. exp. Biol. (1977), 66, 6S-8i 6 5 ^ih 9 figures
Printed in Great Britain
THE MAUTHNER-INITIATED STARTLE RESPONSE
IN TELEOST FISH
BY ROBERT C. EATON, ROCCO A. BOMBARDIERI AND DIETRICH L. MEYER
Neitrobiology Unit, Scripps Institution of Oceanography and Department of Neuro-sciences, University of California, San Diego, Lajolla, CA. 92093, U.S.A.
Department of Psychiatry, University of Goettingen, Goettingen, West Germany
(Received 29 June 1976)
SUMMARY
1. A characteristic behaviour, the 'Mauthner-initiated startle response', was recorded and quantitatively analysed with high-speed cinematography (200 frames/sec) after vibrational stimulation in 11 of 13 teleost species which possess Mauthner cells.
2. The latency of the response is 5-10 msec. This behaviour has: (a) an initial phase, the 'fast-body-bend', lasting about 20 msec and consisting of a stereotyped displacement of the head and tail to one side and (i), a second phase, the 'return-flip', consisting of a non-stereotyped flip of the tail to the opposite side.
3. Within 100 msec after the start of the Mauthner-initiated startle response, most fish were displaced o ^ - i ^ body lengths from their initial position. The variability of the animal's location after 100 msec suggests that the behaviour is adaptively non-predictable.
4. In goldfish, the Mauthner-initiated startle response could also be elicited by visual stimulation.
5. We conclude that the fast-body-bend is the direct result of activation of one Mauthner cell and its spinal motor neurone pool.
6. In four species we described examples of apparently non-Mauthner initiated startle responses.
INTRODUCTION
Other cinematographic studies (Hertel, 1966; Webb, 1975) have been done to develop mechanical models of rapid acceleration in trout (Salmo gairdneri). How-ever, these provide only limited insight into the underlying neurophysiological mechanisms because the filming rate of 1 frame/15-6 msec was not sufficient to ade-quately document a response mediated by the M-cells. That is, in larval zebrafish, the M-cell can fire within 6 msec from the start of a vibrational stimulus and the mechanical muscular response begins 2 msec later (Eaton & Farley, 1975). In adult goldfish, the Mauthner spike (M-spike) has a minimal latency of 2 msec following the acoustic stimulus, and the electrical response in the muscle begins 2 msec later (Zottoli, 1977). In addition, the use of an electrical stimulus in the studies on trout further complicates the physiological interpretation.
In the present study we used a fast frame rate (1 frame/5 msec) and replicate trials to quantitatively analyse the startle behaviour of goldfish and zebrafish to acoustical-vibrational stimulation. Our purpose was to develop insights into the neurophysio-logical basis and functional importance of the startle response. This analysis revealed the presence of a characteristic response pattern, probably mediated by the M-cell, and prompted a further analysis of examples of startle behaviour in 11 other teleost species to determine whether a characteristic behaviour exists among diverse fish with M-cells. Species studied are from seven orders, including both primitive and advanced groups (Gosline, 1971), a variety of body forms (Breder, 1926), and diverse habitats.
MATERIALS AND METHODS
Experimental animals were obtained from laboratory stocks and were chosen to maximize phylogenetic, ecological and body-form diversity (Table 1). Previous studies established the presence of the M-cells in five species of this study (Table 1), and we used histological techniques (see below) to identify the M-cells in the remaining eight species. Both freshwater and marine fish are included. Several individual zebra-fish, hatchetzebra-fish, catzebra-fish, garfish and calico rockfish were studied whereas only one individual was used in replicate trials for other species. Tropical specimens were identified according to the descriptions of Sterba (1962).
