Helgol~inder Meeresunters. 41,371-376 (1987)
Spatial and environmental c o m p o n e n t s of evolutionary
change: interactive effects of salinity and temperature
on Fucus v e s i c u l o s u s as an e x a m p l e
G.
Russell
Department of Botany, The University; Liverpool L69 3BX, England, U. K.
ABSTRACT: Intertidal algae experience aerial temperatures as well as those of ambient seawater and, during their periods Of emergence, are subject to considerable variation in salinity. The eastern Atlantic distribution of Fucus vesiculosus L. (Phaeophyta) lies within the 5 ~ and 20 ~ August isotherms. Experiments indicate that this plant can survive temperatures above and below these at normal salinity (34 %0). However, at extreme temperatures it is evid.enfly much more susceptible to saline changes than at those of the limiting isotherms. Thus the temperature limits for surviving acute saline change appear to give a better biogeographical fit than temperature alone. Neverthe- less, the presence of F. vesiculosus in estuaries at or near both geographical limits is inconsistent with the experimental results obtained from British plants. Some population divergence may therefore have occurred.
INTRODUCTION
Local p a t t e r n s of sPecies distribution are not, as a nile, of p r i m a r y c o n c e m to the algal b i o g e o g r a p h e r ; a n d the n u m e r o u s , v a r i e d factors that i n f l u e n c e local p o p u l a t i o n s are seldom discussed p r o m i n e n t l y i n b i o g e o g r a p h i c a / literature. M a r i n e b i o g e o g r a p h y is c o n c e r n e d , a b o v e all, with s p a t i a / p a t t e r n s on a global scale, a n d the chief e n v i r o n m e n t a l d e t e r m i n a n t of species hmits is u s u a l l y h e l d to b e m e a n surface s e a w a t e r t e m p e r a t u r e . This m a p - t h e r m o c l i n e a p p r o a c h has successfully d e m o n s t r a t e d the existence of major floristic p a t t e r n s a n d has, at the same time, g e n e r a t e d h y p o t h e s e s c o n c e r n i n g the t e m p e r a t u r e tolerance of species. In several cases, these h y p o t h e s e s h a v e b e e n tested experimentaUy a n d u p h e l d (see for e x a m p l e Lfining, 1984; Yarish eta/., 1984; C a m b r i d g e et el., 1984).
A separate, b u t related, activity is to d e d u c e e v o l u t i o n a r y e v e n t s from b i o g e o g r a p h i - c a / o b s e r v a t i o n s . If this a i m is i n t e n d e d , t h e n it b e c o m e s n e c e s s a r y to i n v o k e t e m p e r a t u r e histories of a n c i e n t seas a n d the processes of c o n t i n e n t a l drift. In both its spatial a n d e v o l u t i o n a r y respects, s e a w e e d b i o g e o g r a p h y has m a d e n o t e w o r t h y a d v a n c e s in r e c e n t years (Hoek, 1982a, b; Lfining, 1985).
r e l a t e m o r e to t h o s e of s e a t h a n air. It is also p e r t i n e n t to a s k if t e m p e r a t u r e o p e r a t e s s i n g l y as a n a g e n t of n a t u r a l selection, or jointly.
