Copyright 0 1988 by the Genetics Society of America
Chain-Bias
of
Escherichia coli
Rec-Mediated
X
Patch Recombinants Is
Independent of the Orientation
of
X
cos
Susan
M.
Rosenberg’
Institute of Molecular Biology, University of Oregon, Eugene, Oregon 97403
Manuscript received October 2, 1987
Revised copy accepted April 28, 1988
ABSTRACT
Chi is a hotspot for homologous recombination mediated by the RecBCD (Rec) pathway of
Escherichia coli. For Rec-mediated recombination of phage X, the orientation of X cos in the X
chromosome dictates the direction of travel of RecBCD enzyme through DNA and dictates which orientation of Chi or Chi-like sequences will be active in stimulating recombination. I previously found that Rec-mediated X patch heteroduplexes, stimulated by Chi or not, are chain-biased; at the X
P locus, recombinant information resides on the X r chain. This bias exists in the presence or absence of Chi sites. Reported herein is the finding that r-chain-bias at the P locus is independent of the orientation of X cos and thus also independent of the orientation of active Chi’s or Chi-like sequences and of the direction of travel of RecBCD enzyme. These results disprove previously elaborated models in which a chain-specific nick at Chi initiates recombination, and imply that some other chain- distinguishing process is involved with recombination. Replication and transcription are candidates for such a process.
B
Y definition, generalized genetic recombination can occur at any region along homologous chro- mosomes. However, recombination does not occur at all regions of homology with equal probability. Special sites called “recombinators” stimulate homologous recombination in their locales [see STAHL (1979) for review]. Crossover botspot instigator, Chi [5’GCTGGTGG (or its complement or both)] is a recombinator in the Escherichia coli wild-type RecBCD (Rec) pathway for homologous recombination (LAM etal. 1974; STAHL and STAHL 1977) [SMITH et al. (1 98 1) for sequence]. A single Chi sequence confers to phage
X a five- to tenfold higher rate of recombination than is experienced by X lacking Chi (X Chi’; LAM et al.
1974). T h e elevated Rec-mediated recombination is manifested in the vicinity of Chi (LAM et al. 1974; STAHL, CRASEMANN and STAHL 1975). Recombinators presumably facilitate a rate-limiting step in recombi- nation. Understanding Chi function will likely illumi- nate key features of the molecular mechanism of Rec- mediated recombination.
Rec-mediated, Chi-stimulated recombination is studied with X phage lacking the X recombination functions red and gam. T h e E. coli gene products RecA and RecBCD are essential for the process [STAHL and STAHL (1977); see BIEK and COHEN (1986) and AMUNDSEN et al. (1986) for the “new” recD gene and D. S. THALER, E. SAMPSON, I. SIDDIQI, S. M. ROSEN- BERG, M. M. STAHL and F. W. STAHL (in preparation) and THALER et al. (1988) for a view of RecD func-
of Medicine, 50 North Medical Drive, Salt Lake City, Utah 84132. Genetics 119: 7-21 (September, 1988)
’
Present address: Department of Biochemistry, University of Utah Schooltion]. Only the stages of recombination in which RecBCD enzyme finds the Chi site currently seem clear. These are controlled by the X packaging origin,
X cos dictates when and which Chi’s will be active by acting as an asymmetric entry site for RecBCD enzyme [KOBAYASHI, STAHL and STAHL (1984); see STAHL (1986) and SMITH and STAHL (1985) for re- views of supporting experiments]. When cleaved at
cos by the X packaging enzyme terminase (ter), X is linearized. One end of the X chromosome remains bound by ter [the “left” end (on the conventional X
map) of X carrying wild-type cos]. The other end is free for loading of RecBCD which enters DNA at a duplex end and travels unidirectionally through the DNA. In X carrying wild-type cos, RecBCD enters the free X right end and travels leftward. Leftward trav- eling RecBCD can recognize only Chi sequences ori- ented “leftward” (or “frontward,” 5’GCTGGTGGS’);
i.e., the enzyme recognizes Chi approached from the 3’ side of GG3’. RecBCD interacts directly with a properly oriented Chi, resulting in the (observed) stimulation of recombination at Chi and downstream from Chi in RecBCD’s travel path, past Chi’s 5’GC end. If cos is inverted in the X chromosome, RecBCD travels rightward and recognizes only “rightward” (or inverted) Chi’s (3’GGTGGTCG5’). RecBCD is blind to Chi’s oriented incorrectly with respect to its direc- tion of travel.
Contact of RecBCD with Chi results, by an un- known mechanism, in local elevation of recombina- tion. Currently pertinent questions include the follow- ing. (1) What is the rate-limiting step facilitated by
8 S. M. Rosenberg
Chi? Recombination can be imagined to proceed through stages such as those below:
PARENTAL MOLECULES
3-
INITIATION INTERMEDIATE1
RESOLUTION PRODUCTS.If the reactions flow more or less as drawn, does RecBCD-Chi interaction promote initiation of the in- termediate or the intermediate's resolution to prod- ucts? (2) Does RecBCD alter DNA at Chi to make it a more recombinogenic substrate? Or, does Chi change RecBCD into a more potent recombinase, one that acts more efficiently after contact with (at and down- stream from) Chi? Described herein are experiments addressed to specific molecular models elaborated from subsets of these general possibilities.
Beside the polarity/directionality imposed on Rec- mediated X recombination by X cos, two other relevant phenomena exhibit polarities: first, in vitro nicking of the Chi site by RecBCD is chain-specific. Under cer- tain in vitro conditions, RecBCD enzyme loses much of its exonuclease (ExoV) activity and travels through duplex DNA which it enters at a double-chain end
(ROSAMOND, TELANDER and LYNN 1979; TAYLOR and
SMITH 1980). Under such conditions the enzyme can
nick Chi sequences on one chain, that written 5' GCTGGTGG3'; the nick occurs just 3' of the termi- nal GG3' (PONTICELLI et al. 1985). This chain-polar nicking in vitro is subject to the right-left polarity described above. Leftward traveling RecBCD nicks only "leftward" (frontward) Chi's (5'GCTGGTGGS') on the chain shown, which corresponds to the X 1 chain, the chain ending 3' at the X right end. Right- ward traveling RecBCD nicks only "rightward" (in- verted) Chi's on the chain written 3'GGTGGTCG5' which corresponds to the X r chain, the chain ending 5' at the X right end (TAYLOR et al. 1985). Therefore, chain-polarity of nicking is dictated by the right-left polarity of RecBCD travel direction. This orientation-
dependence implies that RecBCD and Chi make direct
contact in vitro as was indicated in vivo (KOBAYASHI et
al. 1982; SCHULTZ, TAYLOR and SMITH 1983; CHAU-
DHURY and SMITH 1984). The orientationdependent
choice of which chain is nicked will be important in consideration of models, below.
