Copyright2000 by the Genetics Society of America
Quantitative Analysis of Gene Function in the Drosophila Embryo
William D. Tracey, Jr., Xiangqun Ning, Martin Klingler,
1Sunita G. Kramer
2and J. Peter Gergen
Department of Biochemistry and Cell Biology and the Institute for Cell and Developmental Biology, State University of New York, Stony Brook, New York 11794-5215
Manuscript received July 8, 1999 Accepted for publication September 15, 1999
ABSTRACT
The specific functions of gene products frequently depend on the developmental context in which they are expressed. Thus, studies on gene function will benefit from systems that allow for manipulation of gene expression within model systems where the developmental context is well defined. Here we describe a system that allows for genetically controlled overexpression of any gene of interest under normal physiological conditions in the early Drosophila embryo. This regulated expression is achieved through the use of Drosophila lines that express a maternal mRNA for the yeast transcription factor GAL4. Embryos derived from females that express GAL4 maternally activate GAL4-dependent UAS transgenes at uniform levels throughout the embryo during the blastoderm stage of embryogenesis. The expression levels can be quantitatively manipulated through the use of lines that have different levels of maternal GAL4 activity. Specific phenotypes are produced by expression of a number of different developmental regulators with this system, including genes that normally do not function during Drosophila embryogenesis. Analysis of the response to overexpression of runt provides evidence that this pair-rule segmentation gene has a direct role in repressing transcription of the segment-polarity gene engrailed. The maternal GAL4 system will have applications both for the measurement of gene activity in reverse genetic experiments as well as for the identification of genetic factors that have quantitative effects on gene function in vivo.
T
HE Drosophila embryo provides a remarkable dem- Horowiczand Pinchin1987; Morrissey et al. 1991; onstration of the productive interplay between de- Fitzpatricket al. 1992;ManoukianandKrause1992, velopmental genetics and mechanistic studies on gene 1993;Tsai and Gergen 1994, 1995; John et al. 1995; function. For example, the central role of transcrip- Aronsonet al. 1997; Dong et al. 1998). One concern tional regulation in patterning the embryo resulted in the interpretation of all of these experiments is the from studies on genes identified through the pioneer- fact that the regulatory responses may be nonphysiologi-ing mutational studies of Nu¨ sslein-Volhard and cal as they are obtained in embryos that have beenheat-Wieschaus(1980). These studies have generated a solid shocked.
framework for functional studies on the transcription We have been using the Drosophila embryo as a model factors that participate in these processes. Indeed, ex- to investigate the function of the pair-rule segmentation periments with the Drosophila embryo provide several gene runt. Runt is the founding member of the Runt of the most elegant and well-understood examples of domain family of transcriptional regulators (
Kago-transcriptional regulation in developmental biology. Al- shimaet al. 1993) and has pivotal roles in sex determina-though the roles of many genes in the embryo were tion, segmentation, and neurogenesis in the Drosophila initially deduced from loss-of-function phenotypes, stud- embryo (Gergen and Wieschaus 1986; Duffy and ies on gene function have also benefited from analysis Gergen 1991;Duffyet al. 1991;TorresandSanchez
of gain-of-function phenotypes produced by overexpres- 1992;DormandandBrand1998;Krameret al. 1999). sion. For example, numerous investigations on the regu- Lozenge, a second Runt domain protein in Drosophila, latory interactions and mechanisms of transcriptional has postembryonic roles in patterning in the antenna regulation that are involved in the segmentation path- and eye (Dagaet al. 1996; Flores et al. 1998; Gupta way have used the Drosophila heat-shock promoter to et al. 1998). In vertebrates, Runt domain genes have induce ectopic gene expression (Struhl 1985; Ish- important roles in the normal development of blood
and bone, and mutations in these genes are associated with defects in these processes in humans (Okuda et Corresponding author: J. Peter Gergen, Department of Biochemistry al. 1996;Wanget al. 1996;Ducyet al. 1997;Komoriet
and Cell Biology, SUNY, Stony Brook, NY 11794-5215.
al. 1997;Ottoet al. 1997;Tracey et al. 1998).
E-mail: [email protected]
In Drosophila, runt is required for the proper tran-1Present address: Zoologisches Institut der Universita¨t Mu¨nchen,
Luissenstrasse 14, 80333 Munich, Germany. scriptional regulation of a number of different genes
2Present address: Department of Molecular and Cell Biology, 519
during early embryogenesis. In some instances runt
Life Sciences Addition, University of California Berkeley, Berkeley,
CA 94720. functions to activate transcription (Tsai and Gergen
1995;Krameret al. 1999), whereas on other targets runt fied as being subject to regulation by runt using the heat-shock assay system also respond when runt expression is plays a role in transcriptional repression (Manoukian
andKrause1993;TsaiandGergen1994). Recent stud- driven with the maternal GAL4 system. However, experi-ments with the GAL4 system reveal clear quantitative ies using heat-shock expression assays have further
re-vealed two independent modes of transcriptional re- differences in the sensitivity of different targets. The potential applications of this system for quantitative pression by the Runt protein. The repression of the
pair-rule genes even-skipped (eve) and hairy involves re- studies of gene function in the Drosophila embryo are discussed.
cruitment of the corepressor protein Groucho through a C-terminal VWRPY motif that is conserved on other Runt domain proteins (Aronson et al. 1997). In
con-MATERIALS AND METHODS trast, repression of the segment polarity gene engrailed
(en) and the head gap gene orthodenticle (otd) is achieved Plasmid construction:A CaSpeR P-element transformation vector was constructed that contains the coding region of yeast through a VWRPY- and Groucho-independent pathway
GAL4 immediately downstream of the nanos promoter and (Aronson et al. 1997; Tsaiet al. 1998). Although the
upstream of DNA containing the 39UTR of the Drosophila mechanisms that account for the various regulatory ac- aTub84B gene. The nanos promoter andz250 bp of 59UTR tivities of the Runt protein are not understood, our are contained in a 1.1-kb HindIII1NdeI restriction fragment that was isolated from plasmid pBS-P[nos], generously pro-working model is that Runt activity is modulated
vided by L. Gavis (Princeton University). One step of the through specific interactions with other
context-depen-cloning strategy included destruction of an initiator methio-dent transcriptional regulators.
