OF STEM CUTTINGS OF DALBERGIA SISSOO AND THEIR RELATIONSHIP WITH BIOCHEMICAL
CHANGES
ADARSH BALA, V. K. ANAND and K. K. NANDA
Botany Department, Panjab University, Chandigarh-14
SUMMARY
Studies were made of the rooting response of stem cuttings taken from seedlings and trees of Dalbergia sissoo in August, October and December and treated with lOOmgjl each of IAA and IBA. The changes in the activity of hydrolysing enzymes and the contents of starch, soluble carbohydrates and total and protein nitrogen were also investigated at periodic intervals.
It was found that seasonal rooting response of stem cuttings was related to the disappearance of starch. Thus, low rooting of cuttings in October corresponded with high and profuse rooting in August with low content of starch. The hydrolytic activity was high in August and October when rooting occurred but was not detected in December when cuttings did not root. The content of soluble carbohydrates increased within 2 days indicating hydrolysis of starch, and decreased subsequently indicating utilization of sugars so produced in the initiation and development of roots.
The content of total and soluble nitrogen increased earlier showing degradation Qf proteins and decreased subsequently showing utilization of soluble nitrogen during rooting.
Auxin effects in rooting are considered to be mediated through their effect on mobilization of starch caused by enhanced activity of hydrolysing enzymes and are determined by the morpho-physiological status of branches that governs the production of endogenous auxin and inhibitors.
INTRODUCTION
SEASONAL CHANGES rN ROOTING OF STEM CUTTINGS 155
these were taken from the trees in an active season. Shapiro (1957) found that in Populus nigra roots appeared on the whole length when cuttings were taken from dormant trees but only at the base when these were taken from actively growing trees in spring. Vieitez and Pena (1968) found that the rooting activity of Salix atrocinera cuttings was rhythmical with strong and weak rooting phases alternating during the annual growth cycle. Nanda et al. (1968, 1970) reported that even the effectiveness of exogenously applied auxins varied with the season. This investigation was undertaken with a view to studying the effect of auxins on seasonal changes in the rooting response of stem cuttings of Dalbergia sissoo taken both from the tree and the seedlings and studying also some of the metabolic changes that are involved in the initiation and development of adventitious roots .
. MATERIALS AND METHODS
Stem cuttings from seedlings raised in pots and also from trees of Dalbergia sissoo were collected in August, October and December and were divided into 3 groups with 100 cuttings in each group each time. While group 1 was treated with water to serve as control, groups 2 and 3 were treated with 100 mg/1 each of IAA and IBA, respectively for 24 hrs and were planted in earthen-ware pots in a 1:1 mixture of garden soil and farmyard manure. Observations on the number of cuttings that rooted and the number and length of roots produced were recorded at weekly intervals.
Plant samples were collected at periodic intervals and determinations of starch and soluble carbohydrates were made by the methods described by Whelan (1955) and Somogyi (1945), respectively and of total and protein nitrogen by micro Kjeldahl method.
solution (6 per cent KI and 0 · 6 per cent I2 V /M) was added
to the solution and its optical density was measured with the help of Bausch and Lomb photo-colorimeter, using a yellow filter. The amount of starch hydrolysed was determined by comparing the optical density of the mixture against the standard curve prepared with known concentrations of starch. The percent hydrolytic activity was calculated by dividing the amount of starch hydrolysed by the amount taken initially and multiplying the value by 100.
RESULTS
The results are presented in Tables I-V.
Rooting response.-Table I shows that in August, tree cuttings rooted after 14 days in control as compared to 10 days with auxin treatment and the number of roots was higher on auxin-treated than on control cuttings. In October rooting occurred on only I out of 10 auxin-treated cuttings and was delayed to 28 days, the control ones not rooting at all. In December cuttings failed to root even with auxin application.
R<;>oting in seedling cuttings was earlier than on tree cuttings and occurred on 2-6 cuttings out of 10 even in October. Roots emerged after 6 days in August, after 20 days in October and not at all in December in spite of auxin application (Table I). It may be noted that the number of roots was much higher on seedling than on tree cuttings (Table I).