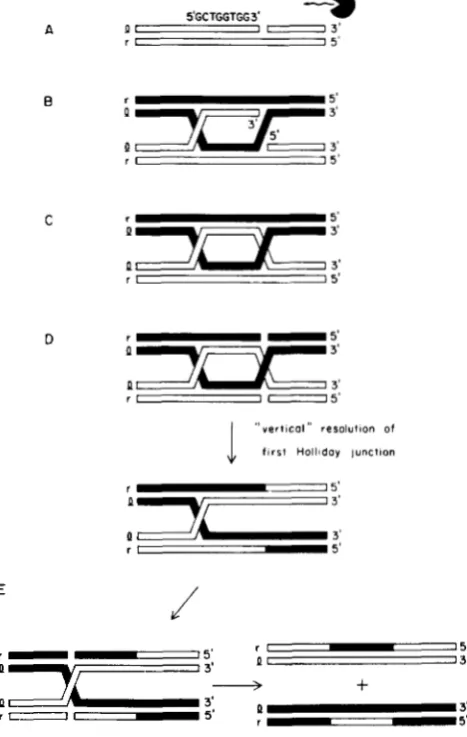
Does the chain-specific nick play a role in recombi- nation in vivo? In the course of testing an economical model in which such a nick at Chi initiates recombi- nation (nick-at-Chi initiation model, Figure 1, from SMITH et al. 1984) I discovered a second chain-polar
phenomenon: that heteroduplex Rec-mediated re-
combinants not crossed over for flanking markers (patches) are chain-biased. The simple nick-at-Chi ini- tiation model (Figure 1) predicted that Chi-stimulated patches would be heteroduplex with recombinant in- formation exclusively on the X 1 chain (ROSENBERG
S'GCTGGTGG
6
n
-
-3;I I5
B r 5 '
Q -fy- L
:.LT
r P L " L ! ! ~ z~ z1 n 5 '
P
2
-;;
A
C r 5'
p-r -3'
r I - --
-resolutton of
Hollldoy lunctton
D
r 5 ' I
(1 I ~ """m 3' . ~ I
,,I..
D O t C h s p l l c e
FIGURE 1 .--Simple nick-at-Chi initiation model of S wet 01. (1984). (A) Chi is nicked at the location determined in vitro (3' to
the terminal GG3') on the chain corresponding to the XI chain. (B) The 3' end left at the nick invades a homologous duplex forcing out a singlechain D-loop which pairs with the free singlechain region of the initiator molecule. (C) The Dloop is clipped to give a classic HOLLIDAY junction intermediate (HOLLJDAY 1964). which can be resolved in either of two "senses" to yield splices (crossovers)
or patches. This model predicted that patches would be heterodu- plex with recombinant information on the X 1 chain and was previ- ously ruled out by the demonstration that patches are strongly biased to the X r chain (ROSENBERC 1987).
1987). Contrary to this prediction, heteroduplex patches at the X P locus, stimulated by Chi (Chi+) or
not (Chi'), have recombinant information strongly biased to the X r chain; at least 90% of all heteroduplex
patches at P are r-chained (ROSENBERG 1987). Thus
the specific model in Figure 1 is incorrect.
The fact that patches at P are r-chain-biased whether stimulated by Chi or not implies that the mechanism of Rec-mediated recombination is identi- cal in both cases, that Chi merely enhances the rate of recombination. This view is also implied by the observation that Chi stimulates patches and splices equally (compared with their ratio in Chi' crosses, STAHL et al. 1982) and by studies of Chi-like se- quences, sequences that differ from Chi by one nu- cleotide pair. Three of such sequences tested have partial Chi-activity, stimulating recombination from
6% to 38% as strongly as Chi (CHENG and SMITH
1984). The ability to stimulate recombination in vivo,
cos Orientation and Patch Chain-Bias 9
the magnitude of X Rec-mediated Chi' recombination is accounted for by the Chi-like sequences present. When cos is wild-type only frontward Chi-like se- quences could act; when cos is inverted only inverted Chi-like sequences could be active. But, by what mech- anism? How do Chi's and Chi-like sequences stimulate recombination?
This report bears on three issues: (1) in the previous examination of heteroduplex patch structure (ROSEN-
BERG 1987) the patches were isolated [as in STAHL,
LIEB and STAHL (1 984)] using X by X crosses in which packageable dimers were generated by the Int system
of phage X. Among Int-mediated recombinants, Rec- mediated patches at the locus P were examined. Here, the previous results are generalized using a different experimental set-up: phage X by Xdv plasmid crosses reveal r-chain-bias of patches at the X P locus. (2)
Previously, heteroduplex patch structure was deter- mined in crosses with and without Chi (ROSENBERG
1987), both of which yielded r-chain-biased patches at P. In the Chi+ crosses, one parent bore Chi and the non-Chi-containing patch recombinant (patch in trans
to Chi) was assayed. Here, previous results are ex- tended by the observation that patches at P in cis to Chi are also r-chain-biased. (3) Because the chain- polarity of nicking Chi's or Chi-like sequences in vitro is determined by the direction of travel of RecBCD down the DNA helix (TAYLOR et al. 1985; CHENG and SMITH 1987) which is controlled in vivo by cos
(Ko-
BAYASHI, STAHL and STAHL 1984; YAGIL and SHTRO-
MAS 1985; STAHL et al. 1986), one can test a second
class of nick-at-Chi initiation models not previously tested. In the simple nick-at-Chi initiation model a nick on the I chain gave rise to I-chain patches; there- fore that model was ruled out by the finding of r- chain patches at X
P
(Figure 1; ROSENBERG 1987). However, models could be imagined in which a nick on the X I chain at Chi (or a Chi-like sequence) initiates recombination which then proceeds through a more complicated set of steps that causes patches to occur on the opposite chain, the X r chain [for example, see Figure 2 (from ROSENBERG 1987)l. Models in which a nick on one chain dictates patches biased to the other predict that switching the chain that is nicked (nicking r instead of I ) should give rise to patches with switched chain-polarity [ I instead of r at XP (for
example see Figure 3)]. This class of models is tested by examining chain-polarity of patches from crosses in which cos is inverted in the X chromosome. I find that unlike chain-polarity of nicking in vitro, r-chain- bias of patches at P is independent of the orientation of cos. I conclude that patch chain-bias is imposed on recombination by some chain-discriminating element other than chain-specific nicking at Chi or Chi-like sequences, the orientation of cos or the direction of travel of RecBCD enzyme. I previously proposed rep- lication and transcription as candidates for such anSGCTGGTGG3'
A i l l - 3 '
r I I 5'
C r
P
I
n 3'5'
P A3'
r I 1 5'
vertical" resoIutjon of
f l r n l Hollldoy juncllon
E
k /
r " 5
' 1 5 I 3' '
TL-"
3'<+
P
r " 5
' f
-
3 ' 5'FIGURE 2.-Alternative nick-at-Chi initiation model. In an effort to generate r-chain-biased patches using a nick such as the nick at Chi seen in vitro, this model (ROSENBERG 1987) adds two extra assumptions to the simple nick-at-Chi initiation model. (A) When RecBCD enzyme (notched circle) enters the X right end and travels leftward, a normally oriented Chi is recognized and nicked 3' to the terminal GG on the X 1 chain. (B) 3' invasion with the X E chain into a homolog. (C) Let the 3' end that invaded be religated with its 5"ended former neighbor (extra assumption No. 1). (D) Topo-
logical rule for resolution of HOLLIDAY junctions (extra assumption No. 2): whichever junction is resolved first, let it be resolved by cutting outside chains, "vertically." (E) Resolution of the second junction in the same Sense as the first gives patches, which are
heteroduplex with recombinant information on the X r chain.
