Macroinvertebrate communities structure in different environments
of the Taim Hydrological System in the state of Rio Grande do Sul,
Brazil.
WÜRDIG1, N. L. , CENZANO1, C. S. S. & MOTTA MARQUES2, D .
1 Universidade Federal do Rio Grande do Sul – UFRGS, Instituto de Biociências, Dep. de Zoologia,
Lab. de Invertebrados Bentônicos. Av. Bento Gonçalves, n° 9.500, CEP: 91501970 – Porto Alegre – RS, Bra s i l . w ü r d i g n l @ u f r g s . b r ; c s s c e n z a n o @ y a h o o . c o m . b r
2 Universidade Federal do Rio Grande do Sul – UFRGS, Instituto de Pesquisas Hidráulicas – IPH.
Av. Bento Gonçalves, n° 9.500, CEP: 91501970 – Porto Alegre – RS, Brasil. [email protected]
ABSTRACT: Macroinvertebrate communities structure in different environments of the Taim Hydrological System in the state of Rio Grande do Sul, Brazil. The structure and functional feeding groups of the benthic macroinvertebrate communities, from wetlands and freshwater lakes within the Taim Hydrological System, in the southern coastal zone of the state of Rio Grande do Sul - Brazil, were studied. Macroinvertebrates were collected during three sampling campaigns in summer of 2000 and winter and autumn of 2001, at seven sampling points in lakes Flores, Nicola and Jacaré and in North, Central and South areas in Mangueira Lake and at Mangueira Lake/ Taim wetland interface. The taxa composition and the community structure showed a large diversity of habitats in the system. The ANOVA test indicated that mean densities of invertebrate community and family richness was significantly higher in north a n d s o u t h M a n g u e i r a L a k e s a m p l i n g s t a t i o n s . T h e c l u s t e r a n a l y s i s , b a s e d o n a v e r a g e density values (ind/m2) and macroinvertebrate composition, defined 4 groups. G r o u p 1 ,
with samples from the Central Mangueira station, which distinguished itself from the other groups by the predominance of Cumacea and Oligochaeta. Group 2 was formed by sampling stations from Taim Wetlands’ inner lakes, Nicola and Jacaré, presenting lower average individual densities and family richness. Group 3 joined the Lake Mangueira/Taim Wetland interface and the North Lake Mangueira sampling stations, which are characterized by the presence of Tanaidae and Corophiidae. Group 4 was formed by Flores Lake and South Mangueira Lake sampling stations, which were characterized by a greater abundance of Hidrobiidae. Functional groups composition of benthic macroinvertebrates was characterized b y a p r e d o m i n a n c e o f g a t h e r i n g c o l l e c t o r s i n t h e s a m p l e s t a t i o n s t h a t c o m p o s e d t h e groups 1, 2 and 3 and by a predominance of scrapers in the points that formed the group 4.
Key-words: m a c r o i n v e r t e b r a t e s , s t r u c t u r e , f u n c t i o n a l f e e d i n g g r o u p s , w e t l a n d s , T a i m Hydrological System, South Brazil.
RESUMO: Estrutura das comunidades de macroinvertebrados em diferentes ambientes do Sistema Hidrológico do Taim, Rio Grande do Sul, Brasil. O presente trabalho estuda a estrutura e os grupos tróficos funcionais de macroinvertebrados bentônicos de um conjunto de lagos na área do Sistema Hidrológico do Taim, na parte sul da região costeira do estado do Rio Grande do Sul – Brasil. Os macroinvertebrados foram coletados durante três campanhas no verão de 2000 e no inverno e outono de 2001, em uma estação amostral nos lagos: Flores, Nicola e Jacaré, nas áreas norte, central e sul da Lagoa Mangueira e na interface da Lagoa Manguei-ra com o subsistema do Banhado do Taim. A composição dos táxons e a estrutuManguei-ra das comunidades mostraram diferenças entre as estações amostrais do sistema. Os resulta-d o s resulta-d o t e s t e A N O VA i n resulta-d i c a r a m q u e a s resulta-d e n s i resulta-d a resulta-d e s m é resulta-d i a s resulta-d a c o m u n i resulta-d a resulta-d e resulta-d e macroinvertebrados e a riqueza de famílias foram significativamente altas nas partes norte e sul da Lagoa Mangueira. A análise de agrupamento baseada nas médias de densidade (ind/m2) e composição de macroinvertebrados definiram 4 grupos. Grupo 1, constituído
pela estação amostral Mangueira centro, se distinguiu dos demais grupos pela predomi-nância de Cumacea e Oligochaeta. Grupo 2, formado pelas estações dos lagos internos do Banhado do Taim, Nicola e Jacaré, que apresentaram baixas densidades médias de indiví-duos e baixa riqueza de famílias. Grupo 3, reuniu a estação de interface Lagoa Mangueira/ Banhado do Taim e a da Lagoa Mangueira norte, que foram caracterizadas pela presença de Tanaidae e Corophiidae. Grupo 4, formado pelas estações amostrais do Lagoa Flores e
da parte sul da Lagoa Mangueira, as quais foram caracterizadas por uma grande abundân-cia de Hidrobiidae. A composição dos grupos funcionais de macroinvertebrados bentônicos foi caracterizada pela predominância de coletores de depósito nas estações amostrais que compuseram os grupos 1, 2 e 3 e pela predominância de raspadores nas estações que formaram o grupo 4.
Palavras-chave: macroinvertebrados, estrutura, grupos tróficos funcionais, terras úmidas, Sis-tema Hidrológico do Taim, sul do Brasil.
Introduction
B e n t h i c m a c r o i n v e r t e b r a t e s a r e a n i m p o r t a n t c o m p o n e n t o f a q u a t i c environments taking part in ecosystem pro-cesses. They play an essential role in the f o o d c h a i n , p r o d u c t i v i t y , n u t r i e n t c y c l i n g , a n d d e c o m p o s i t i o n . L i v i n g o n a n d i n t h e sediments or associated with macrophytes, t h e b e n t h o n i c c o m m u n i t i e s c a n e x h i b i t d i f f e r e n t c o m p o s i t i o n a n d f u n c t i o n a l s t r u c t u r e a s a n s w e r t o b i o l o g i c a l a n d p h y s i c o c h e m i c a l v a r i a b l e s a t m u l t i p l e spacial scales. M a c r o i n v e r t e b r a t e s a r e d i r e c t l y i n f l u e n c e d b y s u b s t r a t e t y p e , a v e r a g e sediment grain size, benthic organic matter abundance, macrophytic bed composition a n d s t r u c t u r e , w a t e r n u t r i e n t s , d i s s o l v e d o x y g e n l e v e l , a n d d e p t h ( D o w n i n g , 1 9 9 1 ; Szalay & Resh, 1996; Würdig et al., 1998, Shieh et al., 1999 and Stewart et al., 2000). Because the environmental heterogeneity, these variables can change in the distincts areas of lakes, wetlands and streams and determining changes in the structure and distritution of macrobenthos. M o s t n a t u r a l e c o s y s t e m s e x h i b i t e x t r e m e h e t e r o g e n e i t y i n e n v i r o n m e n t a l c o n d i t i o n s a n d b i o t i c c o m m u n i t i e s a t m u l t i p l e s p a c i a l s c a l e s , r a n g i n g f r o m m i c r o h a b i t a t s t o w h o l e l a n d s c a p e s a n d ecoregions (Heino et al., 2004). Knowledge of the interplay between ecological scale a n d a q u a t i c c o m m u n i t y c o m p o s i t i o n a n d function are needed to detect and interpret m o r e e f f e c t i v e l y c h a n g e s i n b i o d i v e r s i t y , thus improving our awareness of ecosystem response, resistance and resilience to na-t u r a l a n d h u m a n - i n d u c e d p e r na-t u r b a na-t i o n (Johnson et al., 2004).
