Helgol~nder Meeresunters. 33, 443--451 (1980)
R e d u c U v e m i c r o b i a l d i s s o l u t i o n of m a n g a n e s e n o d u l e s
as a p o s s i b l e h a z a r d of d e e p - s e a m i n i n g
C. Schiitt & J. C. G. Ottow
Institut fiir B o d e n k u n d e und Standortslehre, Universit~t, Hohenheim;
D-7000 Stuttgart-Hohenheim-70, Federal Republic of Germany
ABSTRACT: The microbial lysis of deep-sea nodules as a possible result of large-scale, deep-sea mining is considered. It is assumed that the Mn (IV} and Fe (III) compounds of the manganese nodules are reduced by the numerous aerobic bacteria at the sediment/water interface as soon as the adjacent nodule area is buried by sedimentation of the disturbed deposits and the organic-rich debris from the blooming surface plankton. Intensive mineralization processes in the resettled sediments cause oxygen depletion. Subsequently, the aerobic (and anaerobic) microorganisms will switch to Mn (IV) and Fe (III) oxides as alternative electron acceptors in order to continue their energy-conserving (ATP synthesis) reactions (anaerobic respiration). The higher the amount of decomposable organic matter, the more intensive are these processes. Consequently, buried manganese nodules may be dissolved, thereby liberating mobile Mn (II), He (II) and several trace elements (Ni, Cu, Co and others). This possible hazard and its ecological consequences should be evaluated carefully before deep-sea mining is started on a large scale.
I N T R O D U C T I O N
A p p r o x i m a t e l y 100 y e a r s ago, t h e first m a n g a n e s e n o d u l e s w e r e d i s c o v e r e d b y t h e British r e s e a r c h v e s s e l " C h a l l e n g e r " on t h e b o t t o n of t h e A t l a n t i c O c e a n at d e p t h s b e t w e e n 4000 a n d 5000 m. H o w e v e r , not u n t i l t h e m i d - s i x t i e s w a s t h e e c o n o m i c i m p o r t a n c e of t h e s e - i n fact p o l y m e t a l l i c - c o n c r e t i o n s r e c o g n i z e d (Mero, 1962). B e c a u s e of t h e r e l a t i v l y h i g h c o n t e n t of n i c k e l , copper, cobalt a n d m a n g a n e s e (Table 1), t h e s e m i n e r a l a c c r e t i o n s m a y b e c o m e i n c r e a s i n g l y i m p o r t a n t , s i n c e t h e t e r r e s t r i a l sources of t h e s e e s s e n t i a l e l e m e n t s m a y r u n short w i t h i n a v e r y l i m i t e d p e r i o d of t i m e . At p r e s e n t , t h e r e s e r v e s of C u h a v e b e e n e s t i m a t e d 38 times, t h o s e of Ni e v e n 200 t i m e s h i g h e r t h a n t h e a c t u a l c o n t i n e n t a l s u p p l i e s ( S c h n e i d e r , 1977). B a s e d on c o n s e r v a t i v e c a l c u l a t i o n s t h e r e s o u r c e s r e c o v e r a b l e from t h e Pacific n o d u l e b e l t h a v e b e e n e s t i m a t e d to a c c o u n t for 170 m i l l i o n tons of n i c k e l , 155 m i l l i o n tons of copper, 30 m i l l i o n tons of c o b a l t a n d e v e n 3.7 b i l l i o n tons of m a n g a n e s e (Pellerer, 1975). T h e e c o n o m i c signifi- c a n c e of t h e s e p o t e n t i a l s o u r c e s of r a w m a t e r i a l has b e e n r e c o g n i z e d a n d i n t e r n a t i o n a l a s s o c i a t i o n s h a v e b e e n e s t a b l i s h e d (Hubred, 1975; M e r o , 1977; S c h n e i d e r , 1977). S u c h c o n s o r t i a are r e q u i r e d to d e v e l o p a p p r o p r i a t e m i n i n g t e c h n o l o g i e s as w e l l as to s h a r e t h e c o n s i d e r a b l e e c o n o m i c a l risks. M e a n w h i l e , d i f f e r e n t p i l o t p r o j e c t s for t e s t i n g t h e d e e p - s e a m i n i n g s y s t e m s w i t h m i n i n g ships s u c h as " S e d c o 445" w e r e u n d e r t a k e n success- fully d u r i n g 1978 {Summerer, 1978). T h e r e s e e m s to b e no d o u b t t h a t w e m a y e x p e c t d e e p - s e a m i n i n g on a l a r g e s c a l e w i t h i n t h e n e x t 10-30 years.
