0095-1137/06/$08.00⫹0 doi:10.1128/JCM.00934-06
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Candida albicans
Strain Maintenance, Replacement, and Microvariation
Demonstrated by Multilocus Sequence Typing
F. C. Odds,
1* A. D. Davidson,
1M. D. Jacobsen,
1A. Tavanti,
2J. A. Whyte,
1C. C. Kibbler,
3D. H. Ellis,
4M. C. J. Maiden,
5D. J. Shaw,
1and N. A. R. Gow
1Aberdeen Fungal Group, Institute of Medical Sciences, Aberdeen AB25 2ZD, United Kingdom1; Department of Biology, University of
Pisa, Pisa, Italy2; Department of Medical Microbiology, Royal Free and University College Medical School, Royal Free Hospital,
London, United Kingdom3; Department of Microbiology and Immunology, Australian Candidaemia Study, Women’s and
Children’s Hospital, North Adelaide, SA, Australia4; and Peter Medawar Building for Pathogen Research and
Department of Zoology, University of Oxford, Oxford, United Kingdom5
Received 4 May 2006/Returned for modification 18 May 2006/Accepted 24 July 2006
We typed 165Candida albicansisolates from 44 different sources by multilocus sequence typing (MLST) and ABC typing of rRNA genes and determined their homozygosity or heterozygosity at the mating-type-like locus (MTL). The isolates represented pairs or larger sets from individual sources, which allowed the determination of strain diversity within patients. A comparison of replicate sequence data determined a reproducibility threshold for regarding isolates as MLST indistinguishable. For 36 isolate sets, MLST and ABC typing showed indistinguishable or highly related strain types among isolates from different sites or from the same site at different times from each patient. This observation included 11 sets with at least one isolate from a blood culture and a nonsterile site from the same patient. For one patient, strain replacement was evidenced in the form of two sets of isolates from different hospital admissions where the strain types within each set were nearly identical but where the two sets differed both by MLST and ABC typing. MLST therefore confirms the existing view of C. albicans strain carriage. Microvariation, evidenced as small differences between MLST types, resulted in most instances from a loss of heterozygosity at one or more of the sequenced loci. Among isolate sets that showed major strain type differences, some isolates could be excluded as likely examples of handling errors during storage. However, for a minority of isolates, intermittent differences in ABC type for tightly clustered MLST types and intermittent appearances of MTL homozygosity lead us to propose that someC. albicansisolates, or all isolates under yet-to-be-determined conditions, maintain a high level of genetic diversity by mechanisms such as recombination, gene conversion, or chromosomal ploidy change.
Candida albicansis a normal commensal of the human gut
microflora that can cause invasive superficial and disseminated infections in immunologically susceptible hosts (11). A number of different technical approaches to typingC. albicansisolates have been developed since the early 1980s (78). ForC. albi-cans, as for all microbial pathogens, a reproducible and dis-criminatory strain typing system is of benefit for clinical and epidemiological studies to provide information on sources, car-riage, and transmission of infection and on relations between strain types and properties such as virulence and antimicrobial resistance. In many publications, multipleC. albicansisolates from longitudinal surveys of patients or from surveillance cul-tures of different anatomical sites have been typed. Most work of this nature has confirmed a tendency towards the persis-tence of a unique strain in each human host, which was sug-gested even in the earliest studies based on phenotypic strain typing (45, 53). Minor variations in genetically determined strain types in surveys of multiple C. albicans isolates from individual patients have been described as “microevolution” (33, 35, 37, 64, 65, 76, 78) or genotypic shuffling (37, 69). (This is not the usual sense in which the term “microevolution” is used by population geneticists.) Evidence from strain typing work confirms the view ofC. albicansas a species that
repro-duces predominantly in a clonal manner but with a tendency for occasional genetic variation that may arise by mechanisms such as recombination, gene replacement, or cryptic mating (22–24, 29, 30, 34, 38, 39, 42, 50, 66, 79, 80, 84, 98).
Most longitudinal and/or multiple-site epidemiological work demonstrating strain maintenance inC. albicanshas been done with superficial isolates, particularly with cultures from the oral cavities of human immunodeficiency virus (HIV)-infected pa-tients (2, 3, 6, 19, 31, 48, 51, 61, 71, 72, 90, 96), with vaginal isolates (1, 37, 49, 64, 65, 73, 76, 91), with other sources of superficial isolates (4, 17, 74, 97), or with superficial surveil-lance cultures from hospitalized patients (60, 81, 89). Some surveys of oral isolates have shown the same strain persisting for months to years (4, 17). Occasionally, authors have sug-gested that multiple strain types coexist in samples from some patients (27, 69, 83), but such findings are in the minority and may reflect technical sensitivities and variabilities in the typing methodologies used in some cases. The epidemiology of dis-seminated C. albicans infection has been studied much less often than that of superficial sites, but the results of surveys based on karyotype electrophoretic patterns (62, 63, 93), ran-domly amplified PCR fragments (92), DNA fingerprinting with oligonucleotide probes (44, 67), and multilocus enzyme elec-trophoresis (12) consistently indicate that isolates from blood are highly similar to, or indistinguishable from, isolates from superficial sites in the same patient.
The overall picture ofC. albicanscolonization and infection * Corresponding author. Mailing address: Aberdeen Fungal Group,
Institute of Medical Sciences, Aberdeen AB25 2ZD, United Kingdom. Phone and fax: 44 01224 555828. E-mail: [email protected].
3647
on May 16, 2020 by guest
http://jcm.asm.org/
is therefore mainly of clonal reproduction of strains, infection by the spread of endogenous, colonizing strain types, but with sporadic changes at the level of sequences of individual DNA fragments, which we shall call microvariation rather than mi-croevolution. Microvariation refers to small but detectable changes in DNA sequences among isolates obtained longitu-dinally or as separate clones from individual patients (35, 65). Although the original reports of microvariation suggested that changes arose from the reorganization of repetitive DNA se-quences in the genome, later work has consistently associated the loss of heterozygosity (LOH) as a common mechanism for microvariation changes (16, 21, 70, 85, 88). LOH may result from chromosome deletion or loss, recombination, and/or gene conversion events. Conjugational mating and nuclear fu-sion without subsequent meiosis can occur betweenC. albicans
cells (24, 34, 42), and such mating is dependent on homozygous alleles, i.e., LOH, at the mating-type-like locus (MTL) on chromosome 5 (42, 79).
Multilocus sequence typing (MLST) is a highly discrimina-tory and portable approach to distinguishing strains within a microbial species (43). DNA sequences from six or seven gene fragments are compared to establish levels of similarity be-tween isolates and results, expressed with unique numbers for individual sequences (genotypes) and for diploid sequence types (DSTs), unique combinations of the seven genotypes in each isolate (43). Several statistical procedures have been de-vised for the analysis of MLST data, including eBURST (20), which compiles clusters of isolates that differ from each other at only one of the sequenced loci. MLST schemes are now available for several fungi, and the approach is well developed for the typing ofC. albicansstrains (7–9, 68, 84, 86). The C.
albicansdata are stored on a central internet database (http:
//test1.mlst.net/) to allow free access to investigators. Results from MLST are comparable with those obtained by DNA fingerprinting with the moderately repetitive sequence Ca3, since both typing approaches assign the same sets of isolates to the same clusters of highly related strains, with the exception of DNA fingerprinting clade E, which is split between two MLST clades (84).
In the course of our ongoing work to determine DSTs by MLST for the analysis of C. albicans population structures (84), we have now typed sets of multiple isolates of the fungus from 44 different sources, including instances of superficial and blood isolates from the same patient, isolates that have been maintained and passaged through different culture collections, and isolates that have been experimentally exposed to flucon-azole. We have determined the level of sequencing errors in replicate experiments to establish criteria for regarding isolates as indistinguishable. We have looked for evidence of micro-variation changes and LOH in these isolates and in isolates recovered from experimental infections. In addition to MLST, we have typed the isolates for the presence or absence of an intron in the internal transcribed spacer 1 (ITS1) region of DNA sequences encoding rRNA (ABC typing) (46, 47) and for homozygosity or heterozygosity at the MTL (86). These extra data augment the MLST results by providing extra typing in-formation for each isolate. Our findings confirm the prevailing views of strain maintenance, predominant clonality, and occa-sional microvariation in C. albicans and that isolates from blood cultures match those from nonsterile sites in the same
patient. We hypothesize that LOH may occur among some but not all cells in the population colonizing or infecting a patient so that heterozygosity remains established in a proportion of the population. Our data also raise some caveats concerning the maintenance and distribution of isolates within and be-tween laboratories.