T h e p r e s e n t s t u d y i s p a r t o f t h e B r a z i l i a n P r o g r a m f o r a L o n g – Term Ecological Research (MCT/CNPq), site 7. The objective of this study is to investigate the invertebrate macrofauna composition and s t r u c t u r e a l o n g t h e T a i m H y d r o l o g i c a l System, verifying the distribution patterns o f m a c r o i n v e r t e b r a t e f u n c t i o n a l f e e d i n g
groups in the sampled areas, comparing the l a c u s t r i n e e c o s y s t e m s a n d t r a n s i c i o n a l environments in the Taim wetland.
Study area
The Taim Hydrological System with an area of 2,254km² is located in the southern coastal zone of Rio Grande do Sul – Brazil, a n d m a k e s a c h a i n o f a q u a t i c a r e a s c h a r a c t e r i z e d b y w e t l a n d s a n d s h a l l o w freshwater lakes (Motta Marques et al., 2002). It represents an area with a great diversity of habitats, where in the northern part Flo-res Lake and the Taim Wetlands are covered w i t h e x t e n s i v e m a c r o p h y t i c b a n k s . L a k e s N i c o l a a n d J a c a r é a r e s m a l l f r e s h w a t e r bodies associated with the Taim Wetlands. Mangueira Lake makes up a vast extension of free waters located in the southern region of the Taim Wetlands. The northern part of this lake establishes an important interface w i t h t h e w e t l a n d s , i n t e r a c t i n g i n t h e hydrodynamic processes of the system.
Villanueva et al. (1998) recognize three subsystems: the North Subsystem, formed b y L a k e C a i u b á , L a k e F l o r e s , a n d t h e M a ç a r i c o W e t l a n d , w h i c h i s L a k e F l o r e s influent; the Wetland Subsystem, made up of Nicola Lake, Jacaré Lake, and the Taim Wetlands; and the South Subsystem, formed b y M a n g u e i r a L a k e a n d i t s c o n t r i b u t i n g basin. The South and Wetland Subsystems communicate by the lake-wetland interface through diffuse flow and by a channel along t h e f e d e r a l r o u t e , B R 4 7 1 . F r o m a hydrological perspective, North Subsystem’s i n f l u e n c e o n t h e W e t l a n d S u b s y s t e m i s m i n i m a l . W a t e r f l o w i n t h e w e t l a n d i s characterized by low surface velocities due t o t h e p r e s e n c e o f a g r e a t q u a n t i t y o f macrophytes. Lakes Flores, Nicola, and Ja-c a r é a r e Ja-c h a r a Ja-c t e r i z e d b y s m a l l s u r f a Ja-c e areas corresponding to 11.30km², 2.58km², and 1.45km², respectively. Average depths a r e a r o u n d 1 . 6 8 m , 1 . 1 0 m a n d 1 . 5 7 m , respectively. Lake Mangueira has a surface area of 802 km² and an average depth of 2.49m (Fig. 1).
F i g u r e 1 : Study area, sampling points location, in the Taim Hydrological System, southern coast of Rio Grande do Sul, Brasil. FL= Lake Flores, NI= Lake Nicola, JA= Lake Jacaré, TB= Lake Mangueira-Banhado Taim, TN= Lake Mangueira-north, TC= Lake Mangueira-center, TS= Lake Mangueira-south.
Material and methods
Data collection
B e n t h i c m a c r o f a u n a s a m p l i n g w a s c a r r i e d o u t i n t h e s u m m e r o f 2 0 0 0 (December), autumn 2001 (March), and winter 2001 (June), at 7 sampling points in Lakes Flores (FL), Nicola (NI), Jacaré (JA), North, Central, and South Mangueira, (TN, TC, and T S ) , a n d a t t h e M a n g u e i r a L a k e / T a i m Wetland interface (TB), as displayed in Fi-gure 1. At each area, three samples were collected for the macroinvertebrates using a n E k m a n B o t t o m G r a b ( w i l d c o , 6 ’ x 6 ' , s t a n d a r d ) . S a m p l e s w e r e f i x e d i n a 1 0 % formaldehyde solution buffered with sodium b o r a t e . T h e n t h e y w e r e r i n s e d t h r o u g h a 0.250mm mesh sieve and macrofauna was p r e s e r v e d i n 7 0 % a l c o h o l . S o r t i n g a n d s e p a r a t i o n w e r e c a r r i e d o u t u n d e r a s t e r e o m i c r o s c o p e . T h e o r g a n i s m s w e r e identified at family level and when possible some groups were identified in tribes, genus or species or even morphotypes by means of taxonomic keys: Brinkhurst & Marchese (1989), Epler (2001), McCafferty (1981) and Trivinho-Strixino & Strixino (1995).
D u r i n g t h e m a c r o f a u n a s a m p l i n g , simultaneous water samples were collected using a Van Dorn bottle. Water samples field h a n d l i n g a n d l a b o r a t o r y m e a s u r e s o f s u s p e n d e d s o l i d s , c h l o r i d e s , c h e m i c a l oxygen demand and chlorophyll followed A S P H A ( 1 9 9 5 ) . M e a s u r e m e n t s o f t o t a l n i t r o g e n a n d t o t a l p h o s p h o r o u s f o l l o w e d Mackereth et al. (1978).
W a t e r c o l u m n d e p t h a n d w a t e r transparency were measured using a depth sensor and a Secchi disc, respectively. The w a t e r p H , t e m p e r a t u r e , s a l i n i t y , a n d d i s s o l v e d o x y g e n w e r e m e a s u r e d i n t h e field using a YSI 6600 multiprobe.