Fucus vesiculosus L. is p r o b a b l y t h e m o s t c o m m o n fucoid of N. A t l a n t i c r o c k y shores. O n the e a s t e r n s e a b o a r d its distribution e x t e n d s from E. G r e e n l a n d a n d N. Russia to M o r o c c o (Ppwell, 1963}. Its n o r t h e r n limit lies a little to t h e n o r t h of t h e 5 ~ A u g u s t i s o t h e r m w h i l e its s o u t h e r n b o u n d a r y a p p r o x i m a t e s to the 20 ~ A u g u s t i s o t h e r m . This s p e c i e s m a y t h e r e f o r e b e i d e n t i f i e d as a m e m b e r of t h e a m p h i a t l a n t i c t e m p e r a t e g r o u p (Hoek, 1982b). F. vesiculosus is i n t e r t i d a l a n d e m e r g e n t for m o s t of its r a n g e , b u t its a b i l i t y to live in a p e r m a n e n t l y s u b m e r g e d state is e v i d e n t from its p r e s e n c e in r o c k pools (Lubchenco, 1982} a n d from its strictly sublittoral distribution in t h e Baltic S e a (Powell, 1963). T h e g e o g r a p h i c a l r a n g e of this s p e c i e s t h e r e f o r e involves a b r o a d b a n d of s e a w a t e r t e m p e r a t u r e s w h i l e its i n t e r t i d a l h a b i t a t s e m b r a c e a v e r y c o n s i d e r a b l e v a r i e t y of e m e r - g e n t t e m p e r a t u r e conditions. C o n s e q u e n t l y , it is a p p r o p r i a t e m a t e r i a l for a c o m p a r a t i v e e x a m i n a t i o n of t h e factors t h a t m a y limit its distribution, l o c a l l y a n d g e o g r a p h i c a l l y .
MATERIALS A N D M E T H O D S
All m a t e r i a l w a s collected from t h e m i d d l e of t h e eulittoral z o n e o n H i l b r e Island, R. Dee, N. W. E n g l a n d . This is a n o u t e r e s t u a r i n e locality, w i t h o n - s h o r e s u r f a c e s e a w a t e r t e m p e r a t u r e s v a r y i n g s e a s o n a l l y from 1.5 ~ to 18.5~ Surface w a t e r salinities v a r y similarly from 27-33 o/~ (D. Thomas, pets. comm.}. Fucus p l a n t s w e r e t r a n s p o r t e d to Liverpool w i t h i n two hours of collecting a n d p l a c e d in a n a q u a r i u m c o n t a i n i n g a e r a t e d Irish S e a w a t e r (I.S.W.) o b t a i n e d from M e n a i Straits, N. W a l e s (salinity ca 34 %o). All e x p e r i m e n t s w e r e i n i t i a t e d w i t h i n 48 h of collection a n d w e r e c a r r i e d out on a p i c a l p o r t i o n s of v h g e t a t i v e thallus s e l e c t e d for a b s e n c e of d a m a g e d t i s s u e a n d v i s i b l e e p i p h y t e s .
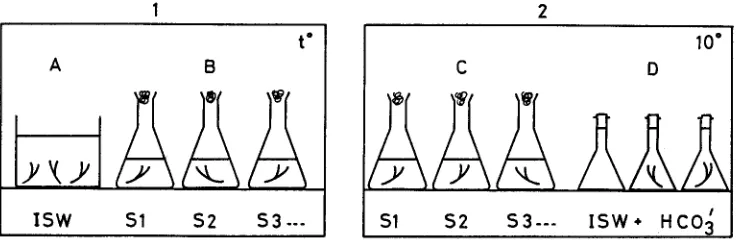
T h e m e t h o d , e x p l a i n e d d i a g r a m a t i c a l l y in F i g u r e 1, u s e d p h o t o s y n t h e t i c o x y g e n e m i s s i o n {net} as a n i n d e x of cell d a m a g e . E x c i s e d tissues w e r e first a c c l i m a t e d to t h e s e l e c t e d , e x p e r i m e n t a l t e m p e r a t u r e in I.S. w a t e r overnight. T h e p i e c e s w e r e t h e n t r a n s - f e r r e d i n d i v i d u a l l y to flasks c o n t a i n i n g w a t e r of salinity 0, 6, 11, 34, 68 or 102%o. T h e
1 2
t ~
A B
| S W $1 $ 2 $ 3 .--
10"
C D
s
$1 $ 2 5 3 - - - I S W * H C O 3
Fig. 1. Diagram showing experimental procedure for measuring temperature-sahnity damage to