Starch content.-In August the starch content of tree cuttings waslowprior to planting,increased somewhat in control andiAA• treated ones after 2 days but decreased after 4 days (Table III). No starch could be detected in IBA-treated cuttings even after 2 days. In October the content before planting was higher than in August, and decreased after 2 days, the decrease being more marked in auxin-treated than in control cuttings. In December the starch content prior to planting was higher than even in October and did not decrease much with time (Table III).
The trends of changes in the starch content of seedling cuttings were more or less similar to the tree cuttings except that the content before planting was higher in August but lower than that of tree cuttings in October and December (Table II).
Table I. Auxin effects on seasonal changes zn the rooting response
of
tree and seedling cuttings of Dalbergia sissoo. First figure in each column shows the numberof
cuttings that rooted out of 10; figure within parenthesis the numberof
roots per cutting and that in italics the mean lengthof
roots in em U> Seedling cuttings Tree cuttings1:%1
>
Month of Auxin treatment Days after planting
"'
0 plantingz
0 6 20 30 6 10 14 28> t"'
0 4
-0 0 5 -0 Control (0) (0·4) -(0) (0) (0·7)-1!: >
0 0·21 -0 0 0·41
-z
August IAA, 100 mg/1 0 6 -(l 5 0 -c;) (0) (2 ·6) -(0) (1. 1) (2·0)-1:%1 "'
0 0·13
-0 0·4 0·36 -.... 0 6 -0 5 6-z
IBA, 100 mg/1 (0) (4·8) -(0) (0·8) (1·6) -::>;! 0 0·18 -0 0·27 0·39 -0 0 0 2 -0 0 0 00 o-,3
Control (0) (0) (2 ·I) -(0) (0) (0) (0)
....
z
0 0 0·31
-0 0 0 0 c;) 0 0 4 -0 0 0 1 0 October IAA, 100 mg/1 (0) (0) (3·8) -(0) (0) (0) (0·5) "1 0 0 0·38 -0 0 0 0·12"'
0 0 6 -0 0 0 I o-,3 IBA, 100 mg/1 (0) (0) (4·6) (0) (0) (0) (0·3) 1:%1 -a;: 0 0 0·32 -0 0 0 0·08 0 0 G 0 0 0 0 0 0 d Control (0) (0) (0) (0) (0) (0) (0) (0)o-,3 o-,3
0 0 0 0 0
·o
0 0 ... 0 0 0 0 0 0 0 0z
pDecember IAA, 100 mgfl (0) (0) (0) (0) (0) \0) (0) (0)
"'
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 IBA, 100 mg/1 (0) (0) (0) (0) (0) (0) (0) (0) ... 0 0 0 0 0 0 0 0Table II. Auxin effects on seasonal changes in the contents of starch (first figure), soluble carbohydrates (within parentheses) and on the
percentage activity of hydrolysing en::,ymes (in italics) in seedling cuttings
cif
Dalbergia sissoo.Days after planting Month of Auxin treatment
planting 0 2 4 6 20 30
14·0 22·5 7·5 2·7 Control (1·26) (1·43) (I· 73) (2·05)
98·4 98·7 98·9 98·4
14·0 10·5 8·5 4·8 August IAA, I 00 mg/1 (1·26) (1·26) (1·26) (1·45)
98·4 98·6 98·7 98·6
14·0 13·5 2·5 2·5 IBA, I 00 mgfl (1·26) (0·95) (1·16) (J.58)
98·4 98·8 98·7 98·7
18·0 10·8 8·2 4·8 4·2 Control ('1·30) (J.52) (1·81) (2. 05) (1·38)
97·3 91·3 90·3 93·5 95·1
18·0 11·5 6·5 4·2 2·8 October IAA, I 00 mgfl (1·30) (1·32) (1·06) (1·40) (1·38)
97·3 98·7 92·8 .99·1 96·3
18·0 7·5 5·0 6·5 3·2 IBA, 100 mg/1 (1·30) (1·01) (1·32) (I· 51) (1·56)
.97·3 99·5 99·7 98·2 .94·6
19·8 20·5 21·5 20·4 21·2 Control (0·92) (1·18) (J.35) (0·93) (1·03)
0 0 0 0 0
19·8 25·0 23·0 21·5 21·5 December IAA, I 00 mg/1 (0 ·92) (1·30) (0·91) (1·50) (1·18)
0 0 0 0 0
19·8 23·5 20·8 20·9 21·5 IBA, 100 mg/1 (0·92) (0·96) (0·98) (1·18) (1·20)
0 0 0 25·2 10·2
-~--- - - - - - - - - --~---
-till 4 and 2 days, respectively and increased subsequently. In October the trends of results were similar to those in August. In December the content was lower than in October and while in control it increased with time, in IAA- and IBA-treated cut-tings it did not change much (Table III).