Data analyses
I n d i v i d u a l s ’ a v e r a g e d e n s i t y ( i n d / m ² ) and relative abundance (%) was carried out based on a bottom-grab area of 332 cm2.
Richness (S), diversity (H’) and equitability (J) were calculated for the number of families present in the sampling areas, employing the statistics program PRIMER version 5.2.9. ( C l a r k e & W a r w i c k , 2 0 0 1 ) . T h e S h a n n o n -Weaver (H’) diversity index is calculed with n a t u r a l l o g a r i t h m ( n l ) , a n d e m p l o y i n g t h e statistics program PRIMER version 5.2.9.
Variance analysis (ANOVA) was applied between the sample points and seasons,
ta k i n g i n t o c o n s i d e r a t i o n t h e m a c r o f a u n a m e a n d e n s i t y t r a n s f o r m e d b y l o g ( x + 1 ) , R i c h n e s s f a m i l y ( S ) , S h a n n o n - W i e n e r diversity (H’) and Pielou equitability (J). The t e s t o f S c h e f f é w a s a p p l i e d w h e n s i g n i f i c a n t d i f f e r e n c e s w e r e d e t e c t e d ( p < 0 . 0 5 ) , b y S TAT I S T I C ® p r o g r a m 5 . 0 v e r s i o n . S t a t i s t i c a l t r e a t m e n t w a s c a r r i e d o u t u s i n g t h e a v e r a g e i n d i v i d u a l s d e n s i t y ( i n d / m2) u n d e r g o i n g a l o g a r i t h m i c
transformation. The Bray-Curtis dissimilarity m e a s u r e w a s c h o s e n f o r d e t e r m i n i n g t h e p r e s e n c e o r a b s e n c e o f f a m i l i e s i n t h e s t u d y a r e a , a s w e l l a s t h e i r q u a n t i t a t i v e e x t e n s i o n ( L e g e n d r e & L e g e n d r e , 1 9 9 8 ) . The hierarchical grouping analysis through t h e U n w e i g h t e d P a i r G r o u p M e t h o d w i t h Arithmetic Mean (UPGMA) was carried out for the distribution analysis of the benthos macrofauna, with the help of the statistical p r o g r a m P R I M E R v e r s i o n 5 . 2 . 9 . ( C l a r k e & Warwick, 2001). From the groups obtained in the Cluster analysis were calculated the p e r c e n t u a l o f f u n c t i o n a l f e e d i n g g r o u p s o f t h e c o m m u n i t y o f b e n t h i c m a c r o i n v e r t e b r a t e s .
Results
Environmental variables
P h y s i c a l a n d c h e m i c a l p a r a m e t e r s were recorded at the 7 sampling points in t h e T a i m H y d r o l o g i c a l S y s t e m d u r i n g autumn 2001(March) and winter 2001(June). I n s u m m e r o f 2 0 0 0 o n l y b i o t i c d a t a w a s collected. Temperatures during the autumn f l u c t u a t e d a r o u n d 2 3 ° C , d r o p p i n g t o 1 2 ° C i n w i n t e r. D i s s o l v e d o x y g e n v a l u e s r e m a i n e d h i g h , w i t h s a t u r a t i o n o v e r 8 0 % a t a l l c o l l e c t i o n p o i n t s a n d p H r e m a i n e d a b o v e 7 . I n a u t u m n , c o n d u c t i v i t y r e g i s t e r e d t h e h i g h e r v a l u e s , w h e r e a s s u s p e n d e d s o l i d s , c h l o r i d e s , a n d c h l o r o p h y l l a w e r e h i g h e r i n w i n t e r throughout the entire system (Tab. I). Wind direction in the region was predominantly S S E i n a u t u m n a n d E N E i n w i n t e r . T h e w a t e r c o l u m n w a s h i g h e r i n w i n t e r 2 0 0 1 , w i t h a v e r a g e v a l u e s a r o u n d 2 . 5 m w h i l e in the autumn was around 2m. According Vo l k m e r - R i b e i r o e t a l . ( 2 0 0 6 ) s a m p l i n g stations sediment varied from mud to sand (Tab. II).The macrofauna structure
T h e b e n t h i c m a c r o f a u n a i n t h e T a i m H y d r o l o g i c a l S y s t e m i s r e p r e s e n t e d b y
f o r t y t w o t a x o n o m i c g r o u p s , b e i n g C h i r o n o m i d a e t h e m o r e r i c h i n s p e c i e s (Tab.III). The most abundant taxons were r e p r e s e n t e d b y C r u s t a c e a ( C o r o p h i i d a e , 22%; Sphaeromatidae, 11%; Tanaidae, 11%; and Cumacea, 7%); by Mollusca (Hidrobiidae, 18% and Corbiculidae, 5%); by Insecta ( C h i r o n o m i d a e , 9 % ) a n d O l i g o c h a e t a , 9 % , corresponding to more than 90% of the macroinvertebrate community.
M a c r o i n v e r t e b r a t e c o m m u n i t y c o m p o s i t i o n a n d a b u n d a n c e i n t h e T a i m system varied among the sampling areas. In Flores Lake, Hidrobiidae predominated; i n N i c o l a L a k e , C h i r o n o m i d a e a n d Tubificidae were most common; and in Ja-caré Lake, Polymitarcyidae and Tubificidae were most found. The Mangueira Lake /Taim W e t l a n d p o i n t w a s c h a r a c t e r i z e d b y a n a l t e r n a t i o n o f f a m i l i e s d u r i n g t h e t h r e e
Table I: Environmental characteristics at the sampling stations in the Taim Hydrological System – RS in Autumn (March) and Winter (June ) 2001.
