motor domain. For example, overexpression of the wild-type Myo2p globular tail has a dominant-negative effect and disrupts secretory ves-icle movement, which results in cell death (7, 38, 44). Similarly, overexpression of the myosin Va globular tail in melanocytes disrupts melanosome movement. Also, recombinant proteins of green fluorescent protein fused to the myosin Va globular * Corresponding author. Mailing address: Department of
Biochem-istry, The University of Iowa, Iowa City, IA 52242. Phone: (319) 335-8581. Fax: (319) 384-4770. E-mail: [email protected].
† N.P. and N.L.C. contributed equally.
‡ Present address: 737 Snapdragon St., Encinitas, CA 92024.
787
on September 8, 2020 by guest
http://ec.asm.org/
tail colocalize with myosin Va cargoes, such as centrosomes, and melanosomes, even in the absence of the motor domain (10, 51).
Notably, distinct regions of the myosin V globular tail ap-pear to bind different organelle-specific receptors. For example, seven single point mutations in the Myo2p globular tail were isolated that cause a vacuole or lysosome inheritance defect with-out affecting secretory vesicle movement (7, 8). Six of these mu-tations are single amino acid changes at D1297, L1301, N1304, or N1307. These amino acids lie along a face of a predicted␣ -helix (7). Subsequent studies have strongly suggested that these residues directly bind to the vacuole-specific receptor, Vac17p (6, 20). Likewise, point mutations that were identified in the globular tail of mouse myosin Va (I1510N, M1513K, or D1519G) appear to specifically affect melanosome movement (18). These mutations were later found to partially impair the binding of the myosin Va globular tail to Slac2-a/melanophilin (13).
Conversely, mutations in the Myo2p globular tail have been identified that affect secretory vesicle movement without af-fecting vacuole movement. Several conditional lethal myo2 alleles, which result from point mutations in the globular tail, do not affect the ability of Myo2p to move the vacuole. However, at the nonpermissive temperature, these mutant alleles are defective in secretory vesicle movement (44). Likewise, myo2-⌬AflII, which contains a small deletion in the Myo2p globular tail, cannot support yeast viability, presumably due to a defect in moving secretory vesicles; however,myo2-⌬AflIIfunctions in vacuole movement (7). Analysis of the above vacuole-specific and secretory vesicle-specific point mutants demonstrates that the Myo2p globular tail can be divided into a vacuole-specific region and secretory vesicle-specific region (7, 44).
These two functional regions reside within separate structural subdomains (see Results and reference 35). The vacuole-specific region is localized to subdomain I, while the secretory vesicle-specific region resides in subdomain II. Subdomains I and II tightly associate with each other, and the tight association of these subdomains is required for all Myo2p functions tested (35).
These studies led to the hypothesis that the myosin V glob-ular tail itself plays a regulatory role in specifying cargoes. Specifically, we speculate that occupancy of subdomain II by a cargo might block the binding of a cargo to subdomain I. Here we focus on a point mutant which may represent the proposed inactive conformation of subdomain I. myo2-2(G1248D) lies outside of the region that binds directly to Vac17p. Moreover,
unlike the vacuole-specific point mutations at residues that bind directly to Vac17p (D1297, L1301, N1304, or N1307), myo2-2has multiple defects. Multiple intragenic suppressors of myo2-2 were isolated and characterized. Analysis of myo2-2 and its suppressors strongly suggests that G1248 and the sur-rounding region may contribute to forming both a “closed or inactive” and “open or active” conformation of subdomain I.
MATERIALS AND METHODS
Yeast strains and plasmids.The strains used in this study are listed in Table 1. pGAD-VAC17(1–170) (20), pGAD-KAR9, and pGAD-SMY1 (35) have been described. For the pGBD-myo2(1111–1574) clones, both wild type and mutants, DNA was PCR amplified from relevant pRS416-myo2plasmids by using primer MYO2-Bam3F (7) and the T3 universal primer. PCR products were cut with BamHI and ClaI and then ligated into pGBD-C1 or pGAD-C1. To generate plasmids carrying Myo2p subdomain I (wild type or mutants) fused with GAL4 BD or GAL4 AD, DNA encoding amino acids 1139 to 1345 of Myo2p was PCR amplified from either pMYO2, pRS413-myo2-2(pNLC1) (8), or pRS416-MYO2
containing themyo2-2 suppressor mutations (see below) by using primers MYO2-Bam1F (5⬘-GCCGGATCCACATTGATTCATTG-3⬘) and MYO2-SAL1R (5⬘-CCCGTCGACTTTGTTCAAAAACCCGC-3⬘), which adds a 5⬘BamHI and 3⬘SalI site (shown in boldface). These fragments were ligated into pAS2-CYH2 (GAL4 BD fusion) or pGAD-C1. To generate pGM2-2G, the 924-kb BspEI and BlpI fragment from pGM2G (described in reference 35) was replaced with the corresponding fragment from pRS413-myo2-2.
Yeast two-hybrid assay and library screen.The GAL4 system was used for yeast two-hybrid analysis (23). The yeast strain PJ69-4A was cotransformed with the indicated GAL4 AD fusion construct (LEU2marker) and the indicated GAL4 BD fusion constructs (TRP1marker). Single colonies of each transfor-mant were patched onto plates containing synthetic complete (SC)-Leu-Trp medium. After growth for 2 days at 24°C, the patches were replica plated to the test plates (SC-Leu-Trp-His-Ade medium) containing 30 mM 3-amino-triazole (Sigma-Aldrich). Plates were incubated for 4 to 5 days at 24°C.
For the liquid-galactosidase assays, cells were inoculated into liquid cultures and incubated overnight at 24°C. The-galactosidase activity assays were per-formed at 30°C using a 30-min incubation time.
A yeast two-hybrid screen for the vacuole-specific receptor was performed by using Myo2p residues 1139 to 1345 fused with GAL4 BD (pNLC23) as bait and the yeast genomic DNA library pools PJ1 and PJ2 (23). The transformants were plated on Trp-His plates and replica plated to plates containing SC-Leu-His-Ade plus 3-amino-triazole. The clone A20, which showed a strong positive interaction, was sequenced. The sequence revealed that GAL4 AD was fused to a DNA sequence encoding amino acids 1336 to 1574 of Myo2p.
Screen for intragenicmyo2-2suppressors.The region ofmyo2-2(pNLC1) was randomly mutagenized by PCR amplification withTaqDNA polymerase (Boehr-inger) by using a strategy similar to that used to obtainmyo2mutants defective in vacuole inheritance, with the forward primer MYOR (which will amplify DNA starting with the codon that corresponds to amino acid residue 1064) and the re-verse primer MYOL (which will amplify plasmid DNA starting about 150 bp down-stream from the 3⬘end ofMYO2and amplifying back into the coding region) (7). A strain that had both thepep4⌬andmyo2-2mutations (LWY5518), containing pGAL4-PEP4-HIS3 (pNLC16), was cotransformed with the mutagenized DNA
LWY6665 MAT␣ura3-52 leu2-3,112 his3-⌬200 trp1-⌬901 lys2-801 suc2-⌬9 myo2⌬::TRP1 pmyo2-2-HIS3
LWY6666 MAT␣ura3-52 leu2-3,112 his3-⌬200 trp1-⌬901 lys2-801 suc2-⌬9 myo2⌬::TRP1 pMYO2-HIS3
aUnless noted, strains were constructed during the course of this work.