(Resolution of the second junction in the opposite sense [imagined
to be equally likely since the single HOLLIDAY junction experiences no topological constraint] yields splices.)
element (ROSENBERG 1987). Those models are sup- ported and are further elaborated herein.
MATERIALS AND METHODS
a-
S. M. Rosenberg
TABLE 1
A Q r 1 3' Bacterial strains
Strain Relevant properties Reference
7 0 - 5 '
3 GGTGGTCG
B Q l 13'
r
II 3'
C
R 3'
Q
Q
:=:
3'E
/
kL
P - 1 ' 3 '
r I
'-
5
-
3:5' '
FIGURE 3.-Alternative nick-at-Chi initiation model with cos in- verted. When cos is inverted, RecBCD (notched circle) enters at the
X left end and travels rightward. Rightward traveling RecBCD is blind to normally oriented Chi but will recognize a backwards Chi (A) and, in this model, singlethain nick 3' to the Chi terminal GG3', now on the X r chain. Follow the same rules as in Figure 2 (B), (C), (D), (E). When the initial nick is on the X r chain, the patch products generated through this model are heteroduplex with recombinant information biased to the X 1 chain.
pKC3l [described in THALER, STAHL and STAHL (1987)l was kindly supplied by ROGER MCMACKEN.
X Genetic elements: X Genetic elements are described in Table 2. All were combined using standard methods (ARBER
et al. 1983). MARY STAHL kindly supplied the three X strains (from STAHL et al. 1982) that were crossed with Xdv. In all stocks of phage carrying cos2 cosML, both mutations were verified by the methods of STAHL et al. (1 982).
Preparation of artificial heteroduplex phage: X DNA chains were separated as in SZYBALSKI et al. (1971) and reannealed by the method of MESELSON and YUAN (1968). Annealed mixtures were packaged in vitro by the method of ROSENBERG et al. (1985; ROSENBERG 1985) in extracts of strain SMR34 (ROSENBERG 1987). Phage were assayed on SMR88 cells. "Artificial" heteroduplex phage will refer to those whose heteroduplex state was created in vitro as de- scribed here rather than "natural" heteroduplexes created by recombination in E . coli.
X by X dv crosses: Xdv-carrying SMR86 cells were grown
t o 1.5 x 10' per ml in tryptone broth supplemented with 1% yeast extract, 0.2% maltose, 0.01 mg/ml vitamin B1 and
534 C600 V66(78)
SMR20 SMR34
SMR40
SMR45
SMR48
SMR86
SMR88
Su- rec+ Su+ rec+
recD1009, allows X b1453 to make large plaques groP mutL216::TnlO
cos- mutL in vitro packaging lysogen, does not EcoK re- strict or modify, is mis- match repair-defective, produces no endogenous phage
Su- recBC sbcB mut- L216::TnIO, recBC sbcB al-
lows X b1453 to make large plaques
Su- recBC sbcB mutL2l I::Tn5 hsdRK"K+
groP mutL21I::TnZ
594 A(hsdSMK) mut- hsdRK-MK+ thr::TnlO
L216::TnlO dnaBts22 malB::Tn9 [pKC31], pKC3 1 is a Xdv, A(hsdSMK) confers EcoK restriction- and modificationdeficiency C600 recD1009 A(hsdSMK)
WEIGLE (1 966)
APPLEYARD (1954)
AMUNDSEN et al.
ROSENBERG (1 987) ROSENBERG (1 987)
(1 986)
ROSENBERG (1987)
ROSENBERG (1 987)
ROSENBERC (1 987)
This work
This work
TABLE 2
X Genetic elements
X genetic
element Relevant properties
cos2 22 bp deletion of cohesive ends leaving cos unusable for termi- nase cleavage or packaging cosML Leftward (inverted) cos cloned
into the X b region b1453 Deletion from att rightward, re-
moves red and gam
imm" Substitution of phage 21 immu- nity region into X, allows X to
g r o w well in cells harboring Xdv
tion gene; can be selected posi- tively in certain host strains
(GEORGOPOULOS and HERS-
KOWITZ 1971)
and Q
Pam80 Amber mutation in P , a replica-
x+D Leftward acting Chi, right of P
Reference KOBAYASHI et al.
(1 982)
KOBAYASHI et al. (1 982)
HENDERSON and
WEIL (1975) CAMPBELL (1 97 1)
CAMPBELL ( 196 1)
STAHL, CRASEMAN
and STAHL (1975)
cos Orientation and Patch Chain-Bias 11
Density transfer experiments done in parallel with the crosses and also those in Figure 5 were performed as in
STAHL et al. (1972) using the one-step-growth procedure outlined above. The density-labeled phage (prepared as in
STAHL et al. 1972) did not contain the b1453 deletion, so
splice-recombinants of plasmid with phage generate X chro- mosomes too large to be packaged. (X chromosomes larger than approximately 105% wild-type X are not packageable, see FEISS and BECKER 1983 for review).
Heteroduplex selections: X of the three different genetic backgrounds used here form plaques with different efficien- cies on selections I and 11. Consequently, the selections were modified slightly from ROSENBERG (1987; more discussion on the selections therein) to reduce plaque formation on the nonpermissive bacteria (“leakage”) and enhance proper X
heteroduplex plating efficiency for each background. Ho-
moduplex isogenic X P + and X Pam80 as well as both con- jugate artificial heteroduplex phage were plated in parallel
with the heteroduplex selections to verify that the selections work. For all three X backgrounds, plates for selection I
were incubated at 40” for 2 days before counting. Plates were not overlayed with drug agar. Selection I1 was exactly as reported before (ROSENBERG 1987) for the Chi’ back- ground and the cos2 cosML background. Plates for selection
I1 were incubated at 40” for the Chi+ D background and counted after one day of incubation. Preparation of 6-hr cultures of SMR20, SMR40, SMR45 and SMR48 was as described in ROSENBERG (1 987).
RESULTS
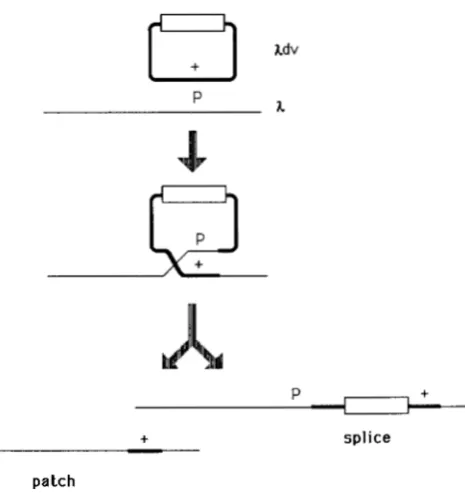
Experimental strategy: In this report, the chain- polarity of heteroduplexes is examined in Rec-me- diated X patch recombinants. T h e operational defini- tion of a patch used here is a recombination event that transfers genetic information from the Xdv plas- mid to X without accompanying crossover (splice) of the plasmid into the phage (see Figure 4). Xdv pIasmids are small, circular, autonomously replicating deriva- tives of X containing the X replication ori and X 0 and P genes (BERG 1974). T h e Xdv employed here, pKC31, was constructed by ligation of a HindIII- BamHI fragment of X, including the X cro, cZZ, 0 and P genes, with a kanamycin drug-resistance element derived from T n 5 a n d is roughly 6.2 kilobase-pairs (kb) long [the plasmid which was constructed by R. N. RAO and supplied by R. MCMACKEN, is similar to those in WOLD et al. (1 982) and was used by THALER, STAHL and STAHL (1987)l. A reciprocal crossover (splice) of plasmid (circle) into X incorporates the plasmid into the X chromosome, adding 6.2 kb of DNA to that phage chromosome (Figure 4). Patches of genetic information from plasmid to X generate recombinant
X whose chromosomes are no longer than those of the parental phage but which have picked up a marker from the plasmid (Figure 4). KELLENBERG-GUJER (1 97 I), BERG (1 97 I), STAHL et
d .