MATERIALS AND METHODS
C. albicansisolates.Details of the 165 isolates studied are shown in Table 1. In our laboratory, all the isolates were stored on beads in 50% glycerol at⫺80°C. For experimental use, a bead was placed onto a plate of CHROMagar Candida medium (M-Tech Diagnostics, Warrington, United Kingdom) and incubated at 37°C for 48 h. All the isolates formed green colonies on this medium, verifying their prior identification asC. albicans.
The isolates came from 44 separate sources and are grouped in numbered sets according to source in Table 1. Most of the isolates were pairs or triplet isolates from a human subject or, in one instance, an animal. The subjects included healthy volunteers, patients with symptomatic vaginitis, patients undergoing che-motherapy for hematological malignancy, and patients with proven candidemia. Many of the isolates came from previously published studies, as indicated in Table 1. The oldest isolates in the panel were originally cultured in the 1970s. Many of the isolates have been maintained in the first author’s collection of fungal strains, which was initiated in Leeds, United Kingdom, in 1973 and transported to Leicester, United Kingdom, and Beerse, Belgium, and which is now in Aberdeen, United Kingdom. With each move, all isolates in the collection were subcultured and restocked at least once (52, 57). The histories of these isolates are indicated in Table 1.
Some sets of isolates merit special comment. In set 1, isolate RV4688 was a strain used in experiments by J. Van Cutsem in Beerse, Belgium, that had become spontaneously avirulent (58). It was originally isolated in Zaire by R. Vanbreuseghem. Records at the culture collection of the Scientific Institute of Public Health (SIPH) (formerly the Institute of Hygiene and Epidemiology) in Brussels, Belgium, show that isolate IHEM3742 was received in 1988, having passed from R. Vanbreuseghem to J. Van Cutsem to the American Type Culture Collection (as strain ATCC 28516). IHEM16346 is not listed in the public SIPH catalog but came directly from the original Zaire isolate from R. Vanbreuseg-hem, according to SIPH collection manager N. Nolard (personal communica-tion). 81/005 was the number given to an isolate that was received by the first author from J. Van Cutsem in 1981. From their histories, these isolates should be indistinguishable by MLST. The isolates in set 15 share a similar complex history. An oral isolate from an AIDS patient in Leicester, United Kingdom, in 1988 was sent to J. Schmid at the University of Iowa and designated OD8826. A subculture of the isolate was taken by J. Schmid to Massey University in New Zealand, and the isolate was supplied from New Zealand for the present study. Two other oral isolates from the same patient in 1988 were removed to Beerse, Belgium, and were then added to the SIPH collection and were supplied as isolates IHEM20491 and IHEM20493 for the present study. The isolates in set 18, all originally from an AIDS patient in London, United Kingdom, had also been subcultured and relocated in different culture collections before inclusion in the present study. Set 23 comprises examples of isolates derived from T118 that had been passaged for a measured number of generations in the presence of flucon-azole (14, 16). For each isolate, the two digits after the letter “D” indicate the subclone number from T118, and the three final digits indicate the number of generations. Finally, set 44 comprises the isolates still available from a previous study in which examples of the widely usedC. albicansstrain 3153 were submit-ted from several laboratories for comparison by an earlier strain typing meth-odology (41).
Strain typing.The isolates were tested by MLST as previously described (9, 84). The ABC type (A, B, or C) and MTL types (a/␣,a/a, or␣/␣) were deter-mined by PCR as previously described (86). Similarities between sequence data were analyzed in terms ofpdistance with MEGA version 2.1 (28), and results with 500 bootstrap replications are depicted as a dendrogram by the unweighted-pair group method with arithmetic averages (UPGMA). The analyses were based on concatenated data from all known polymorphic sites, duplicated to allow the discrimination of homozygous and heterozygous differences (84). The sequences were also analyzed to determine clonal clusters by eBURST, version 2 (20). Clusters were defined as groups of isolates with six of the seven genotype sequences being identical.
When one or more isolates within a set of isolates from a common source differed from the rest by MLST or ABC type, the sequencing chromatograms for
on May 16, 2020 by guest
http://jcm.asm.org/
TABLE 1. Details ofC. albicansisolates studied
Isolate
set Isolate
Yr isolated
Country
of origina Isolated fromb: Patient detail(s)c Storage historyd DST
ABC type
MTL
type Reference
1 IHEM16346 1970s Zaire Kidney? May be parent of
IHEM3742
Be2 258 C a/␣
1 IHEM3742 1970s Zaire Kidney Unknown Be2 124 C a/␣
1 RV4688 1970s Kidney Originally
IHEM3742?
Be1 87 C a/␣ 58
1 81/005 Subculture of RV4688
received in 1981
Be1, UK2, Be1 480 C a/␣
1 RV2LK Recovered from
experimental mouse infection with RV4688
269 C a/␣
2 73/024 1973 UK Vagina Open-heart surgery UK1, UK2, Be1 145 A a/␣ 54
2 73/027 1973 UK Vagina Open-heart surgery UK1, UK2, Be1 485 A a/␣ 54
3 78/005 1978 UK Oropharynx Healthy volunteer UK2, Be1 96 A a/a
3 78/010 1978 UK Oropharynx Healthy volunteer UK2, Be1 517 A ␣/␣
4 81/066 1980 UK Oropharynx Female, STD clinic UK2, Be1 396 A a/␣
4 81/114 1980 UK Vagina Female, STD clinic UK2, Be1 396 A a/␣
5 90/141 1980 Spain Folliculitis Heroin addict UK2, Be1 547 A a/␣ 56
5 90/142 1980 Spain Abscess Heroin addict UK2, Be1 482 A a/␣ 56
6 L206 1985 UK Feces Patient 11 UK2, Be1 255 B a/␣ 55
6 L249 1985 UK Feces Patient 11 UK2, Be1 255 B a/␣ 55
6 L258 1985 UK Oropharynx Patient 11 UK2, Be1 255 B a/␣ 55
6 L332 1985 UK Feces Patient 11 UK2, Be1 255 B a/␣ 55
6 L343 1985 UK Oropharynx Patient 11 UK2, Be1 80 B a/␣ 55
6 L475 1985 UK Feces Patient 11 UK2, Be1 255 B a/␣ 55
6 L485 1985 UK Feces Patient 11 UK2, Be1 256 B a/␣ 55