Order Table I. Taxonomic relations of experimental species and source for Mauthner cell identijication Family Species Common name Lower teleost fish
Salmoniformes Cypriniformes
Intermediate teleost fish Beloniformes Belonidae Xenetodon cancila (H.-B.) Garfish Higher teleost fish Scorpaeniformes Scorpaenidae Sebasies dalli (E. & B.) Calico rockfish Mastacembeliformes Mastacembelidae Mastacembelus loennbergi Boulinger Spiny eel Perciformes Serranidae Paralabrcuc clathratus (Girard) Kelp bass Pleuronectiformes Bothidae Paralichthys californicus (Ayres) California halibut Salmonidae Salmo gairdneri (Gibbons) Cyprinidae Carassius auratus (L.) Brachydanio rerio (H.-B.) Gasteropelecidae Carnegiella strigata Giinther Gasteropelecus sternicla (L.) Ictaluridae Ictalurus nebulosus (Leseur) Characidae Hydrocinos maculatus (C. & V.) Apteronotidae Apteronotus albifrons (L.) Rainbow trout
Goldfish Zebrafish Marbled
hatchetfish
Common
hatchetfish
Catfish Characid Black
the photographic field. The subject was visually isolated from the stimulus and signal light. Control experiments, without stimulus presentation, verified that the subject did not respond to the falling of the magnet or the turning on of the camera. The signal light also provided a calibration for the film speed.
Photography was done with a Milliken DMB5-2, 16 mm movie camera with 1 msec exposure at 200 frames. sec"1. These characteristics, and the 10 msec signal light, permitted latency measurements with an accuracy of ±4 msec. Four 300 watt flood-lamps provided approximately 1000 footcandles of illumination (incident light measured at water surface with a Gossen Luna-Pro light meter). The floodlamps were turned off between trials to prevent heating the water. In separate experiments, filming was done from above and in front of the aquarium. In general, fish were allowed to swim freely in the tank and were usually filmed in midwater at various orientations.
The responses of goldfish to visual stimulation were studied by dropping a golf ball into the experimental aquarium from 17-5 cm above the water surface. A small magnet was glued to the ball and, as above, we used a timing circuit and electro-magnet to release the ball and to synchronize filming. Only those startle responses which preceded the stimulus striking the water were counted as positive to visual stimulation. Latency measurements were not possible in these experiments because the time when the stimulus was first seen by the fish is unknown.
For single-frame data analysis we used an L-W Photo Inc., 224-A projector and made drawings and measurements from the projected images. To determine displace-ment speed we measured the position of the rostral-most extremity of the head at consecutive 5 msec intervals when the animal was viewed from above. A negligible error is introduced because the pathway of the head follows a very small arc. To measure angular velocities we determined the angle of the midline of the head in dorsal view sequences at consecutive 5 msec intervals. The error in this technique is estimated to be ± 5°.
For histological analysis, animals were anaesthetized with tricaine methanesulphon-ate and transcardially perfused with fish saline followed by 10% formalin. Brains were removed and stored in 10% formalin until they were embedded in paraffin and sectioned at 10 to 15 fim. Sections were stained according to either the Kluver-Barrera method for myelin and nerve cells (Luna, i960), or Ziesmer's (1952) modifica-tion of Bodian's silver stain. Light microscopic examinamodifica-tion of all species revealed the presence of a single pair of giant neurones which met the criteria for identification as M-cells: that is, location at the level of the eighth root, presence of an axon cap, conspicuously large ventral and lateral dendrites, and a decussating axon (see Zottoli,
1976 a).
RESULTS
(a) A biphasic startle response to vibrational stimulation in goldfish and zebrafish
Table 2. Quantitative characteristics of startle responses" in teleostfih (Values shown as mean f S.E. ; parentheses contain n's) ;5* Time to max. Time to max. Max. angular Max. Tempera- Latency angular velocity displacement velocity displacement Length of ture
7
(msec) (msec) velocity (msec) (dedsec) speed (Lls) fish (cm) (C"s.
3
Goldfish 6.5 f 2.4 (10) 10 + o (6) 11.7+2'6 (6) 8833 f 1961 (6) 34'1 f 5.4 (6) 8.5 24 Zebrafish 5 f0 (6) 8.3 f 2.6 (6) 9.2 f 2.0 (6) I 1400 f 1780 (6) 41.9 f I I .6 (6) 2'5 26
Ei
Rainbow trout 9.2 f 3.8 (6) 16.7 f 2'9 (3) 11.7 f 2.9 (3) 5733 f 61 1 (3) 23.6 f 0 (3) 13.0 20&
Catfish 10.0 f 2.9 (9) 7.5 f 3.4 (2) 10.0 f o (2) 6600 f 849 (2) 29.5 f 8.3 (2) 9.2 25 h Calico rockfish 7.0 f 2.6 (10) 6.7 f 2.9 (3) 13.3 f 7.6 (3) 3600f 917 (3) ZI.O+ 82 (3) 4.4 22 Garfish 5.8 f 2.0 (6) 11.3 f 2.5 (4) 12.5 f 5.0 (4) 3900+ 503 (4) 19'7 f 3.2 (4) 12'0$
CL 23 2 Characid 10.0 f 5 (3) 12.5 f 3.5 (2) 12.5 f 3.5 (2) 2700 f 1273 (2) 14.2 f 5.5 (2) 18.4 23 Marbled hatchet- 6.0 f 2.2 (5) 5 (1) 10 (1) 7600 (1) 74.8 (1) 2.8 24 Yfish Common
10.0 (I) 10.0 (I) 10 (1) 7800 (1) 67.0 (1) 2.1
$.
