1
Channels control flow in and out of cells
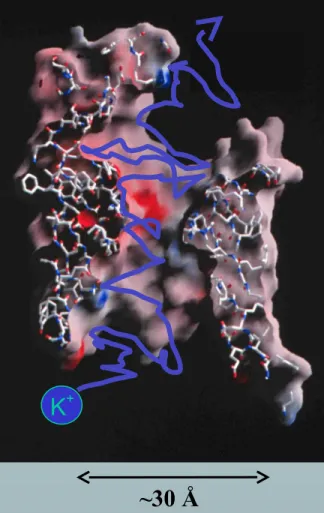
~5 µm
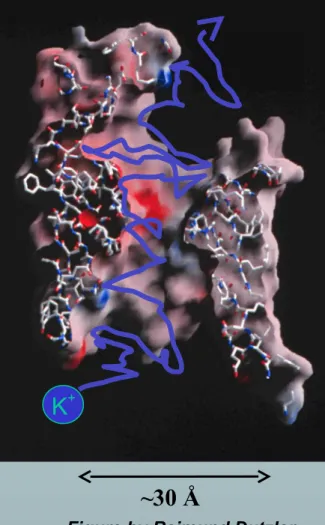
2 Figure of ompF porin by Raimund Dutzler
Ion Channels
are the
Valves* of Biological Function
Main Controllers of Cellular Function
*Pun:
Valve
=
Vacuum
Tube
≈
PNP
Transistor
≈
FET
Transistor
BritSpeak
~30 Å
Flow time scale is 0.1 msec to min
K+
Chemical Bonds are lines Surface is Electrical Potential
Red is negative (acid)
3
Natural nano-valves* for atomic control of biological function
Ion channels
control all electrical activity in cells produce signals of the nervous system coordinate contraction in skeletal musclecoordinate contraction in the heart, allowing the heart to function as a pump
control secretion and absorption
are proteins whose genes (blueprints) can
be manipulated by molecular genetics have structures shown by x-ray
crystallography in favorable cases are involved in thousands of diseases and
many drugs act on channels studied by thousands of scientists
with >20,000 AxoPatch or other amplifiers
*nearly pico-valves: diameter is 400900 pico-meters
Ion Channels are Biological Devices
~30 Å
K+
Conflict of Interest
4
Natural nano-valves* for atomic control of macroscopic flow
Ion channels
have SelectivityCalcium channel selects Ca++ over Na+ by ~104
Ion channels
respond to very small stimuli: chemicals, voltage, pH, and mechanical force.Control macroscopic flow with Atomic structures.
Gate/Switch from conducting to nonconducting state
Ion channel
proteins allow Atomic Scale Mutations to control electrical charge and biological functionIon channels
are biologicalDevices
that self-assemble into perfectly reproducible arrays
Ion channels
are Templates for design of bio-devices and biosensorsIon channels
have VERY Large Charge Densities critical to I-V characteristics and selectivity (and gating?)*nearly pico-valves: diameter is 400900 pico-meters
Ion Channels are Engineering Devices
~30 Å
K+
5
Ion Channels are Physical Devices
Channels control flow of Charged Spheres
Channels have Simple Invariant Structure
on the biological time scale
Why can’t we predict the movement of
Charged Spheres through a Hole?
Physics of Ionic Movement under biological conditions is
MUCH SIMPLER
than the physics of Fluids, Plasmas, or Semiconductors,
6
Mathematics
describes only a tiny part of life,
But
Mathematics* Creates
our
Standard of Living
*e.g.,
electricity, computers, fluid dynamics, optics, structural mechanics, ….
u
7
How can we use mathematics to describe
biological systems?
I believe
Some biology can be described by
Physics and Mathematics ‘as usual’
Guess, Solve, Check
But you have to know which biology!
u
8
Channels are only Holes
Why can’t we have a fully successful theory?
Must know physical basis to make a good theory
Where do we start?