FL
NI
JA
TB
TN
TC
TS
Environmental
Variables
Aut 01 Win 01 Aut 01 Win 01 Aut 01 Win 01 Aut 01 Win 01 Aut 01 Win 01 Aut 01 Win 01 Aut 01 Win 01 Depth (m) 2.8 3.5 1.1 2.1 1.9 2.5 2.3 2.5 2.2 2.8 2.1 2.4 2.3 2.51 ZSD (m) 0.23 0.35 0.32 0.3 0.58 0.3 1.55 0.6 1.2 0.6 0.8 0.88 1.2 1.2 Temperature (°C) 24.44 13.6 24 13.31 24.17 13.55 22.81 10.99 23.13 11.78 23.5 14.81 22.42 14.12 Conductivity (mS/cm) 0.1 0.063 0.379 0.171 0.387 0.263 0.334 0.266 0.388 0.275 0.38 0.279 0.36 0.266 Saturated Oxygen (%) 100 105.1 82.9 108.2 81.7 100.9 93.5 103.8 99.8 106.3 99 105.4 99.1 106.6 pH 7.53 7.48 7.81 7.93 7.74 7.79 7.9 7.86 8.12 8.19 8.51 8.16 8.6 8.32 Total Solids (mg/L) 22.5 52.91 40 43.98 42 54.8 20 41.2 20.5 54.7 31.5 141.8 25 38.7 Total DQO (mg O2/L) 15.2 27 36.8 22 26.8 23 35.4 24 26.4 17 18.2 11 16.7 17 Total N (mg N /L) 0.189 0.4039 0.103 0.360 0.091 0.852 0.103 0.404 0.147 0.261 0.123 0.788 0.177 0.675 Total P (mg P /L) 0.06 0.069 0.049 0.042 0.0198 0.011 0.083 0.086 0.047 0.047 0.045 0.049 0.072 0.064 Chloride (mg Cl-/L) 17.95 89.184 69.03 98.155 68.38 86.035 69.44 42.23 65.81 108.518 65.02 115.289 74.4 88.937 Chlorophyll a (ug/L) 0.9 5.1 0.3 4.5 0.2 7.5 0.6 7.7 2.1 7.3 0.9 6.7 0.3 8.1 * F L = F l o r e s L a k e , N I = N i c o l a L a k e , J A = J a c a r é L a k e , T B = M a n g u e i r a L a k e / T a i m w e t l a n d , T N = N o r t h M a n g u e i r a L a k e , T C = C e n t r a l M a n g u e i r a L a k e , T S = S o u t h M a n g u e i r a - L a k e , A u t = a u t u m n , W i n = w i n t e r .Sampling station
Textural Classification*
Organic matter %
NI Mud with sand 27.70%
TB Sand with mud 22.70%
FL Sand with mud 9.95%
JA Mud 40.90%
TC Sand 0
TS Sand 0
TN Sand 12.90%
Table II: Sediment classification and organic matter percentage.
* F r o m : V o l k m e r - R i b e i r o e t . a l . 2 0 0 6 .
F L = F l o r e s L a k e , N I = N i c o l a L a k e , J A = J a c a r é L a k e , T B = M a n g u e i r a L a k e - T a i m w e t l a n d , T N = N o r t h M a n g u e i r a L a k e , T C = C e n t r a l M a n g u e i r a L a k e , T S = S o u t h M a n g u e i r a L a k e .
T a b l e I I I : C o m p o s i t i o n a n d c l a s s i f i c a t i o n b y f u n c t i o n a l g r o u p s o f b e n t h i c m a c r o f a u n a f o u n d i n T a i m Hydrological System lakes – RS.
Class
Order
Family
Genus or species
Functional
Group
Oligochaeta Tubificida Naididae Unidentified* Gathering Collector
Tubificidae Unidentified * Gathering Collector
Enchytraidae Unidentified * Gathering Collector
Hirudínea Rhynchobdellida Glossiphonidae Unidentified * Predator
Hirudinea morphotype A Unidentified * Predator
Turbellaria Planariidae Unidentified * Predator
Bivalvea Hyriidae Diplodon sp. Filterer
Veneroida Corbiculidae Neocorbicula sp. Filterer
Corbicula fluminea
Müller, 1774 Filterer
Sphaeridae Pisidium sp. Scraper
Gastropoda Mesogastropoda Hidrobiidae Heleobia sp. Scraper
Heleobia parchappii
Orbigny, 1835 Scraper
Chilinidae Chilina fluminea parva
Martens, 1868 Scraper
Crustacea Amphipoda Corophiidae Leptocheirus sp. Gathering Collector
Tanidacea Tanaidae Sinelobus stanfordi
Richardson, 1901
Gathering Collector
Isopoda Sphaeromatidae Unidentified * Gathering Collector
Cumacea Unidentified * Gathering Collector
Decapoda Aeglidae Aegla prado Schmith,
1942
Gathering Collector
Insecta Diptera Chironomidae Procladius sp. Shredder
Djalmabatista pulcher Tennessen & Gottfried, 1983 Predator Coelotanypus sp. Shredder Clinotanypus sp. Shredder Ablabesmyia (Karelia) group Shredder
Aedokritus sp. Gathering Collector Axarus sp. Gathering Collector Chironomus sp. Gathering Collector Polypedilum (Tripodura)
group
Gathering Collector Harnischia sp. Gathering Collector Saetheria sp. Gathering Collector Eifeldia sp. Gathering Collector Cladopelma sp. Gathering Collector
c o l l e c t i o n p e r i o d s , h i g h l i g h t i n g T a n a i d a e , C o r o p h i i d a e , a n d P o l y m i t a r c y i d a e . I n t h e n o r t h e r n p o i n t o f M a n g u e i r a L a k e , Corophiidae, Tanaidae, and Chironomidae p r e d o m i n a t e d ; i n t h e c e n t r a l p o i n t , C u m a c e a , O l i g o c h a e t a , j u v e n i l e f o r m s o f Ve n e r o i d a , a n d S p h a e r o m a t i d a e w e r e t h e most abundant; and in the southern point, Hidrobiidae and Sphaeromatidae composed the majority of the community (Tab. IV).
The average benthic macroinvertebrate density in the lakes at north of Mangueira Lake during the three study periods varied f r o m 2 9 7 t o 1 , 2 5 5 i n d / m2 ( T a b . I V ) . T h e
sampling point that represents the interface between the Taim Wetlands and Mangueira L a k e h a d a n a v e r a g e d e n s i t y o f 1 , 6 3 3 ind/m² a nd Mangueira Lake registered the h i g h e s t a v e r a g e s , v a r y i n g f r o m 5 , 2 9 7 t o 7,547 ind/m2 (Tab. IV).
C o m p a r i n g t h e a v e r a g e b e n t h i c organism density values during the three s a m p l i n g p e r i o d s i n t h e T a i m s y s t e m , a s well as family richness, a tendency toward h i g h e r d e n s i t i e s a n d f a m i l y r i c h n e s s w a s observed in the points situated at Mangueira Lake. T h e d i v e r s i t y i n d e x d i d n o t e x h i b i t a n y g r e a t a l t e r a t i o n s t h r o u g h o u t t h e system, though higher value was observed a t t h e M a n g u e i r a L a k e / T a i m W e t l a n d i n t e r f a c e . T h e r e s u l t s o f A N O VA t e s t i n d i c a t e d t h a t m e a n d e n s i t i e s o f i n v e r t e b r a t e c o m m u n i t y a n d f a m i l y richness was significantly higher in north
a n d s o u t h M a n g u e i r a L a k e s a m p l i n g s t a t i o n s . A m o n g s e a s o n s t h e r e w a s n o significant differences, but the interaction results among sample points and seasons were significant (Tab.VI).