hatchetfish 242
n Black ghost 10.0 (I) 10.0 (I) 10.0 (I) 5800 (1) 19.7 (1) I 6.0 25 Kelp bass 7.5 k 2.9 (4) 10.0 (I) r 0.0 (I) 4200 (1) 23.6 (1) 14'5 19s.
Spiny eel 7.9 f 2.7 (7) 43'3 f33.3 (3) 11'7f2.9 (3) 4000 f I744 (3) 8.53 f 3.9 (3) 25.5 27 22'0 21F
California 10.0 (I) 2 halibut2
With the exception of the rockfish and spiny eel, this table contains values only from the biphasic startle responses.Fast-body-bend Return-flip
IJLLHii
"1 I I < ( ( < < " C
0")
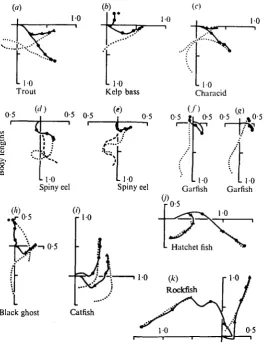
Fig. i. Dorsal view silhouettes offish during startle behaviour: (a) trout, (6) goldfish, (c) zebra-fish, (d) catzebra-fish, («) marbled hatchetzebra-fish, (/) chaxacid, (g) black ghost, (h) garzebra-fish, (<) calico rockfish, (J) spiny eel. Silhouettes are drawn at 5 msec intervals beginning 5 msec before onset of the vibrational stimulus. Successive silhouettes were drawn after moving the figure a standard distance to the left. The first horizontal bar above each species indicates the fast-body-bend, the second bar, the return-flip. Note that the general characteristics of the responses in (a-h) are similar except that the hatchetfish (<) shows the effects of a rather in-flexible body on the response and the garfish (h) did not give the return-flip within the 100 msec test period. Rockfish (1) gave a response which consisted of a weak turn to one side followed by a body bend to the other. The spiny eel (j) responded by retracting its head.
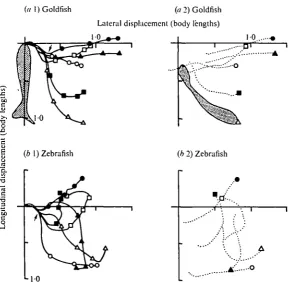
(1974) for the adult zebrafish. As seen from Fig. ib, c and further substantiated in Figs. 2-4 this startle response is divided into two phases. The initial phase consists of a short-latency, unilateral contraction of the trunk and tail. This contraction results in the fish assuming a C-like shape with both the head and tail displaced to one side. This is the most constant phase of the response and we name it the 'fast-body-bend' (Fig. ib,c). The fast-body-bend is followed by the second phase which consists of a more variable movement pattern resulting from a straightening of the tail. During this phase the two species were propelled on the average 0-98 and 0*97 L (body lengths) from their initial position (Fig. 2). This variable, second phase we term the 'return-flip' (Fig. ib, c).
Mauthner-initiated startle response in teleost fish
(« 1) Goldfish (a 2) Goldfish
Lateral displacement (body lengths)
1 0 .
I {b 1) Zebrafish o
(b 2) Zebrafish
Q.