444 C. Schfitt & J. C. G. Ottow DISTRIBUTION AND C O M P O S I T I O N
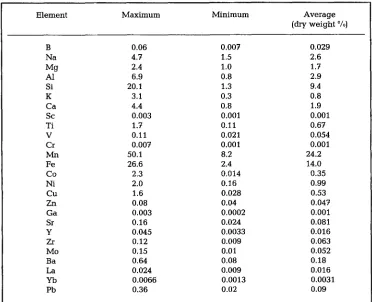
M a n g a n e s e n o d u l e s are w i d e l y d i s t r i b u t e d (25-80 % coverage) o n the b o t t o m of the d e e p sea, p a r t i c u l a r l y i n the Pacific Ocean. G e n e r a l l y s p e a k i n g , t h e y m a y occur at almost all d e p t h s a n d l a t i d u d e s b o t h i n the o c e a n s a n d i n c e r t a i n f r e s h - w a t e r lakes. However, the most a b u n d a n t deposits are f o u n d o n the surface of t h e p e l a g i c s e d i m e n t s i n the d e e p s e a (Hubred, 1975; C r o n a n , 1978). T h e s e areas of g e n e s i s are c h a r a c t e r i z e d b y (a) a n e x t r e m e l y low s e d i m e n t a t i o n rate, (b) a r e l a t i v e l y low b i o l o g i c a l p r o d u c t i v i t y a n d (c) b y a c o n t i n u o u s s u p p l y of o x y g e n b y (arctic) b o t t o m waters. T h e s e features are u s u a l l y restricted to the c e n t r a l r e g i o n s of o c e a n s far from t e r r i g e n i c (organic) supply. It is only at a l o w m i n e r a l i z a t i o n rate, a restricted s u p p l y of M n a n d Fe, a n d a p e r m a n e n t a e r o b i c e n v i r o n m e n t {reflected b y the r e l a t i v e l y h i g h Eh of + 400 to + 565 mV) that n o d u l e s m a y d e v e l o p at a slow, b u t s t e a d y a c c r e t i o n rate (a few ram/106 years). T h e s e ecological p r e r e q u i s i t e s m a y e x p l a i n w h y t h e most a b u n d a n t (and e c o n o m i c a l l y i n t e r e s t - ing) deposits h a v e b e e n r e c o r d e d n o r t h a n d south of the e q u a t o r i n t h e Pacific Ocean. So far, the m i n i m u m a b u n d a n c e of n o d u l e s r e q u i r e d to start m i n i n g has b e e n e s t i m a t e d as Table 1. Survey of the average dry-weight percentages of 26 different elements*) in 54 manganese
nodules collected from the Pacific Ocean
Element Maximum Minimum Average
(dry weight ~
B 0.06 0.007 0.029
Na 4.7 1.5 2.6
Mg 2.4 1.0 1.7
AI 6.9 0.8 2.9
Si 20.1 1.3 9.4
K 3.1 0.3 0.8
Ca 4.4 0.8 1.9
Sc 0.003 0.001 0.001
Ti 1.7 0.11 0.67
V 0.11 0.021 0.054
Cr 0.007 0.001 0.001
Mn 50.1 8.2 24.2
Pe 26.6 2.4 14.0
Co 2.3 0.014 0.35
Ni 2.0 0.16 0.99
Cu 1.6 0.028 0.53
Zn 0.08 0.04 0.047
Ga 0.003 0.0002 0.001
Sr 0.16 0.024 0.081
Y 0.045 0.0033 0.016
Zr 0.12 0.009 0.063
Mo 0.15 0.01 0.052
Ba 0.64 0.08 0.18
La 0.024 0.009 0.016
Yb 0.0066 0.0013 0.0031
Pb 0.36 0.02 0.09
8000 tons/km 2 with metal contents (weight % of dry material) of 1.25-1.40 % Ni, 0.05-1.35 % Cu, 0.18-0.25 % Co and 25-28 % Mn (Pellerer, 1975~ Cronan, 1978). These trace e l e m e n t s are intimately intermixed with the concentric layered amorphous and (crypto)-crystalline Mn (IV) and Pe (III) oxides as well as with other Mn and Be phases (Crerar & Barnes, 1974; Hubred, 1975; Priedrich et al., 1977). The interior of the manganese nodules is characterized by high porosity (Halbach et al., 1975) and a great variety of small-scale columnar cups orientated with their long axes in the radial plane of the nodules (Hubred, 1975~ Cronan, 1978). This high porosity and internal surface area makes post-depositional redissolution and migration of metals possible. Because of the lower activation energy required to reduce Mn (IV) compounds (mainly birnes- site = delta-MnO 2 and todorokite) compared with the amorphous and crystalline Fe (III) oxides (McKenzie, 1972~ Crerar & Barnes, 1974), we may expect the Mn (IV) components to be dissolved much more rapidly and earlier than the corresponding Fe (III) structures. Such a selective dissolution of different constituents would disintegrate the nodule considerably. Indeed, such internal "erosional features" have been observed repeatedly (Hubred, 1975; Cronan, 1978).