(1 982) and THALER, STAHL and STAHL (1987) used equilibrium density gradient centrifugation to separate progeny of X by Xdv crosses into these two size classes. From the frac- tions containing parental-sized progeny, they identi- fied patches at the loci 0 and P genetically. Here, heteroduplex chain-polarity of Rec-mediated patchesP
I +
+ splice
patch
FIGURE 4.-Recombination of the plasmid Xdv with X. Patches transfer genetic information from plasmid to X without adding extra length to the X chromosome. Splices (crossovers) of plasmid into X add an extra 6.2 kb of DNA (for the Xdv used here) to the X
chromosome. The white box represents a DNA segment not ho- mologous with the X (a kanamycin-resistance element) that is part of the Xdv. See Figures 6-8 for cesium formate equilibrium density gradients in which these two size classes are separated.
at the P locus is examined. T h e chain bearing recom- binant information is identified. T h e plasmid carries the P + allele. T h e phage used carry Pam80. From fractions containing parental-sized phage, heterodu-
plex I, 1-3’, and 11, 1-3’, phage were geneti-
tally selected using slight modifications of my previous method (ROSENBERG 1987).
Replication and mismatch repair were inhibited to help preserve heteroduplexes created by recombina- tion. T h e crosses of X by Xdv were performed in cells carrying the mutL mutation. mutL inactivates mis- match repair by the E. coli DNA adenine methyl- directed system (PUKILLA et al. 1983; MESEISON 1987) and the very short patch repair system (LIEB 1985) acting on Pam8O/P+ heteroduplexes (Fox and RAPOSA
1983; MUELSON 1987). In mut+ cells, mismatch repair acts on Rec-mediated heteroduplex patch recombi- nants to generate homoduplex patches (ROSENBERG
1987). Experiments reported here were conducted prior to the finding that full replication does not affect the frequency of heteroduplex patches in Rec-me- diated X by Xdv crosses (S. M. ROSENBERG unpublished data) under what seemed, a priori, the most “cautious” conditions. Allowing full replication during Rec-me- diated X by X crosses also does not affect the frequency of heteroduplexes among progeny (ROSENBERG 1987). Presumably, the heteroduplex recombinants have no opportunity to replicate before encapsidation
r L 5 ’ r L 5
’
12 S. M. Rosenberg
because of the close coupling of packaging with Rec- mediated recombination (KOBAYASHI et al. 1984).
T o inhibit replication, crosses were conducted in dnaBts, Xdv-harboring SMR86 cells (Table 1) with Pam80 phage at 42
”
.
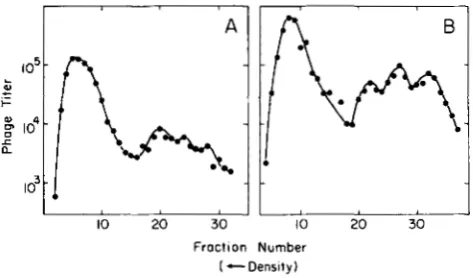
At 4 2”, dnaBts has previously been shown to block X replication fully regardless of whether the X carries a mutant or wild-type allele ofP (STAHL et al. 1972), but the extent of synthesis in Xdv-harboring dnaBts cells was not known. Density transfer experiments (Figure 5 ) indicate that only a partial replication-block was achieved at 42” in the dnaBts cells that carry the Xdv plasmid. Apparently, overproduction of one or both of the X replication proteins partially compensates for the mutant dnaB function at 4 2 ” . Thus, in the experiments reported here, the extent of replication is similar to that shown in Figure 5A.
Genetic selections for heteroduplexes at
P:
The light peak in each gradient from a X by plasmid cross contains X phage whose chromosomes have not spliced-in a plasmid. These include nonrecombinant, parental-type X plus a small fraction (=6%-0.6%) of Rec-mediated patch recombinants (STAHL et al. 1982) (Figures 6-8). T o identify which chain of X DNA carries recombinant information in heteroduplex patches, fractions from the light peaks were submitted to genetic selections for both possible heteroduplex types. Selection I selects for heteroduplex I (het I,r+5
’
1-3’). Selection I1 selects for heteroduplex I1 (het
P
7 - 2 - 5
’
+
11, I-3’). Both selection schemes are described in detail elsewhere (ROSENBERG 1987). The properties of the P gene that allow these selections to work are the following: (1) P is a replication gene. In cells restrictive for a given P allele (e.g., Su- cells and Pam80 phage), replication of the X chromosome is inhibited. (2) P message is transcribed from the X r chain [see FRIEDMAN and GOTTESMAN (1 983) for review]. Thus,
if X is heteroduplex at P , the DNA will replicate only if the P allele permitted by the host is present on the r (transcribed) chain. (3) A host mutation called groP
blocks replication of W + but allows W a m 8 0 to repli- cate (GEORGOPOULOS and HERSKOWITZ 197 1). Thus, either allele can be positively selected.
r L 5 ’
T o select 1-3’, het I, phage are adsorbed to SU-
mutL cells. Only those X with
“+”
information on the r (transcribed) chain can replicate and produce a phage burst. Before bursting, the infected cells are plated on lawns of groP cells. If some Pam80 particles are released from the first burst (of the Su- cell), a plaque is formed on the groP lawn. This procedure is called selection I. Selection I1 is the reciprocal: phage are adsorbed to groP mutL cells; so, only those withP
A
I I I
10 20 30 IO 20 30
Froctlon Number C-Denslty)
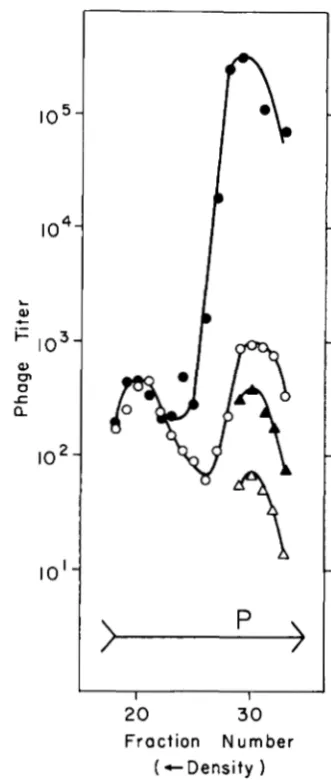
FIGURE 5.-Density transfer experiments to examine extent of replication-block in dnaBts cells carrying hdv. Density labeled X
COSML int29 Pam80 nin5 was passaged through SMR86 cells at 42’ (A) or 39” (B) and centrifuged in cesium formate (see MATERIAIS AND METHODS). The X chromosome is t o o large to allow packaging of any chromosomes that have spliced-in the plasmid. The heaviest peaks in (A) and (B) represent phage with heavy coats and heavy DNA, unadsorbed phage that did not enter the cells. The three lighter peaks represent phage that shed their heavy coats upon entering the cell and emerged with coats made from light isotopes. From left to right they are: heavy/heavy, fully unreplicated “free- loader” phage chromosomes; heavy/light, phage chromosomes that have replicated at least once and have one heavy and one newly- synthesized, light chain; light/light, phage chromosomes that have replicated at least twice and have both chains of light isotopes. Phage were assayed with SMR88 cells. Phage titer is expressed on a log scale.