.a
X) C
[image:7.451.81.372.67.349.2]•5
Fig. a. Displacement pathway (a 1 and b 1) and final body orientation (az and 62) of goldfish (a) and zebrafish (6). Lines in (a 1) and (6 i), respectively, describe the pathway of the head of the goldfish and zebrafish relative to initial orientation in six trials. The body length, direction of body bend, and the initial orientation at the beginning of the response are standardized as shown by the silhouette in (a 1) and pathways are determined by marking successive positions at 5 msec intervals. At the time of stimulus presentation the rostral-most tip of the head is at the origin. During the initial 15-20 msec period (the fast-body-bend) the pathways among trials are essentially identical, and subsequent to that, during the return-flip, the pathways among trials are quite variable (a 1 and 61). The arrows indicate the end of the fast-body-bend phase. The dotted lines in graphs (02) and (62) show the orientations of the midline of the body at the end of the 100 msec test period as illustrated by the silhouette in (02).
four points) corresponds to the fast-body-bend shown in Fig. 1 b, c. During the return-flip the fish turned at different rates so that after 100 msec the animals' positions were quite variable with respect to the initial position (Fig. 2).
The biphasic nature of the response is clearly seen in graphs of displacement speed and angular velocity (Fig. 4). The fast-body-bend is characterized by a sudden in-crease in both displacement speed and angular velocity. During the fast-body-bend phase the animal achieves its greatest angular velocity and displacement speed (see Table 2). The end of the fast-body-bend occurs when the angular velocity returns to approximately o° per sec, whereas displacement speed, although decelerating, remains relatively high for a period of time as the fish executes its return-flip.
(b) Startle responses in other teleost fish
0-25
0-5
Lateral displacement (lengths)
025 0 5 0-75
a o
m
075 L Goldfish
Fig. 3. Large-scale plot of consecutive positions of the head during the first 35 msec of each of the six startle responses of goldfish in Fig. a (a 1). Each symbol represents the positions, at 5 msec intervals, of individual fish during the response. In this figure the latencies were standardized so that the responses were initiated at the same frame (latency 5 msec). Note that during the fast-body-bend, the first four points, the position of the head was quite con-sistent between trials.
(a) Goldfish (b) Zebrafish
20 r
8
u
c
, • • » • . .
"I
H -J J
1 1
-1
J
1
0 20 40 60 80 100
-I 50
-I
J
1
H
j • • • • • .
v# . ' . .
-1 -1
X4
0 20 40 60 80 100
Time (ms)
Fig. 4. Displacement speed and angular velocity for goldfish (a) and zebrafish (6). Solid lines represent angular velocity and dotted lines represent displacement speed. Each point represents the mean of six trials corrected so that the responses were initiated at the same frame (latency 5 msec). See Table 2 for mean value of actual latencies. Note that the peak angular velocity and displacement speed are achieved simultaneously. Arrow indicates approximate end of fast-body-bend phase and onset of return-flip. In four of the six trials, zebrafish executed a turn after the return-flip (Fig. 1 c), but this movement averaged out among trials because the animal did not always turn the same direction relative to the direction of the fast-body-bend.
Mautkner-initiated startle response in teleost fish
73
10 Trout
r
100-5 0-5 o-5
1 0 Kelp bass
t?
0-51-1-0
Spiny eel Spiny eel10
10 Churacid
I (e)
0-5 0-5 . . 0 - 5
10
Garfish Garfish10
L
[image:9.451.94.359.78.422.2]Hatchet fish
| 1-0 (A.) r l O Rockfish
Black ghost Catfish
Fig. 5. Displacement pathways and final body orientation of diverse fish: (a) trout, (6) kelp bass, (c) characid, (<f) and (e) spiny eel, (/) Sc(g) garfish, (A) black ghost, (i) catfish, (j) common (triangles) and marbled hatchetfish (filled circles), (k) rockfish. Format is similar to Fig. 2 except that the fish's pathway (solid line) and final body orientation (dotted line) are shown on the same graph. Asterisks in (6) and (h) indicate fast-forward-displacement responses also seen in the rockfish (k). The responses of the spiny eel (d, e) are also different from the biphasic response characteristic of most of the other species. For the spiny eel the initial position is indicated by the dashed line.
body musculature resulting in a retraction of the head (Fig. if). All these responses had similar latencies, but as discussed below, their angular velocities and displace-ment speed characteristics were quite different.
74
(c) _ V. /
-Characid
i
i - i -i -i
4U
Spiny eel
; ^
. Kelp
V" '-... | 1
Bass -n
I
40
g
I
• Q .