ROLE OF MICROORGANISMS IN GENESIS
In general, areas with high nodule densities have lower metal contents and vice versa. However, nodules of a single restricted region may also differ considerably in their chemical composition, although some remarkable relationships between certain elements have been observed (Friedrich et al., 1973~ Marchig and Gundlach, 1976~ Friedrich et al., 1977~ Halbach et al., 1979). Thus, Fe, Mg (and Co) on the one hand, and Mn, Ni, Cu and Co on the other, are usually well correlated. The highly significant correlation between Mn and Ni (with a Mn/Ni ratio of 23) indirectly supports the view that positively charged ions (such as Ni +) are adsorbed physico-chemically onto the primarily negatively loaded Mn colloids by extracting these cations from the sea water. Although this scavenging theory of Goldberg {1954) has been generally accepted (Crerar & Barnes, 1974), the mechanism of Mn (IV) oxide formation has not yet been completely elucidated. This is caused by the fact that Mn (II) is both undersaturated (with an average concentration of 2 ppb) and chemically stable (at pH 7.7-8.1 and Eh values between
+400 to 565 mV) (Crerar & Barnes, 1974~ Hartmann et al., 1975; Miiller, 1977) at the
446 C. Schiitt & J. C. G. Ottow
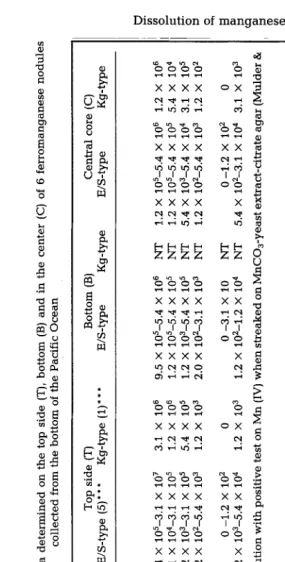
Schiitt, 1979). T h e s e s t u d i e s further clearly d e m o n s t r a t e d the p r e s e n c e of a great n u m b e r of bacteria, p o t e n t i a l l y c a p a b l e of p r e c i p i t a t i n g M n (II) to M n (IV}, both i n a n d o n the n o d u l e s as w e l l as i n the s u r r o u n d i n g w a t e r a n d s e d i m e n t s . However, n o n e of the isolated o r g a n i s m s d e v e l o p e d w i t h M n (II) only, b u t all p r e c i p i t a t e d M n (IV) r e a d i l y i n the p r e s e n c e of a low a m o u n t of glycerol as the c a r b o n source (Schiitt, 1979}. A p o r t i o n of the m a n g a n e s e - p r e c i p i t a t i n g b a c t e r i a (such as Vibrio spp.) e v e n d i s s o l v e d n o d u l e m a t e r i a l b y u s i n g M n (IV) as a n a l t e r n a t i v e e l e c t r o n acceptor if the a m o u n t of o r g a n i c c a r b o n w a s i n c r e a s e d significantly. O b v i o u s l y , it is the a m o u n t of d e g r a d a b l e o r g a n i c m a t t e r r a t h e r t h a n the a c t u a l p r e s e n c e of t h e s e b a c t e r i a that d e t e r m i n e w h e t h e r M n (II} p r e c i p i t a t i o n or M n (IV) r e d u c t i o n w i l l occur.