Pam80 information on the r chain replicate. Infected cells are plated before bursting on lawns of Su- cells.
r P 5
’
This procedure selects heteroduplex I1 (1-3’)+
phage. (“Plaques on selection I and 11” will refer to plaques arising from assay with each selection, respec- tively.) Unadsorbed phage must be removed from either population of (first) infected cells so that any homoduplexes with the genotype permitted by the lawn bacteria do not form plaques. This is accom- plished using genetical and physical methods de- scribed previously (ROSENBERG 1987).
The definition of each plating procedure as a selec- tion is that homoduplex phage of either type and heteroduplex phage of the type not selected do not form plaques when submitted to that selection. Three
X genetic backgrounds were used in crosses here: one has no Chi, another has Chi+D (a Chi situated such that it stimulates recombination at P ) , the third carries a mutant cos (cos2, see Table 2) in the standard posi- tion and bears inverted cos (cosML, Table 2) cloned into the middle of phage X (see MATERIALS AND METH-
ODS for other mutations that all three phage share,
cos Orientation and Patch Chain-Bias 13
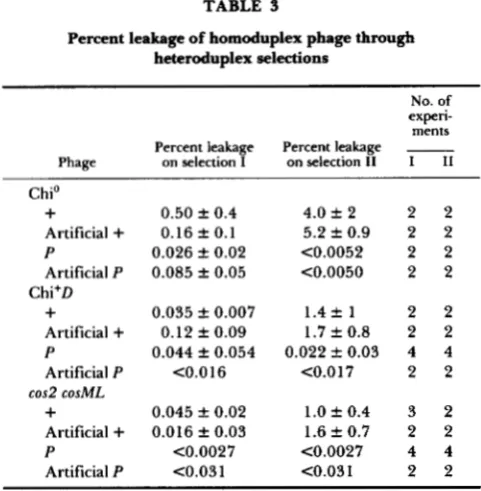
TABLE 3
Percent leakage of homoduplex phage through heteroduplex selections
experi- No. of ments Percent leakage Percent leakage -
Phage on selection I on selection I1 I I1
Chi'
+
0.50 f 0.4 4.0 f 2 2 2Artificial + 0.16 f 0.1 5.2 f 0.9 2 2
P 0.026 k 0.02 C0.0052 2 2
Artificial P 0.085 f 0.05 <0.0050 2 2
+
0.035 f 0.007 1.4 f 1 2 2Artificial + 0.12 f 0.09 1.7 f 0.8 2 2
P 0.044 f 0.054 0.022 f 0.03 4 4
Artificial P C0.016 C0.017 2 2
+
0.045 k 0.02 1.0 f 0.4 3 2Artificial
+
0.016 f 0.03 1.6 f 0.7 2 2P C0.0027 C0.0027 4 4
Artificial P <0.03 1 C0.031 2 2
Chi+D
cos2 cosML
Percentages are relative to the titer of each phage sample assayed with the nonselective host, SMR88. "Chi'" phage contain 6 1 4 5 3 imm21. "Chi+D" phage have b1453 imm2l x+D. 'cos2 COSML" phage bear cos2 cosML b1453 imm21. Artificial
"+"
and -P" phage are homoduplexes made in parallel with the artificial heteroduplex phage used here by chain separation, reannealing and packaging in vitro of X DNAs. These controls show that the chain separation, reannealing and packaging in vitro does not alter plating behavior on selections I and 11. f values are standard deviations for the number of experiments shown, when that number is greater than two. When the number of experiments is two, k values represent measures of variability calculated exactly as standard deviations but with two data.trols. Artificial heteroduplex I and I1 phage of each background were prepared by chain separation and reannealing of X DNAs, followed by packaging in vitro in a mutL packaging system (ROSENBERG 1987). [Mis- match repair in vitro (e.g., LU et al. 1984) could hom- ogenotize artificial heteroduplex phage.] See Table 4 and legend for a biological assay of purity of the separated chains used. The efficiencies of plating of the six artificial heteroduplex phage on selections I
and I1 are given in Table 5. The percentage of het I phage that form plaques inappropriately ("leak") on selection
I1
(II/I leakage, Table 5) will be compared withII/I
titers from actual crosses, below. The as- sumption that the "heteroduplexing process" (chain- separation, reannealing and packaging in vitro) does not affect efficiencies of plating on selections I and I1 is verified in Table 3.Rec-mediated, Chi' patches are heteroduplex with recombinant information biased to the X r chain: X
b1453 imm21 Pam80 Chi' was crossed with Xdv P+ in
SMR86 cells at 42". Figure 6 displays a cesium for- mate equilibrium density gradient of the total cross progeny. The smaller, more dense (heavy) peak rep- resents splices of the Xdv plasmid into the X chromo- some. The larger, less dense (light) peak represents all phage progeny that do not contain a spliced-in
TABLE 4
Purity of separated chains and artificial heteroduplex phage
Titer
per
A genotype DNA packaged packaging
Chi'
Chi+D
r
+
5' 1 = 1 3'P
P 5' I1 = 1 r- 3'
+
r
+
5'1
+
3'r P 5'
1 P 3'
r-
+
5'1 = 1 3'
P
7 - P 5'
I1 = I 3'
+
r+
5'1
+
3'r- P 5'
1 P 3'
cos2 cosML
+
5' 1 = 1 r- 3'P
r- P 5'
I1 = 1 3'
+
r-
+
5'1
+
3'r- P 5'
1 P 3'
1.4 X lo5
2.9 X 10'
C1
220
C1
1.5 X 10'
1.3 X 10'
2.8 X 10'
<10
850
e 1 0
1 . 1 X 103
2.4 X 10'
1.7 X 104
<1
10
<1
130
All of the separated chains and heteroduplex DNA mixtures were treated for reannealing, then precipitated, brought up in TEK buffer for in vitro packaging (ROSENBERC et al. 1985) then in vitro packaged in a single batch of S M R 3 4 extract. Phage were assayed with S M R 8 8 cells. The same amount of DNA was used in each prep, so the titer out of the packaging reaction assays for the presence of packageable, duplex DNA. Note that all r chain preps are not detectably contaminated with their complementary 1 chain and that 1 chain contamination with r is always less than 10%. Since only 1 chains are ever detectably contaminated, any contaminating phage in the heteroduplex phage preps would be homoduplex (of the genotype from which the 1 chain came), not heteroduplex of
the opposite type.