The trends of results with seedling cuttings were similar to those with tree cuttings (Table II).
Hydrolytic en::,ymes.-The activity of hydrolyzing enzymes
SEASONAL CHANGES IN ROOTING OF STEMS CUTTINGS 159
Table III. Auxin effects on s~asonal changes in the contents of starch
and on the (first figure), soluble carbohydrates (within parentheses)
percentage activity
of
hydrolysing en;;,ymes (in italics) tree cuttingsof
Dalbergia sisso.zn
---~--- ---~--- ---~--- ---~--- ---~--- ---~--- ---~--- ---~--- ---~--- ---~--- - -- - -- - - . - - - - - ·
:Month of Auxin treatment----· Days after planting
planting 0 2 4 6 10 14 30
Control 4·5 8·5 2·0 2·5 2·5 1·5 (1·34) (1·43) (1·43) (1·85) (1·34) (1·26)
97·1 97·1 97·2 98·0 98·8 98·4
4·5 7·0 1·5 1·5 1·5 2·0 August IAA, 100 mg/1 (1·34) (1·38) (1·06) (1·34) (1·58) (1·38)
97·1 .97·5 .97·0 98·5 97·4 .98·6
4·5 0 0 0·5 0·5 1·5 IBA, IOC mg/1 \1·34) (1·19) (1·81) (1·38) (1·39) (1·38)
97·1 97·5 97·0 98·9 99·1 98·0
-21·5 17·5 10·5 8·0 8·0 Control (1·42) (1·50) (I-50) (2·00) (1·42)
96·5 99·0 92·5 98·4 92·0
21·5 12·0 8·0 5·5 4·8 October IAA, 100 mgjl (1·42) (1·42) (1·08) (1·38) (1·52)
96·5 .98·8 91·5 99·5 .98·2
21·5 12·5 7·5 5·8 4·1 IBA, 100 mgjl (1·42) (1·20) (1·81) (1-40) (1·58)
96·5 98·7 .98·6 99·8 .97·4
-23·9 22·5 20·1 18·0 18·0 Control (1·01) (1·17) (1·20) (1·30) (1·41)
0 0 0 0 0
23·9 20·1 21·5 20·0 20·3 December IAA, 100 mgjl (1·01) (1·03) (0·83 (0·96) (1·30)
0 0 0 0 0
23·9 22·0 22·5 16·0
IBA. 100 mgjl (1·01) (1·08) (1·18) (I· 10) (1·20)
0 0 0 0 0
to this the hydrolysing enzymes remained completely inactive in December (Tables II and III).
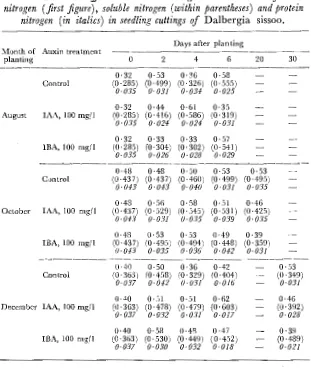
Total nitrogen.-In August the total nitrogen content of
Table IV. Auxin e.ffects on seasonal c~anges in the contents of total nitrogen (first figure), soluble nitrogen (within parentheses) and protein
nitrogen (in italics) in seedling cuttings of Dalbergia sissoo.