9
Proteins Bristle with Charge
Cohn (1920’s) & Edsall (1940’s)
We start with
Electrostatics
because of biology
We will soon add
10
We will soon add
Finite Size Effects
Chemically Specific Properties
of ions
come from their
Diameter and Charge
not vaguely defined hydration shells or ‘chemical’
bonds
11
Active Sites
of Proteins are
Very
Charged
e.g. 7 charges ~
20 M net charge
Selectivity Filters and Gates of Ion Channels
are
Active Sites
1 nM
=
10
Å
= 1.2×10
22
cm
-3
-+ -+ -+
+
+
-
-Ions are
Crowded
12
We start with Langevin equations of
charged particles
Simplest stochastic trajectories
are
Brownian Motion of Charged Particles
Einstein, Smoluchowski, and Langevin ignored charge
and therefore
do not describe Brownian motion of ions in solutions
Once we learn to count Trajectories of Brownian Motion of Charge,
we can count trajectories of Molecular Dynamics
Opportunity
and Need
We use
Theory of Stochastic Processes
to go
from Trajectories to Probabilities
13
James Clerk Maxwell
“I only count molecules ….,
avoiding all personal enquiries
which would only get me into trouble.”
Slightly rephrased from Royal Society of London, 1879, Archives no. 188
14 Schuss, Nadler, Singer, Eisenberg
Langevin
Equations
Electric Force
from
all charges
including
Permanent charge of
Protein
,
Dielectric Boundary charges,
Boundary condition charge
;
k
2
p
k
x q
p
p
p
k
k
k
f
kT
m
m
x
x
w
Positive
cat
ion
,
e.g.,
p
=
Na
+
;
Newton's Law
Friction & Noise
2
k
n
k
x q
n
n
n
k
k
k
f
kT
m
m
x
x
w
15
01
p k p nprotein j j
j j
f
eP
e
t
e
t
x
= div
x grad
x
x
x x
x x
Trajectories of
Negative Ions
Trajectories of Positive
Ions
Electric Force
from all charges including
P
ermanent charge ofP
rotein, Dielectric Boundary charges,Boundary condition charge
Electric Force
in each Trajectory
from Poisson Equation
Schuss, Nadler, Singer, Eisenberg
16 Schuss, Nadler, Singer, Eisenberg
0
0
p
p
p n
k
k
xs
k
i i
i
i
e
e
f
f
q
z
x
div
E
x
x
Electric Force from
AVERAGED Poisson Equation
Excess
‘Chemical
’
Force
Implicit Solvent
‘Primitive’ Model
or
Primitive Solvent Model
Electric Force
from all charges including
P
ermanent charge ofP
rotein, Dielectric Boundary charges,Boundary condition charge
17
Conditional PNP
Schuss, Nadler, Singer, Eisenberg
0
|
|
|
y
y
e
y
y x
y
y x
y x
P
Electric Force depends on Conditional Density of Charge
Nernst-Planck gives
UN
conditional Density of Charge
1
|
0
y
x e
y
y x
y x
m
x
Other Forces
Mass
Friction
Closures or Approximations
Needed
18
Finite OPEN System
Fokker Planck Equation
Boltzmann Distribution
Trajectories
Configurations
Nonequilibrium
Equilibrium
Thermodynamics
Thermodynamics
Device Equation
lim ,
N V
Statistical Mechanics
Theory of Stochastic Processes
19
Poisson’s Equation
Drift-diffusion & Continuity Equation
0;
J x
i
D
i
i
1
i
kT
J x
x
x
x
0
i i
i
e
e
z
x
x
x
x
Chemical Potential
ln
i
* ex
ii
z e
ikT
x
x
x
x
Chemical Correlations
Poisson-Nernst-Planck
(PNP)
Channel
P
rotein Positive Permanent Charge is Basic20
Semiconductor Equations:
One Dimensional PNP
i
i
i
i
d
J
D x A x
x
dx
Poisson’s Equation
Drift-diffusion & Continuity Equation
0
ii i
d
d
x A x
e
x
e
z
x
A x dx
dx
0
idJ
dx
Chemical Potential
ex
*
x
x
x
ln
ix
i
z e
ikT
i
Special Chemistry
Chemical Correlations ‘by hand’: finite size
Channel
P
rotein Positive Permanent Charge is Basic21
Poisson-Nernst-Planck
(PNP)
Counting at low resolution gives
‘Semiconductor Equations’
Gouy-Chapman, (nonlinear) Poisson-Boltzmann,
Debye-Hückel,
are siblings with similar resolution
but at equilibrium, without current or flux of any species
Devices do not exist at equilibrium
22
Compare with Biological Function!