Fucus tissue. Stage I involves temperature acclimation (A) in Irish Sea water (I.S.W.) before incubation for 48 h (B) in the experimental salinities (St, $2, $3. 9 .). In the second stages (2), the treatment flasks are brought to 10~ (C) before transfer to Winkler flasks (D) containing Irish Sea
v a r i o u s salinlties w e r e o b t a i n e d b y diluting I.S. w a t e r w i t h distilled water, or b y e v a p o r a t - i n g o v e r g e n t l e h e a t (80-90 ~ The flasks w e r e i n c u b a t e d for 48 h t h e n t r a n s f e r r e d to a 10 ~ r o o m until the t e m p e r a t u r e h a d once a g a i n ~tabilized (ca 2 h). T h e tissue portions w e r e t h e n r e m o v e d a n d p l a c e d in g l a s s - s t o p p e r e d b o t t l e s filled w i t h I.S. w a t e r , also at 10~ This w a t e r h a d b e e n d e g a s s e d b e f o r e h a n d b y b o i l i n g a n d then, w h e n cool, e n r i c h e d b y t h e a d d i t i o n of NaHCO3 to g i v e a final c o n c e n t r a t i o n of 5 ~ tool. T h e s e flasks w e r e i n c u b a t e d for 2.5 h u n d e r cool w h i t e f l u o r e s c e n t l i g h t at a p h o t o n f l u e n c e r a t e of 94 tool m -2 sec -1. T h e tissues w e r e t h e n r e m o v e d a n d o v e n - d r i e d b e f o r e w e i g h i n g , a n d t h e a m o u n t of d i s s o l v e d o x y g e n p r e s e n t in t h e flasks w a s d e t e r m i n e d b y s t a n d a r d W i n l d e r t e c h n i q u e . All t r e a t m e n t s w e r e a p p l i e d in triplicate a n d e a c h set of W i n l d e r bottles i n c l u d e d a trio of Fucus-free controls. T h e o n l y d e p a r t u r e from this m e t h o d o c c u r r e d in the c a s e of t h e l o w e s t t e m p e r a t u r e ( - 1 8 ~ T h e m a t e r i a l in this e x p e r i m e n t w a s k e p t o v e r n i g h t at 10~ in t h e various salinities, t h e n r e m o v e d from the w a t e r , p l a c e d s e p a - r a t e l y in l a b e l l e d p o l y t h e n e b a g s a n d t r a n s f e r r e d to the f r e e z i n g c o m p a r t m e n t for 48 h. T h e tissues w e r e t h a w e d at 10 ~ a n d the e x p e r i m e n t c o m p l e t e d as d e s c r i b e d a b o v e .
4 0 .
T.c
30,T
20.
IM 0E 10.
.
.~0
s.,.o
, ~
> , T ~ J
0 I - ' - 2 0
Fig. 2. N e t oxygen production in m g g-1 dry w e i g h t h - t p e r litre I.S.W. at 10~ in t e m p e r a t u r e - salinity stressed Fucus vesiculosus. Salinity (Too) a n d t e m p e r a t u r e (~ values are i n d i c a t e d on the
It is e x p e r i m e n t a l l y m o r e satisfactory to m e a s u r e o x y g e n o u t p u t in r e c o v e r y c o n d i - tions, as here, t h a n in the salinity t r e a t m e n t flasks. A l g a l cells t h a t e x p e r i e n c e s a l i n e s h o c k u n d e r g o c o n s i d e r a b l e c h a n g e s "in v o l u m e . T h e s e c h a n g e s m a y affect t h e p e r m e a - bility of the cell m e m b r a n e and, p a r t i c u l a r l y in the c a s e of d o w n s h o c k , r e s u l t in m i s l e a d i n g l y h i g h d i s s o l v e d o x y g e n values. T h e fact t h a t o x y g e n s o l u b i l i t y in w a t e r v a r i e s w i t h t e m p e r a t u r e a n d salinity a r g u e s also for m e a s u r e m e n t of o x y g e n o u t p u t u n d e r c o n s t a n t conditions. A n o t h e r a d v a n t a g e of this m e t h o d is t h a t it d e m a n d s a s m a l l e r n u m b e r of controls a n d is h e n c e m o r e e c o n o m i c a l in b e n c h time.