on September 8, 2020 by guest
http://ec.asm.org/
encoding theMYO2tail and pRS416-myo2-2cut with EcoRI to remove the region encoding the globular tail (residues 1117 to 1574). For pRS416-myo2-2(pNLC18), the 5-kb ClaI fragment containing themyo2-2gene was subcloned into pRS416 from pNLC1 (8). Transformants were replica plated to galactose-containing me-dium (SC-Ura-His plus 3% galactose) to induce proteinase A (PEP4) expression and then transferred to glucose-containing medium (SC-Ura plus 0.5% casamino acids) and assayed for carboxypeptidase Y activity, as described previously (7). Red and pink colonies, indicating carboxypeptidase Y function and wild-type vacuole inheritance, were further assayed by microscopy by using FM4-64 label-ing. After assessment of vacuole inheritance of the suppressors, the strains were replicated onto 5-fluoroorotic acid-containing media to remove the URA3-based plasmid containing the putativemyo2suppressor allele and thus to test that the observed suppression was due to the mutagenizedmyo2plasmid. To test whether the intragenic suppressors ofmyo2-2could function as the sole copy of Myo2p, the corresponding pRS416-myo2plasmids were cut with XhoI and SacI. The resulting fragments were subcloned into pRS413. The pRS413-myo2plasmids were introduced as the sole copy ofMYO2into LWY2949 as described (7).
Protein analysis.Expression, purification, and mild proteolysis of the recom-binant myo2-2p globular tail was performed mostly as described for Myo2p globular tail (35) with some modifications. Briefly, glutathione transferase (GST)-myo2-2p tail fusion protein was expressed inEscherichia coliBL21(DE3) from pGM2-2G. Cells were grown in Luria-Bertani medium with 100 mg of ampicillin per liter at 37°C to an optical density at 600 nm of⬃0.4. Protein expression was induced with 0.1 mM IPTG (isopropyl--D-thiogalactopyranoside). After 20 h of incubation at 22°C, cells were harvested and frozen at⫺80°C. Protein was purified by using affinity chromatography followed by cleavage from GST tag with thrombin and gel filtra-tion. For mild proteolysis experiments, purified protein was concentrated to⬃0.5 mg/ml and incubated at 0°C with 3g of trypsin (Sigma-Aldrich) per ml, with ali-quots removed at 10, 20, and 40 min. To stop the reaction, 0.1 mg of Pefabloc SC (Roche) per ml was added, and the mixture was incubated for 10 min on ice before analyzing by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) or by gel filtration on a Superose 12 HR 10/30 column (Amersham Bio-scienes Corp.). To determine the molecular weight of the polypeptides, protein sam-ples were analyzed by using matrix-assisted laser desorption ionization–time of flight (MALDI-TOF) mass spectrometry by the University of Iowa facilities. Before anal-ysis the purified samples were concentrated and desalted with a ZipTipC4pipette
tip (Millipore) containing reverse-phase media, according to vendor instructions. Levels of protein expression from pGAD or pGBD constructs were assessed by Western blot analysis. Cell extracts were separated on SDS-PAGE (10 or 4 to 15% gel) and transferred to nitrocellulose membranes overnight. The mem-branes were probed with goat anti-Myo2p tail antiserum (1:2,000) (7) or rabbit anti-GAL4 activation domain antiserum (1:2,000; Upstate Biotechnology, Inc.).
Observation of vacuoles and nuclei.Yeast vacuoles were labeled in vivo with
N-(3-triethylammoniumpropyl)-4-(6(4-(diethylamino)phenyl)hexatrienyl) pyri-dinium dibromide (FM4-64; Molecular Probes, Inc.) as described previously (7). To detect nuclei, yeast cells were fixed in ethanol and stained with DAPI (4⬘,6⬘ -diamidino-2-phenylindole) (25).
RESULTS
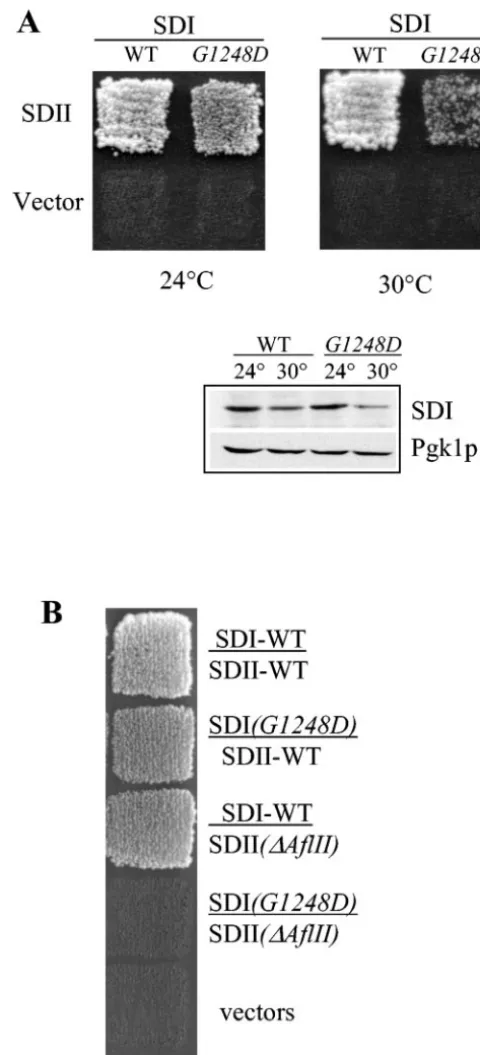
The globular tail of Myo2p contains two functional and struc-tural subdomains that interact with each other.Mild trypsin digestion of a recombinant protein that corresponds to the Myo2p globular tail (residues 1087 to 1574) results in three major polypeptides, residues 1131 to 1574, 1131 to 1345, and 1346 to 1574; the latter two polypeptides are subdomain I and subdomain II, respectively (35). Notably, subdomain I contains all of the known vacuole-specific point mutations, while sub-domain II contains six out of seven known secretory vesicle mutations. In order to identify vacuole-specific proteins that bind to the Myo2p globular tail, a yeast two-hybrid screen was performed testing a random yeast genomic DNA library with Myo2p residues 1139 to 1345 (subdomain I) as the bait. To fur-ther screen for the putative receptor, candidate positives were subsequently tested with a mutant clone encoding Myo2p resi-dues 1139 to 1345 (G1248D), which is defective in vacuole in-heritance. Of approximately 50,000 clones screened, 1 clone was obtained that interacted strongly with wild-type but not mutant subdomain I. Interestingly, this clone encoded Myo2p
subdo-FIG. 1. Myo2p globular tail subdomains I and II interact with each other. (A) In a yeast two-hybrid test, subdomain I (wild-type or myo2-2p) fused to GAL4 AD interacts with subdomain II (residues 1336 to 1574) fused to GAL4 BD, at 24 or 30°C (upper panel). A Western blot analysis of the GAL4 AD fusion proteins with Myo2p subdomain I is shown in the lower panel. Pgk1p was used as a loading control. (B) Subdomains I or II from the partially functional mutantsmyo2-2
(G1248D) andmyo2-⌬AflII(⌬1459–1491) interact with the correspond-ing wild-type subdomain in a yeast two-hybrid assay. Plasmids encod-ing a fusion protein of GAL4 BD fused to the Myo2p tail subdomain I (residues 1131 to 1345) and GAL4 AD fused to Myo2p tail subdo-main II (residues 1346 to 1574) were cotransformed into the yeast strain PJ69-4A. SDI, subdomain I; SDII, subdomain II; WT, wild type;
G1248D,myo2-2(G1248D);⌬AflII,myo2-⌬AflII(⌬1459–1491). Plates were incubated at 24°C and photographed on day 4.
on September 8, 2020 by guest
http://ec.asm.org/
main II plus an additional 10 residues (1336 to 1574) (Fig. 1A), which strongly suggested that Myo2p subdomains I and II interact.