nine for the Nanos protein that is encoded at the NdeI site of The modular GAL4 system is an alternative and impor- this fragment. The GAL4-encoding fragment was isolated as tant tool that has been developed for the manipulation a HindIII fragment from plasmid pGAT-B (Brandand
Perri-mon 1993). The aTub84B 39 UTR was contained within an of gene expression in Drosophila (Brand and
Perri-850-bp XhoI 1 HindIII fragment from plasmid pTa1-59-39 mon 1993). This system takes advantage of the ability
(Theurkaufet al. 1986). These fragments were inserted into
of the yeast transcriptional activator GAL4 to function
the BamHI site of pCaSpeR (Thummelet al. 1988). The NGT in Drosophila cells (Fischer et al. 1988). Drosophila fusion gene is oriented within this vector such that it is tran-strains that express GAL4 specifically at one stage of scribed from the opposite DNA strand as the white marker gene in a divergent manner. Further details of the cloning development or in a specific cell type can be mated to
strategy used to generate this plasmid are available on request. strains carrying a GAL4-responsive transgene that
ex-A Uex-AS-runt construct was made by cloning a 3.2-kb BamHI presses any gene of interest (UASG-X ) in order to induce
fragment isolated from pCaSpeR:hs-runt (Tsaiand Gergen
expression of gene X in a genetically controlled manner. 1994) into the BglII site of the pUAS-T vector (Brandand A number of investigators have used an enhancer-trap Perrimon1993). The portion of the runt mRNA contained within this segment begins four nucleotides upstream of the version of a GAL4-expressing P-element transposon to
initiator methionine and extends through the polyadenylation generate a large number of lines that are useful in
differ-signal to the poly(A) tail of a full-length runt cDNA. ent developmental contexts (Brand and Perrimon
Fly strains and crosses:Drosophila strains were maintained 1993;Yanget al. 1995;GustafsonandBoulianne1996; on standard cornmeal/yeast/sugar and agar media. General
Manseauet al. 1997). Notable by their absence in these information on marker mutations and balancer chromosomes is available throughFlyBase(1999). Germ-line transformants collections are lines that express GAL4 at significant
carrying the P{w[1mC] Scer\GAL4[nos.PG]5GAL4-nos.NGT} levels during the early stages of embryogenesis, the
and P{UAS-runt.T} transposons were recovered by standard stages that would be most useful for studies on
segmenta-P-element-mediated germ-line transformation protocols using
tion gene products. the p:D2-3 helper plasmid. The P{UAS-runt.T}232 line was
Here we describe a system that allows for GAL4-driven recovered as 1 of 14 initial independent lines and was retained as a representative strong line based on experiments with a ectopic expression in the early embryo. We constructed
Kr-GAL4 driver (M. KlinglerandJ. P. Gergen,unpublished a GAL4 expression vector that uses the promoter from
results). Preliminary characterization based on the lethality the maternally expressed gene nanos and a 39
untrans-obtained with various UAS transgenes indicated that 2 of the lated (UTR) region from a tubulin mRNA to drive ma- initial NGT lines, P{GAL4-nos.NGT}9 and P{GAL4-nos.NGT}11, ternally dependent uniform activation of GAL4-depen- had the highest levels of activity. These 2 lines were mapped to the third and second chromosomes, respectively, and stocks dent transgenes during the blastoderm stage of
homozygous for each of these chromosomes were established. embryogenesis. Through the use of NGT (nanos-G
AL4-P-transposition mediated by theD2-3 third chromosome (
Rob-tubulin) lines that have different levels of GAL4 activity,
ertsonet al. 1988) was used to generate hops from the original
it is possible to quantitatively manipulate expression in P{GAL4-nos.NGT}11 line. All of the lines reported here were the embryo over a range that extends up to an estimated obtained as hops on chromosome II that are associated with a change in eye color. Experiments to generate further NGT 125,000 molecules/blastoderm cell. Specific
pheno-lines on both the second and third chromosomes, as well as types are produced by expression of different
develop-on different balancer chromosomes, are in progress. mental regulators with this system, including genes that
identi-42B.4), en (P{UAS-en.Y}4-1), and the different isoforms of oogenesis (Al-Atia et al. 1985; Gavis and Lehmann pointed (pnt[P1], P{UAS-pnt.P1}3), and pnt[P2], P{UAS-pnt.P2}2) 1994;Salleset al. 1994;Kim-Haet al. 1995;Markussen were obtained from the Bloomington stock center. The
et al. 1995). Furthermore, there is also substantial evi-transposon for the P{UAS-runt.T}232 line maps on
chromo-dence indicating that maternally expressed mRNAs con-some II and is homozygous viable. The P{UAS-runt.T}U15 line,
which also maps on chromosome II, was generated byD2- tain signals that mediate transport from their site of 3-mediated mobilization. Quantitation by RNase protection synthesis in the nurse cells to the developing oocyte indicates that the U15 line is expressed at approximately three- (MacDonald and Struhl 1988; Ephrussi and Leh-fold higher levels than the 232 line (Li1999). The relative
mann1992). This transport is important if the maternal viability of flies carrying different UAS transgenes was
deter-mRNA is to be available for translation during the early mined by mating males heterozygous for the transgene and
the appropriate balancer chromosome (either CyO or TM3) stages of embryogenesis. On the basis of these observa-to virgin females from the pertinent NGT sobserva-tock. In experi- tions, we reasoned that GAL4 transgenes that contained ments with P{UAS-en.Y}4-1 (which is on the TM6 balancer), appropriate cis-regulatory elements for transcription, males were heterozygous for the ruPrica marker chromosome.
translation, and mRNA transport during oogenesis These crosses were carried out in vials at 258in uncrowded
might be active during oogenesis and early embryogene-conditions.
Embryo manipulation:For cuticle preparations, nonhatch- sis. The two elements that we used to test this hypothesis ing embryos were dechorionated in bleach, rinsed with dis- were the promoter from the maternal effect gene nos, tilled water, and mounted in a 1:1 mixture of lactic acid and and the 39UTR from theaTub84B gene. nos is specifi-Hoyer’s. In situ hybridization was carried out as described
cally expressed in germ cells with high expression levels previously (KlinglerandGergen1993). Digoxigenin-labeled
in the ovary in germ-line cysts from region 2 of the (Boehringer Mannheim, Indianapolis) RNA riboprobes to
de-tect the lacZ, runt, eve, and fushi tarazu (ftz) mRNA transcripts germarium (Wang et al. 1994; Forbes and Lehmann were synthesized as described previously (Tsaiand Gergen 1998). TheaTub84B gene product is abundant during
1994). Expression of en was detected using a probe synthesized both oogenesis and early embryogenesis (Matthewset from HindIII digested pB:en (gift of D. Ish-Horowicz, ICRF)
al. 1989), suggesting that the 39 UTR of this mRNA template with T7 RNA polymerase.
would allow both for transport into the oocyte and for The quantitative measurement of b-galactosidase activity
was done using a luminescent substrate and the Galacto-Light translation during early embryogenesis. The anticipated Plus detection kit (Tropix, Bedford, MA). Single living em- properties of the NGT fusion gene that we constructed bryos of the appropriate stage were identified by observation are schematically presented in Figure 1.
under oil (Halocarbon Products), transferred into a
micro-A number of transgenic lines carrying the P{Gmicro-AL4- P{GAL4-fuge tube, and homogenized in 50 ml of lysis buffer (100
nos.NGT} were obtained by P-element-mediated germ-mmpotassium phosphate, pH 7.8, 0.2% Triton X-100, 1 mm
dithiothreitol, 100mg/ml bovine serum albumin). One-fifth line transformation. P-element remobilization was also of this homogenate was diluted into 70ml of reaction buffer used to obtain additional derivatives of these initial lines. (100 mmsodium phosphate, pH 8.0, 1 mmmagnesium chlo- Characterization of these lines by a number of criteria ride) containing a 1:100 dilution of the Galacto-Light Plus
indicated that they did indeed express GAL4 maternally, substrate stock solution (Tropix) and incubated at room
tem-and that there were differences in the level of expression perature for 30–60 min. Activity was measured after addition
of 100ml of Light Emission Accelerator (Tropix) containing in the different lines. We used P{GAL4-nos.NGT}40, a 10 mmH2O2on an Optocomp I luminometer (MGM Instru- relatively strong line (see below), to investigate the activ-ments, Inc.). The relative light units reported were all from ity of the NGT transgenes. In situ hybridization reveals readings taken over a 30-sec interval. This assay was calibrated
that embryos derived from homozygous P{GAL4-by adding purifiedb-galactosidase (Sigma, St. Louis) into
ex-nos.NGT}40 females express a paternally inherited UAS-tracts of control embryos. The assay was linear over the full
range tested, from 0.46 to 330 pg with an average measure- lacZ reporter gene during the syncytial blastoderm stage, ment of 153103light units/pg of enzyme.