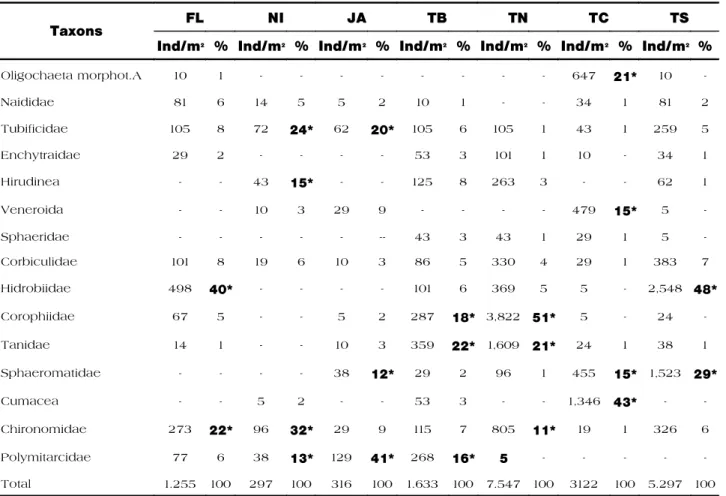
T h e c l u s t e r a n a l y s i s c o n s i d e r i n g t h e a v e r a g e d e n s i t y v a l u e s ( i n d / m2) a n d
b e n t h i c m a c r o i n v e r t e b r a t e c o m p o s i t i o n defined 4 groups (Fig. 2). Group 1 united samples from the Central Mangueira station ( T C ) , w h i c h d i s t i n g u i s h e d i t s e l f f r o m t h e o t h e r g r o u p s b y t h e a b u n d a n c e o f c r u s t a c e a n f a m i l i e s ( C u m a c e a 4 3 % a n d Sphaeromatidae 15%). Group 2 was formed by sampling stations from the interior lakes of the Taim Wetland, Nicola Lake (NI) and J a c a r é L a k e ( J A ) , a n d h a d i n c o m m o n a g r e a t e r a b u n d a n c e o f P o l y m i t a r c y i d a e , T u b i f i c i d a e a n d C h i r o n o m i d a e , w h i c h r e p r e s e n t e d 6 9 % o f t h e c o m m u n i t y. F u r t h e r m o r e , t h e t w o l a k e s p r e s e n t e d l o w e r a v e r a g e i n d i v i d u a l d e n s i t i e s i n comparison to the other groups, with 297 and 316 ind/m², respectively (Tab. IV).
G r o u p 3 u n i t e d t h e M a n g u e i r a L a k e / Taim Wetland interface (TB) and the North M a n g u e i r a L a k e ( T N ) s a m p l i n g s t a t i o n s , which presented differentiated average in-dividual densities, 1,630 and 7,547 ind/m², r e s p e c t i v e l y. N o n e t h e l e s s , t h e y w e r e s i m i l a r i n r e g a r d s t o i n v e r t e b r a t e c o m p o s i t i o n w i t h a p r e d o m i n a n c e o f Corophiidae (18 and 51%, respectively) and
Ta b l e I I I : C o n t .
Class
Order
Family
Genus or species
Functional
Group
Insecta Diptera Chironomidae
Cryptochironomus sp2 sensu Trivinho-Strixino & Strixino, 1995
Predator Glyptotendipes sp. Gathering Collector
Rheotanytarsus sp. Filterer
Caladomya sp. Gathering Collector
Lopescladius sp. Shredder
Nanocladius sp. Shredder
Ephemeroptera Polymitarcyidae Unidentified * Gathering Collector
Baetidae Unidentified * Gathering Collector
Caenidae Unidentified * Gathering Collector
Ephemeridae Unidentified * Gathering Collector
FL
NI
JA
TB
TN
TC
TS
Taxons
Ind/m
2% Ind/m
2% Ind/m
2% Ind/m
2% Ind/m
2% Ind/m
2% Ind/m
2%
Oligochaeta morphot.A 10 1 - - - 647 21* 10 - Naididae 81 6 14 5 5 2 10 1 - - 34 1 81 2 Tubificidae 105 8 72 24* 62 20* 105 6 105 1 43 1 259 5 Enchytraidae 29 2 - - - - 53 3 101 1 10 - 34 1 Hirudinea - - 43 15* - - 125 8 263 3 - - 62 1 Veneroida - - 10 3 29 9 - - - - 479 15* 5 - Sphaeridae - - - -- 43 3 43 1 29 1 5 - Corbiculidae 101 8 19 6 10 3 86 5 330 4 29 1 383 7 Hidrobiidae 498 40* - - - - 101 6 369 5 5 - 2,548 48* Corophiidae 67 5 - - 5 2 287 18* 3,822 51* 5 - 24 - Tanidae 14 1 - - 10 3 359 22* 1,609 21* 24 1 38 1 Sphaeromatidae - - - - 38 12* 29 2 96 1 455 15* 1,523 29* Cumacea - - 5 2 - - 53 3 - - 1,346 43* - - Chironomidae 273 22* 96 32* 29 9 115 7 805 11* 19 1 326 6 Polymitarcidae 77 6 38 13* 129 41* 268 16* 5 - - - - - Total 1.255 100 297 100 316 100 1.633 100 7.547 100 3122 100 5.297 100 Table IV: Average total density values (ind/m2) and relative abundance (%) of the benthic macroinvertebrate
community in the Taim Hydrological System, southern coastal region of Rio Grande do Sul.
F L = F l o r e s L a k e , N I = N i c o l a L a k e , J A = J a c a r é L a k e , T B = M a n g u e i r a L a k e - T a i m W e t l a n d , T N = n o r t h M a n g u e i r a L a k e , T C = c e n t r a l M a n g u e i r a L a k e , T S = s o u t h M a n g u e i ra L a k e .
Among environment
Among season
Interaction
Biologic
Descriptors
F
Scheffé
Results
F
Scheffé
Results
F
Scheffé
Results
Mean Density
7.46 TN, TS> FL (p=0.0001) 1.54 NS (p=0.225) 5.61 p=0.0001S
11.20 TN, TS> NI, JA FL>NI, JA (p=0.0001) 0.81 NS (p=0.450) 1.16 NS (p=0.338)J’
1.46 NS (p=0.216) 1.16 NS (p=0.323) 0.47 NS (p=0.923)H’
2.67 ND (p=0.027) 3.10 NS (p=0.055) 0.69 NS (p=0.753)Table V: ANOVA test results considering mean density of taxonomic groups, family richness, equitability and diversity index, among sample points and seasons and interaction among them. (QM = m e a n s q u a r e ; F = d i s t r i b u t i o n o f t h e v a r i a n c e ; N S = n o n s i g n i f i c a n t d i f f e r e n c e a n d N D = n o n detected difference).