Biochemical analysis of the Myo2p globular tail confirmed that indeed subdomains I and II tightly associate with each other and further showed that this association does not require additional proteins. Most importantly, in vivo analyses demon-strated that the tight association of these subdomains is re-quired for all tested functions of Myo2p (35). Thus, we further tested whether themyo2-2mutant is defective in subdomain I and II interactions.
As described below, subdomain I and II interactions in myo2-2are similar to those observed with the wild-type pro-tein. We initially observed a defect in subdomain I and II interactions in themyo2-2protein in a yeast two-hybrid test. While the difference in interaction strengths between wild-type andmyo2-2subdomains I and II was modest when assessed by growth on plates containing SC-His-Ade-Trp-Leu medium at 30°C (Fig. 1A), liquid-galactosidase assays showed a three-fold difference (not shown). However, these differences in the yeast two-hybrid test appear to be due to differences in protein stability and/or expression. At 30°C, where a difference was observed, lessmyo2-2subdomain I fusion protein was present (Fig. 1A). At 24°C the protein levels of wild-type andmyo2-2 subdomain I fusion proteins were similar. Under these condi-tions reproducible differences in the yeast two-hybrid test were not observed with the growth assay (Fig. 1). The subdomain II fusion protein obtained in the yeast two-hybrid screen (resi-dues 1336 to 1574) contained an additional 10 resi(resi-dues in comparison to subdomain II as defined by mild proteolysis (residues 1346 to 1574). Thus, in further two-hybrid studies we used a plasmid that encoded the GAL4 binding domain fused with Myo2p residues 1346 to 1574.
We also examined the ability of subdomains I and II to interact in the secretory vesicle-specific mutant, myo2-⌬AflII (⌬1459 to 1491). Subdomain I and II interactions occurred in this partially functional mutant. However,myo2-2subdomain I did not interact withmyo2-⌬AflIIsubdomain II (Fig. 1B). One possibility is that each of these mutants individually has a minor defect in subdomain I and II interactions, which be-comes more pronounced when both subdomains are mutated.
Subdomains I and II are present within the myo2-2p glob-ular tail.We tested whether themyo2-2globular tail has struc-tural features similar to those found within the wild-type glob-ular tail. A bacterially expressed GST fusion protein of the myo2-2p globular tail (residues 1087 to 1574) was generated. The GST tag was removed by digestion of the fusion protein at the engineered thrombin cleavage site. The resultant polypep-tide was incubated with trypsin at 0°C, and aliquots were re-moved at 10, 20, and 40 min. The results were compared with those obtained for the wild-type globular tail. Similar to results with the wild type, mild proteolysis of the myo2-2p tail resulted in three major stable polypeptides of approximately 50, 26, and 24 kDa (Fig. 2). The 50-kDa fragment resulted from trypsin digestion at residue 1130. This 50-kDa fragment was subse-quently cleaved by trypsin into subdomains I and II. MALDI-TOF mass spectrometry demonstrated that these fragments corresponded to residues 1131 to 1574 (50 kDa), 1131 to 1345 (24 kDa), and residues 1346 to 1574 (26 kDa) (Table 2). The frag-ments generated by mild trypsin digestion of the myo2-2p globu-lar tail were identical with those obtained with the wild-type tail. There were small but reproducible differences in the rate and extent of trypsin digestion of the wild-type and mutant globular tails (Fig. 2). These results suggest that while both myo2-2p and wild-type globular tails have two stable subdo-mains, there are likely to be some conformational differences between the globular tails. These potential small differences in conformation may account for the major differences observed in wild-type versusmyo2-2function.
FIG. 2. Mild proteolysis of the Myo2p and myo2-2p globular tails reveals that each protein contains two stable subdomains I (SDI) and II (SDII). The myo2-2p and wild-type globular tails were incubated with trypsin at 0°C. Samples were collected at intervals of 10, 20, and 40 min and analyzed by SDS-PAGE (15% gel). Data shown are representative of three independent experiments. Proteins were detected with the Coomassie blue R-250 stain.
TABLE 2. MALDI-TOF mass spectrometry analysis of the myo2-2p globular tail polypeptides generated
by mild trypsin digestion
Polypeptide (residues)
Predicted molecular mass (kDa)a
Experimental molecular mass (kDa)
Difference between predicted and
experimental values (Da)
Globular tail (1131–1574) 50.606 50.685 79
Subdomain I (1131–1345) 24.248 24.268 20
Subdomain II (1346–1574) 26.376 26.409 33
aCalculated from primary sequences by using ExPASy web site at http://www
.us.expasy.org/.
on September 8, 2020 by guest
http://ec.asm.org/
myo2-2subdomains I and II interact in vitro.One possibility is that conformational differences between the wild-type and myo2-2p globular tails are due to a defect in the association of subdomains I and II. To further test for potential defects in subdomain I and II interactions of myo2-2p, we tested their interaction in vitro. The subdomains were generated by mild proteolysis of the myo2-2p tail and analyzed by gel filtration. As with the wild-type protein, the subdomains generated by proteolysis of the myo2-2p globular tail migrated together on gel filtration (Fig. 3). When the resultant trypsin fragments were treated with urea and chromatographed in the presence
of urea, they migrated as two partially resolved peaks. Upon removal of urea they rapidly reassociated (Fig. 4). Therefore, by both the yeast two-hybrid test and biochemical analysis, it appears that subdomains I and II derived from the myo2-2p tail interact similarly to wild type.
myo2-2has multiple defects.myo2-2and the other
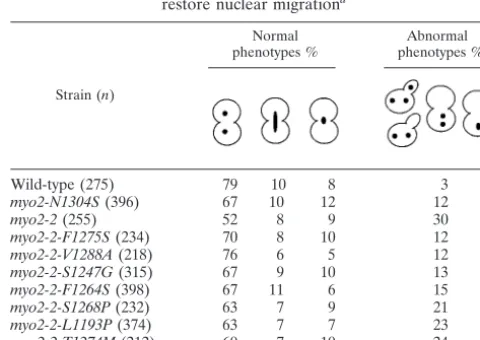
vacuole-related point mutants are defective in vacuole inheritance and do not interact with Vac17p (7, 8, 20) (Table 3). We tested wheth-er these mutants have defects in othwheth-er Myo2p-related functions. First, we tested whether interaction occurred between the cor-responding mutant myo2p globular tails with two globular tail binding partners, Kar9p and Smy1p. Neither Kar9p nor Smy1p are required for vacuole inheritance. Kar9p is required for accurate nuclear partitioning (1, 19, 29). Mutations at the Vac17p binding site, residues D1297, L1301, N1304, or N1307, did not disrupt the ability of the Myo2p globular tail to bind to Kar9p (Table 3), and a further test of one of them, myo2-N1304S, showed a very modest defect in nuclear partitioning (Table 4). In contrast,myo2-2(G1248D) was defective in its interaction with Kar9p (Table 3) and, furthermore, was de-fective in accurate nuclear partitioning (20) (Table 4).