ROLE OF O R G A N I C SUPPLY
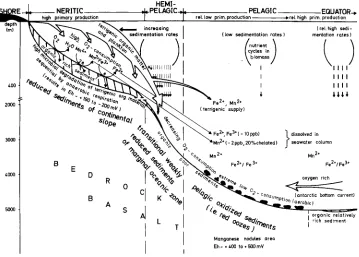
In F i g u r e 1 the c o n d i t i o n s a n d processes i n the n e r i t i c a n d p e l a g i c s e d i m e n t s are s u m m a r i z e d a n d compared. T h e r e is n o d o u b t that the differences are c a u s e d p r i m a r i l y b y the s i g n i f i c a n t v a r i a t i o n i n the s u p p l y of d e g r a d a b l e o r g a n i c matter. I n the n e r i t i c zone, a c o n t i n u o u s s u p p l y of o r g a n i c m a t t e r (due to c o n t i n e n t a l e r o s i o n a n d h i g h p r i m a r y
.~J=LQRE ~ NERITIC ~ l l h , J high primary production
depth
(m)
h00
2000
3000
&000
5OOO
HEM]-
PELAGIC j PELAGIC EO.UATOR-~
rel. Low prim.production I~reLhigh prim, production
~ 4 1 m ~ = increasing ( reL high sedl-
~eote~ sedimentation rates (Low sedimentation rates) mentatlon rates}
% 6
L ' N . b "+ ~176 +o,. in
, , , , ,
~O~ @ ~'A er~ 9 t
~' . , . ~
; ~ - ~ ' - . & 1
~
i ~ + . + +" ~ , ~ : ",~O,o ratio n " " ~ote~t ~ Fe2§ Mn 2.
"'~/'/t~ Of ~ ~ ~ % ~ I ( te:igenic supp[y )
0 . ' ~ ~'~1 ~>~0~ L "~Mn 2 r c
,~ "o+ "%, ~ \ +.
"0,~ v ~/ ~ 0o0 Mn2+ Mn2*
B E ~ ^ ~(~.-~e. +'~"+%- Fe2+/Fe 3+ Fe2+/F, 3+
V'f ~' (~. ~ o r oxygen rich
o
~ %
0 "&" zb ++ o <'4
C i/ 0 . ~ X ~ / ~ ' O , + ' ~ 0 2 " c o o s ~ [antarctic bottom current)
B I r~ ~/)0 "YI~ ,~,. ~ u r n _ _ _ pt'~176 laerobic}
A c I (i.v~'/~i>_ ~ - -
'J AJ '~. /-~,. <e(~ I I organi~c- reiotive~[y
L - v ct... ~eq//.~_ ' rich sediment
I T "UZe~ } 'neOt s i
Manganese nodules oreo
i Eh- *400 to +500mV
Fig. 1. Fate of the Mn and Fe compounds in the neritic zone compared with the pelagic sediments. In the neritic zone Mn (IV) and Fe (III) oxides (and hydroxides) are reduced microbiologically by acting as alternative electron acceptors. In the pelagic sediments far from the continental slope Mn (If) and Fe (II, III) compounds are immobilized (nodule genesis) by chemical and/or microbial activity9 These opposite processes are essentially caused by the significant differences in organic
448 C. Schiitt & J. C. G. O t t o w
p r o d u c t i o n ) w i l l l e a d to i n t e n s i v e m i n e r a l i z a t i o n p r o c e s s e s . A s soon as o x y g e n h a s b e e n r e s p i r e d , a e r o b i c a n d a n a e r o b i c b a c t e r i a s w i t c h to nitrate, M n (IV) a n d F e (III) a s a l t e r n a t i v e e l e c t r o n a c c e p t o r s in o r d e r to c o n t i n u e t h e i r e n e r g y c o n s e r v i n g r e a c t i o n s (ATP synthesis) (Trimble & Ehrlich, 1968; O t t o w & G l a t h e , 1973; M u n c h & Ottow, 1977; M u n c h et al., 1978). T h e s e p r o c e s s e s m a y b e s u m m a r i z e d as follows.