6 L527 1985 UK Feces Patient 11 UK2, Be1 257 B a/␣ 55
6 L531 1985 UK Feces Patient 11 UK2, Be1 255 B a/␣ 55
6 L548 1985 UK Oropharynx Patient 11 UK2, Be1 255 B a/␣ 55
6 L586 1985 UK Feces Patient 11 UK2, Be1 255 B a/␣ 55
6 L607 1985 UK Oropharynx Patient 11 UK2, Be1 255 B a/␣ 55
7 L344 1985 UK Oropharynx Patient 5, March 1985 UK2, Be1 273 C a/␣ 55
7 L345 1985 UK Sputum Patient 5, March 1985 UK2, Be1 273 C a/␣ 55
7 L367 1985 UK Oropharynx Patient 5, March 1985 UK2, Be1 272 C a/␣ 55
8 L479 1985 UK Feces Patient 5, May 1985 UK2, Be1 125 A a/␣ 55
8 L562 1985 UK Sputum Patient 5, May 1985 UK2, Be1 271 A a/␣ 55
8 L572 1985 UK Stomach Patient 5, May 1985 UK2, Be1 271 A a/␣ 55
8 L580 1985 UK Feces Patient 5, May 1985 UK2, Be1 270 A a/␣ 55
9 L217 1985 UK Oropharynx Patient 8 UK2, Be1 89 B a/␣ 55
9 L252 1985 UK Oropharynx Patient 8 UK2, Be1 89 B a/␣ 55
9 L261B 1985 UK Oropharynx Patient 8 UK2, Be1 267 B a/␣ 55
9 L283 1985 UK Oropharynx Patient 8 UK2, Be1 268 B a/␣ 55
9 L289 1985 UK Oropharynx Patient 8 UK2, Be1 89 B a/a 55
9 L441 1985 UK Oropharynx Patient 8 UK2, Be1 89 B a/␣ 54
9 L463 1985 UK Feces Patient 8 UK2, Be1 89 B a/␣ 54
10 L121 1985 UK CSF UK2, Be1 141 B ␣/␣
10 L123 1985 UK Urine UK2, Be1 515 B a/␣
10 L124 1985 UK CSF UK2, Be1 542 B ␣/␣
11 L220 1985 UK Feces Patient 6 UK2, Be1 544 A a/␣ 55
11 L262 1985 UK Oropharynx Patient 6 UK2, Be1 545 A a/␣ 55
11 L353 1985 UK Oropharynx Patient 6 UK2, Be1 544 A a/␣ 55
12 85/005 1985 UK Blood UK2, Be1 83 A a/a 10
12 85/006 1985 UK Sputum UK2, Be1 541 A a/␣ 10
13 L513 1986 UK Sputum UK2, Be1 546 B a/␣
13 L516 1986 UK Blood UK2, Be1 134 B a/␣
14 L517 1986 UK Anus Male, STD clinic UK2, Be1 216 A a/␣
14 L518 1986 UK Oropharynx Male, STD clinic UK2, Be1 216 A a/␣
15 OD8826 1988 UK Oropharynx UK2, Iowa, NZ 24 A a/␣ 72
15 IHEM20491 1989 UK Oropharynx UK2, Be1, Be2 373 A a/␣ 72
15 IHEM20493 1990 UK Oropharynx UK2, Be1, Be2 372 A a/␣ 72
16 B59626 1991 Germany Oropharynx Be1 585 A a/␣
16 B59630 1991 Germany Oropharynx Be1 585 A a/␣
17 C19 1993 UK Sputum 429 B a/␣
17 C25 1993 UK Catheter tip 429 B a/␣
17 C26 1993 UK Blood 429 B a/␣
17 C27 1993 UK Catheter tip 429 B a/␣
18 IHEM20486 1993 UK Oropharynx AIDS patient UK2, Be1, Be2 309 B a/␣
Continued on following page
on May 16, 2020 by guest
http://jcm.asm.org/
TABLE 1—Continued
Isolate
set Isolate
Yr isolated
Country
of origina Isolated fromb: Patient detail(s)c Storage historyd DST
ABC type
MTL
type Reference
18 IHEM20487 1993 UK Oropharynx AIDS patient UK2, Be1, Be2 389 B a/␣
18 IHEM20488 1993 UK Oropharynx AIDS patient UK2, Be1, Be2 302 B a/a
18 J932570 1993 UK Oropharynx AIDS patient UK2, Be1 489 C a/␣
18 J932571 1993 UK Oropharynx AIDS patient UK2, Be1 379 B a/␣
18 J932575 1993 UK Oropharynx AIDS patient UK2, Be1 380 C a/␣
18 J941383 1994 UK Oropharynx AIDS patient UK2, Be1 378 A a/␣
19 J942148 1994 Germany Oropharynx AIDS patient Be1 381 A ␣/␣
19 J942149 1994 Germany Oropharynx AIDS patient Be1 382 A ␣/␣
20 TW2-76 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW2-80 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW2-81 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW2-82 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW2-83 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW2-84 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW2-85 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW2-86 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW3-55 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW8-43 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW8-44 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW8-45 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW8-46 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW8-47 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
20 TW12-79 1994 USA Oropharynx AIDS patient 254 A a/␣ 95
20 TW12-99 1994 USA Oropharynx AIDS patient 79 A a/␣ 95
21 S01 1995 Italy Oropharynx Liver transplant 88 C a/␣ 87
21 S02 1995 Italy Oropharynx Liver transplant 264 C a/␣ 87
21 S03 1995 Italy Oropharynx Liver transplant 88 C a/␣ 87
21 S04 1995 Italy Oropharynx Liver transplant 264 C a/␣ 87
21 S05 1995 Italy Sputum Liver transplant 265 C ␣/␣ 87
21 S06 1995 Italy Sputum Liver transplant 88 C a/␣ 87
21 S07 1995 Italy Sputum Liver transplant 88 C a/␣ 87
21 S08 1995 Italy Sputum Liver transplant 266 C a/␣ 87
21 S09 1995 Italy Sputum Liver transplant 265 C ␣/␣ 87
21 S10 1995 Italy Bronchoalveolar lavage Liver transplant 88 C a/␣ 87
22 T63 1995 Canada Oropharynx AIDS patient 222 B a/␣ 15
22 T65 1995 Canada Oropharynx AIDS patient 222 B a/␣ 15
22 T68 1995 Canada Oropharynx AIDS patient 223 C a/␣ 15
23 T118 1995 Canada Oropharynx AIDS patient 151 A a/␣ 15
23 D08-330 T118 population exposed
to fluconazole
278 A a/␣ 16
23 D09-330 T118 population exposed
to fluconazole
278 A a/␣ 16
23 D11-330 T118 population exposed
to fluconazole
278 A a/␣ 16
23 D12-165 T118 population exposed
to fluconazole
278 A a/␣ 16
23 D12-260 T118 population exposed
to fluconazole
280 A a/␣ 16
23 D12-330 T118 population exposed
to fluconazole
279 A a/␣ 16
24 J990102 1999 Belgium Vagina Symptomatic vaginitis Be1 45 B a/␣
24 J990103 1999 Belgium Cervix Symptomatic vaginitis Be1 490 B ␣/␣
25 AM2001/0019 2001 UK Sternal abscess 527 A a/␣
25 AM2001/0022 2001 UK Sternal abscess 527 A a/␣
26 WC01-202761 2001 Australia Blood 390 B a/␣
26 WC01-202762 2001 Australia Wound 391 B a/␣
27 WC02-202964 2002 Australia Blood 420 A a/␣
27 WC02-202965 2002 Australia Urine 421 A a/␣
28 WC02-202291 2002 Australia Blood 232 A a/␣
28 WC02-202292 2002 Australia Blood 232 A a/␣
28 WC02-202293 2002 Australia Blood 430 A a/␣
29 WC02-202294 2002 Australia Blood 416 B a/␣
29 WC02-202295 2002 Australia Blood 417 B a/␣
29 WC02-202296 2002 Australia Blood 418 B a/␣
30 WC02-201987 2002 Australia Urine 414 A a/␣
30 WC02-201988 2002 Australia Catheter tip 415 A a/␣
30 WC02-202225 2002 Australia Blood 66 A a/␣
Continued on following page
on May 16, 2020 by guest
http://jcm.asm.org/
the set were rescrutinized and/or the isolates were resequenced to ensure that the differences were reproducible.