Days after planting Month of Auxin treatment
planting 0 2 4 6 20 30
0·32 0·53 0· 36 0·58 Control (0·285) (0 ·499) (0. 326) (0 ·555)
0·035 0·031 0·031: 0·025
0·32 0·44 0·61 0·35 August IAA, 100 mg/1 (0·285) (0·416) (0·586) (0·319)
0·035 0·021: 0·021: 0·031
0·32 0·33 0·33 0·57 lBA, 100 mgjl (0·285) (0. 304) (0·302) (0· 541)
0·035 0·026 0·028 0·029
-0·48 0·48 0·50 0·53 0·53
Control (0 ·437) (0·437) (0·460) (0. 499) (0. 495)
0. 01:3 0·01:3 0·01:0 0·031 0·035
0·48 0·56 0·58 0·51 0·46 October IAA, 100 mg/1 (0 ·437) (0·529) (0· 545) (0·531) (0·425)
0. 01:3 0·031 0·035 0·039 0·035
0·48 0·53 0·53 0·49 0·39 IBA, 100 mgjl (0·437) (0·495) (0·494) (0·448) (0·359)
0·043 0·035 0·036 0·042 0·031
-0 ·4-0 0·50 0·36 0·42 0·53
Control (0·363) (0·458) (0·329) (0·404) (0·349)
0·037 0·042 0·031 0·016 0·031
0·4.0 0·51 0·51 0·62 0·46
December IAA, 100 mg/1 (0. 363) (0·478) (0 ·479) (0·603) (0·392)
0·037 0·032 0·031 0·017 0·028
0·40 0·58 0·48 0·47 0·39
IBA, 100 mg/1 (0·363) (0. 530) (0·449) (0·452) (0 ·489)
0·037 0·030 0·032 0·018 0·021
The total nitrogen content of seedling cuttings in August was higher than that of tree cuttings. It increased after 2 days in control and IAA-treated cuttings but after 6 days in lEA-treated ones. In October the content was initially higher than in August and increased with auxin application within 2 days. While in controls the content remained high even after 20 days, it decreased in IBA-treated cuttings. In December the content in all cases increased after 2 days and was a little lower than in October (Table IV).
SEASONAL CHANGES IN ROOTING OF STEM DUTTINGS 161
Table. V. Auxin effects on seasonal changes in the contents
of
totalnitrogen (first figure), solube nitrogen (within parentheses) and
protein nitrogen (in italics) in tree cuttings
of
Dalbergia sissooMonth of Auxin treatment-- Days after planting
planting 0 2 4 6 10 14 30
0·23 0·26 0·51 0·41 0·40 0·34 Control (0·189) (0·223) (0 ·475) (0·387) (0·373) (0·317)
0·011 0·037 0·035 0·023 0·027 0·023
0·23 0·42 0·43 0·42 0·38 0·41 August IAA, 100 mg/1 (0·189) (0 ·392) (0 ·405) (0·390) (0·364) (0·385)
0·041 0·028 0·025 0·030 0·016 0·025
0·23 0·32 0·48 0·48 0·39 0·25 IBA, 100 mgfi (0·189) (0·289) (0·461) (0·449) (0·372) (0·222)
0·041 0·031 0·019 0·031 0·018 0·028
-0·41 0·43 0·45 0·49 0·36 Control (0·354) (0 ·382) (0·399) (0·457) (0·320)
0·056 0·018 0·051 0·033 0·010
0·41 0·44 0·56 0·36 0·45 October IAA, 100 mg/1 (0·354) (0. 399) (0·522) (0. 324)
(0·424)
0·056 0·011 0·038 0·036 0·026
0·41 0·39 0·51 0·45 0·57 IBA, I 00 mg/1 (0·354) (0·348) (0·471) (0·408) (0 ·549)
0·056 0·042 0·039 0·042 0·021
0·28 0·31 0·51 0·28 0·50 Control (0·254) (0·284) (0·494) (0·257) (0 ·475)
0·026 0·026 0·016 0·023 0·025
0·28 0·38 0·56 0·37 0·53 December IAA, 100 mgfl (0·254) (0·353) (0·542) (0·352) (0·507)
0·026 0·027 0·018 0·018 0·023
0·28 0·47 0·41 0·42 0·38 IBA, 100 mg/1 (0·254) (0·450) (0·393) (0·403) (0 ·352)
0·026 0·020 0·017 0·017 0·028
soluble nitrogen were similar to those of total nitrogen (Tables IV and V).