How do we check the theory?
Our task is to
Discover & Understand, Control & Improve
Biological Function
Inverse Problem
Heinz Engl (Vienna, Linz), Martin Burger (Muenster, Linz)
Must use realistic concentrations
because they are the
Power Supply
23
Selectivity
Differs
in Different Types of
Channels
24
Selectivity
depends on Correlations
I put in
Correlations ‘by hand’
as do the chemists
because
I do not know how to do better!
25
Chemically Specific Properties
of ions come from
Correlations produced by
Diameter & Charge
(much) more than anything else.
Established by Regensburg School, Josef Barthel leader,
Learned from Doug Henderson, J.-P. Hansen, Stuart Rice, among others…Thanks!
Physical Models are Enough
Vaguely defined hydration shells
are
26
PNP/DFT
Selectivity
filter
DDDD
4
−
charges
RyR
RyR
Channel
Channel
PNP/DFT
Monte Carlo
PNP/DFT
Monte Carlo
Selectivity
filter
Various
many - charges
Selectivity
filter
DEKA
2
−, 1+ charge
Selectivity
filter
EEEE
4
−
charges
Synthetic
Synthetic
Ca Channel
Ca Channel
Sodium
Sodium
Channel
Channel
Calcium
Calcium
Channel
Channel
Selectivity of Different Channel Types Studied in Many Solutions
K channel Model of Benoit Roux is very Similar
Quantum Water/K
+Model of Susan Rempe is very Similar,
but
Neither K model has yet been computed in a range of solutions
27
Understand Selectivity
well enough to
Make a Calcium Channel
using techniques of
molecular genetics,
site-directed Mutagenesis
Goal:
28 30 60 -30 30 60 0
pA
mV
LECE (-7e)
LECE-MTSES- (-8e)
LECE-GLUT- (-8e)
E
CaE
ClWT (-1e)
Calcium selective
Experiment
Two Synthetic Calcium Channels
Designed by Theory
As charge density increases, channel becomes calcium selective
Erev ECa
Unselective Wild Type
built by Henk Miedema, Wim Meijberg of BioMade Corp.,Groningen, Netherlands Miedema et al, Biophys J 87: 3137–3147 (2004)
MUTANT ─ Compound
29 -6 -4 -2 0 2 4 6
-100 -50 0 50 100
5 nm diameter
1.0 mM
KCl 10 mM KCl
-2 0 2 4 6
-150 -100 -50 0 50 100 150
pA
mV
-3 -1 1 3 5-100 -50 0 50 100
pA
mV
K+ selectivity Ca2+ selectivity 10 mM CaCl2 0.1 mM CaCl2 1.0 mM KCl 10 mM CaC2 10 mM KCl 0.1 mM CaCl2pA
mV
Ca2+ selectivityVr ~ 10 mV
Double Gradients
Zuzanna Siwy & Yan He, UC Irvine
Polyethylene terephthalate
200 nm
30
Understand Selectivity
Goal:
For L-type Ca channel
Selectivity = Binding Curves
31
Binding Curve
32
O
½Selectivity
Filter
Selectivity Filter
Crowded with Charge
Wolfgang Nonner
+
33 Dielectric Protein
Dielectric Protein
6 Å
μ
mobile ions
=
μ
mobile ions
Ion ‘Binding’ in Crowded Channel
Classical Donnan Equilibrium of Ion Exchanger
Side chains move within channel to their equilibrium position of minimal free energy. We compute the Tertiary Structure as the structure of minimal free energy.