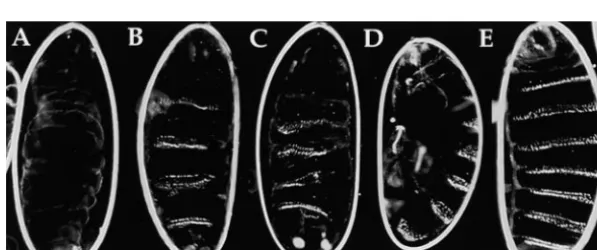
prior to cellularization (Figure 2B). Expression is ob-served at uniform levels throughout the embryo with the exception that no lacZ mRNA accumulates within RESULTS
the pole cells (Figure 2C). Expression levels increase through the completion of cellularization, gastrulation,
A maternal effect GAL4 mRNA:The two-component
GAL4 system (Brand and Perrimon 1993) has been and the process of germband extension (Figure 2, D and E). Reciprocal matings between females that are extremely useful for targeting gene expression to a
num-ber of different tissues and a numnum-ber of different stages homozygous for the UAS-lacZ transgene and homozy-gous P{GAL4-nos.NGT}40 males confirm that NGT-during Drosophila development. Notable by their
ab-sence has been the availability of GAL4 lines that drive driven expression is strictly maternally dependent. Thus, the increasing accumulation of UAS-lacZ mRNA expression during oogenesis and the earliest stages of
embryogenesis. It has been suggested that this is due to during the postblastoderm stages is due to perduring maternal GAL4 activity at either the mRNA and/or pro-the developmental regulation of factors that specifically
affect either the translation of the GAL4 mRNA or the tein level.
A sensitive photometric assay forb-galactosidase activ-activity of the GAL4 protein during these stages.
Figure1.—GAL4-dependent activation with a maternal mRNA. GAL4-dependent transcriptional regulation is achieved in early embryos that contain maternally expressed GAL4 mRNA. The nos promoter is used to drive expression of the GAL4 mRNA specifically during oogenesis. Inclusion of the 39untranslated region of a maternally expressed tubulin mRNA allows for uniform deposition of the GAL4 mRNA in the developing oocyte. GAL4-dependent transgenes can be inherited either from the male in the cross (as shown) or from the female.
allows for detection of UAS-lacZ expression in aliquots of lacZ.B} transgene, but also translation of the lacZ mRNA prior to the completion of cellularization. Calibration extracts prepared from single embryos (seematerials
and methods). To generate a developmental time experiments indicate that the level of activity detected per embryo at this stage is equivalent to that of 67 pg course, we performed assays on single embryos that were
staged based on the time at which they initiated gastrula- of b-galactosidase. This corresponds to z1.25 3 105 molecules ofb-galactosidase in each of the 6000 blasto-tion. This is a readily observed and rapid morphogenetic
process that occurs immediately upon completion of derm cells. As found for the mRNA, b-galactosidase enzyme levels continue to increase during the germ-cellularization. The activity detected in early gastrula
stage embryos is.100-fold greater than the background band extension stages (Table 1). Decreases in enzyme activity levels, which require degradation of both the levels detected in control crosses (Table 1). This enzyme
activity depends on not only transcription of the P{UAS- lacZ mRNA and protein, do not become apparent until
Figure 2.—Developmental time course of
TABLE 1
Developmental time course ofNGT-driven gene expression
b-Galactosidase
Stage activity
Maternal genotype Paternal genotype (hr)a (31023)b
NGT40/NGT40 UAS-lacZ/UAS-lacZ 3.5 201666
NGT40/NGT40 UAS-lacZ/UAS-lacZ 4.5 8646237
NGT40/NGT40 UAS-lacZ/UAS-lacZ 5.5 13656181
NGT40/NGT40 UAS-lacZ/UAS-lacZ 7.25 17766212
NGT40/NGT40 UAS-lacZ/UAS-lacZ 8.5 14796120
NGT40/NGT40 UAS-lacZ/UAS-lacZ 10.5 11766455
UAS-lacZ/UAS-lacZ NGT40/NGT40 7.25 2.160.3
y w/y w UAS-lacZ/UAS-lacZ 7.25 3.860.4
aEmbryos were selected at the onset of gastrulation based on appearance of the cephalic and ventral furrows. Embryos at this stage were defined as 3.5 hr old. Older embryos were obtained by aging them from the time of gastrulation.
bb-Galactosidase activity was measured on aliquots prepared from single staged embryos as described in
materials and methods.The numbers shown indicate the relative light units6SD for measurements made
on at least five embryos from each stage.
several hours later, when the embryos are in the process More severe patterning defects, including elimination of the entire head skeleton, are observed in embryos of germband retraction.
Unique phenotypes produced byNGT-driven expres- with NGT-driven runt expression (Figure 3C). The
mo-lecular basis of the phenotype produced by runt
overex-sion of different UAS transgenes: We examined the
effects of using the strong P{GAL4-nos.NGT}40 line to pression is discussed below.
We also obtained specific embryonic lethal pheno-drive expression of several different UAS transgenes in
the blastoderm embryo. Embryonic lethality was ob- types upon ectopic expression of lz and pnt, two genes that are most well characterized for their roles in pattern served for all but one of the different UAS lines tested.
Cuticle preparations of the inviable embryos indicate formation during eye development (Brunner et al. 1994;O’Neillet al. 1994;Dagaet al. 1996). NGT-driven that unique phenotypes are associated with NGT-driven
expression of each of these genes. Ectopic expression expression of lz causes a dorsal closure defect similar to that observed in embryos mutant for the “tail-up” of dpp produces embryos that lack ventral denticle belts
(Figure 3A). This resembles the phenotypes obtained class of recessive lethal mutants (Figure 3D;Frankand
Rushlow1996). This tail-up phenotype seems unlikely when dpp is ectopically expressed by other means in the
early embryo and is consistent with the role that dpp has to be relevant to a normal regulatory function, as lz shows only extremely limited expression during Dro-in dorsal-ventral patternDro-ing (FergusonandAnderson
1992;Staehling-Hamptonet al. 1994). Patterning de- sophila embryogenesis (S. G. KramerandJ. P. Gergen,
unpublished results). However, it is notable that this fects along the anterior-posterior axis are produced by
NGT-driven expression of the segmentation genes en phenotype is distinct from that produced by expression of runt (compare Figure 3, C and D). These two genes and runt. Uniform expression of the segment-polarity
gene en in this manner generates a reproducible pair- encode members of the Runt domain family of tran-scription factors. The differential response of the em-rule phenotype (Figure 3B). Similar effects have been
observed when en expression is induced by heat shock at bryo to these two related proteins provides a clear indi-cation of their functional specificity, presumably due to the onset of gastrulation (PooleandKornberg1988).