B i o l o g i c a l o x i d a t i o n (-- d e h y d r o g e n a t i o n ) :
m e t a b o l i c a c t i v i t y H +
o r g a n i c m a t t e r ~ 4- e 4- b i o m a s s 4- A T P 4- p r o d u c t s
d e h y d r o g e n a s e s
M n (IV) or F e (III) o x i d e s as H - a c c e p t o r s ( h y d r o g e n a t i o n b y r e d u c t a s e s ) ;
M n O 2 4- 2e § 4 H + ~ M n (II) § 2I-I20
F e ( O H ) 3 § e 4 - 3 H + ) F e ( I I ) §
O x y g e n d e p l e t i o n a n d t h e a c c u m u l a t i o n of r e d u c e d p r o d u c t s a r e r e s p o n s i b l e for t h e r a p i d d r o p in Eh (in t h e r a n g e of § 190 to 220 mV). It is i m p o r t a n t to r e a l i z e t h a t M n (IV} a n d Fe (III) are r e d u c e d d i r e c t l y b y m e t a b o l i c a c t i v i t y r a t h e r t h a n i n d i r e c t l y b y a l o w e r e d r e d o x p o t e n t i a l . A d r o p in Eh is the r e s u l t of m i c r o b i a l r e d u c t i o n p r o c e s s e s a n d not a m e c h a n i s m of r e l e v a n t c h e m i c a l t r a n s f o r m a t i o n s . Its s i g n i f i c a n t r o l e is r e s t r i c t e d to t h e c r e a t i o n of a c e r t a i n r e d o x l e v e l t h a t m a y s u p p o r t t h e s t a b i l i t y of M n (II) or Fe (II) c o m p o u n d s .
B e y o n d a t r a n s i t i o n a l a r e a (with m i n o r or m o d e r a t e o r g a n i c s u p p l y a n d t h u s r e l a t i v e l y l o w m i c r o b i o l o g i c a l activity) w e u s u a l l y f i n d t h e c e n t r a l p e l a g i c z o n e (red oozes) w i t h e x t r e m e l y l o w o r g a n i c s e d i m e n t a t i o n r a t e s (Volkov et al., 1975). T h e s e a r e a s w i t h e x t e n d e d f i e l d s of m a n g a n e s e n o d u l e s are r e l a t i v e l y p o o r in o r g a n i c m a t t e r ( 0 . 1 - 0 . 3 % o r g a n i c carbon) (Mfiller, 1977), strictly a e r o b i c a n d c h a r a c t e r i z e d b y e x t r e m e l y l o w o x y g e n - c o n s u m p t i o n rates. T h e r e s t r i c t e d m i c r o b i a l a c t i v i t y is s u p p o r t e d b y l o w t e m p e r a t u r e s ( n e a r l y -- 1 ~ a n d t h e e n o r m o u s h y d r o s t a t i c p r e s s u r e (5000-m d e p t h c o r r e s p o n d s to 500 bar). Due to t h e l o w b a c t e r i a l activity, t h e r e d o x p o t e n t i a l r e m a i n s at l e v e l s of 4-400 a n d 4-565 m V (Miiller, 1977). S u c h c o n d i t i o n s s h o u l d b e c o n s i d e r e d as t h e m a i n p r e r e q u i s i t e for n o d u l e formation.