The in vivo functions of Smy1p have not been determined, but overexpression of Smy1p suppresses the growth defects at the nonpermissive temperature of several temperature-sensi-tive alleles ofmyo2(30, 44). As was observed with Kar9p, the myo2 point mutations at residues D1297, L1301, N1304, or N1307 had no effect on the ability of Smy1p to interact with the Myo2p globular tail. In contrast,myo2-2(G1248D) was defec-tive in its interaction with Smy1p (2 and Table 3).
In addition to defects in binding Smy1p and Kar9p, myo2-2p does not accumulate at sites of polarized growth, whereas the localizations of D1297G, N1304D, and myo2p-N1307D are close to the localization of wild-type Myo2p (7). Together, these observations suggest that while Myo2p resi-dues D1297, L1301, N1304, and N1307 define a region that binds directly to Vac17p, residue G1248 may be part of a region responsible for a more global regulation of Myo2p at-tachment to cargo. Alternatively, G1248 may be in a region where the binding sites for Smy1p and Kar9p overlap with the site for Vac17p.
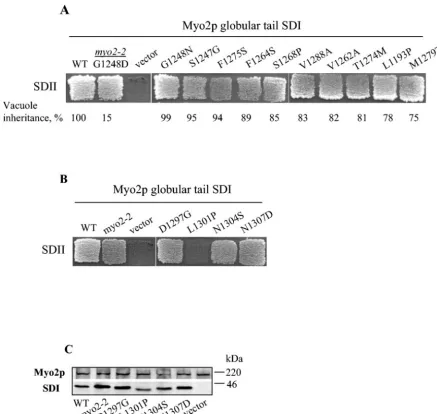
Isolation of intragenic suppressors of myo2-2. To test the hypothesis that the myo2-2 mutation is within a regulatory region of subdomain I, we sought intragenic suppressors that restore vacuole inheritance and then subsequently tested them for other Myo2p-related functions. The screen was designed so that the suppressors did not necessarily need to function in the essential functions of Myo2p; a genomic copy ofmyo2-2 was present in the strain used in the screen. Tenmyo2-2suppressor alleles were isolated that restored the vacuole inheritance de-fect from 15% to more than 75% of wild-type levels (Fig. 5A). Each of the 10 suppressors identified supported yeast viability and functioned as the sole copy ofMYO2. Notably, all of these mutations mapped to subdomain I (Fig. 5B) and restored Vac17p-Myo2p interactions (2) (Table 3). There is unlikely to be an intrinsic bias toward mutagenesis within subdomain I. When the same region of Myo2p was PCR amplified under highly mutagenic conditions, the resulting vacuole-specific mu-tants (myo2-4,myo2-5,myo2-6, andmyo2-7) contained multi-ple point mutations spanning both subdomains (7). Thus, the location of the intragenic suppressors further suggests that FIG. 3. Subdomains I and II from the myo2-2p globular tail tightly
associate with each other. The globular tails of Myo2p (panel A and red in panel C) and myo2-2p (panel B and blue in panel C) were analyzed by using mild proteolysis and gel filtration. Purified proteins (lanes 1) were cleaved with trypsin into two subdomains (lanes 2) and applied on a Superose 12 column. The subdomains from either the Myo2p tail (lane 3 in panel A) or the myo2-2p tail (lane 3 in panel B) coeluted. Protein samples were analyzed by SDS-PAGE (15% gel) and visualized with Coomassie blue R-250 (A) or silver staining (B). (C) During gel filtration, similar to the wild-type protein, the myo2-2p tail migrated as⬃55-kDa polypeptide (upper panel). After trypsin digestion of the myo2-2p tail followed by gel filtration, subdomains I and II coeluted as a single peak at 13 ml, corresponding to⬃50-kDa polypeptide. The column was calibrated with the indicated protein standards (Bio-Rad) shown as vertical arrows. SDI, subdomain I; SDII, subdomain II.
on September 8, 2020 by guest
http://ec.asm.org/
residues 1193 to 1307 define a region that is important for Myo2p interaction with Vac17p. In addition, the absence of suppressor mutations within subdomain II further suggests that subdomain I and II interactions inmyo2-2are similar to those in wild type.
Some intragenic suppressors globally restore multiple Myo2p functions.We tested whether any of these intragenic suppressors globally restore all known defects ofmyo2-2. We used a yeast two-hybrid test or functional analysis of nuclear migration (Tables 3 and 4) as a measure of Myo2p interaction with Kar9p, a yeast two-hybrid test to measure Myo2p inter-action with Smy1p (Table 3), and vacuole inheritance and a yeast two-hybrid test to measure interaction with Vac17p (Ta-ble 3).
Notably, three of the suppressors globally restored Myo2p interactions with Vac17p, Kar9p, and Smy1p. One of them was the pseudo-revertantmyo2-2-G1248N, suggesting that either glycine or asparagine can be accommodated at this position. Two second-site suppressors, myo2-2-V1262A and myo2-2-F1275S, also restored Myo2p globular tail interactions with all
binding partners tested (Table 3). We predict that themyo2-2 suppressors that restored all functions may have restored the myo2-2p globular tail to a conformation that is similar to that of the wild type.
myo2-2-M1279T, myo2-2-V1288A, myo2-2-S1247G, and myo2-2-F1264Srestored vacuole inheritance and nuclear mi-gration and/or interaction with Kar9p.myo2-2-S1268P, myo2-2-L1193P, and myo2-2-T1274M restored interaction with Vac17p alone. These partial suppressors may restore binding to a subset of proteins either by restoration of some of the original interaction sites or by generation of new interaction sites in the myo2p globular tail. This latter type of suppression is more commonly observed (43).
Subdomain I and II interactions occur within all myo2-2
intragenic suppressors and is likely required for Myo2p func-tion.The 10myo2-2intragenic suppressors provided us with an additional set of functionalMYO2alleles. Therefore, to further test the importance of subdomain I and II interactions for myo2p globular tail function, we tested interactions in each of the suppressors (Fig. 6A). We also tested the vacuole-specific FIG. 4. Subdomains I and II from the myo2-2p tail partially separate in the presence of 6 M urea, then rapidly reassociate upon the removal of urea. (A) Undigested Myo2p (red) and myo2-2p (blue) globular tails migrate as 50-kDa polypeptides on a Superose 12 column. The leftyaxis and rightyaxis indicate absorbance for Myo2p or myo2-2p, respectively. (B) Myo2p and myo2-2p subdomains I and II partially separate in the presence of 6 M urea. Subdomains I and II were generated by trypsin digestion (20 min at 0°C). The resultant polypeptides were incubated with 6 M urea for 2 h on ice then centrifuged at 12,000⫻gfor 10 min. One half of each sample was loaded on a Superose 12 column, equilibrated with HEPES column buffer containing 6 M urea. (C) Upon removal of urea, subdomains I and II reassociate and migrate as a single polypeptide of approximately 50 kDa. Urea was removed with an Ultrafree-4 MW30 spin filter (Millipore); the samples were immediately applied to a Superose 12 column, equilibrated with HEPES column buffer without urea. (D) Fractions of myo2-2p tail indicated by arrows in panels A, B, and C were analyzed by SDS-PAGE (15% gel). Lane 1, myo2-2p tail before trypsin digestion (peak 1 in panel A); lane 2, protein after trypsin digestion and incubation in 6 M urea; lane 3, subdomain I (peak 3 in panel B); lane 4, subdomain II (peak 4 in panel B); lane 5, after removal of urea subdomains I and II comigrate as a single polypeptide (peak 5 in panel C). The same gel was sliced and stained with Coomassie blue R-250 (lanes 1, 2, and 5) or silver stain (lanes 3 and 4) to properly visualize the different concentrations of proteins. SDI, subdomain I; SDII, subdomain II.