.S3
Q
- (fc)
Black ghost Rockfish
Time (msec)
i 4 0
0 40 80 Kelp bass
Fig. 6. Displacement speeds and angular velocities for diverse fish: (a) trout, (6) catfish, (<;) characid, (d) garfish, (e) spiny eel, (/) and (i) black ghost, (g) marbled hatchetfish, (A) and (A) kelp bass, (j) rockfish. Format is same as Fig. 4 except that each graph represents a single trial. The data in (a) to (g) and (j) are from the same trials as shown in Fig. 1. Graphs (1) and (k) are fast-forward-displacements. Comparing those with (/) and (A) shows how they differ in the same species from the biphasic responses. In (6) and (/) either the angular velocity or dis-placement speed could not be determined over the entire 100 msec response time because part of the fish was out of view. In (k) the kelp bass hit the side of the tank at 60 msec into the response.
Mauthnet-initiated startle response in teleost fish
75
+
[image:11.451.56.400.46.202.2]+
Fig. 7. Fin erection during a startle response in the kelp baas. The stimulus occurs at (1) and successive figures are at 5 msec intervals. Splaying of the dorsal fin and caudal fin occur simultaneously with the initiation of the response at a latency of 5 msec. Crosses mark a fixed reference point, and the bar indicates 4-0 cm.
had the same latencies as the biphasic startle responses observed for the majority of species. However, the animals failed to achieve velocities comparable with those of their biphasic responses (Fig. 6 / vs. i and h vs. k). In addition, there was no early peaking in the angular velocity or displacement speed (Fig. 6 i, k) and in the case of the response of the black ghost, the angular velocity maximum did not correspond to a single displacement maximum (Fig. 61). These differences justify the distinction between the two categories of responses, and suggest that different neurophysiological processes underlie them.
All the responses of the spiny eel consisted of bilateral contractions of the body musculature (Fig. 17) which resulted in a withdrawal of the head (Fig. $d, e). The response is not biphasic, although there are simultaneous peaks in the angular velocity and displacement speed curves (Fig. 6c).
In some species, the animal's position in the tank may have influenced the type of response that was obtained. In halibut, which we filmed from the side, only a slight movement of the fins and a withdrawal of the eyes could be obtained when the animal was resting at the bottom in five trials. Lack of responsiveness in these cases seems consistent with the adoption in this species of protective coloration which matches the background. However, a biphasic startle response was obtained when the animal was stimulated while swimming. Similar conditions may have been responsible for our obtaining only the fast-forward-displacement, or head retraction in calico rock-fish and spiny eel, respectively. During these trials the animals were resting on the bottom of the tank.
(c) Fin movements during the biphasic startle response.
7
6
R. C. EATON AND OTHERS15
Fig. 8. Fin and body movements of the marbled hatchetfish in response to vibrational stimula-tion. Drawings are made at 5 msec intervals, and the crossed lines represent standard reference points 1 -6 cm apart. The stimulus was delivered in the first frame. In the last frame the fish penetrated the water surface (dotted line). The fins are shown only where they could be seen. See text for details.
Eye and opercular movements, probably involved in the response (Diamond, 1971) could not be clearly seen.
The pectoral fins play an important, though variable, part in the startle response of hatchetfish. In some cases the initiation of the fast-body-bend and flexion of these fins occurred simultaneously, whereas in other cases the pectorals were not involved in the initial phases of the response, but were activated later. Fig. 8 shows an example of a marbled hatchetfish startle response where pectoral fin movements precede (frames 2 and 3) and follow (frames 13 and 14) a fast-body-bend (frames 5-9). Following the second pectoral fin response the animal left the water (frame 15) for 175 msec and landed 8-4 cm away. The fish was not in view while above the water surface.
(d) Visual activation of the startle response in goldfish
Mauthner -initiated startle response in teleost fish
(«) (b)
i A+ 11
77
J>W^+ 3 JU 13
^2>
£ a
+ 6e
+ 7
i
+ 8 ,
+ 9
C=^^+ 14
-<S^"16
•-^= + 17
^ ^ 8
. ^+19
/^\
+1
°
Fig. 9. Side views of goldfish responses to vibrational (a) and visual (6) stimulation. The stimulus in (a) occurred at frame 1 and successive frames in (a) and (6) show body orientation at s msec intervals. Crosses mark a fixed reference point. The responses to the two stimuli do not differ significantly. Fin rays were drawn when they could be discerned and thus their number and spacing provide some indication of splaying, which is particularly evident in (6) 2-6. The bar indicates 4-5 cm.