T h e e q u a t o r r e g i o n ,
however,
is c h a r a c t e r i z e d b y r e l a t i v e l y h i g h p r i m a r y p r o d u c t i o n a n d thus h i g h e r s e d i m e n t a t i o n r a t e s of o r g a n i c m a t t e r w h i c h r e s u l t in e n h a n c e d m i c r o - b i a l a c t i v i t y a n d r e d u c t i o n p r o c e s s e s . C o n s e q u e n t l y no n o d u l e s m a y b e g e n e r a t e d in this p a r t i c u l a r a r e a of the c e n t r a l ocean.N O D U L E LYSIS A S A N E N V I R O N M E N T A L I M P A C T
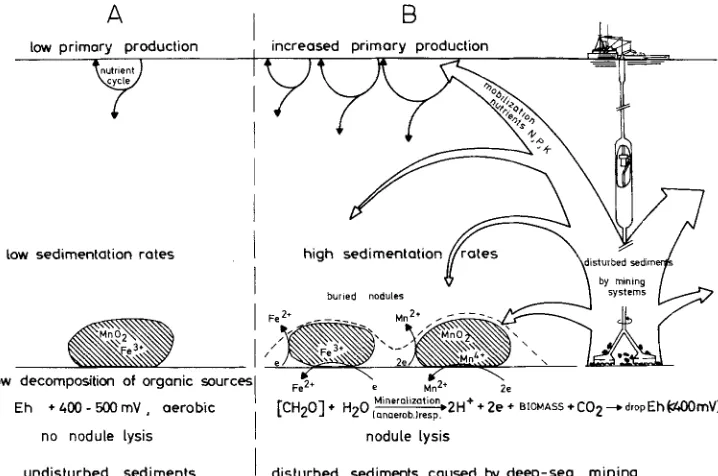
b l o o m i n the surface r e g i o n s ( i n c r e a s e d p r i m a r y production). U n m i n e d m a n g a n e s e n o d u l e s w i l l b e b u r i e d b y s e t t l i n g of t h e s u s p e n d e d m a t e r i a l , w h i c h is s i g n i f i c a n t l y e n r i c h e d b y p h y t o p l a n k t o n i c debris. This a d d i t i o n a l e n e r g y s u p p l y will i n c r e a s e the activity of the m i c r o o r g a n i s m s a n d the rate of o x y g e n c o n s u m p t i o n . B e c a u s e of o x y g e n d e p l e t i o n , the b a c t e r i a will switch to other e l e c t r o n acceptors such as NO~, M n (IV) a n d Fe (III) c a u s i n g d e n i t r i f i c a t i o n as w e l l as the r e d u c t i v e d i s s o l u t i o n of m a n g a n e s e n o d u l e s (Fig. 2B). T h e h i g h e r the a m o u n t of d e c o m p o s a b l e o r g a n i c debris, the more i n t e n s i v e a n d c o m p l e t e is t h e lysis of b u r i e d m a n g a n e s e n o d u l e s i n the s e d i m e n t . T h e s e i n t e n s i v e m i n e r a l i z a t i o n processes cause the Eh to drop rapidly. At this stage, the processes a n d c o n d i t i o n s i n the u p p e r layers of the d i s t u r b e d s e d i m e n t s are c o m p a r a b l e w i t h the n e r i t i c s i t u a t i o n at the c o n t i n e n t a l slope. A l t h o u g h the m o b i l i z e d M n (II) a n d Fe (II) c o m p o u n d s m a y b e r e p r e c i p i t a t e d i n the a e r o b i c a r e a n e a r the h a u l i n g p l a n t , the fate of the l i b e r a t e d trace e l e m e n t s is u n k n o w n .
A
low primary production
nutrient
tow s e d i m e n t a t i o n r o t e s
tow decomposition of organic sources Eh +400-500mV, aerobic
no nodule [ysis
undisturbed sediments
B
hincreased primary production ~ . . , ~ .
h i " "
Fe 2* e Nn 2. 2e
Miner olizotion §
[CH20]+ H20 anoerobJresp. ~. 2H +2e+mO~ass+CO2--*dropEh(,~d?OmV) nodule [ysis
d i s t u r b e d s e d i m e n t s c a u s e d by d e e p - s e a m i n i n g
Fig. 2. The lysis of manganese nodules as a possible result of deep-sea mining. Because of mining activity (B), sediments are disturbed, nutrients become mobilized and primary production will occur. Nodules next to the hauling plant are buried by resettled sediments enriched with organic debris. In the following intensive mineralization processes oxygen may become exhausted, and microorganisms will switch to Mn (IV) and Fe (III) oxides as alternative electron acceptors in order
to continue their energy-conserving reactions (ATP synthesis by anaerobic respiration)
4 5 0 C. S c h i i t t & J. C. G. O t t o w
m e n t a l l y e x a m i n e d b e f o r e d e e p - s e a m i n i n g is s t a r t e d . T h e " w a t e r d e s e r t " of t h e c e n t r a l
P a c i f i c is - l i k e m o s t o t h e r l i v i n g s y s t e m s - t h e r e s u l t of a s e n s i t i v e b i o l o g i c a l b a l a n c e . W e s h o u l d l e a r n i n f u t u r e t o c o n s i d e r e c o l o g i c a l k n o w l e d g e a s a n e s s e n t i a l p r e r e q u i s i t e f o r r e s p o n s i b l e e n v i r o n m e n t a l m a n a g e m e n t .
Acknowledgements. We gratefully a c k n o w l e d g e the skillful technical assistance of Miss. E. Feier- tag. This w o r k was s u p p o r t e d by a r e s e a r c h grant from the Deutsche Forschungsgemeinschaft, Bonn
- Bad Godesberg, Germany.