on September 8, 2020 by guest
http://ec.asm.org/
myo2mutants D1297G, L1301P, N1304S, or N1307D for sub-domain I and II interactions (Fig. 6B). As judged with the yeast two-hybrid test, subdomain I and II interactions were normal within the globular tails of all the suppressors and also in myo2-D1297G, myo2-N1304S, and myo2-N1307D. Of the 15 alleles tested, only one,myo2-L1301P, potentially had a defect in subdomain I and II interactions (Fig. 6B). Note, however, that the expression level of the GAL4 BD fusion of myo2-L1301p tail subdomain I was lower than the other alleles and that this mutant subdomain I had altered mobility on SDS-PAGE (Fig. 6C). This aberrant behavior makes it difficult to evaluate the apparent loss of interaction.
To further study the L1301P variant, the mutant globular tail was expressed as a GST fusion protein in bacteria. Unfortunately, the myo2-L1301Pp tail was very unstable and degraded during purification. Thus, we were not able to test subdomain I and II interactions of L1301P in an in vitro assay. We suspect that the absence of the interaction detected in the yeast two-hybrid test was due to the instability of the fusion proteins rather than a defect in binding per se. We tentatively conclude that the associ-ation of subdomains I and II is required for Myo2p function.
Subdomain I and II interactions are required for the wild-type tail to interact with Vac17p. Thus, we tested whether any of themyo2-2suppressor mutants allow subdomain I alone to interact with Vac17p. In the yeast two-hybrid test, for most of the suppressors there was no interaction between Vac17p (res-idues 1 to 170) and subdomain I (data not shown). A weak potential interaction may have occurred with a few suppressor mutants; growth was finally observed at day 11. This growth is far weaker than that of the interaction of the full globular tail with Vac17p (3 days) or wild-type subdomain I and II
ex-pressed separately interacting with Vac17p (4 days). Thus, this potential interaction is likely not an accurate representation of a biological association of subdomain I alone with Vac17p.
Subdomain I and II interaction occurs within a single poly-peptide.The interaction between subdomains I and II occurs within a single globular tail. The recombinant globular tail is a monomer as shown by equilibrium analytical ultracentrifu-gation (35). Likewise, in the yeast two-hybrid test the Myo2p globular tail does not interact with itself (2) (Fig. 7A). It is likely that the tight intratail association of the subdomains prevents dimer formation.
If the association of the two subdomains in the myo2-2, myo2-⌬AflII, ormyo2-L1301Pmutants is not as tight as in the wild type, then subdomain I from one tail could potentially interact with subdomain II from a second tail. Using combina-tions of wild-type and mutant tails frommyo2-2,myo2-⌬AflII, and myo2-L1301P, we tested for the potential interaction of two full-length globular tails (Fig. 7A and data not shown). In every case, no interaction was detected.
We also tested whether intramolecular association of the two subdomains of the above mutants may be weakened and allow subdomain I alone to interact with subdomain II within a full-length mutant tail. As was seen with wild type, none of the mutant subdomains interacted with the wild-type full-length globular tail (Fig. 7B). Moreover, the myo2-2p tail did not interact with either mutant or wild-type subdomains I or II (Fig. 7C). Similarly, no interaction was observed between the full-length tails ofmyo2-L1301Pand wild-type or mutant sub-domains I or II (data not shown). Thus, in all of the mutants tested, the globular tail is likely to be a monomer.
DISCUSSION
Myosin V molecular motors transport multiple cargoes to distinct places at different times. Therefore, there is likely to be TABLE 3. Interaction of the Myo2p tail mutants
with Vac17p, Kar9p, and Smy1p
Myo2p globular tail
Vac17p Kar9p Smy1p Two-hybrid
testd Vacuole
inheritanceb,d Two-hybrid
test
Nuclear
distributionc Two-hybrid
test
Wild-type ⫹ ⫹⫹ ⫹ ⫹ ⫹
⌬AfIII ⫹ ⫹⫹ ⫺ ND ⫺
D1297N ⫺ ⫺ ⫹ ND ⫹
L1301P ⫺ ⫺ ⫹ ND ⫹/⫺
N1304S ⫺ ⫺ ⫹ ⫹/⫺ ⫹
N1307D ⫺ ⫺ ⫹ ND ⫹
2-2(G1248D) ⫺ ⫺ ⫺ ⫺ ⫺
G1248N ⫹ ⫹⫹ ⫹ ND ⫹
V1262Aa ⫹ ⫹ ⫹ ND ⫹
F1275Sa ⫹ ⫹⫹ ⫺ ⫹/⫺ ⫹
M1279Ta ⫹ ⫹ ⫹
ND ⫺
V1288Aa ⫹ ⫹ ⫺ ⫹
/⫺ ⫺
S1247Ga ⫹ ⫹⫹ ⫺ ⫹
/⫺ ⫺
F1264Sa ⫹ ⫹⫹ ⫺ ⫹
/⫺ ⫺
S1268Pa ⫹ ⫹⫹ ⫺ ⫺ ⫺
L1193Pa ⫹ ⫹ ⫺ ⫺ ⫺
T1274Ma ⫹ ⫹ ⫺ ⫺ ⫺
a
These intragenic suppressors also contain the G1248D mutation. All sup-pressors grouped as follows: global supsup-pressors (restored all Myo2p tail functions tested), suppressors of the defect in interaction with Kar9p and Vac17p, and suppressors of the defect in interaction with Vac17p.
b
Percent vacuole inheritance is shown in Fig. 6.⫹⫹, 100 to 85% vacuole inheritance;⫹, 84 to 70% vacuole inheritance.
c
Nuclear distribution data are from Table 4. Abnormal nuclear distributions of less than 5, 10 to 20, and 20 to 30% cells are indicated as⫹,⫹/⫺, and⫺, respectively. ND, not determined.
d
Previously assessed and described in reference 20, excludingmyo2-2-L1193
andmyo2-2-T1274M.
TABLE 4. myo2-2intragenic suppressors partially restore nuclear migrationa
Strain (n)
Normal phenotypes %
Abnormal phenotypes %
Wild-type (275) 79 10 8 3
myo2-N1304S(396) 67 10 12 12
myo2-2(255) 52 8 9 30
myo2-2-F1275S(234) 70 8 10 12
myo2-2-V1288A(218) 76 6 5 12
myo2-2-S1247G(315) 67 9 10 13
myo2-2-F1264S(398) 67 11 6 15
myo2-2-S1268P(232) 63 7 9 21
myo2-2-L1193P(374) 63 7 7 23
myo2-2-T1274M(212) 60 7 10 24
aThe percentage of each cell phenotype with the indicated nuclear
distribu-tion in medium- and large-budded yeast is shown.myo2-2has a severe nuclear migration defect, while the vacuole-specific mutant,myo2-N1304S, has modest defect in nuclear migration. One group ofmyo2-2suppressors partially restores nuclear migration (10 to 20% cells with abnormal phenotypes). The other group of suppressors reveals little restoration of nuclear migration (20 to 30% cells with abnormal phenotypes). All strain genotypes are listed in Table 1.n, number of cells examined.
on September 8, 2020 by guest
http://ec.asm.org/
stringent regulation of when myosin V moves an individual cargo. This regulation could occur either via the activation or deactivation of a motor protein that is constitutively attached to its cargo and/or via the attachment or detachment of the motor protein from a specific cargo. Multiple lines of evidence indicate that this latter type of regulation occurs. For example, melanosome transport inXenopus laevismelanophores is reg-ulated by reversible association with myosin V in a cell cycle-coordinated manner (40). In addition, the discovery of cargo-specific receptor complexes (6, 20, 52, 53) and the fact that regulated degradation of the myosin V receptor directs vacuole movement in yeast (47) clearly indicate that this type of regu-latory mechanism occurs. Note that these findings do not ex-clude an additional regulatory mechanism involving the acti-vation of the motor domain.