Animal experiments.Single colonies of isolates S09 and WC02-202294 (Table 1) were grown for 20 h at 30°C in NGY medium (0.1% neopeptone [Difco, Sparks, MD], 0.4% glucose, and 0.1% yeast extract [Difco]) (40). These isolates were chosen to represent low (S09) and high (WC02-202294) levels of heterozy-gosity in the MLST data. The yeast cells were washed twice in sterile saline and
[image:5.585.51.539.80.601.2]resuspended in saline, and concentrations were adjusted by hemocytometer count results. For each isolate, a single female BALB/c mouse weighing 20⫾2 g was injected intravenously at a dose of 2⫻105yeast cells/g body weight, confirmed by subsequent viable counts. Three days after challenge, the mice were humanely terminated, the kidneys and brains were removed and homogenized in saline, and 10-fold dilutions were plated onto Sabouraud agar (Oxoid, Basing-stoke, United Kingdom) for incubation at 35°C. From each organ culture, six TABLE 1—Continued
Isolate
set Isolate
Yr isolated
Country
of origina Isolated fromb: Patient detail(s)c Storage historyd DST
ABC type
MTL
type Reference
31 WC02-202861 2002 Australia Ileal conduit 419 A a/␣
31 WC02-202862 2002 Australia Blood 419 A a/␣
32 WC03-200970b 2003 Australia Catheter tip 422 B a/␣
32 WC03-200970d 2003 Australia Blood 423 B a/␣
32 WC03-200970e 2003 Australia Blood 424 B a/␣
32 WC03-200970h 2003 Australia Thigh 425 B a/␣
33 WC03-202178 2003 Australia Urine 427 A a/␣
33 WC03-202179 2003 Australia Blood 427 A a/␣
33 WC03-202180 2003 Australia Blood 427 A a/␣
34 WC03-200975 2003 Australia Blood 426 B a/␣
34 WC03-201397 2003 Australia Urine 426 B a/␣
35 WC03-203422 2003 Australia Blood 37 A a/␣
35 WC03-203424 2003 Australia Urine 428 A a/␣
36 AM2003/0044 2003 UK Vagina 172 C a/␣
36 AM2003/0045 2003 UK Vagina 172 C a/␣
37 AM2003/0108 2003 UK Blood 212 A a/␣
37 AM2003/0109 2003 UK Blood 212 A a/␣
38 AM2005/0131 2005 UK Blood 522 A a/␣
38 AM2005/0134 2005 UK Blood 522 A a/␣
38 AM2005/0135 2005 UK Blood 522 A a/␣
39 AM2005/0163 2005 UK Sputum 537 A a/␣
39 AM2005/0165 2005 UK Blood 537 A a/␣
40 AM2005/0150 2005 UK Blood 536 A a/␣
40 AM2005/0151 2005 UK Catheter tip 536 A a/␣
41 AM2005/0211 2005 UK Blood 564 A a/␣
41 AM2005/0212 2005 UK Blood 564 A a/␣
41 AM2005/0213 2005 UK Blood 565 A a/␣
41 AM2005/0218 2005 UK Blood 564 A a/␣
42 AM2005/0192 2005 UK Blood 66 A a/␣
42 AM2005/0195 2005 UK Blood 66 A a/␣
43 IHEM17156 Belgium Guinea pig penis Be1, Be2 307 B a/␣
43 IHEM17157 Belgium Guinea pig nose Be1, Be2 305 B a/␣
44 3153Abstl Recently received as
3153, Bristol, UK
369 A a/␣
44 3153Alds Recently received as
3153, Leeds, UK
369 A a/␣
44 3153ALndn2 Recently received as
3153, London, UK
286 A a/␣
44 3153ALndn1 1980s culture of 3153
from London laboratory
UK2, Be1 286 A a/␣
44 90/189 Authentic 3153 UK2, Be1 54 A a/␣ 41
44 90/195 3153, CA05 in previous
study
UK2, Be1 261 A a/␣ 41
44 90/198 3153, CA08 in previous
study
UK2, Be1 262 A a/␣ 41
44 90/202 3153, CA12 in previous
study
UK2, Be1 260 B a/␣ 41
44 90/206 3153, CA16 in previous
study
UK2, Be1 263 B a/␣ 41
44 IHEM20523 3153, CA01 in previous
study
UK2, Be1, Be2 370 A a/␣ 41
44 IHEM20535 3153, CA14 in previous
study
UK2, Be1, Be2 369 A a/␣ 41
aUK, United Kingdom; USA, United States. bCSF, cerebral spinal fluid.
cSTD, sexually transmitted disease.
dUK1, Department of Microbiology, University of Leeds, United Kingdom; UK2, Department of Microbiology, University of Leicester, United Kingdom; Be1,
Janssen Research Foundation, Belgium; BE2, Scientific Institute of Public Health, Brussels, Belgium.
on May 16, 2020 by guest
http://jcm.asm.org/
well-separated colonies were randomly selected, subcultured, and subjected to MLST.
RESULTS
Reproducibility of MLST data.Data were obtained from 17 isolates fully sequenced in duplicate. Sequence reproducibility was perfect for 10 duplicates. For the other seven isolates, differences were at the level of a homozygous versus a het-erozygous result for a single nucleotide polymorphism (SNP) in five instances, two homozygous/heterozygous discrepancies at a single locus in one instance, and a base-for-base (homozy-gous) SNP difference in one instance. All but one of the errors therefore resulted from a misinterpretation of double peaks, an occasional difficulty to be expected with sequencing chro-matograms from a diploid organism.
The eight replicate differences occurred in a total of 2,329 bases sequenced—an accuracy of 99.66%. Since a heterozy-gous/homozygous difference would be scored at half the value of a homozygous difference in the calculation of distances for UPGMA analyses, the sequencing accuracy could be expressed as nine error scores among 4,658 polymorphic sites sequenced (99.8% accuracy). In the context of this study, the worst inac-curacy seen, one homozygous change or two homozygous/het-erozygous changes, would erroneously suggest nonidentity be-tween strains with a score difference of 1 across all SNPs, which corresponds with ap-distance difference of 0.005 by UPGMA nucleotide distance analysis. We have used as a worst-case value that is four times this magnitude, i.e., ap-distance dif-ference of 0.02, as a basis for describing isolates as “indistin-guishable,” although the sequence chromatograms for all iso-lates included in this study have been rescrutinized or the sequencing has been repeated to ensure that differences be-tween isolates are genuine.
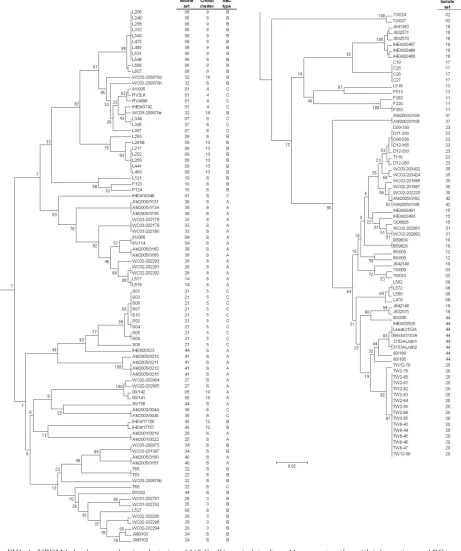
Similarities and differences between multiple isolates from single sources.The DSTs, ABC types, and MTL types of all the isolates studied are shown in Table 1. Figure 1 shows the relationships between the isolates in the form of a UPGMA dendrogram based on MLST sequence data together with their isolate set numbers, ABC types, and eBURST clonal cluster assignments. In the majority of instances, isolates from the same source coclustered, often with the same DST or with very high levels of similarity by MLST (Fig. 1). Isolates in 36 of the 44 sets (121 of 165 isolates) were considered indistinguishable within each set because they clustered in the MLST dendro-gram within our chosen limitingpdistance of 0.02, they had the same ABC type, and they fell within the same eBURST clonal cluster (when a set of isolates showed sufficient sequence vari-ation to generate a clonal cluster). This list includes 11 isolate sets (sets 12, 14, 18, 27, 28, 31, 32, 34, 35, 36, and 40) in which samples came from the bloodstream and a nonsterile site in the same patient. It also includes sets 7 and 8, which came from a patient sampled on two separate admissions to the same hos-pital, 2 months apart. The set from each admission comprised isolates clustering at the level of indistinguishability, but the two sets were widely separated in the UPGMA dendrogram (Fig. 1), unequivocally indicating strain replacement at all sites sampled for this patient.
For the sets of isolates that could not be designated indis-tinguishable by the above-described criteria, the interisolate
differences were of various types. Isolates from set 32 clustered closely, but one isolate differed from the other two in more than one genotype, so it did not conform to a clonal cluster (Fig. 1). The two isolates in sets 13 and 19 differed, with ap
distance of⬎0.02, but were matched in ABC type (Fig. 1). In both cases, we interpret these data to be consistent with mi-crovariation occurring in the isolate sets, which raises the num-ber of sets of isolates that were indistinguishable or very closely related to 40/44 (125/165 isolates).