DISCUSSION
Our data provide quantitative evidence that there is a characteristic biphasic startle response behaviour found in a wide variety of teleost fish having M-cell systems. This behaviour consists of a short latency (Table 2), unilateral contraction of the trunk and tail, such that the animal assumes a characteristic C-like shape with the head and tail displaced to one side (Fig. 1 a-h). The initial movement is followed by a less extensive contraction to the opposite side (Fig. 1 a-g). The result of the behaviour is that within 100 msec the animal is usually displaced from 0-5-1-5 L from its initial position (Figs. 2, 5). Two species did not display the characteristic response pattern under our conditions (Fig. 11,7).
We propose that the biphasic startle behaviour we observed in the majority of
species is initiated by the M-cell. This hypothesis is strongly supported by a combina-tion of previous physiological and anatomical data from goldfish and zebrafish. The M-cell systems in these two species are alike, and all evidence indicates that the system mediates a rapid reflexive movement. The M-cells receive excitatory electrical input from fibres of the eighth root and in turn directly activate the spinal motor neurones (Diamond, 1971; Eaton & Farley, 1975; Eaton, Farley, Kimmel & Schabtach, 1977).
Examination of species without M-cell systems would not provide a crucial test of this hypothesis because in the present study, some fish with M-cell systems did not display the characteristic behaviour and others displayed it only part of the time or under certain conditions. The best test of the hypothesis would be to record from the M-cell and simultaneously photograph the response with high-speed cinemato-graphic equipment. Such a study has not yet been attempted.
However, the available data demonstrate that although other cells are involved, the M-cell appears to play an essential role in the production of most examples of startle responses studied. In restrained goldfish, Yasargil & Diamond (1968) recorded a strong contralateral contraction of the body musculature in response to electrical stimulation of the eighth root and activation of the M-cell. Eaton & Farley (1975) recorded a single M-spike in 116 out of 118 startle responses in restrained zebrafish larvae responding to vibrational stimulation. No M-spikes were recorded i n n cases when the animal gave no response. More recently, Zottoli (1977) recorded from the M-cell and trunk musculature in free-swimming adult goldfish. Zottoli recorded single M-spikes in response to acoustic stimulation in 35 out of 36 cases when there was a maximal contraction of the contralateral body musculature, and the M-spike was absent when the ipsilateral musculature contracted in 38 cases. No M-spike was recorded in 52 cases when the goldfish failed to give a tail-flip. In the goldfish and zebrafish, a variety of neural units fire during the startle response, but the most constant feature of such records is the presence of an initial M-spike.
The physiological and anatomical data from common hatchetfish are also consistent with the idea that the M-cells initiate a rapid reflex movement (Auerbach & Bennett, 1969). In other species we studied there are no previous physiological experiments but the major anatomical features of the M-cell systems are similar to those of the goldfish and zebrafish. The anatomical and behavioural consistency support our hypothesis that the M-cells initiate a common type of behaviour in these species as well.
Therefore, we propose the name 'Mauthner-initiated startle response' for the characteristic startle behaviour described for the majority of species in this study. The previous designation 'tail-flip' is misleading because the most characteristic feature of the response is the initial contraction of the body which leads to a lateral displacement of both head and tail. The terminology' Mauthner reflex' or' Mauthner-mediated response' is unjustified at present because the data are not sufficient to determine at what point the influence of the M-cell ends (see below).