T h e p r i n c i p a l source of error is s e l f - s h a d i n g at the final s t a g e of t h e m e t h o d , if o v e r l a r g e thallus p i e c e s a r e used. As far as possible, W i n k l e r flasks s h o u l d b e e q u a l l y a n d l i g h t l y l o a d e d .
RESULTS
T h e results a r e g i v e n in F i g u r e 2, w h i c h c o m b i n e s o x y g e n output, salinity a n d t e m p e r a - ture d a t a in a t h r e e - d i m e n s i o n a i plot. T h e results i n d i c a t e t h a t at t e m p e r a t u r e s w h i c h c o r r e s p o n d w i t h t h e s u m m e r i s o t h e r m s of its g e o g r a p h i c a l limits (4~ 21 ~ Fucus vesiculosus is p r o d u c t i v e of o x y g e n o v e r a w i d e salinity r a n g e . B e y o n d t h e s e t e m - p e r a t u r e s ( - 1 8 ~ 26~ t h e r e is e v i d e n c e of d a m a g e d p h o t o s y n t h e t i c systems. In t h e c a s e of frozen tissues, p r e t r e a t m e n t in 0 Too S a n d 6 ~'oo S .was a s s o c i a t e d w i t h t h e g r e a t e s t d a m a g e . At 26 ~ h o w e v e r , i m p a i r e d p r o d u c t i o n w a s e v i d e n t at b o t h h i g h e s t a n d l o w e s t saiinities. T h e h i g h e s t t e m p e r a t u r e of all (31~ w a s lethal, i r r e s p e c t i v e of sahnity, confirming a n o b s e r v a t i o n b y S c h r a m m ( p e r s o n a l c o m m u n i c a t i o n r e p o r t e d b y G e s s n e r , 1970) t h a t t h e u p p e r t o l e r a n c e limit of s u b m e r g e d F. vesiculosus is 30 ~
D I S C U S S I O N A N D C O N C L U S I O N S
I n t e r a c t i v e effects of salinity a n d t e m p e r a t u r e u p o n m a r i n e a l g a e a r e w e l l - d o c u m e n t e d : (see Yarish & E d w a r d s , 1982, for a discussion of these, a n d for e x a m p l e s in
Caloglossa a n d Bostrychia species). F r o m t h e p r e s e n t results it is e v i d e n t t h a t Fucus
vesiculosus emits o x y g e n over a g r e a t e r r a n g e of t e m p e r a t u r e s t h a n t h o s e a s s o c i a t e d w i t h the i s o t h e r m s at its g e o g r a p h i c a l limits. W h e n t r e a t e d with s e a w a t e r of s a h n i t y 34 Too, this p l a n t s e e m s m o s t indifferent to t e m p e r a t u r e , a n d it is only at the v e r y e x t r e m e s of t h e t e m p e r a t u r e s e m p l o y e d t h a t t h e r e is e v i d e n c e of serious d a m a g e to p h o t o s y n t h e t i c systems. H o w e v e r , t h e ability to survive for 48 h t e m p e r a t u r e s o u t s i d e t h o s e of t h e limiting i s o t h e r m s is p l a i n l y i m p a i r e d w h e n salinities also b e c o m e e x t r e m e . A t 26 ~ n e t p r o d u c t i o n is m a r k e d l y r e d u c e d in tissues w h i c h h a d e x p e r i e n c e d h i g h e s t a n d l o w e s t salinities. F r e e z i n g w a s l e t h a l to tissues p r e t r e a t e d w i t h the l o w e s t b u t n o t t h e h i g h e s t saiinities. This o b s e r v a t i o n is c o n s i s t e n t w i t h t h e r e p o r t in G e s s n e r (1970) t h a t f r e e z i n g r e s i s t a n c e in Porphyra spp. is i n c r e a s e d b y prior r e m o v a l of w a t e r (cf. also Bird & McLachlan, 1974, for i n f o r m a t i o n on l o w e r t e m p e r a t u r e limits of Fucus spp.). P e r h a p s t h e p r e s e n c e of o r g a n i c solutes, m o b i l i z e d in r e s p o n s e to h i g h salinities, h a s also c o n f e r r e d s o m e p r o t e c t i o n a g a i n s t f r e e z i n g d a m a g e (cf. S t e p o n k u s , 1984, for role of s o l u t e s in cold h a r d i n e s s in plants).