Protein nitrogen.-ln August and October the protein
con-tent of tree cuttings decreased gradually with time, the decrease being more with auxin application. In December the protein content was much lower than in October and August and decreas-ed after 4 days in all cases (Table V).
after 6 days. In December the content decreased after 6 days m all cases as compared to 4 days in tree cuttings (Table 4).
DISCUSSION
The results presented in this paper demonstrate that rooting response of stem cuttings of Dalbergia sissoo varies with the season.
Thus, rooting was profuse in August, scarce and considerably delayed in October and did not occur at all in December regard-less of whether the cuttings were taken from the seedlings or from the tree.
It may be noted that in August rooting was earlier and the number of roots was more on seedling than on tree cuttings and in October the tree cuttings did not root even with auxin applica-tion. Evidence is available in literature to show that ability of cuttings to root decreases with the age of the mother plant (Mirov, 1944; Allen and McComb, 1955; Sax, 1962 and Quamme and Nelson,1965). Beakbane (1961) and Goodin (1965) considered that decrease in rooting response with age was due to some anatomical changes in stem structure. A high degree of sclerifi-cation and the presence of pericycle and bast fibres in tree cuttings of many species has been reported from this laboratory (Singh, 1967). Gorter (1961) attributed it to a decrease in the reactivity of auxins with the age of the mother plant.
The complete inhibition of rooting in December closely corresponds with high and profuse rooting during August with low starch content, clearly indicating a close relationship with mobilization of reserve food materials. A negative correlation between starch content and seasonal changes in rooting response of stem cuttings of Populus nigra has also been reported (Nanda and
SEASONAL CHANGES IN ROOTING OF STEM CUTTINGS 163
relationship that is found to exist between the starch content of stem cuttings and the activity of hydrolysing enzymes. Thus, hydrolytic activity of plant extract was very high in August but very low in December.
The more rooting of seedling cuttings in August and the lack of rooting of tree cuttings even in October in spite of the high activity of cambial cells may be due either to the sclerification of tissue or to the decrease in reactivity of the tissue to auxins, as is stated in an earlier paragraph. The food materials, therefore, remain unutilized even though these are present in soluble form.
That the rooting response of stem cuttings is closely related to the mobilization of reserve food materials is also apparent from seasonal changes in the content of soluble carbohydrates, the trends of changes being contrary to those of starch. Thus, the content of soluble carbohydrates was low prior to planting but increased "Within 2 days, indicating mobilization of starch into soluble carbohydrates. It decreased subsequently indicating their utilization in rooting. The subsequent decrease in general, occurred earlier in IBA- than in IAA-treated or control cuttings.
The changes that occurred in the contents of total, soluble and protein nitrogen also demonstrate that the reserve food materials are involved in rooting. Thus, protein nitrogen was high earlier but decreased subsequently, clearly showing that the degradation of proteins occurred during the initiation of roots. This is supported by an increase in the contents of total and soluble nitrogen during this period. The fall in the level of
total and soluble nitrogen contents after 2-4 days was due to the utilization of soluble nitrogen in the initiation and development of roots.
through a change in the activity of hydrolysing enzymes influenc-ing the mobilization of reserve food materials.
AcKNOWLEDGEMENT
This research has been financed by a grant from the United States Department of Agriculture.
REFERENCES
Alexander, T. R. (1938). Carbohydrates of bean plant after treatment with indole-acetic acid. PL. Physiol., 13: 845-56.
Allen, R. H. and McComb, L. (1955). Uber Faktoren die Bewurzelung der Steck-linge von der Populus deltoides Bartt. Beeinfiussen Zen tralbl. Forstwesen,
74: 199-220.
Ali, 'N. and Westwood, M. N. (1966). Rooting of pear cuttings as related to car-bohydrates, nitrogen and rest period. Proc. Amer. Hart. Sci., 88: 145-50. Bausor, S. C. (1942). Effect of growth substances on reserve starch. Bot. Gaz.,
104: 115-21.
Beakbane, B. A. (1961). Structure of the plant in relation to adventitious rooting.
Nature, Lond., 192: 954-55.
Beal, J. M. and Whiting, A. G. (1945). The effect of indole-acetic acid in inhibiting stem abscission in Mirabilis jalapa. Bot. Gaz. 106: 420-31.