Mauthnet--initiated startle response in teleost fish 79
appears to involve an extensive unilateral contraction of the body musculature (Fig. 3). These features lead to the proposition that the fast-body-bend phase is the direct result of the activation of a single M-cell and its spinal motor neurone pool. A test of this hypothesis is to cinematographically analyse startle behaviour following uni-lateral and biuni-lateral lesions of the M-cells. This hypothesis predicts that the fast-body-bend to one side would be absent after destruction of the contralateral M-cell. Webb (1975) did not find stereotyped acceleration responses in trout probably because he used a slower frame rate. Using similar measurement techniques our mean maximum displacement for zebrafish (42 L/sec, n = 6) is approximately the same as the maximum of 37 L/sec seen in the example of Kimmel et al. (1974) who used a frame rate comparable to ours (1 frame/6-7 msec vs. 1 frame/5 msec.) However, for trout we recorded a maximum of 24 L/sec, 12 msec after the start of the response (Table 2) whereas previous studies with slower frame rates (1 frame/i5-6 msec) obtained maxima of 9-5 and 15-5 L/sec, 78 and 75 msec into the response (Hertel, 1966; Webb, 1975). In previous studies the average displacement speed, which is not as sensitive to frame rate, is close to the value we obtained. Over the first 100 msec of the response, we obtained values of 6-9 L/sec compared with 5-7 L/sec in earlier studies (Hertel, 1966; Webb, 1975). These comparisons show how slow frame rates can obscure important details of the startle response behaviour.
The second phase of the Mauthner-initiated startle response, the return-flip, is a variable contraction of the body in the direction opposite to that of the fast-body-bend. The return-flip appears to begin when the angular velocity returns to zero, and has an elevated displacement speed. In garfish, the return-flip appeared to be delayed or inhibited (Figs. 1 h, sf,g). This suggests that the fast-body-bend and return-flip are controlled by separate, although perhaps interdependent, physiological mechanisms. The garfish examples support the idea that the return-flip is an active process and not simply a passive mechanical consequence of the fast-body-bend. The return-flip is probably not mediated by a second ipsilateral or contralateral M-spike because in the combined 151 examples reported for larval and adult fish, the startle response was always accompanied by one M-spike only (Eaton & Farley, 1975; Zottoli, 1977).
It is our view that other startle response behaviour, such as the fast-forwaid-displacement of the black ghost, kelp bass and rock fish, or the rapid contraction of the head of the spiny eel, are probably not initiated by the M-cell. In the case of the fast-forward-displacement, there is no maximal body contraction, as is expected for behaviour produced by the M-cell. In the species so far examined, the M-cell appears to predominantly innervate the contralateral motor neurones (Diamond, 1971), so it also seems unlikely that the M-cell mediates a behaviour involving an extensive bilateral contraction such as seen in the spiny eel.
In adult fish, the Mauthner-initiated behaviour is thought to be important in the avoidance of predators, such as birds, which strike the water surface from above (Diamond, 1971; Zottoli, 19766). Our observation that the response is also activated by visual stimulation supports this idea. Diamond (1971) has argued that the Mauthner-initiated response is non-directional in terms of whether the left or right cell fires in the presence of an acoustic stimulus in the near field. Our data indicate that there is an additional source of unpredictability built into the system; between separate trials,
20 msec after the animal starts contracting, its position begins to become variable. Such variability is consistent with the strategy of escape, and could be useful to prevent a predator from compensating for the response. Unpredictability is also a feature of rapid escape responses of other animals such as the hawk moth (Roeder, 1962).
The fast-body-bend probably has two functions in predator avoidance. It may be a preparatory movement (see Webb, 1975; Weihs, 1973) in which the fish initiates a turn and prepares to swim away by a flip of the tail. At the same time, the most vulnerable part of the animal's body, the head, is abruptly displaced and is no longer a stationary target. According to this view, the return-flip represents the main propulsive action (see Webb, 1975) which begins the fish's escape from a threatening stimulus. This idea is supported by the response of the garfish, which under our condi-tions gave a fast-body-bend but not a return-flip, and therefore remained close to its original position (Fig. 5/,^).
The few examples of startle behaviour which did not appear to be Mauthner-initiated responses provide some insight into the possible significance of the M-cell system. Since those behaviours had latencies the same as the Mauthner-initiated responses, it appears unlikely that this giant fibre system was evolved simply to bring about a fast response. This is consistent with observations of Bullock & Horridge (1965) and Kennedy (1975) on giant fibre systems. It is likely that the importance of M-cell system lies in activating, roughly simultaneously, a large body of musculature to bring as strong a body bend as possible.