TABLE 5
Efficiencies of plating of artificial heteroduplex phage on heteroduplex selections
Plating Plating
on on
Heteroduplex selection I selection I1
phage (%) (%)
II/I
experiments No. of selection
with I I I
Chi’
+
5‘r-
P
1 = 1 3’ 220 f 4 48 f 6
Chi+D
r
+
5’I = Z 3’ 1 3 0 f 5 0 1 3 + 5
P
r- P 5’
I1 = 1 3’ 0.35 f 0.07 36 f 9
+
cos2 cosML r-
+
5‘1 = 1 3’ 1 0 3 f 4 0 1 6 2 1
P
r- P 5‘
I1
= z
3’ 0.41 f 0.2 31 f 022
10
16
+
The Chi’ X background is X b 1 4 5 3 i m m 2 l . The Chi+D back- ground is X b1453 imm21 x+D. The cos2 cosML background is X cos2
cosML b 1 4 5 3 . Percentages are relative to the titer of each phage assayed on the nonselective indicator, SMR88. Plating conditions for each genotype are described in MATERIALS AND METHODS. Results discussed in text. -C values are standard deviations for the number of experiments shown, when that number is greater than two. When the number of experiments is two, f values represent measures of variability calculated exactly as standard deviations but with t w o data.
Chi’ crosses of X by Xdv (STAHL et uZ. 1982).
Heteroduplex patch structure is deduced from assay of the light peak fractions with selections I and I1 as follows: (1) 0.15% k 0.06% of total peak phage form plaques on selection I (filled triangles, mean of five peak fractions f standard deviation). The percentages of inappropriate plaque formation (leakage) of Pam80 Chi’ phage and P + Chi’ phage on selection I (Table 3) are too small for either total phage (Pum80) or any homoduplex P + patches to account for the titer from assay with selection I. Therefore, I conclude that heteroduplex I phage, patches of
“+”
onto the X rchain, are represented by the titer with selection I. Table 5 shows that Chi’ heteroduplex I phage plate efficiently on selection I. (2) The titer from selection 11 is compared with that from selection I: 18.0% f 1 .O% of the titer from selection I formed plaques on selection I1 (open triangles, mean of five fractions &
standard deviation). Since Chi’ het I X leak on selec- tion I1 at roughly 22% of their efficiency of plating on selection I (II/I leakage, Table 5), the titer from
I O ’
IO‘
I
c 0,
i= 1 0 3
Q,
0
c 0
a
IO
I O ’
P
>
1 I
20 30
F r a c t i o n N u m b e r (“Density
1
FIGURE 6.-Cross of Chi’ X by Xdv. X b1453 imm21 Pam80 was crossed by Xdv and the total progeny centrifuged to equilibrium in cesium formate (see MATERIALS AND METHODS). The heavy peak represents X chromosomes that spliced-in one copy of Xdv. (0) Total phage, assayed on SMR88. (0) P+recombinantsassayed on SMR45. (A) Titers from assay with selection I . (A) Titers from assay with selection 11. Phage titer is expressed on a log scale (see text).
selection I1 from the light peak in Figure 6 is inter- preted as a leakage of the heteroduplex I patches through selection 11. How many heteroduplex I1 ( I -
chain) patches might exist, at most? If as much as half of the titer from selection I1 represents actual het I1 phage (rather than het I “leakers”), and if Chi’ het I1 phage plate with 59% efficiency on selection I1 (data from Table 5), then there could be no more than 16% het I1 ( I chain) patches relative to heteroduplex I ( r
chain) patches. Thus, I conclude that patches are strongly biased to the X r chain. There may be no
Z-
chain patches at all. This result is in agreement with previous results from X by X crosses (ROSENBERG 1987) and generalizes the finding with a X by plasmid cross protocol in which patches are isolated in a different way.cos Orientation and Patch Chain-Bias 15
I
o6
lo5
L t
Q) F
Q)
P,
2
lo4
alo3
h
)
P
%+
I I
30 40
Fraction Number ( * D e n s i t y )
FIGURE 7.-Cross of X Chi+D by Xdv. X 6 1 4 5 3 i m m 2 l P a m 8 0
x+D was crossed by Xdv in SMR86 cells at 42" and the total progeny centrifuged to equilibrium in cesium formate. Symbols are as in Figure 6. Phage titer is expressed on a log stale (see text).
with recombinant information biased to the A r chain: Figure
7
displays a density gradient of progeny of a cross of X b1453 imm2l Pam80 X'D by Xdv P+. That Chi+D acted is evidenced by the fact that both splices (smaller, heavy peak) and total patches (homo- duplex plus heteroduplex, open circles in light peak) are a greater fraction of the total yield (filled circles) than in the Chi' cross (Figure 6). [STAHL et aZ. (1982) showed that Chi stimulates patches and splices equally. Their observation is repeated here.] Total patches (P' homoduplex plus heteroduplex) comprise 6% f 2% of the total phage present in fractions of the light peak (mean of eleven fractions f standard deviation).The structure of heteroduplex patches is deduced as above. (1) The titer from assay with selection I includes 2.6% f 0.4% of the total light peak phage (filled triangles, mean of six fractions f standard deviation). Since this number is too high to be caused by leakage of X Chi+ Pam80 total peak phage (Table 3) or X Chi+P+, homoduplex patch phage (Table 3), I conclude that the titer obtained with selection I rep- resents heteroduplex I, r-chain patches. (2) 12.0% f
1.1 % of the titer of phage plating on selection I formed plaques on selection I1 (open triangles, mean of six fractions f standard deviation). Since Chi+ het
1 phage leak onto selection I1 at roughly 10% of their efficiency of plating on selection I (Table 5), the selection I1 plaque-formers among the light peak frac- tions are interpreted as het I "leakers." How many het I1 (1 chain) patches might there be at most? If as much as half of the titer on selection I1 represents het I1 phage which form plaques at 36% efficiency on selec- tion I1 (Table 5), then no more than 17% heterodu- plex I1 phage could be present, relative to heterodu- plex I phage and there may not be any het 11, Z-chain patches. Thus, I conclude that patches in cis to Chi are strongly r-chain-biased. The r-chained patches (2.6 f 0.4% of total light-peak phage in the Chi+ cross) are stimulated by Chi roughly 1 O-20-fold above the level in the Chi' cross (0.15 f 0.06% of total light- peak phage), illustrating that Chi stimulates formation of r-chained patches as was previously observed (Ro-
SENBERG 1 9 8
7).
r-chain-bias of patches at A
P
is independent of the orientation of A cos: To examine whether chain- bias of Rec-mediated heteroduplex patches is deter- mined by the orientation of cos, heteroduplex chain polarity was assayed among patches onto X with in- verted cos sites. cos control over chain-bias could in- dicate that either the direction of travel of RecBCD enzyme, the orientation of active Chi sites, chain- specific nicking at Chi or Chi-like sequences (or polar- ity of injection or packaging) in some way causes patch chain-bias. The lack of cos control over chain-bias would indicate that none of the above factors do, and thus imply the existence of some other chain-discrim- inating participant in Rec-mediated X recombination.T o address this question, a X with mutant cos at the standard position (cos2, see Table 2) and inverted cos
situated in the middle of X (cosML, see Table 2) and b1453 imm21 Pam80 was crossed with P + Xdv in SMR86 cells at 42". A cesium formate equilibrium density gradient of the total progeny is illustrated in Figure 8. Total P + patches (homoduplex plus hetero- duplex, open circles in the light peak) account for 2% f 1% of the total phage in the light peak (mean of eight fractions f standard deviation).