Borthwick, H. A., Hamner, K.C. and Parker, M. W. (1937). Histological and micro-chemical studies of the reaction of tomato plants to indole-acetic acid.
Bot. Gaz., 98: 491-519.
Doak, B. W. (1939). Amino-acids and rooting of cuttings. Nature, Land., 44: 379.
Goodin, J. R. ( 1965). Anatomical changes associated with Juvenile-to-mature growth transition in Hedera. Nature (Land.) 208: 504--05.
Gorter, C. J. (1961). Morphogenetic effects of synthetic auxins. Encycl. Pl. Physiol., XIV: 1085-95.
Hartmann, H. T. and Loreti, F. (1965). Seasonal variation in rooting leafy olive cuttings under mist. Proc. Amer. Soc. Hart. Sci., 87: 194.
Hess, C. E. (1969). A physiological comparison of rooting in easy-to-root and difficult-to-root cuttings. Proc. Pl. Proc. Soc., 13: 51-7.
Hess, C. E. (1968). Internal and external factors regulating root initiation. Proc.
XV Easter School in Agricultural Science University of Nottingham, 42. Jusufov, A. G., Tylik, L. N. and Ahlakar, R. (1965). Some anatomical and
physiological changes in cuttings during rooting. Fiziol. Rast., 12: 732-35. Marcavillaca, M. C. and Montaldi, E. R. (1963). Rooting of Eucalyptus rostrata. FAO World Consult. For Genet., Stockholm No. FAO/FORGEN 63: 518. - - - , and - - - , E. R. (1964). Rooting of Eucalyptus rostrata. ldia.
Suppl. No. 12: 65-72.
Morov, N. T. (1944). Experiments in rooting pines in California. Jour. Forest.,
42: 807-08.
Nanda, K. K. and Anand, V. K. (1970). Seasonal changes in auxin effects on rooting of stem cuttings of Populus nigra and its relationship with mobilization of starch. Ph_ysiol. Plant., 23: 99-107.
---,Purohit, A. N., BaJa, A. and Anand, V. K. (1968). Seasonal rooting response of stem cuttings of some forest tree species to auxins. Indian
Forester, 94: 154--62.
Pearse H. L. (1943). The effect of nutrition and phytohormones on the rooting of vine cuttings. Ann. Bot. N.S. 7: 123-32.
Quamme, H. A. and Nelson, S. H. (1965). Root promoting substances for Juvenile phase of Malus robusta. Canad. Jour. Pl. Sci., 45: 509-11.
Sax, K. (1962). Aspects of ageing in plants. A. Rev. Pl. Physiol., 13: 489-507. Shapiro, S. (1967). The role oflight in the growth of root primordia in the stem
t
SEASONAL CHANGES lN ROOTING OF STEM CUTTINGS 165
Singh, K. (1967). Effect of age and auxins on seasonal rooting response of cuttings of
Dalbergia sissoo Roxb. Thesis submitted to Pan jab University for the degree ofM.Sc.
Stuart, N. W (1938). Nitrogen and Carbohydrate metabolism of kidney bean cuttings as affected by treatment with indole-acetic acid. Bot. Gaz.,
100: 298-311.
Somogyi, M. (1945). A new reagent for the determination of sugars. J. Biol. Chem.,
160:61-68.
Vieitez, E., Scone, E., Gesto, D. V., Mato, C., Vazquez, A. and Curnicer, A. (1964). First step in the isolation of plant hormones associated with rooting capacity of wood cuttings. AnEdafol. Agrabiol., 23: 777.
- - - a n d Pena, J. (1968). Seasonal rhythm of Salix atrocinera cuttings. Physiol.
Pl., 21: 544-55.
Wareing, P. F., El-Antably, H. M. M., Good,]. and Manuel,]. (1967). The possible role and mode of action of abscisin (Dormin) in the regulation of plant growth and development. Wissenschaftliche Zeitschrift Der Universitat Rostock-!6 Jahrgang, 4(5: 667.
- - - and Smith, N. G. (1963). Physiological studies on the rooting of cuttings. Extr. from Rept. Res. For. Com. London.
Waxman, S. Cl962). The physiology of an evergreen cutting from the time it is taken until the time it is rooted. Proc. Int. Pl. Prop. Soc., 27: 55-6].