Figure3.—Embryonic lethal phenotypes
TABLE 2
Effects of expressing various gene products with the maternal GAL4 system
Relative viability of P{UAS-X} progeny (% female:% male)b
lacZ runt.T en.Y dpp.S Pnt.P1
NGT females activitya (232) (4-1) (42B.4) (3)
11 129648 100:100 50:20 80:100 100:100
31 307691 90:10 — 70:60 60:50
40 603678 10:,1 0:0 — —
11/11 7146302 10:0 0:0 10:,5 ,5:0
31/31 7806104 20:0 0:0 ,5:,5 0:0
40/40 17766212 0:0 0:0 0:0 —
ab-Galactosidase activity is measured on staged embryos (7.25 hr) using the luminescent Galacto-Light Plus substrate (Tropix). The numbers reported are31023and represent the average and standard error of measurements done with portions (1/10) of lysates prepared from.10 single embryos.
bRelative viability was determined by counting the number of UAS carrying adult progeny relative to their siblings that carry a balancer (or other marked chromosome). The relative viability of P{UAS-runt.T}232 was measured relative to the second chromosome balancer CyO. The P{UAS} transposons for en, dpp, and pnt[P1] are all on the third chromosome. Viability crosses with these transgenes involved TM3 or ruPrica. The data shown are reported to the nearest decile, except for crosses that produced,5% of the expected number of viable UAS-X progeny. These data are all from crosses that yielded at least 50, and typically .200, sibling progeny.
differential interactions with other factors in the Dro- that are homozygous for any particular NGT chromo-some show approximately twice the level of activity as sophila embryo. Our experiments with pnt provide
an-other example of a differential response, in this case heterozygous females (Table 2). The relative strengths of these different NGT genotypes correlate with the to protein isoforms produced by differential splicing.
Expression of the Pnt[P1] protein results in fully pene- lethality observed in crosses with different UAS transgenes (Table 2). Importantly, these data show that trant embryonic lethality with patterning defects in the
head skeleton (Figure 3E). In contrast, the P{UAS- for a number of the toxic UAS transgenes tested in our assays, the difference between no discernible effect on pnt.P2}2 transgene has no discernible effect on the
via-bility or phenotype of embryos from homozygous viability and fully penetrant embryonic lethality occurs over a range that involves only a 4- to 5-fold increase in P{GAL4-nos.NGT}40 females. The principal difference
in these two transgenes is an N-terminal exon that con- the level of NGT-driven expression.
We further characterized the basis of the lethality fers constitutive activity on the Pnt[P1] protein, whereas
Pnt[P2] is activated in response to receptor tyrosine obtained in crosses with UAS-runt. One observation of particular interest was a pronounced sex bias in lethality kinase signaling pathways (Brunneret al. 1994;O’Neill
et al. 1994). On the basis of these observations, we specu- that was not obtained with the other UAS transgenes. Males are consistently more sensitive than females to late that the defects caused by ectopic Pnt[P1]
expres-sion do not occur with Pnt[P2] because the level of the lethal effects of NGT-driven runt expression. One explanation for this sex-biased lethality is that NGT-receptor tyrosine kinase signaling activity within the cells
that are affected by Pnt[P1] is insufficient to activate driven runt expression leads to inappropriate activation of the Sex-lethal (Sxl ) gene in male embryos. This would Pnt[P2].
Quantitative effects of NGT-driven expression:The be consistent with runt’s role as a regulator of Sxl and
would both confirm and extend recent results indicat-above experiments are with P{GAL4-nos.NGT}40, one
of the stronger maternal GAL4 drivers. We also exam- ing that increased runt activity can result in transcrip-tional activation of Sxl in male embryos (Krameret al. ined the activities of other lines using similar assays.
The results for three representative lines, P{GAL4- 1999). However, we found no evidence that NGT-driven runt expression was capable of activating a Sxl[Pe]lacZ nos.NGT}11, P{GAL4-nos.NGT}31, and P{GAL4-nos.
NGT}40, are shown in Table 2. Quantitation of b-galac- reporter gene in male embryos (data not shown). Fur-thermore, the preferential sensitivity of males to UAS-tosidase activity allows for a direct measurement of the
relative levels of expression produced by these different runt expression is not suppressed in males hemizygous for the Sxl mutations Sxl[F#1] and Sxl[7BO] (data not lines. Even a single copy of P{GAL4-nos.NGT}11, the
weakest line characterized in these experiments, drives shown). If Sxl activation contributed to male lethality, then males carrying these loss-of-function mutations expression of lacZ at levels that are 100-fold over
Sxl activation does not account for the enhanced sensi- ity (Table 2). Embryos from this cross show a consistent repression of the odd-numbered stripes of the segment-tivity of males to UAS-runt expression.
An alternative explanation is that the activity of the polarity gene en (Figure 4F). In contrast, expression of the pair-rule genes eve and ftz is not altered in these P{UAS-runt.T} transgene is dosage compensated, i.e., it
is twice as active in males as in females. Using the results same embryos (Figure 4, G and H). Similarly, this level of ectopic runt activity has little to no effect on the of theb-galactosidase assays as a scale, we estimate that
males are approximately twice as sensitive as females to expression of the pair-rule genes hairy, paired, odd-skipped, and sloppy-paired (data not shown). This strongly the toxic effects of runt overexpression. For example,
male viability is reduced to 10% in crosses with heterozy- suggests that the repression of en is not mediated indi-rectly through alterations in the expression of these gous P{GAL4-nos.NGT}31 mothers. A comparable
re-duction in female viability is obtained in crosses either other pair-rule genes, and it provides evidence that runt directly represses the odd-numbered en stripes. As ex-with heterozygous P{GAL4-nos.NGT}40 mothers or ex-with
homozygous P{GAL4-nos.NGT}31 mothers. These two pected, some variation is also observed in the patterns of en expression. All embryos show defects in the initia-maternal genotypes are estimated to have twice the
activ-ity of the heterozygous P{GAL4-nos.NGT}31 mothers tion of the odd-numbered stripes during the early stages of germband extension. In later stage embryos, the ex-(Table 2). Similarly, fully penetrant lethality of the
P{UAS-runt.T} transgene in males is obtained at half the pression of the odd-numbered en stripes is partially or even fully restored. Approximately 10% of the embryos level of NGT activity that is required for fully penetrant
lethality in females (homozygous P{GAL4-nos.NGT}11 at full germband extension show apparently normal en expression. This corresponds well to the proportion of females vs. homozygous P{GAL4-nos.NGT}40 females,
respectively). These results agree extremely well with a progeny from this cross that will survive to adulthood. We also examined segmentation gene expression pat-model whereby the embryonic activities of the
P{UAS-runt.T} transgenes are dosage compensated. Previous terns in embryos from a cross of homozygous P{GAL4-nos.NGT}40 females with homozygous P{UAS-runt.T} work demonstrated that runt’s activity during
segmenta-tion is dosage compensated (Gergen1987). These re- U15 males. This combination is estimated to allow for an approximately sixfold higher level of ectopic expression sults presented here strongly suggest that control
ele-ments responsible for dosage compensation of runt at than that obtained in the above experiment and is equal to or greater than that obtained within the pair-rule this stage of development are contained within the
P{UAS-runt.T} transgene. stripes produced by the endogenous runt gene (Figure
4I). As observed at lower levels, the odd-numbered
Quantitative response of target genes to UAS-runt
during segmentation:The expression of genes in both stripes of en are repressed by this higher level of ectopic
runt expression (Figure 4J). The effects of high-level the pair-rule and segment-polarity classes of
segmenta-tion genes is altered in runt mutant embryos (Carroll NGT-driven runt expression on the pair-rule genes mimic what has been described previously in hs-runt and Scott 1986; DiNardo and O’Farrell 1987;
Inghamand Gergen 1988; Baumgartnerand Noll embryos. Expression of eve is reduced and there is a difference in the sensitivity of the different stripes, stripe 1990). The identification of the genes that are direct
targets for transcriptional regulation by the Runt pro- 2 being the most sensitive to repression by runt (Figure 4K). Similar stripe-specific repression of hairy stripe #1 tein is complicated by a number of cross-regulatory
in-teractions between these different genes. One approach is also observed in these embryos (data not shown). Expression of ftz is increased with the broader stripes that has been used extensively to investigate the
regula-tory circuitry in the segmentation pathway has involved fusing, especially in the more posterior regions (Figure 4L). These findings validate previous results with hs-runt ectopic expression using heat-inducible transgenes. For
example, runt’s roles in activating the pair-rule gene ftz embryos and indicate that these different regulatory interactions are not a result of the physiological pertur-and repressing the pair-rule genes eve pertur-and hairy have
been investigated using hs-runt transgenes (Tsai and bations associated with the heat-shock response. The localized effects of NGT-driven runt expression on the
Gergen1994, 1995). Additional experiments with these
hs-runt lines have also suggested that runt directly regu- pair-rule genes contrast with the relatively uniform re-pression of the odd-numbered en stripes. This provides lates segment-polarity gene expression (Manoukian
andKrause1993). a further indication that the repression of en is direct and not mediated by other pair-rule genes.