F L = F l o r e s L a k e , N I = N i c o l a L a k e , J A = J a c a r é L a k e , T B = M a n g u e i r a L a k e - T a i m W e t l a n d , T N = n o r t h M a n g u e i r a L a k e , T C = c e n t r a l M a n g u e i r a L a k e , T S = s o u t h M a n g u e i r a L a k e .
Tanidae (22 and 21%, respectively) families. G r o u p 4 w a s f o r m e d b y F l o r e s L a k e ( F L ) a n d S o u t h M a n g u e i r a L a k e ( T S ) s a m p l i n g s t a t i o n s , w h i c h w e r e c h a r a c t e r i z e d b y a greater abundance of Hidrobiidae (40 and 48%, respectively).
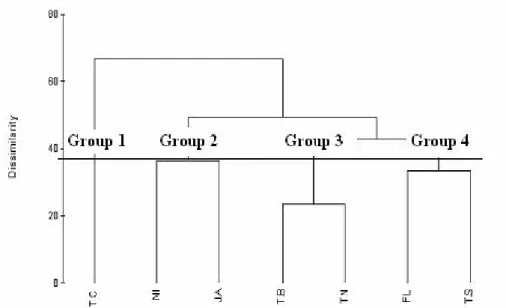
From the results obtained in the Cluster analysis were calculated the percentual of f u n c t i o n a l f e e d i n g g r o u p s o f t h e T a i m benthic macroinvertebrates community, that w a s c h a r a c t e r i z e d b y a p r e d o m i n a n c e o f g a t h e r i n g c o l l e c t o r s f o l l o w i n g i n o r d e r o f abundance by scrapers and filterers (Fig.3).
F i g u r e 2 : C l u s t e r a n a l y s i s c o n s i d e r i n g t h e a v e r a g e t o t a l d e n s i t i e s ( i n d / m2) a n d m a c r o i n v e r t e b r a t e composition at the THS sampling stations – southern coast of Rio Grande do Sul.
GROUP 1 Gathering Collectors 76,4% Scrapers 3,6% Predators 0,4% Filtering Collectors 19,6% GROUP 2 Predators 19,6% Scrapers 5,1% Filthering Collectors 17,5% Gathering Collectors 57,8% GROUP 3 Gathering Collectors 81,7% Filthering Collectors 5,3% Scrapers 9,1% Predators 3,9% GROUP 4 Scrapers 48,1% Gathering Collectors 39,4% Predators 4,4% Filthering Collectors 8,1%
Figure 3: Percentage composition of invertebrates functional feeding groups according distribution in c l u s t e r a n a l y s i s .
F L = F l o r e s L a k e , N I = N i c o l a L a k e , J A = J a c a r é L a k e , T B = M a n g u e i r a L a k e - T a i m W e t l a n d , T N = n o r t h M a n g u e i r a L a k e , T C = c e n t r a l M a n g u e i r a L a k e , T S = s o u t h M a n g u e i r a L a k e .
G r o u p s 1 , 2 a n d 3 a r e v e r y s i m i l a r , t h e s e c o n d d i f f e r i n g s o m e t h i n g b y t h e abundance of predators. Group 4 differed of the others by the abundance of scrapers.
Discussion
T h e T a i m H y d r o l o g i c a l S y s t e m i s compound by small lakes such as Flores Lake, interior lakes of the Taim Wetland, like Nicola and Jacaré and the large Mangueira Lake. We can expected that the macrofauna c o m m u n i t y d i f f e r e d a c c o r d i n g t h e environmental changes observed in these ecosystems. Communities are structured by both abiotic and biotic factors nested along s p a t i a l a n d t e m p o r a l s c a l e s a n d t h a t knowledge of relationships and ecological l i n k a g e s i s i m p o r t a n t f o r u n d e r s t a n d i n g large- and small- scale variability (Johnson & G o e d k o o p , 2 0 0 4 ) . S t o f f e l s e t a l . ( 2 0 0 5 ) c o m m e n t s t h a t a t f i n e r s c a l e s , t h e c o m p o s i t i o n o f s u b s t r a t e s , p a r t i c u l a r l y macrophytes and inorganic sediments, may be drivers of community spacial structure. T h e r e s u l t s o f A N O VA t e s t i n d i c a t e d t h a t mean densities of invertebrate community a n d f a m i l y r i c h n e s s d i f f e r e d s i g n i f i c a n t l y along the Taim Hydrological System. Local c h a r a c t e r i s t i c s l i k e d i f f e r e n c e s i n substratum and the proximity of the wetland v e g e t a t i o n p r o b a b l y c a n e x p l a i n t h e c h a n g e s i n t h e c o m p o s i t i o n , d e n s i t y a n d richness of the macrobenthos.
Physical and chemical data measured in the studied lakes are relatively similar, b u t s e d i m e n t g r a i n s i z e a n d t h e o r g a n i c matter presented variations (Tab.II). Jacaré and Nicola lakes, have registered 40.95% and 27.7% of organic matter and sedimentar s u b s t r a t u m o f m u d a n d s a n d y m u d , r e s p e c t i v e l y . T h e b e n t h i c c o m m u n i t y s t r u c t u r e i s f o r m e d b y T u b i f i c i d a e , Chironomidae and Polymitarcyidae groups, s u g g e s t i n g a f o o d c h a i n p r e d o m i n a n t l y based on detritus. Lakes Jacaré and Nicola naturally receive a high detritic load from t h e T a i m W e t l a n d ’ s m a c r o p h y t i c b a n k s , which may corroborate with over 70% of t h e d e p o s i t f e e d e r s . A c c o r d i n g S p i e l e s & M i t s c h ( 2 0 0 0 ) , t h e d i s t r i b u t i o n a n d a b u n d a n c e r e g a r d i n g t h e i n v e r t e b r a t e f u n c t i o n a l g r o u p s i n w e t l a n d s a n d o t h e r aquatic systems are influenced by organic material availability as a food source.
I n F l o r e s L a k e , t h e s e d i m e n t i s s a n d w i t h m u d , a n d s c r a p p e r s r e p r e s e n t e d b y t h e H i d r o b i i d a e s h o w e d g o o d r e p r e s e n t a t i v i t y , w i t h 4 0 % o f a b u n d a n c e . A c c o r d i n g t o S z a l a y & R e s h ( 1 9 9 6 ) , t h e p e r c e p t i o n t h a t a l g a l h e r b i v o r e s a r e r e l a t i v e l y u n i m p o r t a n t i n w e t l a n d m a c r o i n v e r t e b r a t e c o m m u n i t i e s m a y b e based on a lack of knowledge on the taxa f e e d i n g e c o l o g y. M a n y s t u d i e s d o n o t distinguish between those taxa feeding on detritus or algae. Ephiphytic habitats and benthic algal biomass available in Flores L a k e p r o b a b l y s u p p o r t H i d r o b i i d a e a n d explain its high abundance.