IHEM16346 did not cocluster with other isolates in set 1 (Fig. 1), but, like the rest of the set, it belonged to the least common ABC type, type C. The data therefore neither confirm nor rule out the possibility that IHEM16346 was the progeni-tor of the other isolates in set 1. Similarly, L527 did not co-cluster with the other 11 isolates in set 6 (Fig. 1), but its ABC type, type B, matched that of the other isolates. Once again, these data do not unequivocally rule in or out an association between L527 and the other isolates in set 6; our records show that, chronologically, L527 was the last isolate received from surveillance cultures of the patient concerned, and the sample was taken 5 days after the previous isolate, so strain alteration or strain replacement is a possible interpretation.
Set 18 presented the most unusual set of findings between isolates. The seven isolates in this set originated from samples taken at different times from the same HIV-infected patient but had histories of storage in different culture collections (Table 1); no detailed information on the isolates is available. Six of the isolates clustered very tightly on the basis of MLST data and formed a single eBURST clonal cluster (Fig. 1), but J932575 was substantially different from these six isolates. Six of the isolates, including J932575, were ABC type B, while J941383 was type A. ABC typing was repeated with these isolates to confirm the different assignments. A similar finding applied to the three isolates T63, T65, and T68, all of which came from the same patient (set 22). T63 and T65, with iden-tical DSTs (Table 1), were type B, but T68, which did not cocluster with the two other isolates, was type C.
Set 44 consisted of 11 isolates that had been supplied as “strain 3153” from various laboratories. Three isolates were recently submitted, and the remaining isolates were part of a study published in 1991 (41). Eight of the isolates (including the three recently obtained isolates) coclustered in the den-drogram (Fig. 1). Five of these isolates belonged to eBURST clonal cluster 1. Isolate 90/206, the most distant in the dendro-gram cluster, was type B, while the other isolates were type A. Of the remaining three isolates in set 44, each of which ap-peared in isolation in the dendrogram (Fig. 1), two were type A and one was type B.
Twelve of the 165 isolates were homozygous at the MTL: nine␣/␣and threea/aisolates (Table 1). The random occur-rence of MTL homozygosity among heterozygous isolates in sets 9, 10, 12, 18, 21, and 24 indicates that this property is not a character of use in strain typing but is a possible indicator of microadaptive change. Both oral isolates in set 3, from a healthy female volunteer in the 1970s, and in set 19, from an AIDS patient in the 1990s, were MTL homozygous; therefore, the possibility that further isolates from these individuals may also have been homozygous at the MTL cannot be excluded.
Loss of heterozygosity associated with strain microvaria-tion.Among the sets of multiple single-source isolates where
on May 16, 2020 by guest
http://jcm.asm.org/
MLST sequence diversity was most evident, differences were seen mainly as heterozygous/homozygous changes between iso-lates rather than as homozygous base exchanges. In set 32, isolate WC03-200970b, which differed most markedly from its three coisolates, had 15 heterozygous SNPs, compared with 4, 9, and 10 for isolates WC03-200970d, WC03-200970e, and
[image:7.585.44.505.72.623.2]WC03-200970h, respectively. In the pair of isolates in set 13, L516 had 20 heterozygous SNPs, compared with 25 in L513. Similarly, in set 19 J942148 had 4 heterozygous SNPs and J942149 had 10 heterozygous SNPs. For isolate set 18, IHEM16346, which may or may not have been the progenitor isolate, had 18 heterozygous SNPs, which was more than those FIG. 1. UPGMA dendrogram showing clustering of 165C. albicansisolates from 44 sources together with information on ABC type and clonal cluster. The dendrogram has been split to facilitate text legibility, with the top half at the left and the continuation at the right.
on May 16, 2020 by guest
http://jcm.asm.org/
for IHEM3742 (11 SNPs) or RV4688 (nine SNPs). Similarly, in set 22, isolate T68 had 33 heterozygous SNPs, which was many more than the 16 heterozygous SNPs in its two sister isolates, T63 and T65.
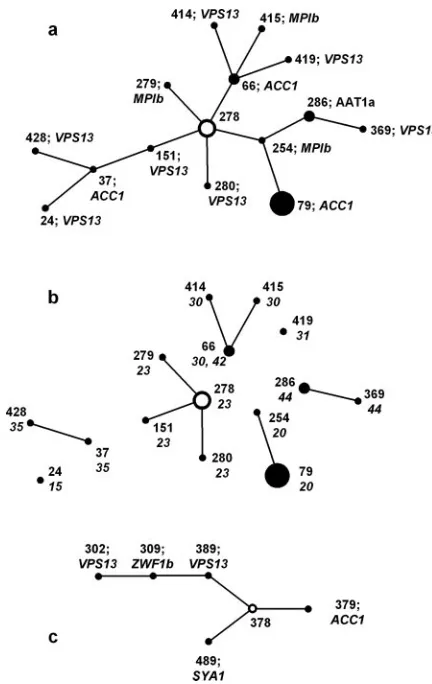
eBURST analysis of MLST data generates “clonal clusters” of isolates in which each differs from another by a single ge-notype among the seven sequenced. For each clonal cluster, the eBURST software determines the founder DST as the one with the greatest number of other DSTs that can be linked in steps by single locus changes (single locus variants). In the present study, most eBURST clonal clusters comprised only two isolates, but clusters 1 and 2 (Fig. 2) contained several isolates. The isolates in clonal cluster 2 (Fig. 2c) all came from a single patient. The isolates in cluster 1 (Fig. 2a) represented eight different isolate sets. The implication of clonal cluster data is of evolutionary patterns that normally show paths of isolate derivation from putative parental isolates. However, the pathways inferred from clonal cluster 1, if correct, indicate both gain and loss of heterozygosity as evolutionary
mecha-nisms forC. albicans. For example, DST 254 in Fig. 2a differs from the putative founder DST 278 by the loss of all heterozy-gous SNPs at theMPIblocus. The step to DST 79 from DST 254, however, involves a gain of two heterozygous SNPs from theACC1locus that was fully homozygous in DST 254. Simi-larly, the path from DST 278 to DST 415 via DST 66 involves the gain of heterozygosity inMPIbin the final step, and the path from DST 278 to DST 37 via DST 151 requires a gain of heterozygosity in DST 37. An alternative explanation for the changes is shown in Fig. 2b, in which connecting lines between isolates from different sets have been removed so that isolates coming from a single patient or other source remain con-nected. The links within each set now show how the isolates could all have evolved within each patient by LOH from a different putative parent, without the clonal relationship im-plied by the eBURST analysis. The validity of disconnecting clonal cluster 1 in this way is emphasized by the sometimes considerable geographical distance between the isolate sources: sets 20 and 23 came from North America; sets 30, 31, and 35 came from came Australia; and sets 15 and 42 came from the United Kingdom. (Isolates in set 44 were laboratory strains.)
[image:8.585.52.268.66.408.2]Table 2 summarizes the nature of the sequence differences between all the sets of isolates that gave nonidentical sequenc-ing results for one or two of the test loci. The table shows that homozygous SNP changes (indicated with the letter C in the table) accounted for only four of the 38 differences found; the remaining changes were all changes from heterozygosity to homozygosity at a polymorphic base position. The loss of a single heterozygous base position, indicated with the letter A in Table 2, included those instances with only a single heterozy-gous base position in the entire genotype. When two or more heterozygosities were found in the sequence for one isolate
TABLE 2. Details of differences in sequences between isolates from the same source
Set Clonal cluster
Nature of differenceabetween isolates in gene fragment
AAT1a ACC1 ADP1 MPIb SYA1 VPS13 ZWF1b Note
1 4 A B
2 15 C
3 6 B
5 10 A
6 9 A
7 A B
8 7 C A
9 13 B
11 14 B
12 B A
18 2 A A A A See Fig. 2
20 1 B See Fig. 2
21 5 C C
23 1 A A See Fig. 2
24 A A
26 3 B
27 A A
28 8 B
29 3 B A
30 1 B A See Fig. 2
32 16 B
35 1 A See Fig. 2
41 A B
43 12 A
44 See Fig. 2
aA, heterozygous/homozygous difference at one SNP; B, difference at all
heterozygous SNPs; C, single homozygous SNP change. FIG. 2. Clonal clusters 1 (a) and 2 (c) generated by eBURST
anal-ysis of the MLST data. For each cluster, the putative founding isolate is shown as a hollow circle. Diameters of the circles are proportionate to the numbers of isolates with the DST indicated. With each DST, the gene locus and SNP difference from the putative predecessor isolate along the path from the founder isolate are specified. The lengths of the joining lines are arbitrary. b shows clonal cluster 1 with the joining lines removed to leave different isolate sets separate; set numbers are shown next to DSTs. Only sets 30 and 42 share a common DST.
on May 16, 2020 by guest
http://jcm.asm.org/
[image:8.585.300.541.468.709.2]and all were homozygous in the other, this was regarded as complete LOH for the locus concerned and is indicated with the letter B in Table 2. Thus, the 34 instances of putative LOH involved 21 instances in which single heterozygosities changed and 13 instances of complete LOH for a sequenced locus.