Boda, Nonner, Valisko, Henderson, Eisenberg & Gillespie
34
We* use
Monte Carlo Simulations
over a wide range of concentrations
based on
Amino Acid Sequence
of the
Selectivity Filter
Three Dimensional Structures not known,
so we use Sieving Data and Side Chain Size
and compute
Tertiary Structure as an Output
35
Ion Diameters
Pauling Diameters
Ca
++1.98 Å
Na
+2.00 Å
K
+2.66 Å
‘
Side Chain’ Diameter
Lysine K
3.00 Å
D or E
2.80 Å
Channel Diameter 6 Å
Parameters are Fixed in all calculations in all solutions for all mutants
Boda, Nonner, Valisko, Henderson, Eisenberg & Gillespie
‘Side Chains’
are Spheres
Free
to move inside channelSnap Shots of Contents
Crowded Ions
6Å
36 10
‘Titration Curve’
Classical Definition of Selectivity
Boda, Nonner, Valisko, Henderson, Eisenberg & Gillespie
Calcium
Channel
6Å
3 electron charges
High Occupancy
Sodium
Channel
6Å
1 electron charge
Low Occupancy
Biology
Na
+2 Å
Ca
++1.98 Å
We believe curves like these must be reproduced by a useful theory
Mutation O cc up an cy
Low
Occupancy
High
Occupancy
37
Charge Selectivity
Na
+vs
Ca
++Depends on Dielectric
Dielectric Boundary Force = DBF
Dielectric is a Charge Amplifier
Na
+
2.0 Å
Small Size
andLarge
Dielectric
Boundary
Force
DEKA
Na Channel,
6 ÅZero
Dielectric
Boundary Force
Large
Dielectric
Boundary Force
2 solvent protein ion solvent protein DBF = f Q
Boda et al
protein solvent
protein solvent
Diameter
/
Å 5 25 15S
el
ec
ti
vi
t
y
# N a +
/
# C a + +1.98
ÅCa
++ε
solvent10
p
solvent6 8 10
38
We turn from Charge Selectivity
to
Size Selectivity
Na
+
vs
K
+
in
the DEKA Na Channel
39 Selectivity Filter Selectivity Filter Selectivity Filter Selectivity Filter Selectivity Filter Selectivity Filter Selectivity Filter Selectivity Filter
log C/Cref
Binding Sites*
*Binding Sites are
outputs of our model,
not
structural inputs
Boda, et al
Ion Diameter
Ca++ 1.98 Å
Na+ 2.00 Å
K+ 2.66 Å
‘Side Chain’ Diameter
3.00 Å
2.80 Å
Na Channel DEKA6Å
1 2
O
+ 4
NH
Lys or K
D or E
[NaCl] = [KCl] = 50 mM
BLACK
=
Depletion=0
Na vs K Size Selectivity is in Depletion Zone
Size
Selectivity
Size
40
Size Selectivity is in the Depletion Zone
K
+Depletion Zone
Na
+Boda, et al
0 K+
in Depletion Zone
[NaCl] = 50 mM [KCl] = 50 mM
Selectivity Filter Channel Protein
Na
+vs.
K
+Occupancy
of the DEKA Na Channel, 6 Å
41
Size Selectivity
Depends on Channel Size,
not
Protein
Dielectric Coefficient
Selectivity
for small ion
Na
+
2.00 Å
K
+
2.66 Å
in DEKA 6Å
Na Channel
K+ Na+
Boda, et al
Small Channel Diameter Large
in Å
42
Size Selectivity does not depend on protein dielectric
Occupancy Depends on Protein Dielectric,
Protein
Dielectric Amplifies Electrostatics
80
solvent
= Dielectric Coefficient =
of Protein
protein
protein
O
cc
up
an
cy
Boda, et al
DEKA Na Channel, 6 Å
Occupancy
Na
+2.0 Å
K
+
43
Binding Sites* are
OUTPUTS
of our Calculations
*Selectivity is in the Depletion Zone,
NOT IN THE BINDING SITE
of the DEKA Na Channel
Our model has no preformed
structural binding sites
but
44
Repulsion
Binding Sites
Electrostatic Attraction
Steric Competition for Space
Location and Strength of Binding Sites
Depend on Ionic Concentration and
Ryanodine Receptor RyR
Model has 5 charged amino acids
found by mutation experiments to
determine selectivity and permeation
The model reproduces >100 data
curves of RyR with 8 adjustable
parameters (geometry and K
+) and
only 1 for non-K
+ions. Parameters
determined by fits to tiny data set.
