INFLUENCE OF GENES CONTROLLING FLOWER COLOR ON RELATIVE QUANTITIES OF ANTHOCYANINS AND FLAVONOLS IN PETALS OF IMPATIENS BALSAMINA
7
0
0
Full text
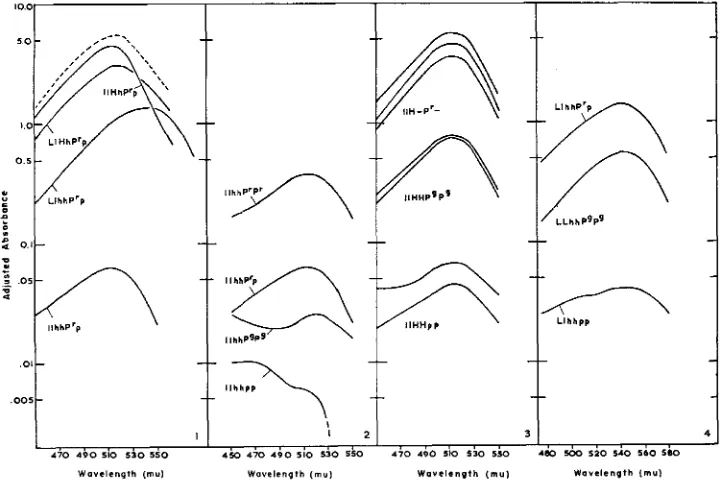
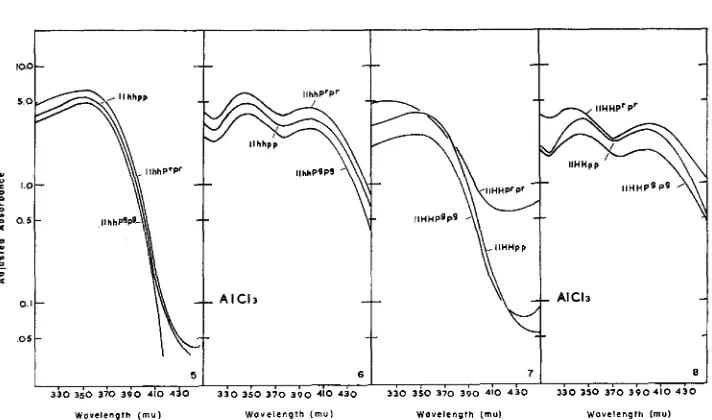
Figure
Related documents