We examined the response of several segmentation genes to different levels of NGT-driven runt expression. To obtain insight on the basis for the lethality associated
DISCUSSION with NGT-driven runt expression, we examined embryos
from a cross of homozygous P{GAL4-nos.NGT}11 fe- A maternal GAL4 system: We have developed and characterized Drosophila strains that express the yeast males with homozygous P{UAS-runt.T} males. The level
viabil-Figure4.—Effects of maternal GAL4-driven runt expression on segmentation gene expression. The top row of embryos shows the normal mRNA expression patterns of runt (A), en (B), eve (C), and ftz (D) as revealed by whole-mount in situ hybridization. The alterations in expression of these same genes in embryos from a cross of homozygous P{GAL4-nos.NGT}11 females with homozygous P{UAS-runt.T}232 males are shown in E, F, G, and H, respectively. Similarly, I, J, K, and L show the respective expression of these four genes in embryos from a cross of homozygousn P{GAL4-nos.NGT}40 females with homozygous P{UAS-runt.T}U15 males. Embryos are oriented anterior to the left, dorsal side up. All embryos are in the blastoderm stage, except for those probed for en expression (B, F, and J), which have completed cellularization and have gastrulated. The circles in F and J indicate regions where the odd-numbered en stripes are absent. Similarly, the circle in K marks the absence of eve stripe 2.
for manipulating gene expression in Drosophila. Many to greatly reduce the postblastoderm expression by in-cluding elements from mRNAs that are turned over of the GAL4 lines that have been characterized
pre-viously are based on the initial expression constructs of rapidly during these stages in both the NGT- and UAS expression constructs. Incorporating the results of
fur-Brand andPerrimon(1993). A key difference in the
GAL4 expression construct described here is the use of ther studies on mRNA control elements should greatly improve the specificity of this type of strategy for manip-the 39 UTR region of the aTub84B mRNA in place of
the hsp70 terminator. The use of the tubulin 39 UTR ulating gene expression.
There are several attractive features of using mater-also distinguishes the NGT strains from GAL4 drivers
that use the 39UTRs of either bicoid (Arnostiet al. 1996) nally driven GAL4 expression to investigate gene func-tion. First, the large body of information on the genetics or nos (Van Doren et al. 1998) to deliver maternally
expressed transcripts specifically to the anterior or pos- and cell biology of the Drosophila embryo makes this a powerful developmental context for interpreting the terior pole of the embryo, respectively. The spatially
uniform activation mediated by the NGT drivers simpli- phenotypes produced by ectopic expression of any tar-get gene of interest. Experiments at this stage of devel-fies the quantitative interpretation of experimental
re-sults both in the entire embryo as well as on the cellular opment also avoid some of the difficulties that can arise with other GAL4 drivers that have low levels of expres-level.
The importance of mRNA control elements in GAL4 sion at stages or in tissues other than the desired devel-opmental context. With a maternal GAL4 driver, the misexpression strategies is also reemphasized by the
re-cent work of Rørth(1998). In this case it was found earliest stage at which a paternally inherited UAS transgene is available for activation by GAL4 protein is that GAL4-driven expression during oogenesis was
quali-tatively altered by including the 39untranslated region within the fertilized egg. Finally, the relative simplicity of the ectopic expression pattern makes it relatively of the maternally expressed K10 gene in place of the
SV40 polyadenylation signal in the UAS expression con- straightforward to interpret the experimental results quantitatively. The three NGT lines characterized here struct (Rørth1998). It seems likely that further
modi-fications of the 59and 39UTRs of both the GAL4 driver allow for control of expression over a range that differs
.10-fold between the weakest line as a heterozygote and UAS responder will allow for even greater control
over gene expression during oogenesis and early em- and the strongest line as a homozygote (Table 2). Using enzyme activity measurements as a benchmark, we esti-bryogenesis. It may be possible to increase blastoderm
stage expression by including the 39UTR from a mRNA mate this range to extend from z10,000 to 125,000 molecules of b-galactosidase/cell at the blastoderm that is efficiently translated during this stage in the UAS
lines. It is also possible to obtain higher expression levels the sex determination pathway is the Sxl gene, which normally is activated in females and repressed in males. by combining different NGT drivers. Altogether, with
the NGT lines that are currently available, we estimate runt plays a role in the transcriptional activation of Sxl, and increasing the dosage of runt activity is sufficient being able to manipulate expression levels over a range
that approaches two orders of magnitude. for triggering the inappropriate activation of Sxl in pre-blastoderm male embryos (Kramer et al. 1999). Inap-The expression levels obtained with the different NGT
drivers are presumed to be due to the sites of transgene propriate activation of Sxl could in principle account for the preferential male lethality observed with NGT-insertion. Similar position effects are, of course, also
observed for different inserts of any given UAS driven runt expression. However, male lethality is not suppressed in males that are mutant for Sxl. Further-transgene. Indeed, the ability to mix and match NGT
drivers with different UAS responders provides an addi- more, NGT-driven runt expression is not capable of acti-vating the full-length SxlPeembryonic promoter in males. tional level of flexibility that can be useful in the design
and interpretation of ectopic expression experiments. Presumably the levels of ectopic runt expression ob-tained during the syncytial blastoderm stage are not Our experiences with several different P{UAS-runt.T}
lines indicate a robust and linear response over the high enough to trigger inappropriate activation of the Sxl embryonic promoter in males. It remains to be deter-ranges tested. The phenotype observed in any given
cross depends on the level of runt expression that is mined whether the levels of NGT-driven UAS-runt ex-pression are sufficient for SxlPeactivation in females. obtained, irrespective of the particular combination of
NGT driver and UAS-runt responder that is used in the The preferential sensitivity of males to NGT-driven runt expression is readily explained if P{UAS-runt.T} cross.