Cargoes bind to the globular tail domain of myosin V, and it is likely that conformational changes within this domain play an active role in regulating cargo attachment (33). Further-more, phosphorylation of the myosin V globular tail results in its release from melanosomes (26); this phosphorylation may act by inducing conformational changes. In addition to a role in specifying cargo, the globular tail of myosin V may also play a role in regulating myosin motor activity. It has recently been shown that myosin Va exists in two conformations, which are
regulated by calcium and calmodulin. In the folded, inactive conformation of full-length myosin V, the globular tail inter-acts with the motor domain, while in the open, extended con-formation the motor domain is free of the globular tail and is active (27, 48). Conformational changes within the globular tail itself may be part of the regulation of motor activity.
Our recent finding that the myosin V globular tail consists of two tightly associated subdomains (35) suggests at least two possible types of conformational changes that could play a role in specifying cargo. First, the regulation of cargo attachment could occur via a reversible interaction between the two sub-domains, where some cargoes bind to the globular tail in an extended conformation, while others bind to the tail in a closed conformation. In an alternative model, the two subdomains may always be tightly associated. In this latter model, binding of cargo to one subdomain may induce a conformational change in the other subdomain that would preclude the bind-ing of additional types of cargoes. This latter model, where subdomains I and II are always tightly associated with each other, is more likely. Note that in virtually all partially func-tional alleles of Myo2p, subdomain I and II interactions were similar to those observed in wild type. Moreover, in order to bind to the globular tail, the binding proteins tested required the presence of both subdomains I and II.
FIG. 5.myo2-2intragenic suppressors simultaneously restore vacuole inheritance and Myo2p-Vac17p interactions. (A) Identification ofmyo2-2
intragenic suppressors. The starting strain,myo2-2, has a defect in vacuole inheritance as determined by the lack of carboxypeptidase Y (yellow colonies). Each suppressor restored vacuole inheritance as measured by the presence of active carboxypeptidase Y (red colonies). Suppressor mutant plasmids encoded the indicated substitution and G1248D (myo2-2) with the exception of the pseudo-revertant G1248N. The F1298L mutant (asterisk) self-activated in the yeast two-hybrid assay and was excluded from further study. (B) Schematic diagram indicating the myo2p globular tail point mutations that suppressmyo2-2. Although the region of DNA that encodes the entire globular tail was subjected to mutagenesis, note that all suppressors obtained reside in subdomain I.
on September 8, 2020 by guest
http://ec.asm.org/
If the occupation of a receptor-binding site on one main precludes the binding of receptors to the other subdo-main, then it may be possible to identify point mutations that cause a subdomain to be constitutively locked in the confor-mation that prevents the binding of a subset of cargoes. Anal-ysis ofmyo2-2(G1248D) strongly suggests that G1248D may be this type of point mutation.
Two lines of evidence support the hypothesis that themyo2-2 mutation causes a conformational change that prevents the binding of a subset of cargoes. First, while secretory vesicle movement (associated with subdomain II) is normal inmyo2-2, this mutant has multiple, severe, nonrelated defects. Themyo2-2
mutant is defective in vacuole inheritance due to an inability to interact with Vac17p, the vacuole-specific Myo2p receptor. The Vac17p binding site has been mapped to subdomain I. Furthermore, the globular tail ofmyo2-2cannot interact with Smy1p. Based on the finding that deletion of the last 117 residues of the Myo2p tail abolished Myo2p-Smy1p interac-tions, the binding site for Smy1p had been proposed to map to the C terminus (subdomain II) (2). However, similar trunca-tions also abolish the ability of Myo2p to interact with Vac17p in a yeast two-hybrid test (6, 20, 35); the primary defect in these truncations is likely due to a loss of interaction between sub-domains I and II, rather than the loss of a receptor binding site FIG. 6. Subdomains I and II interact in wild type and most partially functionalmyo2mutants. (A) As measured with the yeast two-hybrid test, subdomain I (residues 1139 to 1345) from eachmyo2-2intragenic suppressor interacts with subdomain II (residues 1346 to 1574). Each suppressor mutant plasmid encoded the indicated substitution and G1248D (myo2-2), with the exception of the pseudo-revertant G1248N. The degree of vacuole inheritance measured as percent buds with a detectable vacuole is indicated; the vacuole inheritance measurements on L1193P and M1279T were first reported here, whereas the vacuole inheritance of the other mutants has also been reported previously (20). (B) With the exception of L1301P, subdomains I and II interact in each vacuole-specific point mutant. In panels A and B, a plasmid encoding GAL4 AD fused with subdomain II was tested with the two-hybrid assay for interactions with GAL4 BD fused with subdomain I containing the indicatedmyo2point mutants. (C) The relative amounts of subdomain I-GAL4 BD fusion proteins were assessed by Western blotting by using anti-Myo2p tail antiserum. Endogenous Myo2p is shown as a loading control. Subdomain II GAL4 AD fusion proteins could not be detected by using either anti-GAL4 AD antiserum or anti-Myo2p tail antiserum (data not shown). SDI, subdomain I; SDII, subdomain II; WT, wild type.
on September 8, 2020 by guest
http://ec.asm.org/
(see below for further discussion). Thus, the binding region for Smy1p is unknown and may reside in either or both subdo-mains.myo2-2is also defective in nuclear spindle orientation due to an inability to bind to Kar9p; the binding region for Kar9p is also unknown.
In addition to the above defects, myo2-2p does not properly concentrate at sites of polarized growth (7, 8). The binding partner or molecular mechanism required for Myo2p concen-tration is unknown.
The multiple defects observed in myo2-2do not appear to result from a global instability of the mutant protein. First, in cell extracts, Myo2p and myo2-2p are present at the same steady-state levels (8). Moreover, while themyo2-2mutant has multiple defects, the ability of myo2-2p to move some cargoes is the same as that observed for the wild-type protein. This is most obvious for secretory vesicle movement, where a block would cause cell death and a partial block in movement would result in slower than normal growth. Notably,myo2-2is viable FIG. 7. Neither mutant nor wild-type globular tails interact with each other or with subdomains I or II. (A) No interactions were detected between any combination of the full-length tails of Myo2p from wild type,myo2-2, andmyo2-⌬AflII.(B) No interactions were detected between the wild-type Myo2p full-length tail and either wild-type or mutant subdomains I or II. (C) No interactions were detected between themyo2-2
full-length tail and either wild-type or mutant subdomains I or II. Plates were photographed on day 4. Control plates indicate the presence of the relevant plasmids; patches from these plates were replica plated on test media as described in Materials and Methods. SDI, subdomain I; SDII, subdomain II; WT, wild type.
on September 8, 2020 by guest
http://ec.asm.org/
over a wide range of temperatures (18 to 37°C) (8), and we were unable to identify any growth conditions where growth of themyo2-2mutant was more severely affected than growth of wild-type cells (data not shown).