T h e s u b s t r a t u m o f M a n g u e i r a L a k e / T a i m W e t l a n d a n d t h e N o r t h M a n g u e i r a s a m p l i n g s t a t i o n s d i f f e r s s o m e t h i n g f r o m Jacaré and Nicola lakes registering 22.70% a n d 1 2 . 7 0 % o f o r g a n i c m a t t e r a n d s u b s t r a t u m o f m u d y s a n d a n d s a n d , respectively. In the first one the macrofauna a r e c h a r a c t e r i z e d b y t h e p r e s e n c e o f Tanaidae, Corophiidae, and Polymitarcyidae. N o r t h M a n g u e i r a e x h i b i t e d t h e s e s a m e f a m i l i e s , a n d C h i r o n o m i d a e a d d i t i o n a l l y. These two points differed from the others by a greater abundance of Corophiidae and T a n a i d a e f a m i l i e s . T h u s , t h e s a m p l i n g stations also exhibited a food chain made up of detritus. According Merritt & Cummins (1996) depending on the food source, some t a x a c a n c h a n g e t h e i r f e e d i n g h a b i t s . Amphipoda and Tanaidacea could feed on c o a r s e o r g a n i c m a t t e r a n d b e h a v e a s shredders, changing the trophic structure at these points in Mangueira Lake. This area is prone to receive coarse organic matter f r o m t h e T a i m W e t l a n d s u n d e r s p e c i a l hydrodynamic conditions which draw water f r o m t h e w e t l a n d s t o w a r d M a n g e i r a L a k e (Fragoso Jr., 2005).
A t c e n t r a l a n d s o u t h L a k e M a n g u e i r a t h e s e d i m e n t i s s a n d y a n d t h e o r g a n i c matter was not detected.
I n c e n t r a l M a n g u e i r a s t a t i o n t h e crustaceans, Cumacea and Sphaeromatidae corresponded to 58% of the community, and together with Oligochaeta, form a trophic structure of deposit feeders. This point of M a n g u e i r a L a k e i s f a r t h e r f r o m t h e m a c r o p h y t i c b a n k s a n d , t h e r e f o r e , m u s t receive organic material from other areas of the lake. Due to the strong presence of H i d r o b i i d a e , S o u t h M a n g u e i r a L a k e a l o n e h a d a c o m m u n i t y s t r u c t u r e w h e r e t h e d o m i n a n t t r o p h i c g r o u p w a s s c r a p e r s , followed by gathering collectors.
I n t h e T a i m H y d r o l o g i c a l S y s t e m , c o n s i d e r i n g t h e s y s t e m a s a w h o l e , a
p r e d o m i n a n t l y d e t r i t i c f o o d c h a i n w a s o b s e r v e d . T h e p r e d o m i n a n c e o f a detritivore food chain was also observed in most of the lakes in the northern coastal region of Rio Grande do Sul (Würdig et al., 1998). The fauna composition is quite simi-lar, with the exception of the presence of C o r o p h i i d a e a n d t h e g r e a t d e n s i t y o f C u m a c e a , p r e s e n t i n b o t h t h e w e t l a n d zones and principally in the free waters of Mangueira Lake.
C o r o p h i i d a e a n d C u m a c e a a r e characteristics in environments with a saline influence, such as lagoons and estuarine regions, conditions that were not observed on present-day in the Taim Wetlands. The p r e s e n c e o f t h e s e s p e c i e s m i g h t b e e x p l a i n e d b y t h e m a r i n e o r i g i n o f t h i s coastal system (Vilwock, 1984) and by the considerable tolerance of these groups to s a l i n i t y g r a d i e n t s . H i s t o r i c a l p r o c e s s e s s u p p o r t t h e p r e s e n c e o f t h e s e t a x a , considering an ecological and large tempo-r a l s c a l e . J o h n s o n e t a l . ( 2 0 0 4 ) i n t h e i tempo-r c o m p a r a t i v e s t u d i e s a b o u t s p a t i a l s c a l e s a n d e c o l o g i c a l r e l a t i o n s h i p s b e t w e e n m a c r o i n v e r t e b r a t e s f i n d t h a t s p e c i e s distribution is determined fundamentally by conditions prevailing at the local scale and important regional factors such as land use and the role of history which act as strong d e t e r m i n a n t s o f l a r g e - s c a l e p a t t e r n s i n biodiversity.
T h e m a c r o i n v e r t e b r a t e c o m m u n i t y s t r u c t u r e i n t h e w e t l a n d s a n d a s s o c i a t e d lakes of Taim Hidrological System differ in local scale due to many components, like macrophytes presence, detritus abundance, s e d i m e n t g r a n u l o m e t r y , a n d w a t e r h y d r o d y n a m i c s , w h i c h c o n t r i b u t e d t o t h e high variability along the system. However m o r e a n d o b j e c t i v e s t u d i e s d e a l i n g w i t h different spatial and temporal scales that influence the distribution and structure of the macrofauna communities are necessary, i n o r d e r t o e x p l a i n t h e i m p o r t a n c e o f ecological characteristics and processes at different levels in this System, permiting to i n t e r p r e t c h a n g e s i n t h e f a u n a a n d i n ecosystem responses to natural and human-induced perturbation.
Acknowledgements
This part of the work developed in site 7 o f t h e B r a z i l i a n L o n g T e r m E c o l o g i c a l Research network. We thank the research development agency, CNPq (Conselho
Na-c i o n a l d e D e s e n v o l v i m e n t o C i e n t í f i Na-c o e Tecnológico) for the research grants. Special thanks to Dr. Simone Kapusta for her critical reading of the manuscript, to Dr. Maria Te-r e s a B e Te-r a Te-r d o ( U n i v e Te-r s i d a d e P Te-r e s b i t e Te-r i a n a Mackenzie), and Dr. Yoko Wakabara (Univer-sidade Santa Úrsula) for the identification of the Amphipoda genus Leptocheirus, and t o D r . M a r i a C r i s t i n a P o n s f o r t h e identification of the Mollusca species. Also we acknowledge the anonymous reviewers w h i c h c o m m e n t s g r e a t l y i m p r o v e d t h e manuscript.
References
A m e r i c a n P u b l i c H e a l t h A s s o c i a t i o n – A P H A . 1 9 9 5 . S t a n d a r d m e t h o d s f o r e x a m i n a t i o n o f w a t e r a n d w a s t e w a t e r. A m e r i c a n P u b l i c H e a l t h A s s o c i a t i o n , Wa s h i n g t o n . 1 1 9 3 p. B r i n k h u r s t , R . O . & M a r c h e s e , M . R . 1 9 8 9 . G u i d e t o t h e f r e s h w a t e r a q u a t i c O l i g o c h a e t a o f s o u t h a n d C e n t r a l America. Associacion Ciencias Naturales del Litoral, São Tomé, Argentina. 236p. (Coleción Climax, 6).Clarke, K.R. & Warwick, R.M. 2001. Change in marine communities: an approach to statistical analysis and interpretation. 2ª ed. PRIMER- E, Plymouth. 144p.