L I T E R A T U R E C I T E D
Amos, A. F., Roels, O. A., Garside, C., Malone, T. C. & Paul, A. Z., 1977. Environmental aspects of nodule mining. In: Marine m a n g a n e s e deposits. Ed. by G. P. Glasby. Elsevier, Amsterdam, 391-437.
Butkewitsch, W. S., 1928. Die Bildung der E i s e n m a n g a n a b l a g e r u n g e n am M e e r e s b o d e n u n d die daran b e t e i l i g t e n Mikroorganismen. - Trudy morsk, nauch. Inst. 3, 63-80.
Crerar, D. A. & Barnes, H. L., 1974. Deposition of d e e p - s e a nodules. - Geochim. cosmochim. Acta 38, 279-300.
Cronan, D. S., 1977. D e e p sea nodules: Distribution a n d geochemistry. In: Marine m a n g a n e s e deposits. Ed. by G. P. Glasby. Elsevier, Amsterdam, 11-44.
Cronan, D. S., 1978. M a n g a n e s e nodules: controversy u p o n controversy. - Endeavour 2, 80-84. Ehrlich, H. L., Ghiorse, W. C. & Johnson, G. L., 1972. Distribution of microbes in m a n g a n e s e nodules
from the Atlantic a n d Pacific Oceans. - Devs ind. Microbiol. 13, 57-65.
Fellerer, R., 1975. Some technical a n d economical aspects of d e e p s e a mining. - De I n g e n i e u r 87, 634-661.
Friedrich, G. H., Kunzendorf, H. & Pliiger, W. L., 1973. Geochemical investigations of d e e p sea m a n g a n e s e n o d u l e s from the Pacific on board R/V Valdivia. A n application of the EDX- technique. In: The origin a n d distribution of m a n g a n e s e nodules in the Pacific a n d prospects for exploration. Ed. by M. Morgenstein. Univ. Press. Honolulu, Hawaii, 31-44.
Friedrich. G., Pliiger, W., Kunzendorf, H., Roonwal, G., Schmitz-Wiechoski, A., Giirkan, A., Zuleta J. R. & KrSmer, E., 1977. Studies on the g e o c h e m i s t r y a n d g e n e t i c interpretation of m a n g a n e s e n o d u l e deposits. In: Natural resources a n d development. Ed. by Inst. Scient. Coop. Conj. Federal Inst. Geosciences Nat. Res. Laupp & G6bel, Tfibingen, 6, 17-47.
Goldberg, E. D., 1954. Marine geochemistry. - J. Geol. 62, 249-265.
Halbach, P., Ozkara, M., & Hense, d., 1975. The influence of metal content on the physical a n d mineralogical properties of p e l a g i c m a n g a n e s e nodules. - Miner. Deposita 10, 397-411. Halbach, P., Rehm, E. & Marchig, V., 1979. Distribution of Si, Mn, Fe, Ni, Co, Zn, Pb, Mg, a n d Ca in
grain-size fractions of s e d i m e n t s a m p l e s from a m a n g a n e s e n o d u l e field in the central pacific O c e a n . - Mar. Geol. 29, 237-252.
Hartmann, M., KSgler, F. C. Miiller, P. & Suess, E., 1975. Ergebnisse der U n t e r s u c h u n g e n zur G e n e s e der M a n g a n k n o l l e n . ForschBer. Bundesmin. Forsch. Technol. M 75-02, 60-113. Hubred, G., 1975: D e e p - s e a m a n g a n e s e nodules: A r e v i e w of the literature. - Miner. Sci. E n g n g 7,
71-85.
Kuznetsov, S. I., 1975. The role of microorganisms in the formation of lake bottom deposits a n d their diagenesis. - Soil Sci. 119, 81-88.
McKenzie, R. M. 1972. The m a n g a n e s e oxides in softs. - Z. Pflanzenernaehr. Bodenkd. 131, 221-242.
Marchig, V. & Gundlach, H., 1976. Zur G e o c h e m i e yon M a n g a n k n o i i e n aus d e m Zentralpazifik u n d ihrer S e d i m e n t u n t e r l a g e . - Geol. Jb. (D) 16, 59-77.