In contrast to the multiple defects observed inmyo2-2, the globular tail mutations at residues D1297, L1301, N1304, or N1307 are more specifically defective in vacuole inheritance. These residues likely reside within a region of the globular tail that directly interacts with Vac17p (6, 20). Thus, themyo2-2 mutant appears to be unique among mutants identified to date that are defective in vacuole movement.
A second line of evidence suggesting that the globular tail of myo2-2may be in an altered, closed or partially nonfunctional conformation is our finding that there are reproducible differ-ences in the rate and extent of mild proteolysis of the wild-type globular tail compared with the myo2-2p globular tail.
If themyo2-2mutation causes a regulatory conformational change that simultaneously affects multiple cargoes, then it may be possible to identify second site mutations that simul-taneously restore all Myo2p-related functions. Therefore, we screened for second site mutations that specifically restore vacuole inheritance to close to wild-type levels and then sub-sequently analyzed these alleles to determine whether they also suppressed other defects associated withmyo2-2. Notably, 3 of the 10 suppressors identified solely based on their ability to restore vacuole inheritance also restored interaction with Smy1p and Kar9p (Tables 3 and 4).
To date, secretory vesicles are the only known essential cargo moved by Myo2p. While Myo2p is also involved in the inheritance of mitochondria (21, 22), an essential organelle, other mechanisms also play a role in mitochondrial movement (4). In addition, Myo2p moves vacuoles, peroxisomes, and the late Golgi; if Myo2p-based movement of these organelles to the bud is blocked, then the organelles appear in the bud by unknown, independent processes.
Notably, the Myo2p globular tail binding region for secre-tory vesicles resides within subdomain II. This raises an inter-esting possibility that the binding sites for nonessential cargoes may reside within subdomain I, while the binding site for se-cretory vesicles is localized within subdomain II. If occupation of subdomain II by the putative secretory vesicle receptor in-hibits cargo binding to subdomain I, this could provide a mech-anism for secretory vesicle movement to take precedence over the nonessential Myo2p-based movement of other cargoes. Unfortunately, it is not yet possible to test this aspect of the model. While a region on the globular tail that is part of the secretory vesicle binding site has been mapped, the identity of the secretory vesicle-specific receptor has not yet been deter-mined. This makes it impossible to design experiments that would mimic occupancy of the secretory vesicle binding site on subdomain II.
The binding sites for Vac17p, Smy1p, and Kar9p are not identical; mutations at residues D1297, L1301, N1304, or N1307 have little to no effect on the ability of Myo2p to inter-act with Smy1p or Kar9p (Tables 3 and 4). However, despite the fact that the binding sites for these proteins are not iden-tical, the binding sites for Vac17p, Kar9p, and Smy1p could potentially overlap at Myo2p residue G1248. If the binding sites overlap, then an alternative possibility is that the global suppressors restore the binding site that these proteins share in
common. However, it is not yet possible to test this hypothesis. Mapping the binding sites on Myo2p for Kar9p and Smy1p will be complex, because mutations that either interfere with sub-domain I and II interactions or directly reside in the organelle-specific receptor binding site will abolish function. Indeed, either the G1248D mutation within subdomain I or the⌬AflII (residues 1459 to 1491) deletion within subdomain II blocked the ability of the Myo2p globular tail to interact with either Kar9p or Smy1p (Table 3). Identification of the precise binding sites in Myo2p for Smy1p, Kar9p, and other as yet unknown binding partners will clarify whether these sites are structurally separated yet share an overlapping regulatory region or wheth-er the binding sites ovwheth-erlap.
While overlapping binding sites are possible, we favor the idea that the defect inmyo2-2may be due to a conformational defect in subdomain I that affects binding to several organelle-specific receptors. Determination and comparison of the three-dimensional structures of the globular tail from wild type ver-susmyo2-2may provide insights into conformational changes that contribute to the regulation and function of the globular tail.
ACKNOWLEDGMENTS
We thank R. E. Cohen, R. Piper, and S. Ramaswamy for helpful discussions and advice throughout the project, J. Duex and other members of the L.S.W. laboratory for a critical reading of the manu-script, and L. M. Tesch and Y. Li (The University of Iowa Molecular Analysis Facility) for MALDI-TOF mass spectrometry.
This work was supported by National Institutes of Health grant GM62261 (to L.S.W.) and an Established Investigator Award from the American Heart Association (to L.S.W.). N.L.C. was supported by the National Institutes of Health, National Institute on Aging grant T32 AG 00214 to the Interdisciplinary Research Training Program on Aging, University of Iowa.
REFERENCES
1.Beach, D. L., J. Thibodeaux, P. Maddox, E. Yeh, and K. Bloom.2000. The role of the proteins Kar9 and Myo2 in orienting the mitotic spindle of budding yeast. Curr. Biol.10:1497–1506.
2.Beningo, K. A., S. H. Lillie, and S. S. Brown.2000. The yeast kinesin-related protein Smy1p exerts its effects on the class V myosin Myo2p via a physical interaction. Mol. Biol. Cell11:691–702.
3.Berg, J. S., B. C. Powell, and R. E. Cheney.2001. A millennial myosin census. Mol. Biol. Cell12:780–794.
4.Boldogh, I. R., S. L. Ramcharan, H. C. Yang, and L. A. Pon.2004. A type V myosin (Myo2p) and a Rab-like G-protein (Ypt11p) are required for reten-tion of newly inherited mitochondria in yeast cells during cell division. Mol. Biol. Cell15:3994–4002.
5.Brown, J. R., P. Stafford, and G. M. Langford.2004. Short-range axonal/ dendritic transport by myosin-V: A model for vesicle delivery to the synapse. J. Neurobiol.58:175–188.
6.Catlett, N. L.2000. Role of a yeast myosin-V in movement of the vacuole and other cargoes. Ph. D. Thesis. University of Iowa, Iowa City, Iowa. 7.Catlett, N. L., J. E. Duex, F. Tang, and L. S. Weisman.2000. Two distinct
regions in a yeast myosin-V tail domain are required for the movement of different cargoes. J. Cell Biol.150:513–526.
8.Catlett, N. L., and L. S. Weisman.1998. The terminal tail region of a yeast myosin-V mediates its attachment to vacuole membranes and sites of polar-ized growth. Proc. Natl. Acad. Sci. USA95:14799–14804.
9.da Silva Bizario, J. C., A. A. da Cunha Nascimento, L. Casaletti, E. V. Patussi, M. F. Chociay, R. E. Larson, and E. M. Espreafico.2002. Expression of constructs of the neuronal isoform of myosin-Va interferes with the distribution of melanosomes and other vesicles in melanoma cells. Cell Motil. Cytoskeleton51:57–75.
10.Espreafico, E. M., D. E. Coling, V. Tsakraklides, K. Krogh, J. S. Wolenski, G. Kalinec, and B. Kachar.1998. Localization of myosin-V in the centro-some. Proc. Natl. Acad. Sci. USA95:8636–8641.