t u r e t o l e r a n c e c o n d u c t e d at o p t i m a l salinity, a n d t h o s e on salinity t o l e r a n c e c o n d u c t e d at o p t i m a l t e m p e r a t u r e , m a y give results t h a t a r e difficult to i n t e r p r e t e c o l o g i c a l l y or b i o g e o g r a p h i c a l l y . S o m e of t h e s e p r o b l e m s may, of course, b e artifacts, p r o d u c t s of t h e s h o r t - t e r m n a t u r e of the e x p e r i m e n t s . Liirdng (1984) h a s d r a w n a t t e n t i o n to d i s c r e p a n c i e s b e t w e e n his o w n o b s e r v a t i o n s on t h e u p p e r t e m p e r a t u r e limits of c e r t a i n m e m b e r s of the D e l e s s e r i a c e a e , a n d those o b t a i n e d b y Biebl (1958). Biebl's e x p e r i m e n t s w e r e c o n d u c t e d o v e r a 12 h p e r i o d w h i l e those of Ltining w e r e m a i n t a i n e d for 7 days, w h i c h m a y e x p l a i n the h i g h e r t o l e r a n c e s r e p o r t e d b y the former, a l t h o u g h differences in m e t h o d m a y also h a v e b e e n involved. T h e p r e s e n t a u t h o r s h a r e s t h e e x p e r i e n c e of Ltining (1984) in f i n d i n g c o n s i d e r a b l e m i c r o b i a l d e v e l o p m e n t in h i g h - t e m p e r a t u r e t r e a t m e n t s , w h i c h c o u l d b e v i s i b l e to the n a k e d e y e e v e n after as short a time as 24 h. T h e 48 h t r e a t m e n t p e r i o d a d o p t e d h e r e h e s b e t w e e n t h o s e of Biebl a n d Lfining, a n d the m e t h o d of m e a s u r i n g t o l e r a n c e r e s e m b l e s the letter's. T h e r e is a l w a y s a p r o b l e m in e x t r a p o l a t i n g from the results of s h o r t - t e r m l a b o r a t o r y e x p e r i m e n t s to t h e u n c o n t r o l l e d conditions e x p e r i e n c e d b y p l a n t s in nature. N e v e r t h e l e s s , t h e s e e x p e r i m e n t s e n a b l e us to i d e n t i f y optimal, s u b o p t i m a l a n d l e t h a l conditions w h i c h w e m a y c o m p a r e w i t h t h o s e o b t a i n i n g o v e r the g e o g r a p h i c a l r a n g e of a species. In this case, t h e results are in a c c o r d a n c e w i t h the t e m p e r a t u r e e x t r e m e s e x p e r i e n c e d b y F u c u s vesiculosus at its g e o g r a p h i c a l limits.
H o w e v e r , results m a y b e e q u a l l y m i s l e a d i n g if, as in this case, t h e e x p e r i m e n t s are b a s e d u p o n a single p l a n t p o p u l a t i o n . T h e H.fibre p l a n t s u s e d in this e x e r c i s e w o u l d b e l i k e l y to s u s t a i n d a m a g e if e x p o s e d first to f r e s h w a t e r a n d t h e n to s u b z e r o t e m p e r a t u r e s . S u c h conditions m i g h t o b t a i n in a stratified e s t u a r y w h e n a falling t i d e l e a v e s p l a n t s e x p o s e d to f r e e z i n g air t e m p e r a t u r e s . It w o u l d b e t e m p t i n g to c o n c l u d e t h a t F. ves- iculosus at its n o r t h e r n hmit m u s t t h e r e f o r e b e a b s e n t from e s t u a r i n e habitats. It w o u l d also b e w r o n g ; for, in I c e l a n d at least, this p l a n t is to b e found in i n n e r e s t u a r i e s (Munda, 1972).