There is an additional point to be made with respect activity is dosage compensated. A comparison of the NGT expression levels required to produce comparable to our attempts to quantitatively characterize this
ec-topic expression assay system. As indicated in the tables, effects on male and female viability is consistent with a twofold increase in UAS-runt potency in males. These the standard error in the measurements of NGT-driven
b-galactosidase activity averages z20% of the activity observations provide the basis for a model in which Sxl, which is specifically expressed in females, buffers the measured, irrespective of the total absolute enzyme
ac-tivity. These assays were carried out on individual em- embryos from the toxic effects of UAS-runt expression. In addition to its more widely known role in regulating bryos that were staged based on their time of
gastrula-tion, a readily observed and rapid morphogenetic splicing, the Sxl protein represses expression of msl-2 by interacting directly with the 59 and 39 UTR of the change that immediately follows the completion of
cel-lularization. Thus, the embryo-to-embryo variability is msl-2 mRNA (BashawandBaker1997;Gebaueret al. 1998). The runt 39 UTR contains several putative Sxl not likely to be due to differences in developmental
stage. Repeated measurements on extracts from differ- binding sites (Kelleyet al. 1995) which may mediate translational repression by the Sxl protein, and this en-ent single embryos further indicate that differences in
micropipeting do not account for this level of variability. tire region is contained within the P{UAS-runt.T} transgenes used in our experiments. In support of this Thus, the variability may be intrinsic within this
biologi-cal system. This variation somewhat complicates the util- hypothesis, UAS-runt transgenes that use the SV40 39 UTR instead of the runt 39 UTR show similar, though ity of this system for confident measurement of small
(,20%) changes in the level of gene function. However, reduced, levels of activity in both sexes (J. Wheeler, D. Swantek and J. P. Gergen, unpublished results). the ability to manipulate gene expression in a stepwise
manner over one to two orders of magnitude with a Further work is needed to confirm the mechanisms re-sponsible for the enhanced potency of UAS-runt reliability that approaches620% will provide an
impor-tant tool for quantitative analysis of gene function in transgenes in males. Indeed, the activity of UAS-runt transgenes may provide a useful tool for further investi-vivo. Indeed, the importance of quantitative
considera-tions is emphasized by the dose-dependent effects ob- gation of this mode of dosage compensation.
Dose-dependent effects ofrunton segmentation:The
tained with several different toxic UAS transgenes. In
each case we found a relatively sharp threshold in the Runt protein is a pivotal transcriptional regulator in the pathway of segmentation in Drosophila. Previous work biological response, with the difference between no
ap-parent phenotype and a nearly fully penetrant, lethal with heat-inducible hs-runt transgenes has indicated that Runt functions to activate the transcription of some phenotype resulting from a four- to fivefold increase in
the level of NGT-driven ectopic gene expression. downstream targets, such as ftz, while repressing the transcription of targets such as en, eve, and hairy (Tsai
Interactions betweenruntand the sex determination
pathway:Our results reveal that male embryos are repro- and Gergen 1994, 1995). Furthermore, there are at
least two mechanisms for transcriptional repression by ducibly more sensitive than females to the toxic effects
of UAS-runt expression. Previous work reveals there are the Runt protein. The stripe-specific repression of eve and hairy involves interactions with the corepressor pro-multiple interactions between runt and the sex
motif located at the C terminus of the Runt protein mutant phenotypes. Mutations with generally deleteri-ous effects would be expected to increase the severity (Aronson et al. 1997). In contrast, the repression of
the segment polarity gene en and the head gap gene of defects in a sensitized genetic screen based on en-hancement of loss-of-function phenotypes, but in a non-orthodenticle occurs through a VWRPY-independent
mechanism (Aronsonet al. 1997;Tsaiet al. 1998). The specific manner. However, these same mutations also would be expected to enhance, not suppress, the severity results obtained here using maternally provided GAL4
to drive ectopic runt expression confirm these previous of defects produced by GAL4-driven overexpression. From our quantitative analysis, we feel it should be possi-findings and indicate that these varied regulatory effects
are not an artifact due to overexpression by heat-shock ble to identify and characterize mutations that have a twofold effect on the potency of the Runt protein. In treatment. An important additional finding is that the
odd-numbered stripes of the segment-polarity gene en summary, the ability to quantitatively manipulate gene expression in the Drosophila blastoderm embryo should are efficiently repressed in embryos that display normal
pair-rule gene expression patterns. This is strong evi- provide a powerful new tool for genetic studies on the function of runt as well as any other regulatory gene dence that the repression of these en stripes is not
indi-rectly mediated through Runt’s regulatory effects on product that can function within this well-defined devel-opmental context.
the expression of these other pair-rule genes. These
results strongly suggest that the odd-numbered en stripes The excellent and invaluable technical assistance of Claudia Brun-are a direct target for repression by the Runt protein. ner and Deborah Swantek is greatly appreciated. Joe McLean provided the impetus and some assistance with cuticle preparations of embryos
What is the relevance of runt’s ability to repress the
carrying various UAS transgenes. This manuscript was improved by
odd-numbered en stripes for normal segmentation? The
comments from John Wheeler, Kathy Wojtas, and Christine
Vander-even- and odd-numbered en stripes are controlled by two Zwan. Recombinant DNA plasmids containing the nos promoter and distinct regulatory programs (DiNardoandO’Farrell theaTub84B 39UTR were generous gifts from Liz Gavis (Princeton,
1987;DiNardoet al. 1988). The even-numbered stripes NJ) and Bill Theurkauf (University of Massachusetts), respectively. The starting vectors for the GAL4 modular misexpression system,
emerge first and form in the center of the seven stripes
pGAT-B and pUAS-T, were provided by Andrea Brand (Welcome
of runt expression that are present during the
blasto-MRC Institute, Cambridge, United Kingdom). The P{UAS-lz} flies
derm stage. The odd-numbered en stripes emerge dur- were a gift from Utpal Banerjee (UCLA). Sxl mutations were obtained ing the process of cellularization and are not expressed from Tom Cline (Berkeley) and Jim Ericson (Columbia). Many of at levels comparable to the even-numbered stripes until the other Drosophila lines used in this work were obtained from the stock center in Bloomington. This work was supported by a National
after gastrulation (DiNardoandO’Farrell1987). The
Institutes of Health grant GM-53229 to J.P.G.
odd-numbered en stripes arise in the regions between the runt stripes and in fact appear to define the center of the runt interstripes. The runt stripe/interstripe pattern
develops from an earlier broad band of expression LITERATURE CITED
throughout the presegmental region of the embryo Al-Atia, G. R., P. FruscoloniandM. Jacobs-Lorena,1985 Trans-(KlinglerandGergen1993). The levels of NGT-driven lational regulation of mRNAs for ribosomal proteins during early
Drosophila development. Biochemistry 24: 5798–5803.
runt expression in the presumptive interstripe regions
Arnosti, D. N., S. Barolo, M. Levineand S. Small,1996 The
that are required for repression of the odd-numbered eve stripe 2 enhancer employs multiple modes of transcriptional en stripes are below the levels of expression within the synergy. Development 122: 205–214.