MLST differences among isolates recovered from experi-mentally infected animals.Because the MLST result for iso-late RV2LK from set 01, recovered from the left kidney of a mouse infected with RV4688 (set 01), showed a minor differ-ence (two more heterozygous SNPs in the recovered isolate than in the originally typed isolate), we conducted a prelimi-nary investigation into possible MLST variations in isolates recovered from infected tissues. Isolates S09 and WC02-202294 were first spread to single colonies on Sabouraud agar, and six clones on each plate were subjected to MLST. All six clones of S09 were indistinguishable by MLST, whereas three DSTs were found among the six clones of WC02-202294. These resulted from a heterozygous/homozygous difference at one SNP inACC1 in two instances and a similar single-site difference inADP1in one instance. Cloned inocula of the two strains were each injected intravenously into two mice. After 3 days, the mice were sacrificed, and colonies ofC. albicanswere reisolated from homogenates of kidney and brain. In repeats of MLST for five or six colonies from each organ, no differences in the DST of each strain were found.
DISCUSSION
Our study used MLST, the most dependable system forC.
albicansstrain differentiation, with a discriminatory power of
0.9996 (84), to investigate strain relationships in multiple iso-lates from single sources. None of the isoiso-lates included in this study happened to represent the most commonly encountered DST, type 69 (84), or a close relative. MLST has confirmed that sets of such isolates most often belong to a single or highly related strain type; 36 (82%) of 44 strain sets were very closely related by MLST data and had identical ABC types. MLST thus agrees with many other strain typing systems that have similarly shown the persistence of a single strain type per patient to be the most common situation. All 11 sets of isolates that included cultures from blood and nonsterile sites in the same patient fell within the 36 most closely related sets. These results clearly indicate that adult patients (none of the isolates tested came from pediatric sources) with bloodstream infec-tions are usually infected with their own commensal isolates. This observation does not, of course, show whether the portal of endogenous infection was the patient’s digestive tract or another route, such as an intravenous catheter. Results similar to ours were obtained in a recently published study from Taiwan in which both MLST and electrophoretic karyotypes of multiple isolates from nine patients in inten-sive care units showed high relatedness but were not always indistinguishable (13). However, multiple oral isolates taken from three HIV-positive patients over several years showed evidence of strain replacement. Our own findings are sup-ported in the case of isolates from set 21, which were pre-viously found by Ca3 DNA fingerprinting to be highly sim-ilar but not indistinguishable (87).
All but 2 of the 44 sets of isolates that we tested came from human sources. Among these were two sets of surveillance
culture isolates (sets 7 and 8) from a patient who was admitted twice to the same hospital in March and May of 1985. While each of these individual sets of isolates clustered within this study’s bounds of indistinguishability, the two sets were suffi-ciently different, including a change of ABC type, to provide unequivocal evidence of strain replacement between the pa-tient’s hospital admissions. No other similar, unambiguous ex-amples of strain replacement were found. It is possible that isolate L527 (set 1) might also represent strain replacement, but the level of dissimilarity of this isolate from the rest of the set may have resulted from mishandling errors in the course of many years of maintenance. A third possibility is that the patient carried two strain types and that the less populous one was the randomly picked colony from the isolation plate only for the L527 sample.
One finding from this study concerns the storage and trans-mission of isolates between collections. The earliest isolates of certain date in our 44 sets were a pair of successive vaginal isolates first cultured in Leeds, United Kingdom, in 1973. Over 30 years of storage, the with removal and occasional reculture from the first author’s collection in Leicester, United King-dom, Beerse, Belgium, and now Aberdeen, United KingKing-dom, these isolates differed by only a single polymorphism: a change from A to G in theADP1sequence. Twelve other isolate sets dating from the late 1970s to the late 1980s had also been transferred between several locations and in some instances had been returned after storage in other collections. All of these isolate sets coclustered to high levels of similarity by MLST and ABC type, suggesting that their storage and main-tenance had been adequate for long periods of time. By con-trast, several of the isolates that had originally been received into the Leicester, United Kingdom, collection from other laboratories under the label 3153 (set 44 in the present study) were obviously subjected to handling errors at some stage in their history. In the original interlaboratory studies of these isolates (32, 41), restriction fragment length polymorphism and multilocus enzyme electrophoresis data already suggested that several of them differed from the pattern of the majority of the isolates. The MLST results now show that at least three of the isolates cannot be regarded as authentic examples of strain NCPF3153. The data for the most recently received examples of NCPF3153 define the strain as DST 369 and ABC type A. Isolate 90/189, which we received as the “authentic” strain 3153 in the 1980s, differed from this DST in two genotypes but was still type A and remained closely clustered with six other examples of “strain 3153” by MLST (Fig. 1).
The differences between the coclustered NCPF3153 isolates in Fig. 1 are of the same order of magnitude as those we interpret as microvariation in other isolate sets. The nature of these changes most often involves the loss of one or all het-erozygous SNPs from one or two of the sequenced genes (Ta-ble 2). LOH as a general mechanism for microvariation changes has been previously described (16, 21, 70, 85, 88). However, the precise mechanism underlying LOH events is unknown. The list of possible mechanisms includes recombi-nation and gene conversion. The possibility of chromosome ploidy changes must now be added to this list, since aneu-ploidy, including segmental aneuploidy of subchromosomal re-gions, has now been elegantly demonstrated inC. albicans(77). The loss of all or part of a chromosome followed by
on May 16, 2020 by guest
http://jcm.asm.org/
cation has been invoked as a mechanism underlying a range of phenotypic properties in the fungus, including sorbose utiliza-tion and fluoroorotic acid resistance (26, 94) and the genera-tion of MTL-homozygous variants (98). If spontaneous partial ploidy changes occur as frequently in clinical isolates as was shown for the laboratory strain CAI-4 (77), the high frequency of heterozygosity changes seen among the isolates that we sequenced may not be surprising. This comment is further supported by the occasional appearance of isolates homozy-gous at the MTL in a temporal series of isolates that were otherwise MTL heterozygous. Wu et al. previously studied distantly located heterozygous markers on chromosome 5 on both sides of the MTL and found that whole-chromosome loss and replacement accounted for spontaneous conversion to MTL homozygosity in 15/16 isolates studied, with a recombi-national event in only one isolate (98). We speculate that a complete or partial chromosome loss and reduplication, com-bined with occasional genetic exchange through recombination and inefficient DNA repair, account for the apparent genetic plasticity of C. albicansevidenced in our data, including ap-parent diploid homozygous base switches, which would be in-explicable by chance mutation. Future investigations of karyo-types and of sequences neighboring and distant from the MLST genes on the same chromosome will provide evidence for recombination, gene conversion, or aneuploidy in strain sets showing LOH and single-base homozygous changes.