11.Estrada, P., J. Kim, J. Coleman, L. Walker, B. Dunn, P. Takizawa, P. Novick, and S. Ferro-Novick.2003. Myo4p and She3p are required for cortical ER inheritance inSaccharomyces cerevisiae. J. Cell Biol.163:1255–1266. 12.Evans, L. L., A. J. Lee, P. C. Bridgman, and M. S. Mooseker.1998.
on September 8, 2020 by guest
http://ec.asm.org/
979–990.
18.Huang, J. D., V. Mermall, M. C. Strobel, L. B. Russell, M. S. Mooseker, N. G. Copeland, and N. A. Jenkins.1998. Molecular genetic dissection of mouse unconventional myosin-VA: tail region mutations. Genetics148:1963–1972. 19.Hwang, E., J. Kusch, Y. Barral, and T. C. Huffaker.2003. Spindle orientation inSaccharomyces cerevisiaedepends on the transport of microtubule ends along polarized actin cables. J. Cell Biol.161:483–488.
20.Ishikawa, K., N. L. Catlett, J. L. Novak, F. Tang, J. J. Nau, and L. S. Weisman.2003. Identification of an organelle-specific myosin V receptor. J. Cell Biol.160:887–897.
21.Itoh, T., E. A. Toh, and Y. Matsui.2004. Mmr1p is a mitochondrial factor for Myo2p-dependent inheritance of mitochondria in the budding yeast. EMBO J.
22.Itoh, T., A. Watabe, E. A. Toh, and Y. Matsui.2002. Complex formation with Ypt11p, a rab-type small GTPase, is essential to facilitate the function of Myo2p, a class V myosin, in mitochondrial distribution inSaccharomyces cerevisiae. Mol. Cell. Biol.22:7744–7757.
23.James, P., J. Halladay, and E. A. Craig.1996. Genomic libraries and a host strain designed for highly efficient two-hybrid selection in yeast. Genetics
144:1425–1436.
24.Johnston, G. C., J. A. Prendergast, and R. A. Singer.1991. The Saccharo-myces cerevisiaeMYO2 gene encodes an essential myosin for vectorial trans-port of vesicles. J. Cell Biol.113:539–551.
25.Kaiser, C., S. Michaelis, and A. Mitchell.1994. Methods in yeast genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. 26.Karcher, R. L., J. T. Roland, F. Zappacosta, M. J. Huddleston, R. S. Annan,
S. A. Carr, and V. I. Gelfand.2001. Cell cycle regulation of myosin-V by calcium/calmodulin-dependent protein kinase II. Science293:1317–1320. 27.Krementsov, D. N., E. B. Krementsova, and K. M. Trybus.2004. Myosin V:
regulation by calcium, calmodulin, and the tail domain. J. Cell Biol.164:
877–886.
28.Langford, G. M.2002. Myosin-V, a versatile motor for short-range vesicle transport. Traffic3:859–865.
29.Liakopoulos, D., J. Kusch, S. Grava, J. Vogel, and Y. Barral.2003. Asym-metric loading of Kar9 onto spindle poles and microtubules ensures proper spindle alignment. Cell112:561–574.
30.Lillie, S. H., and S. S. Brown.1992. Suppression of a myosin defect by a kinesin-related gene. Nature356:358–361.
31.Mancini, A. J., L. S. Chan, and A. S. Paller.1998. Partial albinism with immunodeficiency: Griscelli syndrome: report of a case and review of the literature. J. Am. Acad. Dermatol.38:295–300.
32.Mercer, J. A., P. K. Seperack, M. C. Strobel, N. G. Copeland, and N. A. Jenkins.1991. Novel myosin heavy chain encoded by murine dilute coat colour locus. Nature349:709–713.
33.Miller, K. E., and M. P. Sheetz.2000. Characterization of myosin V binding to brain vesicles. J. Biol. Chem.275:2598–2606.
34.Nascimento, A. A., R. G. Amaral, J. C. Bizario, R. E. Larson, and E. M.
motors during organelle transport in melanophores. Curr. Biol.8:161–164. 40.Rogers, S. L., R. L. Karcher, J. T. Roland, A. A. Minin, W. Steffen, and V. I.
Gelfand.1999. Regulation of melanosome movement in the cell cycle by reversible association with myosin V. J. Cell Biol.146:1265–1276. 41.Rose, S. D., T. Lejen, L. Casaletti, R. E. Larson, T. D. Pene, and J. M.
Trifaro.2003. Myosins II and V in chromaffin cells: myosin V is a chromaffin vesicle molecular motor involved in secretion. J. Neurochem.85:287–298. 42.Rossanese, O. W., C. A. Reinke, B. J. Bevis, A. T. Hammond, I. B. Sears, J.
O’Connor, and B. S. Glick.2001. A role for actin, Cdc1p, and Myo2p in the inheritance of late Golgi elements inSaccharomyces cerevisiae. J. Cell Biol.
153:47–62.
43.Sandrock, T. M., J. L. O’Dell, and A. E. Adams.1997. Allele-specific sup-pression by formation of new protein-protein interactions in yeast. Genetics
147:1635–1642.
44.Schott, D., J. Ho, D. Pruyne, and A. Bretscher.1999. The COOH-terminal domain of Myo2p, a yeast myosin V, has a direct role in secretory vesicle targeting. J. Cell Biol.147:791–808.
45.Shepard, K. A., A. P. Gerber, A. Jambhekar, P. A. Takizawa, P. O. Brown, D. Herschlag, J. L. DeRisi, and R. D. Vale.2003. Widespread cytoplasmic mRNA transport in yeast: identification of 22 bud-localized transcripts using DNA microarray analysis. Proc. Natl. Acad. Sci. USA100:11429– 11434.
46.Takagishi, Y., S. Oda, S. Hayasaka, K. Dekker-Ohno, T. Shikata, M. Inouye, and H. Yamamura.1996. The dilute-lethal (dl) gene attacks a Ca2⫹store in the dendritic spine of Purkinje cells in mice. Neurosci. Lett.215:169–172.
47.Tang, F., E. J. Kauffman, J. L. Novak, J. J. Nau, N. L. Catlett, and L. S. Weisman.2003. Regulated degradation of a class V myosin receptor directs movement of the yeast vacuole. Nature422:87–92.
48.Wang, F., K. Thirumurugan, W. F. Stafford, J. A. Hammer, 3rd, P. J. Knight, and J. R. Sellers.2004. Regulated conformation of myosin V. J. Biol. Chem.
279:2333–2336.
49.Wei, Q., X. Wu, and J. A. Hammer III.1997. The predominant defect in dilute melanocytes is in melanosome distribution and not cell shape, sup-porting a role for myosin V in melanosome transport. J. Muscle Res. Cell Motil.18:517–527.
50.Weisman, L. S.2003. Yeast vacuole inheritance and dynamics. Annu. Rev. Genet.37:435–460.
51.Wu, X., B. Bowers, Q. Wei, B. Kocher, and J. A. Hammer, 3rd.1997. Myosin V associates with melanosomes in mouse melanocytes: evidence that myosin V is an organelle motor. J. Cell Sci.110:847–859.
52.Wu, X. S., K. Rao, H. Zhang, F. Wang, J. R. Sellers, L. E. Matesic, N. G. Copeland, N. A. Jenkins, and J. A. Hammer, 3rd.2002. Identification of an organelle receptor for myosin-Va. Nat. Cell Biol.4:271–278.
53.Yin, H., D. Pruyne, T. C. Huffaker, and A. Bretscher.2000. Myosin V orientates the mitotic spindle in yeast. Nature406:1013–1015.