D o w n i n g , J . A . 1 9 9 1 . T h e e f f e c t o f h a b i t a t s t r u c t u r e o n t h e s p a t i a l d i s t r i b u t i o n o f freshwater invertebrates populations. In: Bell, S.S., McCoy, E.D. & Mushinsky, H.R. (eds.) The habitat structure: the physical a r r a n g e m e n t o f o b j e c t s i n s p a c e . Chapman and Hall, London. p.87-102. Epler, J.H. 2001. Identification manual for
the larval Chironomidae (Diptera) of North a n d S o u t h C a r o l i n a . E PA Region 4 and H u m a n H e a l t h a n d E c o l o g i c a l C r i t e r i a D i v i s i o n . Ve r s i o n 1 . 0 . N o r t h C a r o l i n a Department of Environment and Natural Resources, Raleigh. p.526.
Fragoso Jr., C.R. 2005. Simulações da dinâ-m i c a d e f i t o p l â n c t o n n o S i s t e dinâ-m a Hidrológico do Taim (RS). Porto Alegre, UFRGS, 137p (Master Thesis).
H e i n o , J . , L o u h i , P. & M u o t k a , T. 2004. I d e n t i f y i n g t h e s c a l e s o f v a r i a b i l i t y i n s t r e a m m a c r o i n v e r t e b r a t e a b u n d a n c e , f u c t i o n a l c o m p o s i t i o n a n d a s s e m b l a g e structure. Freshwater Biol., 49:1230-1239. Johnson, R.K. & Goedkoop, W. 2002. Littoral macroinvertebrate communities: spatial s c a l e a n d e c o l o g i c a l r e l a t i o n s h i p s . Freshwater Biol., 47:1840-1854.
Johnson, R.K., Goedkoop, W. & Sandin, L. 2 0 0 4 . S p a t i a l s c a l e a n d e c o l o g i c a l r e l a t i o n s h i p s b e t w e e n t h e macroinvertebrate communities of stony habitats of streams and lakes. Freshwater Biol., 49:1179-1194.
L e g e n d r e , P. L . & L e g e n d r e , L . 1 9 9 8 . N u m e r i c a l e c o l o g y : d e v e l o p m e n t s i n e n v i r o n m e n t a l m o d e l i n g 2 0 . 2 ª e d . Elsevier, New York. 853p.
Mackereth, F.J.H., Heron, J. & Talling, J.F. 1978. W a t e r a n a l y s i s : s o m e r e v i s e d m e t h o d s f o r l i m n o l o g i s t s . 2n d e d . F r e s h w a t e r
Biological Association, Ambleside. 120p. McCafferty, W.P. 1981. Aquatic entomology: the fishermen’s and ecologists’ illustrated guide to insects and their relatives. Jones and Bartlett Publishers, Boston. 448p. Merritt, R.W. & Cummins, K.W. 1996. Trophic
relations of macroinvertebrates. In: Hauer, F.R. & Lamberti, G.A. (eds.) Methods in s t r e a m e c o l o g y . A c a d e m i c P r e s s , S a n Diego. p.453-474.
Motta Marques, D.M.L., Tucci, C., Calazans, D., Callegaro, V.L.M. & Villanueva, A. 2002. O sistema hidrológico do Taim Site 7. In: Seeliger, U., Cordazzo, C. & Barbosa, F. (eds.) Os sites e o Programa Brasileiro de Pesquisas Ecológicas de Longa Dura-ção. O Lutador, Belo Horizonte. p.127-144. Shieh, S.H., Kondratieff, B.C. & Ward, J.V. 1 9 9 9 . L o n g i t u d i n a l c h a n g e s i n b e n t h i c organic matter and macroinvertebrates in a p o l l u t e d C o l o r a d o p l a i n s s t r e a m . Hydrobiologia, 411:191-209. S p i e l e s , D . J . & M i t s c h , W. 2000. M a c r o i n v e r t e b r a t e c o m m u n i t y s t r u c t u r e i n h i g h - a n d l o w - n u t r i e n t c o n s t r u c t e d wetlands. Wetlands, 20:716-729.
Stoffels, K.R., Clarke, K.R. & Closs, G.P. 2005. S p a t i a l s c a l e a n d b e n t h i c c o m m u n i t y organization in the littoral zones of large o l i g o t r o p h i c l a k e s : p o t e n c i a l f o r c r o s s -s c a l e i n t e r a c t i o n -s . F r e -s h w a t e r B i o l . , 50:1131-1145.
Szalay, F.A. & Resh, V.H. 1996. Spatial and temporal variability of trophic relationship among aquatic macroinvertebratres in a seasonal marsh. Wetlands, 16:458-466. Trivinho-Strixino, S. & Strixino, G. 1995.
Lar-vas de Chironomidae (Diptera) do esta-do de São Paulo: guia de Indentificação e d i a g n o s e d o s g ê n e r o s . P r o g r a m a d e Pós-Graduação em Ecologia e Recursos N a t u r a i s , U n i v e r s i d a d e F e d e r a l d e S ã o Carlos, São Paulo. 229p.
Villanueva, A.O.N., Mendiondo, E.M., Tucci, C.E.M. & Louzada, J.A. 1998.
Determina-ção de critérios de irrigaDetermina-ção para preser-vação do Banhado do Taim. Anais do II Simpósio de Recursos Hídricos do Cone Sul. Ed. UNL, Santa Fé, v.2, p.91-99. Vilwock, J.A. 1984. Geology of the coastal
province of Rio Grande do Sul, Southern Brazil: a synthesis. Pesquisas, 16:5-49. Volkmer-Ribeiro, C., Motta-Marques, D., De
Rosa-Barbosa, R. & Machado, V.S. 2006. S p o n g e s p i c u l e s i n s e d i m e n t s i n d i c a t e evolution of coastal freshwater bodies. J. Coastal Res., 39:469-472.
Würdig, N.L., Ozório, C.P., Rodrigues, G.G. & W i e d e n b r u g , S . 1 9 9 8 . T h e i n f l u e n c e o f e n v i r o n m e n t a l p a r a m e t e r s i n t h e s t r u c t u r e o f t h e b e n t h i c c o m m u n i t y i n coastal lakes and lagoons of Rio Grande do Sul, Brasil. Rev. Int. Verein. Limnol., 26:1514-1517.
Received: 14 October 2005