S.
l o 5
L I O 4
W
.-
+t-
W
0 0
E
l o 3
I O 2
*)
<
)*
I I I
20 30 40
Fraction Number (”Density
1
FIGURE 8.-Cross of X with inverted cos by Xdv. X cos2 cosML b1453 imm2l Pam80 was crossed by Xdv in SMR86 cells at 42‘ and the progeny centrifuged to equilibrium in cesium formate. Symbols are as in Figure 6. (*) mutant cos (cos2). (<) inverted cos (cosML).
Phage titer is expressed on a log scale.
phage “leakers.” I f as much as half of the titer from selection I1 represents actual het 11 (I-chain patch) phage, and if cos2 cosML het I1 phage plate at 31% efficiency on selection I1 (Table 5), then there could be no more than 24% het I1 phage. There may be none at all. Thus, when X cos is inverted, patches remain strongly r-chain-biased.
Could mismatch repair of heteroduplexes in the cross host be responsible for the ?..chain bias of patches in the products of recombination? Here and previously (ROSENBERG 1987), I failed to detect I-
chained, heteroduplex I1 patches. Such results could indicate either that I-chain patches are rare (or non- existent) compared with r-chain patches or that 2-chain patches are selectively removed from products, for example by mismatch repair, either during the cross or during assay of the cross progeny in the heterodu- plex selections. The high efficiencies of plating of heteroduplex I and I1 phage on their respective selec- tions I and I1 (Table 5) indicate that mismatch repair in the mutL plating bacteria does not significantly affect detection of either heteroduplex phage among cross progeny. Could mismatch repair in the cross host obscure the presence of possible I-chain, hetero- duplex I1 patches?
The cross host cells carry the mutL mutation. Nu- merous studies of mismatch repair in which hetero- duplexes of X Pam80 and P+, the markers employed
here, were used indicate that I-3’, het 11, should
not have been selectively repaired in the mutL cross host. The E . coli predominant mismatch repair path- way, the DNA adenine methyl (dam)-directed long patch repair system, does not function in mutL cells [see MESEMN (1987) for a review]. Pam80 is, how- ever, a hotspot (MESELSON 1987; RAPOSA and Fox
1987) for mismatch repair by the yery Short patch (VSP) repair system in which one of the two conjugate heteroduplexes is vigorously corrected whereas the other heteroduplex is unrepaired (LIEB 1985; JONES, WAGNER and RADMAN 1987; MESELSON 1987). How- ever, MESELSON (1987) found that heteroduplex I, the het detected in my experiments, is efficiently repaired and heteroduplex 11, the het that I failed to detect is not VSP repaired. So, VSP repair of Pam80/P+ could not cause r-chain-biased products even if it could occur in mutL cells. Moreover, JONES, WAGNER and RADMAN (1 987) and RAPOSA and Fox (1 987) report that VSP repair at Pam80 is efficiently inhibited by the mutL mutation. [LIEB (1987) finds that mutL di- minishes VSP repair incompletely, but her studies focus on a different locus, the
X
c l gene, and repair efficiencies may be context-dependent as is the case for dam-directed mismatch repair [JONES, WAGNER and RADMAN 19881.) Could other mismatch repair pathways specifically remove het 11’s in mutL cells? RAPOSA and FOX (1987) quantitated the extent to which both conjugate heteroduplexes of Pam80 and P+ are repaired in mutL cells. They report that 83% of all heteroduplexes I and I1 are not repaired in mutL cells. Of the remaining 17% of either heteroduplex, repair is neither chain-directed, nor allele-directed.“+”
and “P” homoduplexes are formed roughly equally. (Also, much of this residual “repair” involves loss of all the markers on one chain and so may actually reflect asymmetric replication or “chain loss”.) Even if17% of het Is were not also repaired in the cross host cells, the 17% of mismatch repaired het 11s alone is not sufficient to generate the extent of r-chain-bias detected previously (at least 90%, ROSENBERG 198’7) or here.
In contrast with the above considerations on (the inefficiency of ) mismatch repair of Pam80/P+ heter- oduplexes in mutL cells, the following proposal [sug- gested by G . R. SMITH (Seattle)] will be considered as an alternative interpretation of the results here and in ROSENBERC (1987). The tenets of this alternative proposal follow. (1) Rec-mediated recombination pro- ceeds via a nick-at-Chi initiation mechanism. (2) As predicted by the simple nick-at-Chi initiation model, I-chain patches are the predominant class of patch
r L 5
’
cos Orientation and Patch Chain-Bias 17
recombinants (when cos is wild-type and nicking would occur on the
1
chain). But l-chain patches are selec- tively removed (here and in ROSENBERG 1987) due to mismatch repair in the cross host cell. (3) The cross host cells used here and in ROSENBERG (1987) do in fact repair Pam80/P+ het IIs, in contrast with themutL cells studied by others (work cited above) and the plating bacteria used here (Table 5, and previously ROSENBERG 1987), due to “special physiological con- ditions” which occur in these mutL cross hosts. (4) The r-chain patches detected here and previously are pro- posed to constitute a minority of patches which is more apparent than the (silent majority o f ) 1-chain patches because het I, r-chain patches are postulated to be refractory to the proposed repair.
A prediction of this proposal is addressed by the data in Figures 6-8. If recombination proceeds via the simple nick-at-Chi initiation model (Figure l), (invisible) 1-chain patches are predicted to predomi- nate only when cos is wild-type and the X 1 chain would be the nicked, invasive chain; only then should r-chain patches be a “minority.” When cos is inverted, inverted Chi-like sequences would be nicked on the X r chain (TAYLOR et al. 1985; CHENG and SMITH 1987) and invasion therefrom should produce a visible majority of unrepairable, r-chain patches. The relative “minor-
ity” or “majority” status of r-chain, het I patches when
cos is wild-type or inverted (filled triangles, Figures 6 and 7 us. Figure 8) can be assessed by comparison with the level of splices (filled and open circles, heavy peaks) for each cross. STAHL et al. (1982) demon- strated that the patch to splice ratio is constant whether Chi is present or absent and whether cos is wild-type or inverted. In Figures 6-8, comparison of the splice- and r-chain patch-peaks reveals a rough equality between the two classes of recombinants in each cross progeny. r-chain patches do not become more numerous relative to splices when cos is inverted, contrary to the prediction made by the alternative proposal. (Also note that r-chain patches, which occur at rough equality with splices, cannot be considered a “minority” product of recombination.) Therefore the alternative proposal (above) cannot explain the ab- sence of detectable 1-chained patches. Taken together with the data on (ineffkiency o f ) mismatch repair of Pam80/P+ hets in mutL cells, I conclude that 1-chain patches are not detected because they are produced rarely or not at all by Rec-mediated recombination at the X P locus.