Once found, parameters were
never changed.
Model predicted all AMFE data of
RyR with no adjustable parameters
Model predicts effect of drastic
mutation (removal of fixed charge)
Fixed charge 13M
0
Water (pure) is 55 M
RyR
Receptor
Gillespie, Meissner, Le Xu, et al,
not
Bob Eisenberg
More than 100 combinations of solutions
&
mutants
7 mutants with significant effects fit successfully
Model reproduces >120 data curves of RyR with
8 adjustable parameters (geometry, K
+) and
1 adjustable parameter, for each other ion type.
Parameters determined by fits to tiny data set.
47
Nonner, Gillespie, Eisenberg
DFT/PNP
vs
Monte Carlo Simulations
Concentration Profiles
48
Divalents
KCl
CaCl
2CsCl
CaCl
2NaCl
CaCl
2KCl
MgCl
2Misfit
49
LiCl
D. Gillespie et al., J. Phys. Chem. 109, 15598 (2005).
Misfit in unequal concentration
50
KCl
Theory fits Mutation with Zero Charge
No parameters adjusted
Gillespie et al
J Phys Chem 109 15598 (2005)
Protein charge density
wild type*
13
M
Water is 55
M*some wild type curves not shown, ‘off the graph’
0
M
in
D4899
Theory Fits Mutant
in K
The model predicted an AMFE for Na
+/Cs
+mixtures
before it had been measured
Gillespie, Meissner, Le Xu, et al
Bulk Solution
Channel
62 measurements Thanks to Le Xu!
The model predicted that AMFE disappears
Prediction made
without any adjustable
parameters.
The Na
+/Cs
+mole
fraction experiment is
repeated with varying
amounts of KCl and
LiCl present in addition
to the NaCl and CsCl.
The model predicted
that the AMFE
disappears when other
cations are present.
This was later
confirmed by
experiment.
Mixtures of THREE Ions
The model reproduced the competition of cations for the pore
without any adjustable parameters.
Li
+&
K
+ &Cs
+Li
+ &Na
+ &Cs
+55
Traditional Biochemistry
(more or less)
Ignores the Electric Field
56
But
Rate Constants depend steeply
on
Electrical Properties
and
Concentration*
because of shielding, a fundamental property of matter,
independent of model,
in my opinion.
57
Electrostatic Contribution
to ‘Dissociation Constant’ is large and is an
Important Determinant of
Biological Properties
Change of
Dissociation ‘Constant’
with concentration is large and is an
58
What does the protein do?
Selectivity arises from
Electrostatics and Crowding of Charge
Certain
MEASURES
of structure are
Powerful
DETERMINANTS
of Function
e.g., Volume, Dielectric Coefficient, etc
.
Precise Arrangement of Atoms is not involved
in the model,
to first order
.
59
Ionic Selectivity in Protein Channels
Crowded Charge Mechanism
4 Negative Charges
of glutamates of protein
DEMAND
4 Positive Charges
nearby
either 4 Na
+
or 2 Ca
++
60
Ionic Selectivity in Protein Channels
Crowded Charge Mechanism
2 Ca
++
are
LESS CROWDED
than 4 Na
+
Ca
++
SHIELDS BETTER
than Na
+
, so
Protein Prefers Calcium
61
2 Ca
++
are
LESS CROWDED
than 4 Na
+
62
Protein maintains
Mechanical Forces
*
Volume of Pore
Dielectric Coefficient/Boundary
Permanent Charge
Precise Arrangement of Atoms is not involved
in the model,
to first order
.
but
Particular properties
(‘measures’)
of the protein are crucial!
What does the protein do?
Nonner and Eisenberg
63
Other
Properties of Ion Channels
are likely to involve
more subtle physics
including
orbital delocalization
and
chemical binding
Selectivity apparently does not!