This research was made possible by the generous loan of cinematographic equip-ment from Dr John Hunter, National Marine Fisheries, La Jolla, CA. We thank Dr Sven O. E. Ebbesson for supplying the technical support for our histological analysis and Dr Theodore H. Bullock for providing facilities. Drs Theodore H. Bullock, Donald S. Faber, Charles B. Kimmel and Steven J. Zottoli provided valuable comments on the manuscript. Support was provided by Individual National Research Service Awards to R. C. Eaton and R. A. Bombardieri from the National Institutes of Health, by research grants to Dr Bullock from the National Science Foundation and the National Institutes of Health, by German Science Foundation grant Me526/1-4 and SFB33 to D. L. Meyer, and by grants to Dr Ebbesson from the National Aero-nautics and Space Administration (NGR 47-005-186) and the National Institutes of Health (Ey 00154-05).
REFERENCES
AUERBACH, A. A. & BENNETT, M. V. L. (1969). Chemically mediated transmission at a giant fiber synapse in the central nervous system of a vertebrate. J. gen. Phytiol. 53, 183-210.
BAFFONI, G. M. & SERRA, L. (195a). La citomorfosi del neurone mauthneriano di un teleosteo durante lo sviluppo. Riv. Biol. 44, 469-91.
BARTHLMEZ, G. W. (1915). Mauthner's cell and the nucleus motorious tegmenti. J. comp. Nettrol. 25, 87-128.
BODIAN, D . (1937). The structure of the vertebrate synapse. A study of the axon endings on Mauthner's cell and the neighboring centers in the goldfish. J. comp. Nexirol. 68, 117-59.
BREDER, C. M. (1926). The locomotion of fishes. Zootogica 4, 159-297.
BULLOCK, T. H. & HORRIDGE, G. A. (1965). Structure and Function in the Nervous SytUnu of
Inverte-brates. San Francisco: W. H. Freeman & Co.
DIAMOND, J. (1971). The Mauthner cell. In Fish Physiology, vol. V (eds W. S. Hoar and D . J. Randall), pp. 265—346. New York: Academic Press.
Mauthnet-initiated startle response in teleostfish 81
EATON, R. C , FARLEY, R. D., KIMMEL, C. B. & SCHABTACH, E. (1977). Functional development in
the Mauther cell system of embryos and larvae of the zebra fish. J. Neurobiol. (in the press). GOSLINE, W. A. (1971). Functional Morphology and Classification of Teleosteon Fishes. Honolulu:
University Press of Hawaii.
HERTEL, H. (1966). Structure, Form, Movement, 251 pp. New York: Reinhold Publishing Corporation. KENNEDY, D. (1975). Comparative strategies in the investigation of neural networks. J. exp. Zool. 194,
35-SO.
KIMMEL, C. B., PATTERSON, J. & KIMMEL, R. O. (1974). The development and behavioral
character-istics of the startle response in the zebra fish. Dev. Psychobiol. 7, 47-60.
LEOHISSA, S. (1943). Le basi anatomiche nella evoluzione del 'comportamento' durante lo sviluppo embrionale e post-embrionale di Trota (Salmo fario, irideus e lacustris). Z. Anat. EntioGesch. I l l , 601-75.
LUNA, L. G. (i960). Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology. New York: McGraw-Hill Book Co.
ROEDER, K. D. (1962). The behaviour of free flying moths in the presence of artificial ultrasonic pulses.
Animal Behav. 10, 300-4.
STERBA, G. (1962). Freshwater Fishes of the World. New York: Viking Press.
WEBB, P. W. (1975). Acceleration performance of rainbow trout Salmo gcarneri and green sunfish
Lepomis cyanellus. J. exp. Biol. 63, 451-65.
WEIHS, D. (1973). The mechanism of rapid starting of slender fish. Biorheology 10, 343-50.
YASARGIL, G. M. & DIAMOND, ] . (1968). Startle-response in teleost fish: an elementary circuit for neural discrimination. Nature 220, 241-3.
ZIESMER, C. (1952). Silberfarbung an Paraffinschnitten. Eine weitere Verbesserung der Bodian-Methode. Mikroskopie 7, 415-17.
ZoTTOLi, S. J. (1976a). A comparison of Mauthner cell size in teleosts and cell function in escape tail-flips of unrestrained goldfish in response to sound. Ph.D. Dissertation, University of Massachusetts. ZOTTOLI, S. J. (19766). Fishing behavior of Common Grackles. Auk. (in the Press).