DISCUSSION
In Rec-mediated crosses of X with Xdv in which the P locus is examined: (1) patches are strongly r-chain- biased, (2) Chi-stimulated patches onto the Chi-con- taining molecule (in cis to Chi) are strongly r-chain- biased, (3) chain-bias of patches is not imposed on recombination by the orientation of X cos. The first
two results extend and generalize previous findings from X by X crosses: (1) Rec-mediated patches from Chi’ crosses are biased to the X r chain, (2) Chi- stimulated patches onto the Chi’ molecule (patches in trans to Chi) are r-chain-biased (ROSENBERC 1987). Results (1) and (2) ruled out the simple version of the nick-at-Chi initiation model in Figure 1. The fre- quency of r-chained patches at P was stimulated roughly 10-20-fold by Chi (here) and 4-fold by Chi in X by X crosses (ROSENBERC 1987), indicating that heteroduplex patches at P experience Chi-stimulation just as do splices and patches not examined for het- eroduplex chain-polarity (STAHL et al. 1982; STAHL, LIEB and STAHL 1984) [see SMITH and STAHL (1 985) and STAHL (1986) for reviews].
Result (3) is important in two contexts: the result constrains some previously elaborated models and indicates the existence of a new element in recombi- nation. Models disproved will be addressed first:
Nick-at-Chi initiation models: In nick-at-Chi initi- ation models, a chain-specific nick similar to the one observed in vitro (PONTICELLI et al. 1985; TAYLOR et al. 1985) provides a single-chain end that invades a homolog, initiating recombination. An economical version of this model predicted patches biased to the same chain on which the nick occurred [see Figure 1 from SMITH et al. (1 984); prediction from ROSENBERG
(1987)l. For frontward Chi’s and Chi-like sequences in X with wild-type cos, nicking would be on the X 1-
chain; the model predicted 1 chain patches; and the model was disproved by the demonstration of r-chain- biased patches at the X P locus [ROSENBERG (1987) and here]. However, one could imagine more compli- cated nick-at-Chi initiation models. A nicked chain might invade a homolog as before, but the following steps in recombination might result in patches biased to the chain opposite that nicked. So, a nick on the 1
chain could, by such models, yield r-chain-biased
patches at P [for example, see Figure 2 from ROSEN-
BERG (1987)l. Since chain-specificity of nicking (at
properly oriented Chi’s and Chi-like sequences) is dictated by the direction of RecBCD travel, such models predict that if cos is inverted, patch chain-bias at any locus should switch because (1) inversion of cos in X causes the direction of travel of RecBCD through X to reverse (KOBAYASHI, STAHL and STAHL 1984;
YAGIL and SHTROMAS 1985; STAHL et al. 1986); (2) when RecBCD travels in the reverse direction, only inverted Chi-like sequences would be nicked, and that nick would occur on the X r chain (TAYLOR et al. 1985; CHENG and SMITH 1987); and (3) therefore, such models (for example, Figure 2) predict that in- version of cos should cause l-chain-biased patches at X P (for example, see Figure 3). This class of models is disproved by the finding that patches at X P remain r- chain-biased when cos is inverted.
18 S. M. Rosenberg
ation model with extra assumptions such that r-chain- bias would still be expected when RecBCD travel direction is reversed? A useful way to restate that question is: is there any rule by which the intermedi- ates (C) in Figures 2 and 3 could both be resolved to yield r-chained patches? There is: stipulate a chain- directed HOLLIDAY junction resolvase that follows the rule: when double HOLLIDAY junctions are encoun- tered one of the two must be resolved by cutting r chains ("vertical" resolution as is shown in Figure 2C, "horizontal" resolution contrary to that shown in Fig- ure 3C). The other junction of each pair would be cut in either sense yielding patches (if cut in the same sense as the first) or splices (if cut in the opposite sense as the first). r-chain patches would result regardless of RecBCD travel direction. T h e extra burden assumed by this model is the demand that resolvase know the
1 from r chain and know it without reference to cos,
direction of RecBCD travel or active Chi's or Chi-like sequences. Thus, this augmentation of the compli- cated nick-at-Chi initiation models in Figures 2 and 3 predicts a sequence on X that directs resolvase. This here-to-for undiscovered site would be important for Rec-mediated recombination, and will not be cos or Chi.
Conversely, such a complicated model will have lost its attractiveness for some. Since more economical nick-at-Chi initiation models (Figures 1-3) have been disproved the question of the relevance of in vitro nicking to in vivo recombination seems pertinent. Thus far, chain-specific nicking at Chi in vivo has not been found. Rec-mediated recombination has not been observed in vitro (under any conditions, includ- ing those in which nicking at Chi is observed). Thus, a conservative conclusion to be drawn from in vitro nicking seems warranted. T h e conservative conclu- sion is that Chi and RecBCD directly interact [as was implied by earlier genetic experiments (SCHULTZ,
TAYLOR and SMITH 1983; CHAUDHURY and SMITH
1984)l. [Elsewhere, we argue that all of the activities of RecBCD enzyme in vitro are antirecombinogenic activities of the enzyme, not partial reactions of Rec- mediated recombination (THALER et al. 1988).]
Viable models: what element causes patch chain- bias? T h e existence of a previously unknown element in Rec-mediated recombination is implied by the find- ing that r-chain-bias of patches at P is independent of the only participants in Rec known to have a polar relationship with the X chromosome: cos orientation, orientation and chain-specificity of nicking of Chi's and Chi-like sequences and the direction of travel of RecBCD. Some chain-discriminating process other than those three must interact with recombination causing chain-biased products. For the sake of econ- omy, I propose two classes of models that use already- known, chain-directed features of DNA metabolism, replication or transcription [ROSENBERG (1987) and
"
o r i P
1
-7
-
%
FIGURE 9.-Abbreviated X map. Straight lines with arrowheads represent the directions of replication. Wavy lines with arrowheads indicate the directions of transcription. ori is the X replication origin, PR and PL represent, respectively, the X major rightward and leftward promoters. [See FRIEDMAN and GOTTESMAN (1983) and
FURTH and WICKNER (1 983) for reviews.]
R E P L l C A T l O N
nri D
T R A N S C R I P T I O N
+
/ uri 3' I P
5'
3'
5' r
+
RNase H -DNase H"+I N I T I A T I O N
"give- or "take-
I I
r
/
Y f
-
r
I
""_
+
I r
r
+
I
FIGURE 10.-Replication and transcription models for r-chain- biased patches at X P. Straight lines represent DNA; wavy lines represent RNA; dashed lines represent newly synthesized DNA. Where "3" or "5" d o not label a DNA end, the polarity of that end is not implied. Thus a particular chain-polarity for the invading chain in "give" initiation is not implied in this model. Models described in text.
further elaborated herein]. Both models suppose that initiations, which are chain-directed by replication or transcription, are not rate-limiting and that RecBCD- Chi interaction promotes resolution of a (HOLLIDAY- type) intermediate.