Aronson, B. D., A. L. Fisher, K. Blechman, M. CaudyandJ. P.
runt stripes (Figure 4, E and F) and are comparable to
Gergen,1997 Groucho-dependent and -independent
repres-the levels obtained during repres-the earlier broad band stage. sion activities of Runt domain proteins. Mol. Cell. Biol. 17: 5581– These observations suggest that clearance of Runt from 5587.
Bashaw, G. J.,andB. S. Baker,1997 The regulation of the
Drosoph-the interstripes is an essential prerequisite for Drosoph-the
forma-ila msl-2 gene reveals a function for Sex-lethal in translational
tion of the odd-numbered en stripes. control. Cell 89: 789–798.
Among the segmentation genes examined here, en is Baumgartner, S., and M. Noll, 1990 Network of interactions among pair-rule genes regulating paired expression during
pri-the most sensitive to ectopic runt expression. Indeed,
mordial segmentation of Drosophila. Mech. Dev. 33: 1–18.
the threshold response of en correlates extremely well Brand, A. H.,andN. Perrimon,1993 Targeted gene expression as a with the threshold for the lethality that is associated means of altering cell fates and generating dominant phenotypes.
Development 118: 401–415.
with NGT-driven runt expression. This lethal phenotype
Brunner, D., K. Ducker, N. Oellers, E. Hafen, H. Scholzet al., provides a powerful entry point for a genetic dissection
1994 The ETS domain protein pointed-P2 is a target of MAP
of Runt function. One approach is to screen for muta- kinase in the sevenless signal transduction pathway. Nature 370: 386–389.
tions that suppress the lethality associated with runt
over-Carroll, S. B.,andM. P. Scott,1986 Zygotically active genes that
expression during this early stage of embryogenesis.
affect the spatial expression of the fushi tarazu segmentation gene
There is an obvious practical advantage of genetic during early Drosophila embryogenesis. Cell 45: 113–126.
Daga, A., C. A. Karlovich, K. DumstreiandU. Banerjee,1996
screens that are based on suppression of a lethal
pheno-Patterning of cells in the Drosophila eye by Lozenge, which shares
type. There is also an important theoretical advantage
homologous domains with AML1. Genes Dev. 10: 1194–1205.
of this kind of approach compared to sensitized screens DiNardo, S., andP. H. O’Farrell, 1987 Establishment and
re-finement of segmental pattern in the Drosophila embryo: spatial
control of engrailed expression by pair-rule genes. Genes Dev. 1: Kelley, R. L., I. Solovyeva, L. M. Lyman, R. Richman, V. Solovyev
et al., 1995 Expression of msl-2 causes assembly of dosage com-1212–1225.
DiNardo, S., E. Sher, J. Heemskerk-Jongens, J. A. KassisandP. H. pensation regulators on the X chromosomes and female lethality in Drosophila. Cell 81: 867–877.
O’Farrell,1988 Two-tiered regulation of spatially patterned
engrailed gene expression during Drosophila embryogenesis. Na- Kim-Ha, J., K. Kerr and P. M. MacDonald, 1995 Translational regulation of oskar mRNA by bruno, an ovarian RNA-binding ture 332: 604–609.
Dong, J., L. H. Hung, R. StromeandH. M. Krause,1998 A phos- protein, is essential. Cell 81: 403–412.
Klingler, M.,andJ. P. Gergen,1993 Regulation of runt transcrip-phorylation site in the ftz homeodomain is required for activity.
EMBO J. 17: 2308–2318. tion by Drosophila segmentation genes. Mech. Dev. 43: 3–19.
Komori, T., H. Yagi, S. Nomura, A. Yamaguchi, K. Sasakiet al.,
Dormand, E. L.,andA. H. Brand,1998 Runt determines cell fates
in the Drosophila embryonic CNS. Development 125: 1659–1667. 1997 Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts
Ducy, P., R. Zhang, V. Geoffroy, A. L. RidallandG. Karsenty,
1997 Osf2/Cbfa1: a transcriptional activator of osteoblast differ- [see comments]. Cell 89: 755–764.
Kramer, S. G., T. M. Jinks, P. SchedlandJ. P. Gergen,1999 Direct entiation [see comments]. Cell 89: 747–754.
Duffy, J. B.,andJ. P. Gergen,1991 The Drosophila segmentation activation of Sex-lethal transcription by the Drosophila runt pro-tein. Development 126: 191–200.
gene runt acts as a position-specific numerator element necessary
for the uniform expression of the sex-determining gene Sex-lethal. Li, L.-H.,1999 Functional differences in the requirements for inter-actions of the Runt protein with the Brother proteins and DNA, Genes Dev. 5: 2176–2187.
Duffy, J. B., M. A. KaniaandJ. P. Gergen,1991 Expression and pp. 162 in Program in Cell and Developmental Biology. SUNY at Stony Brook, Stony Brook, NY.
function of the Drosophila gene runt in early stages of neural
development. Development 113: 1223–1230. MacDonald, P. M.,andG. Struhl,1988 cis-acting sequences re-sponsible for anterior localization of bicoid mRNA in Drosophila
Ephrussi, A.,andR. Lehmann,1992 Induction of germ cell
forma-tion by oskar [see comments]. Nature 358: 387–392. embryos. Nature 336: 595–598.
Manoukian, A. S.,andH. M. Krause,1992
Concentration-depen-Ferguson, E. L.,andK. V. Anderson,1992 Decapentaplegic acts as
a morphogen to organize dorsal-ventral pattern in the Drosophila dent activities of the even-skipped protein in Drosophila embryos. Genes Dev. 6: 1740–1751.
embryo. Cell 71: 451–461.
Fischer, J. A., E. Giniger, T. ManiatisandM. Ptashne,1988 GAL4 Manoukian, A. S.,andH. M. Krause,1993 Control of segmental asymmetry in Drosophila embryos. Development 118: 785–796. activates transcription in Drosophila. Nature 332: 853–856.
Fitzpatrick, V. D., A. Percival-Smith, C. J. Ingles and H. M. Manseau, L., A. Baradaran, D. Brower, A. Budhu, F. Elefantet al., 1997 GAL4 enhancer traps expressed in the embryo, larval
Krause,1992 Homeodomain-independent activity of the fushi
tarazu polypeptide in Drosophila embryos. Nature 356: 610–612. brain, imaginal discs, and ovary of Drosophila. Dev. Dyn. 209: 310–322.
Flores, G. V., A. Daga, H. R. KalhorandU. Banerjee,1998
Loz-enge is expressed in pluripotent precursor cells and patterns Markussen, F. H., A. M. Michon, W. BreitwieserandA. Ephrussi,
1995 Translational control of oskar generates short OSK, the multiple cell types in the Drosophila eye through the control of
cell-specific transcription factors. Development 125: 3681–3687. isoform that induces pole plasma assembly. Development 121: 3723–3732.