Of particular note in our study are the few examples among our isolates of strains that were very closely related by MLST but that differed in ABC type (notably in sets 18 and 44). ABC type, based on the presence of one or bothITS1sequences in rRNA genes, is usually a very stable epidemiological marker in
C. albicansisolates and is sufficient information to demonstrate
geographical and temporal differences amongC. albicans iso-lates (46). It has been suggested that type C, the least common ABC type, may represent an intermediate form that is gaining or losing the type A or BITS1sequence (25). On this basis, and in view of our finding of two examples where types A and B were represented among types similar by MLST, we regard ABC typing as a very helpful confirmatory test in instances where isolates differ by more than one or two SNPs in MLST, but it should not be regarded as a definitive test for isolate relatedness in all instances. The rRNA genes inC. albicansare located on chromosome R; among the MLST genes, only
ACC1is located on this chromosome.
eBURST analysis of MLST data provides a useful basis for tracing putative evolutionary patterns among microbial iso-lates. However, in the present study, this analytical approach may have suggested invalid relationships between strains. The major clonal cluster found among our isolates (Fig. 2) suggests that there is a set of evolutionary paths between isolates that requires successive gains and losses of heterozygosity at one of the sequenced loci to be sustainable. Clonal cluster 1 is there-fore unlikely to represent a true clonal relationship between these isolates of widely disparate geographical origins. One obvious mechanism for the gain of heterozygosity would be mating, but it remains uncertain to what extent mating be-tweenC. albicansisolates occurs naturally (5). If mating is the basis for the regaining of allelic heterozygosity, then the mating process would need to involve strains with dissimilar alleles; in other words, theC. albicanscells colonizing or infecting a site
within a patient would need to include homozygous strains of the opposite MTL type and with different MLST DSTs. This situation appears to be quite different from the high tendency toward clonal reproduction evidenced by most isolate sets in this study as well as those described previously in other publi-cations (66, 85). The disadvantage of the eBURST approach and other statistical methods that examine only genotype and DST assignations is that they do not distinguish whether iso-lates differ in sequence at a locus by a single SNP or by multiple SNPs. Hence, its value becomes limited with an organism such
asC. albicans, which regularly undergoes “mutations” at
mul-tiple nucleotides within a single locus.
One hypothesis that would accommodate the otherwise con-flicting evidence suggesting both clonal strain maintenance and temporal gain and loss of heterozygosities and even ABC types would be thatC. albicanspopulations commonly exist in a state of high genetic plasticity. The phenomenon of hypermutable (“mutator”) strains is well recognized among some species of bacteria, particularly gram-negative bacilli (18, 59, 82). Hyper-mutability is associated with the appearance of multiple colony forms and other phenotypic variations, and it is possible that similarly variable strains exist among pathogenicCandida spe-cies. Almost all the isolates that we have studied began as single clones picked from primary isolation plates in clinical laboratories. When successive isolates from the same site in the same patient are found to be sometimes MTL homozygous and sometimes MTL heterozygous (sets 18 and 21), the most reasonable explanation is not that the clones picked randomly from a given isolation plate represent all colonies from that sample but rather that the sample contained a mixture of types. The possibility that C. albicans colonies on isolation plates often differ has been suggested by the minority of colonies that are able to undergo a white-opaque transition (36), by minor intercolony differences in DNA fingerprint patterns (the orig-inal finding characterized as “microevolution”) (35), and by colony-to-colony differences in azole antifungal susceptibility (75). In the present study, three DSTs were found among the six clones of WC02-202294 (from set 29) randomly sampled from a spread plate. If either some strains at all times or all strains in certain environments undergo recombination, gene replacement, or partial ploidy changes at elevated frequencies, the resulting mixture of strain types in the population isolated would explain the occasional superficially aberrant result in the present study. Our pilot experiment to demonstrate this phe-nomenon by resampling infected tissues infected with cloned
C. albicansisolates failed to confirm the hypothesis; however,
we plan to investigate a broader range of isolates in vivo to assess the reality, or otherwise, of strain diversity as a natural process forC. albicans. We are also investigating natural ge-netic diversity between colonies on primary isolation plates.
ACKNOWLEDGMENTS
This study was supported by grants from the Wellcome Trust (69615 and 74898).
We thank individuals who have provided us with isolates, particu-larly James Anderson of the University of Toronto and Nicole Nolard and Francoise Symoens of the Scientific Institute of Public Health in Brussels.
on May 16, 2020 by guest
http://jcm.asm.org/
REFERENCES
1.Asakura, K., S. I. Iwaguchi, M. Homma, T. Sukai, K. Higashide, and K. Tanaka. 1991. Electrophoretic karyotypes of clinically isolated yeasts of
Candida albicansandC. glabrata. J. Gen. Microbiol.137:2531–2538. 2.Barchiesi, F., L. F. DiFrancesco, P. Compagnucci, D. Arzeni, O. Cirioni, and
G. Scalise.1997. Genotypic identification of sequentialCandida albicans
isolates from AIDS patients by polymerase chain reaction techniques. Eur. J. Clin. Microbiol. Infect. Dis.16:601–605.
3.Barchiesi, F., R. J. Hollis, M. Del Poeta, D. A. McGough, G. Scalise, M. G. Rinaldi, and M. A. Pfaller.1995. Transmission of fluconazole-resistant Can-dida albicansbetween patients with AIDS and oropharyngeal candidiasis documented by pulsed-field gel electrophoresis. Clin. Infect. Dis.21:561– 564.
4.Bartie, K. L., D. W. Williams, M. J. Wilson, A. J. C. Potts, and M. A. O. Lewis.2001. PCR fingerprinting ofCandida albicansassociated with chronic hyperplastic candidosis and other oral conditions. J. Clin. Microbiol. 39:
4066–4075.
5.Bennett, R. J., and A. D. Johnson.2005. Mating inCandida albicansand the search for a sexual cycle. Ann. Rev. Microbiol.59:233–255.
6.Boerlin, P., F. Boerlin-Petzold, J. Goudet, C. Durussel, J. L. Pagani, J. P. Chave, and J. Bille.1996. TypingCandida albicansoral isolates from human immunodeficiency virus-infected patients by multilocus enzyme electro-phoresis and DNA fingerprinting. J. Clin. Microbiol.34:1235–1248. 7.Bougnoux, M.-E., D. M. Aanensen, S. Morand, M. The´raud, B. G. Spratt,
and C. d’Enfert.2004. Multilocus sequence typing of Candida albicans: strategies, data exchange and applications. Infect. Genet. Evol.4:243–252. 8.Bougnoux, M.-E., S. Morand, and C. d’Enfert.2002. Usefulness of
multilo-cus sequence typing for characterization of clinical isolates ofCandida albi-cans. J. Clin. Microbiol.40:1290–1297.
9.Bougnoux, M.-E., A. Tavanti, C. Bouchier, N. A. R. Gow, A. Magnier, A. D. Davidson, M. C. J. Maiden, C. d’Enfert, and F. C. Odds.2003. Collaborative consensus for optimized multilocus sequence typing ofCandida albicans. J. Clin. Microbiol.41:5265–5266.
10.Burnie, J. P., F. C. Odds, W. Lee, C. Webster, and J. D. Williams.1985. Outbreak of systemicCandida albicansin intensive care unit caused by cross infection. Br. Med. J.290:746–748.
11.Calderone, R. A.2002.Candidaand candidiasis. ASM Press, Washington, D.C.
12.Caugant, D. A., and P. Sandven.1993. Epidemiological analysis ofCandida albicansstrains by multilocus enzyme electrophoresis. J. Clin. Microbiol.
31:215–220.
13.Chen, K.-W., Y.-C. Chen, H.-J. Lo, F. C. Odds, T.-H. Wang, C.-Y. Lin, and S.-Y. Li.2006. Multilocus sequence typing for analyses of clonality of Can-dida albicansstrains in Taiwan. J. Clin. Microbiol.44:2172–2178. 14.Cowen, L. E., L. M. Kohn, and J. B. Anderson.2001. Divergence in fitness
and evolution of drug resistance in experimental populations ofCandida albicans. J. Bacteriol.183:2971–2978.
15.Cowen, L. E., A. Nantel, M. S. Whiteway, D. Y. Thomas, D. C. Tessier, L. M. Kohn, and J. B. Anderson.2002. Population genomics of drug resistance in
Candida albicans. Proc. Natl. Acad. Sci. USA99:9284–9289.
16.Cowen, L. E., D. Sanglard, D. Calabrese, C. Sirjusingh, J. B. Anderson, and L. M. Kohn.2000. Evolution of drug resistance in experimental populations ofCandida albicans. J. Bacteriol.182:1515–1522.
17.Cross, L. J., D. W. Williams, C. P. Sweeney, M. S. Jackson, M. A. O. Lewis, and J. Bagg.2004. Evaluation of the recurrence of denture stomatitis and
Candidacolonization in a small group of patients who received itraconazole. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodont.97:351–358. 18.Denamur, E., S. Bonacorsi, A. Giraud, P. Duriez, F. Hilali, C. Amorin, E.