A t its s o u t h e r n limit, F. vesiculosus is f o u n d only in e s t u a r i e s (Gayral, 1958). T h e r e a s o n s for its a b s e n c e from the o p e n coast of M o r o c c o are not k n o w n , a n d c o u l d b e various, b u t it w o u l d h a v e b e e n difficult to d e d u c e its o c c u r r e n c e in e s t u a r i e s from the H i l b r e results, e s p e c i a l l y as M o r o c c a n e s t u a r i e s are l i a b l e to b e h y p e r s a l i n e as m u c h as h y p o s a h n e , a n d w i t h w a t e r t e m p e r a t u r e s t h a t m a y e x c e e d 25~ (D. H o c k i n , pers.
comm.).
T h e g e o g r a p h i c a l r a n g e of F. vesiculosus m a y thus t a k e t h e form of p o p u l a t i o n mosaic, a n d e x p e r i m e n t a l e v i d e n c e d r a w n from one p o p u l a t i o n m a y b e i n a p p l i c a b l e to others. G i v e n the v a r i a b i l i t y a n d versatility of this species, t h e r e is a s t r o n g p o s s i b i h t y that e c o t y p e e v o l u t i o n h a s occurred. So, to conclude, if t h e p h y c o l o g i s t w i s h e s to k n o w a b o u t a s p e c i e s at its g e o g r a p h i c a l (or ecological) limits, t h e n the b e s t a d v i c e is to go t h e r e .
LITERATURE CITED
Biebl, R., 1958. Temperatur- und osmotische Resistenz von h4eeresalgen der bretonischen K~iste. - Protoplasma 50, 217-242.
Bird, C. J. & IvicLachlan, J., 1974. Cold hardiness of zygotes and embryos of Fucus (Phaeophyceae, Fucales). - Phycologia 13, 215-225.
Gayral, P., 1958. Algues de la c6te aflantique marocaine. Soc. Sci. Nat. Phys. Maroc, Rabat, 523 pp. Gessner, F., 1970. Temperature: Plants. In: M a r i n e ecology. Ed. b y O. Kinne. Wfley-Interscience,
London, 1 (1), 363--406.
Hoek, C. v a n den, 1982a. Phytogeographic distribution groups of b e n t h i c m a r i n e a l g a e in the North Atlantic Ocean. - Helgol~inder M e e r e s u n t e r s . 35, 152-214.
Hoek, C. v a n den, 1982b. The distribution of b e n t h i c marine algae in relation to t h e t e m p e r a t u r e regulation of their life histories. - Biol. J. Linn. Soc. 18, 81-144.
Lubchenco, J., 1982. Effects of grazers a n d algal competitors on fucoid colonisation in tide pools. - J. Phycol. 18, 544-550.
Liining, K., 1984. T e m p e r a t u r e tolerance a n d b i o g e o g r a p h y of seaweeds: The m a r i n e algal flora of H e l g o l a n d (North Sea) as a n example. - Helgol~inder Meeresunters. 38, 305-317.
Ltining, K., 1985. Meeresbotanik. Thieme, Stuttgart, 375 pp.
h4unda, I., 1972. G e n e r a l features of the b e n t h i c algal zonation a r o u n d the Icelandic coast. - Acta nat. islandica 2I, 1-36.
Powell, H. T., 1963. Speciation in the g e n u s Fucus L. a n d related genera. In: Speciation in the sea. Ed. b y J. P. H a r d i n g & N. Tebble. System. Ass., London, 63-77.
Steponkus, P. L., 1984. Role of the p l a s m a m e m b r a n e in freezing, injury, a n d cold acclimation. - A. Rev. P1. Physiol. 35, 543-584.
Yarish, C., Breeman, A. M. & Hoek, C. v a n den, 1984. Temperature, light a n d p h o t o p e r i o d r e s p o n s e s of some N o r t h e a s t A m e r i c a n a n d West E u r o p e a n e n d e m i c rhodophytes in relation to their g e o g r a p h i c distribution. - Helgol~inder Meeresunters. 38, 273-304.