Mero, d. L, 1962. O c e a n - f l o o r - m a n g a n e s e nodules. - Econ. Geol. 57, 747-767.
Mero, J. L. 1977. Economic aspects of n o d u l e mining. In: M a r i n e m a n g a n e s e deposits. Ed. by. G. P. Glasby. Elsevier, Amsterdam, 327-355.
Mulder, B. G. & V a n Veen, W. L., 1963. Investigations on the Sphaerotilus-Lepthotrix group. - Antonie v a n L e e u w e n h o e k 29, 121-153.
Miiller, P. J., 1977. C/N-ratios on Pacific d e e p - s e a sediments: Effect of inorganic a m m o n i u m a n d organic n i t r o g e n c o m p o u n d s s o r b e d by clays. - Geochem. cosmochim. Acta 41, 765-776. Munch, J. C. & Ottow, J. C. G., 1977. M o d e l l u n t e r s u c h u n g e n z u m M e c h a n i s m u s der b a k t e r i e l l e n
]Eisenreduktion in h y d r o m o r p h e n BSden. - Z. P f l a n z e n e m a e h r . Bodenkd. 140, 549-562. Munch, J. C., Hillebrand, T. & Ottow, J. C. G., 1978. Transformations in the Feo/Fe a ratio of
p e d o g e n i c iron oxides affected by i r o n - r e d u c i n g bacteria. - Can. J. Soil Sci. 58, 475-486. Nealson, K. H. 1978. The isolation a n d characterization of marine bacteria w h i c h catalyze m a n g a n -
ese oxidation. In: E n v i r o n m e n t a l b i o g e o c h e m i s t r y a n d geomicrobiology. Ed. b y W. E. Krumbein. A n n Arbor Sci. Publ., Michigan, 3, 847-857.
Ottow, J. C. G & Glathe, H. 1973. P e d o c h e m i e u n d Pedomikrobiologie h y d r o m o r p h e r B6den; V o r a u s s e t z u n g u n d U r s a c h e n der Eisenreduktion. - C h e m i e Erde 32, 1-44.
Schneider, J., 1977. G e o w i s s e n s c h a f t l e r u n d ihre V e r a n t w o r t u n g fiir die m e n s c h l i c h e Gesellschaft. - Geol. Rdsch. 66, 740-755.
Schiitt, C., 1979. U n t e r s u c h u n g e n zur q u a n t i t a t i v e n u n d qualitativen Mikroflora yon M a n g a n k n o l - l e n aus d e m z e n t r a l e n Pazifik sowie zur B e d e u t u n g Mangan-(II)-f~llender Bakterien fiir ihre G e n e s e . Hochschul Verl. Freiburg, 1-116.
Schiitt, C. & Ottow, J. C. G., 1977. Mesophilic a n d psychrophilic m a n g a n e s e - p r e c i p i t a t i n g bacteria in m a n g a n e s e n o d u l e s of the Pacific Ocean. - Z. allg. Mikrobiol. 17, 611-816.
Schiitt, C. & Ottow, J. C. G., 1978. Distribution a n d identification of m a n g a n e s e - p r e c i p i t a t i n g bacteria from n o n c o n t a m i n a t e d f e r r o m a n g a n e s e nodules. In: Environmental b i o g e o c h e m i s t r y a n d geomicrobiology. Ed. by W. E. Krumbein. A n n Arbor Sci. Publ., Michigan, 3, 869-878. Summerer, 0., 1978. M a n g a n k n o l l e n - Die r i e s i g e n Erzvorrfite der Tiefsee w e r d e n jetzt angegrif-
fen. - Bild. Wiss. 8, 50-59.
Thiel, H., 1975. Die 6kologische Erforschung der Tiefsee. - U m s c h a u 75, 716.
Thiel, H., 1978. The faunal e n v i r o n m e n t of m a n g a n e s e n o d u l e s a n d aspects of d e e p s e a time scales. In: E n v i r o n m e n t a l b i o g e o c h e m i s t r y a n d geomicrobiology, l~d. b y W. E. Krumbein. A n n Arbor Sci. Publ. M i c h i g a n 3, 887-896.
Trimble, R. B. & Bhrlich, H. L., 1968. Bacteriology of m a n g a n e s e nodules, III. Reduction of M n O 2 by two strains of n o d u l e bacteria. - Appl. Microbiol. 16, 695-702.