64
Remember
We can build them
(reasonably well)
65 30 60 -30 30 60 0
pA
mV
LECE (-7e)
LECE-MTSES- (-8e)
LECE-GLUT- (-8e)
E
CaE
ClWT (-1e)
Calcium selective
Experiment
Two Synthetic Calcium Channels
Designed by Theory
As charge density increases, channel becomes calcium selective
Erev ECa
Unselective Wild Type
built by Henk Miedema, Wim Meijberg of BioMade Corp.,Groningen, Netherlands Miedema et al, Biophys J 87: 3137–3147 (2004)
66
5 nm diameter
1.0 mM
KCl 10 mM KCl
-2 0 2 4 6
-150 -100 -50 0 50 100 150
pA
mV
-3 -1 1 3 5-100 -50 0 50 100
pA
mV
K+ selectivity Ca2+ selectivity 10 mM CaCl2 0.1 mM CaCl2 1.0 mM KCl 10 mM CaC2 10 mM KCl 0.1 mM CaCl2 -6 -4 -2 0 2 4 6-100 -50 0 50 100
pA
mV
Ca2+
selectivity
Vr ~ 10 mV
Double Gradients
Zuzanna Siwy & Yan He, UC Irvine
Polyethylene terephthalate
200 nm
67
Selectivity can be understood
by
Reduced Models
K
channels
Benoit Roux
Susan Rempe
Na
&
Ca
channels
Nonner,
et al,
RyR
channels
Gillespie
&
Meissner
70
Binding Curve
71
We* use
Monte Carlo Simulations
over a wide range of concentrations
based on
Amino Acid Sequence
of the
Selectivity Filter
Three Dimensional Structures not known,
so we use Sieving Data and Side Chain Size
and compute
Tertiary Structure as an Output
72
What about
Traditional View of Selectivity
and
Binding Sites
?
73
Electrodiffusion of
charged, hard spheres
Correlations put in ‘by Hand’
using best available theory of liquids
74
Binding Sites Come From
Forces
in this model.
Forces
in the model are
Electrostatic Attraction
Steric Competition for Space
75
Ryanodine Receptor
one type of Calcium Channel
work of Gillespie, Meissner, Le Xu,
et al
RyR
Receptor
Gillespie, Meissner, Le Xu, et al, studied
More than 100 combinations of solutions
&
mutants
7 mutants with significant effects fit successfully
Model reproduces >100 data curves of RyR with
8 adjustable parameters (geometry, K
+) and
1 adjustable parameter, for each other ion type.
Parameters determined by fits to tiny data set.
77
Theory fits Mutation with Zero Charge
No parameters adjusted
Gillespie et al
J Phys Chem 109 15598 (2005)
Protein charge density
wild type*
13
M
Water is 55
M*some wild type curves not shown, ‘off the graph’
0
M
in
D4899
Theory Fits Mutant
in K
Theory Fits Mutant
The model predicted an AMFE for Na
+/Cs
+mixtures
before it had been measured
Gillespie, Meissner, Le Xu, et al
80
We* use
Monte Carlo Simulations
over a wide range of concentrations
based on
Amino Acid Sequence
of the
Selectivity Filter
Three Dimensional Structures not known,
so we use Sieving Data and Side Chain Size
and compute
Tertiary Structure as an Output
81
We turn from Charge Selectivity
to
Size Selectivity
Na
+
vs
K
+
in
the DEKA Na Channel
82
What about
Traditional View of Selectivity
and
Binding Sites
?
83
Binding Sites Come From
Forces
in this model.
Forces
in the model are
Electrostatic Attraction
Steric Competition for Space
84
Binding Sites* are
OUTPUTS
of our Calculations
Location and Strength of Binding Sites Depend
on Ionic Concentration and Temperature (etc)
*Selectivity is in the Depletion Zone,
NOT IN THE BINDING SITE
of the DEKA Na Channel
Our model has no preformed
structural binding sites
but
Selectivity is very specific
85
Ionic Selectivity in Protein Channels
Crowded Charge Mechanism
Simplest Version: MSAHow does
86
General Biological Theme
(‘adaptation’)
Selectivity Arises in a Crowded Space
Biological case:
0.1 M NaCl and 1 M CaCl2 in the baths