Bingen, A. Andremont, B. Picard, F. Taddei, and I. Matic.2002. High frequency of mutator strains among human uropathogenicEscherichia coli
isolates. J. Bacteriol.184:605–609.
19.Dromer, F., L. Improvisi, B. Dupont, M. Eliaszewicz, G. Pialoux, S. Fournier, and V. Feuillie.1997. Oral transmission ofCandida albicans be-tween partners in HIV-infected couples could contribute to dissemination of fluconazole-resistant isolates. AIDS11:1095–1101.
20.Feil, E. J., B. C. Li, D. M. Aanensen, W. P. Hanage, and B. G. Spratt.2004. eBURST: inferring patterns of evolutionary descent among clusters of re-lated bacterial genotypes from multilocus sequence typing data. J. Bacteriol.
186:1518–1530.
21.Forche, A., G. May, and P. T. Magee.2005. Demonstration of loss of het-erozygosity by single-nucleotide polymorphism microarray analysis and al-terations in strain morphology inCandida albicansstrains during infection. Eukaryot. Cell4:156–165.
22.Hull, C. M., and J. Heitman.2002. Fungal mating:Candida albicansflips a switch to get in the mood. Curr. Biol.12:R782–R784.
23.Hull, C. M., R. M. Raisner, and A. D. Johnson.2000. Evidence for mating of the “asexual” yeastCandida albicansin a mammalian host. Science289:307– 310.
24.Johnson, A.2003. The biology of mating inCandida albicans. Nat. Rev. Microbiol.1:106–116.
25.Jorge, J., M. J. McCullough, and S. R. Porter.2001. Longitudinal in vitro
study of the stability ofCandida albicansgenotypes. Clin. Infect. Dis.33:
1195.
26.Kabir, M. A., A. Ahmad, J. R. Greenberg, Y. K. Wang, and E. Rustchenko.
2005. Loss and gain of chromosome 5 controls growth ofCandida albicanson sorbose due to dispersed redundant negative regulators. Proc. Natl. Acad. Sci. USA102:12147–12152.
27.Kam, A. P., and J. P. Xu.2002. Diversity of commensal yeasts within and among healthy hosts. Diagn. Microbiol. Infect. Dis.43:19–28.
28.Kumar, S., K. Tamura, I. B. Jakobsen, and M. Nei.2001. MEGA2: molec-ular evolutionary genetics analysis software. Bioinformatics17:1244–1245. 29.Lachke, S. A., S. R. Lockhart, K. J. Daniels, and D. R. Soll.2003. Skin
facilitatesCandida albicansmating. Infect. Immun.71:4970–4976. 30.Legrand, M., P. Lephart, A. Forche, F. M. C. Mueller, T. Walsh, P. T. Magee,
and B. B. Magee.2004. Homozygosity at the MTL locus in clinical strains of
Candida albicans: karyotypic rearrangements and tetraploid formation. Mol. Microbiol.52:1451–1462.
31.Leguennec, R., J. Reynes, M. Mallie, C. Pujol, F. Janbon, and J. M. Bastide.
1995. Fluconazole- and itraconazole-resistantCandida albicansstrains from AIDS patients: multilocus enzyme electrophoresis analysis and antifungal susceptibilities. J. Clin. Microbiol.33:2732–2737.
32.Lehmann, P. F., L. C. Wu, and D. W. R. Mackenzie.1991. Isoenzyme changes inCandida albicansduring domestication. J. Clin. Microbiol.29:2623–2625. 33.Lischewski, A., D. Harmsen, K. Wilms, G. Baier, U. Gunzer, H. Klinker, M. Wilhelm, A. Schwinn, and J. Hacker.1999. Molecular epidemiology of Can-dida albicansisolates from AIDS and cancer patients using a novel standard-ized CARE-2 DNA fingerprinting technique. Mycoses42:371–383. 34.Lockhart, S. R., K. J. Daniels, R. Zhao, D. Wessels, and D. R. Soll.2003. Cell
biology of mating inCandida albicans. Eukaryot. Cell2:49–61.
35.Lockhart, S. R., J. J. Fritch, A. S. Meier, K. Schroppel, T. Srikantha, R. Galask, and D. R. Soll.1995. Colonizing populations ofCandida albicansare clonal in origin but undergo microevolution through C1 fragment reorgani-zation as demonstrated by DNA fingerprinting and C1 sequencing. J. Clin. Microbiol.33:1501–1509.
36.Lockhart, S. R., C. Pujol, K. J. Daniels, M. G. Miller, A. D. Johnson, M. A. Pfaller, and D. R. Soll.2002. InCandida albicans, white-opaque switchers are homozygous for mating type. Genetics162:737–745.
37.Lockhart, S. R., B. D. Reed, C. L. Pierson, and D. R. Soll.1996. Most frequent scenario for recurrentCandidavaginitis is strain maintenance with “substrain shuffling”: demonstration by sequential DNA fingerprinting with probes Ca3, C1, and CARE2. J. Clin. Microbiol.34:767–777.
38.Lockhart, S. R., W. Wu, J. B. Radke, R. Zhao, and D. R. Soll.2005. Increased virulence and competitive advantage of a/␣over a/a or␣/␣offspring con-serves the mating system ofCandida albicans. Genetics169:1883–1890. 39.Lott, T. J., R. E. Fundyga, R. J. Kuykendall, and J. Arnold.2005. The human
commensal yeast, Candida albicans, has an ancient origin. Fungal Genet. Biol.42:444–451.
40.MacCallum, D. M., and F. C. Odds.2005. Temporal events in the intrave-nous challenge model for experimentalCandida albicansinfections in female mice. Mycoses48:151–161.
41.Mackenzie, D. W. R., and F. C. Odds.1991. Non-identity and authentication of two major reference strains ofCandida albicans. J. Med. Vet. Mycol.
29:255–261.
42.Magee, B. B., and P. T. Magee.2000. Induction of mating inCandida albicans
by construction of MTLa and MTL␣strains. Science289:310–313. 43.Maiden, M. C. J.2006. Multilocus sequence typing of bacteria. Annu. Rev.
Microbiol.60:561–588.
44.Marr, K. A., C. N. Lyons, K. Ha, T. R. Rustad, and T. C. White.2001. Inducible azole resistance associated with a heterogeneous phenotype in
Candida albicans. Antimicrob. Agents Chemother.45:52–59.
45.McCreight, M. C., and D. W. Warnock.1982. Enhanced differentiation of isolates ofCandida albicansusing a modified resistogram method. Mykosen
25:589–598.
46.McCullough, M., K. V. Clemons, and D. A. Stevens.1999. Molecular epide-miology of the global and temporal diversity of Candida albicans. Clin. Infect. Dis.29:1220–1225.
47.McCullough, M. J., K. V. Clemons, and D. A. Stevens.1999. Molecular and phenotypic characterization of genotypicCandida albicanssubgroups and comparison withCandida dubliniensisandCandida stellatoidea. J. Clin. Mi-crobiol.37:417–421.
48.McCullough, M. J., B. C. Ross, B. D. Dwyer, and P. C. Reade.1994. Geno-type and phenoGeno-type of oralCandida albicansfrom patients infected with the human immunodeficiency virus. Microbiology140:1195–1202.
49.Mercure, S., S. Poirier, G. Lemay, P. Auger, S. Montplaisir, and L. Dere-pentigny.1993. Application of biotyping and DNA typing ofCandida albi-cansto the epidemiology of recurrent vulvovaginal candidiasis. J. Infect. Dis.
168:502–507.
50.Miller, M. G., and A. D. Johnson.2002. White-opaque switching inCandida albicansis controlled by mating-type locus homeodomain proteins and al-lows efficient mating. Cell110:293–302.
51.Nebavi, F., S. Arnavielhe, R. Leguennec, E. Menan, A. Kacou, P. Combe, E. Aoussi, M. Mallie, M. Kone, and J. M. Bastide.1998. Oropharyngeal can-didiasis in AIDS patients from Abidjan (Ivory Coast)—antifungal