FlyBase,1999 The FlyBase database of the Drosophila Genome
Projects and community literature. The FlyBase Consortium. Nu- Matthews, K. A., D. F. MillerandT. C. Kaufman,1989 Develop-mental distribution of RNA and protein products of the Drosoph-cleic Acids Res. 27: 85–88.
Forbes, A.,andR. Lehmann,1998 Nanos and Pumilio have critical ila alpha-tubulin gene family. Dev. Biol. 132: 45–61.
Morrissey, D., D. Askew, L. RajandM. Weir, 1991 Functional roles in the development and function of Drosophila germline
stem cells. Development 125: 679–690. dissection of the paired segmentation gene in Drosophila embryos. Genes Dev. 5: 1684–1696.
Frank, L. H.,andC. Rushlow,1996 A group of genes required
for maintenance of the amnioserosa tissue in Drosophila. Devel- Nu¨ sslein-Volhard, C.,andE. Wieschaus,1980 Mutations affect-ing segment number and polarity in Drosophila. Nature 287: opment 122: 1343–1352.
Gavis, E. R.,andR. Lehmann, 1994 Translational regulation of 795–801.
Okuda, T., J. van Deursen, S. W. Hiebert, G. GrosveldandJ. R.
nanos by RNA localization. Nature 369: 315–318.
Gebauer, F., L. Merendino, M. W. HentzeandJ. Valcarcel,1998 Downing, 1996 AML1, the target of multiple chromosomal
translocations in human leukemia, is essential for normal fetal The Drosophila splicing regulator Sex-lethal directly inhibits
translation of male-specific-lethal 2 mRNA. RNA 4: 142–150. liver hematopoiesis. Cell 84: 321–330.
O’Neill, E. M., I. Rebay, R. TjianandG. M. Rubin, 1994 The
Gergen, J. P.,1987 Dosage compensation in Drosophila: evidence
that daughterless and Sex-lethal control X-chromosome activity at activities of two Ets-related transcription factors required for Dro-sophila eye development are modulated by the Ras/MAPK path-the blastoderm stage of embryogenesis. Genetics 117: 477–485.
Gergen, J. P.,andE. Wieschaus,1986 Dosage requirements for way. Cell 78: 137–147.
Otto, F., A. P. Thornell, T. Crompton, A. Denzel, K. C. Gilmour
runt in the segmentation of Drosophila embryos. Cell 45: 289–299.
Gupta, B. P., G. V. Flores, U. BanerjeeandV. Rodrigues,1998 et al., 1997 Cbfa1, a candidate gene for cleidocranial dysplasia
syndrome, is essential for osteoblast differentiation and bone Patterning an epidermal field: Drosophila lozenge, a member of
the AML-1/Runt family of transcription factors, specifies olfactory development [see comments]. Cell 89: 765–771.
Poole, S. J.,andT. B. Kornberg,1988 Modifying expression of sense organ type in a dose-dependent manner. Dev. Biol. 203:
400–411. the engrailed gene of Drosophila melanogaster. Development 104: 85–93.
Gustafson, K.,andG. L. Boulianne,1996 Distinct expression
pat-terns detected within individual tissues by the GAL4 enhancer Robertson, H. M., C. R. Preston, R. W. Phillis, D. M. Johnson-Schlitz, W. K. Benzet al., 1988 A stable genomic source of P trap technique. Genome 39: 174–182.
Ingham, P.,andP. Gergen, 1988 Interactions between the pair- element transposase in Drosophila melanogaster. Genetics 118: 461–470.
rule genes runt, hairy, even-skipped and fushi tarazu and the
estab-lishment of periodic pattern in the Drosophila embryo. Develop- Rørth, P.,1998 Gal4 in the Drosophila female germline. Mech. Dev. 78: 113–118.
ment 104 (Suppl.): 51–60.
Ish-Horowicz, D.,andS. M. Pinchin,1987 Pattern abnormalities Salles, F. J., M. E. Lieberfarb, C. Wreden, J. P. GergenandS. Strickland,1994 Coordinate initiation of Drosophila develop-induced by ectopic expression of the Drosophila gene hairy are
associated with repression of ftz transcription. Cell 51: 405–415. ment by regulated polyadenylation of maternal messenger RNAs. Science 266: 1996–1999.
John, A., S. T. SmithandJ. B. Jaynes,1995 Inserting the Ftz
homeo-domain into engrailed creates a dominant transcriptional repres- Staehling-Hampton, K., P. D. Jackson, M. J. Clark, A. H. Brand
andF. M. Hoffmann,1994 Specificity of bone morphogenetic sor that specifically turns off Ftz target genes in vivo. Development
121:1801–1813. protein-related factors: cell fate and gene expression changes in Drosophila embryos induced by decapentaplegic but not 60A. Cell
Kagoshima, H., K. Shigesada, M. Satake, Y. Ito, H. Miyoshiet al.,
1993 The Runt domain identifies a new family of heteromeric Growth Differ. 5: 585–593.
or indiscriminate expression of the Drosophila segmentation ila fushi tarazu gene: a nuclear receptor response element medi-ates the opposing regulatory effects of runt and hairy. Develop-gene ftz. Nature 318: 677–680.
Theurkauf, W. E., H. Baum, J. BoandP. C. Wensink,1986 Tissue- ment 121: 453–462.
Tsai, C. C., S. G. KramerandJ. P. Gergen,1998 Pair-rule gene specific and constitutive alpha-tubulin genes of Drosophila
mela-nogaster code for structurally distinct proteins. Proc. Natl. Acad. runt restricts orthodenticle expression to the presumptive head of
the Drosophila embryo. Dev. Genet. 23: 35–44. Sci. USA 83: 8477–8481.
Thummel, C. S., A. M. BouletandH. D. Lipshitz,1988 Vectors Van Doren, M., A. L. WilliamsonandR. Lehmann,1998 Regula-tion of zygotic gene expression in Drosophila primordial germ for Drosophila P-element-mediated transformation and tissue
cul-ture transfection. Gene 74: 445–456. cells. Curr. Biol. 8: 243–246.
Wang, C., L. K. DickinsonandR. Lehmann,1994 Genetics of nanos
Torres, M.,andL. Sanchez,1992 The segmentation gene runt is
needed to activate Sex-lethal, a gene that controls sex determina- localization in Drosophila. Dev. Dyn. 199: 103–115.
tion and dosage compensation in Drosophila. Genet. Res. 59: Wang, Q., T. Stacy, M. Binder, M. Marin-Padilla, A. H. Sharpe
189–198. et al., 1996 Disruption of the Cbfa2 gene causes necrosis and
Tracey, W. D., Jr., M. E. Pepling, M. E. Horb, G. H. Thomsen hemorrhaging in the central nervous system and blocks definitive andJ. P. Gergen,1998 A Xenopus homologue of aml-1 reveals hematopoiesis. Proc. Natl. Acad. Sci. USA 93: 3444–3449. unexpected patterning mechanisms leading to the formation of Yang, M. Y., J. D. Armstrong, I. Vilinsky, N. J. StrausfeldandK.
embryonic blood. Development 125: 1371–1380. Kaiser,1995 Subdivision of the Drosophila mushroom bodies
Tsai, C.,andJ. P. Gergen,1994 Gap gene properties of the pair- by enhancer-trap expression patterns [see comments]. Neuron rule gene runt during Drosophila segmentation. Development 15:45–54.
120:1671–1683.